
Abstract
Generation of induced pluripotent stem cells (iPSCs) from other somatic cells has provided great hopes for transplantation therapies. However, these cells still cannot be used for clinical application due to the low reprogramming and differentiation efficiency beside the risk of mutagenesis and tumor formation. Compared to iPSCs, induced neural stem cells (iNSCs) are easier to terminally differentiate into neural cells and safe; thus, iNSCs hold more opportunities than iPSCs to treat neural diseases. On the other hand, recent studies have showed that small molecules (SMs) can dramatically improve the efficiency of reprogramming and SMs alone can even convert one kind of somatic cells into another, which is much safer and more effective than transcription factor-based methods. In this study, we provide a review of SMs that are generally used in recent neural stem cell induction studies, and discuss the main mechanisms and pathways of each SM.
Introduction
T
Recently, several studies have reported that fibroblasts can be reprogrammed into iNSCs by TFs [2,16 –19], but they are unlikely useful as the source of cell transplantation in clinics because of the risk of transgene integration, and the association with viral transduction vectors have significant safety concerns. The unsafety of using TFs or viral transduction vectors has forced researchers to find nonintegrative ways of generating therapeutic cells. For example, the therapeutic cells generated by the delivery of mRNA, protein, and small molecules (SMs) are comparatively safe and secure [20]. Among these methods, SMs are effective as they can be easily introduced and manipulated. Their effects are specific and reversible, and they are inexpensive and can be used at different concentrations. This makes SMs favorable over protein or TF-based methods in cell therapy. On the other hand, some studies have showed that SMs can effectively improve the efficiency of reprogramming [11,21], and even SMs alone can convert one type of somatic cells into another [22 –24]. Typically, SMs can increase the efficiency or even manipulate cell fate by playing four separate roles: (1) epigenetic modifiers; (2) signaling pathway regulators; (3) metabolic modulators; and (4) nuclear receptor agonists and antagonists, although the first two mechanisms are more widely used in reprogramming researches. The aim of this review is to summarize the recent literature of iNSCs and discuss different mechanisms by which SMs play during the induction of somatic cells into neural stem cells.
Epigenetic Modifiers
Epigenetic landscape has been considered a major obstacle for reprogramming, as it is necessary, but not easy, to reset the existing somatic epigenetic memory in a reprogramming process. Epigenetic landscape is mainly modified by two mechanisms, DNA methylation and histone modifications, which are related to many enzymes, such as DNA methyltransferases (DNMTs), histone acetyltransferases, histone deacetylases (HDACs), and histone demethylases [25,26]. Correspondingly, there are some SMs that can regulate these enzymes, and consequently modify the epigenome and accelerate the reprogramming process.
Valproic acid
Valproic acid (VPA) (Table 1, NO. 1.1) is a short-chain fatty acid that has been used in clinic to treat epilepsy and bipolar disorder, and to prevent migraine headaches. As it can inhibit HDACs, it is also widely used in reprogramming-related studies to erase the original epigenetic memory of cells. It has already been known that VPA could have an important function in iPSC induction process. For example, it increases the efficiency by 100-fold while reprogramming mouse fibroblasts with Yamanaka factors [27], and could replace Klf-4 and c-Myc and increase the efficiency from 10 to 20-fold while reprogramming human fibroblasts [21]. Moreover, VPA has been suggested as the best enhancer of pluripotency in vivo when compared to other reported epigenetic modifiers like Bix01294, BayK8644, and RG108 [28]. Mirakhori et al. [29] converted human foreskin fibroblasts (HFFs) into induced neural progenitor cells (iNPCs) by using SOX2 protein transduction, human LIF, and a cocktail of SMs composed of LDN193189, SB431542, CHIR99021, purmorphamine, and VPA in 3D culture condition, while using SOX2 protein or SMs alone could only partially convert these cells. Zheng et al. [30] converted mouse embryonic fibroblasts (MEFs) to NSCs by only using a combination of VPA, A-83-01, Thiazovivin, and purmorphamine. Moreover, these four molecules could be replaced by other chemical inhibitors or activators of the same pathways, which are NaB (HDAC inhibitor), SB431542 (TGFβ inhibitor), Y-27632 (Rock inhibitor), and 25-hydroxycholesterol (SHH agonist), respectively, indicating the importance of these pathways in NSC induction. Cheng et al. [31] generated NPCs from MEFs, mouse tail-tip fibroblasts (TTFs), and human urinary cells (HUCs) by using a combination of VPA, CHIR99021, Repsox and Parnate under 5% O2 condition, in which VPA was necessary, while Parnate, a histone demethylase inhibitor, was not. It may indicate that VPA alone, at proper doses, is enough to erase the original epigenetic memory of the cells (Table 2). Interestingly, although the hypoxia condition is essential for NPC induction in this study, it cannot be replaced by cobalt chloride, which is known to induce cellular hypoxic response, suggesting that exact pathways activated under this condition remain to be identified. VPA is also known to directly or indirectly inhibit the activity of glycogen synthase kinase 3β (GSK3β) [32,33], which is important for reprogramming, indicating the multiple targets/functions of VPA.
NPC, neural progenitor cell; NSC, neural stem cell.
AHDF, adult human dermal fibroblasts; HUC, human urinary cell; MEF, mouse embryonic fibroblast; SM, small molecule; TTF, tail-tip fibroblast.
It is also worthy to note that VPA could arrest NSC/NPC proliferation and promote their neuronal differentiation [34,35], which may indicate that it is crucial to remove VPA at some time point in reprogramming process to sustain NSC/NPC properties. After all, it can be summarized that VPA could play a very helpful role in NSC/NPC induction processes.
Sodium butyrate
Sodium Butyrate (NaB) (Table 1, NO. 1.2) is also an HDAC inhibitor, which is similar to VPA. It has been reported that NaB could replace VPA in some NSC/NPC induction protocols [30,31]. Actually, VPA has been recommended as a viable alternative to NaB as it is more cost-effective than NaB [36]. However, there are still some differences between these two SMs in improving reprogramming efficiency. Zhang and Wu [37] found that NaB alone could significantly improve HFF-to-iPSC reprogramming efficiency by upregulating the miR302/367 cluster, whereas VPA alone could not, indicating that the inhibition of HDACs is not the only mechanism by which NaB promotes reprogramming. Zhu et al. [38] reported a chemical cocktail that includes NaB could facilitate the reprogramming of adult human dermal fibroblasts (AHDFs) transfected with OCT4 alone into iNSCs.
Tranylcypromine
Tranylcypromine (also named Parnate, see Table 1, NO. 1.3) is a histone demethylase LSD1 inhibitor, which has been shown to enable reprogramming of human keratinocytes induced by Oct4 and Klf4 in combination with CHIR99021 [39]. Besides, Zhang et al. [40] efficiently reprogrammed mouse fibroblasts into iNSCs by using a cocktail of nine compounds (M9), which contained two epigenetic modifiers, RG108 and Parnate. Moreover, removal of either of these two molecules would significantly reduce the reprogramming efficiency, indicating their importance in the process. However, Cheng et al. [31] found that Parnate was removable in their induction protocol when they combined Parnate with VPA, CHIR99021, and Repsox. It is unclear why the importance of these epigenetic modifiers in reprogramming process is variable.
It is also worthy to notice that, like VPA, Parnate has been reported to dramatically reduce NSC proliferation [41], which may affect the success of NSC induction.
Bix01294
Bix01294 (Table 1, NO. 1.4) is a G9a histone methyltransferase (G9a HMTase) inhibitor. Shi et al. reported that Bix10294 could enable reprogramming of NPCs transduced with c-Myc, Klf4, and Sox2, in the absence of Oct4 ectopic expression [42], and could enable reprogramming of Oct4- and Klf4-transduced MEFs [43]. However, Bix01294 is usually combined with other epigenetic modifiers, like RG108 [43] or VPA [44], or an L-type calcium channel activator, BayK8644, to enhance reprogramming efficiency. Han et al. [45] reprogrammed MEFs into NSCs by a combination of SMs containing VPA, Bix01294, RG108, PD0325901, CHIR99021, A83-01, and ascorbic acid (AA), but only Bix01294, RG108, and PD0325901 were indispensable for TTF reprogramming, indicating their importance in this induction system.
RG108
RG108 (Table 1, NO. 1.5) is a nonnucleoside DNMT inhibitor that acts by binding and blocking the active site of the enzyme or by masking DNMT target sequences [46]. It has been reported that RG108 could improve reprogramming efficiency in some iPSC induction studies [22,43]. Moreover, just like Bix01294, RG108 is usually combined with other epigenetic modifiers to fully erase the original epigenetic memory of the cells. Although RG108 has not been widely used as VPA in reprogramming researches, it is essential to include RG108 in some NSC induction protocols. It has been shown that TTFs could be reprogrammed into NSCs by using a combination of Bix01294, RG108, and PD0325901 [45]. Another group reported that MEFs could be transformed into NSCs by using a cocktail of 9 compounds that contained RG108 [40]. Remarkably, removal of RG108 would lead to significant reduction of the reprogramming efficiency in both of these cases, indicating that DNA methylation is a major barrier to NSC reprogramming.
Azacitidine
Azacitidine (AZA, 5-AZ, see Table 1, NO. 1.6) is a nucleoside DNMT inhibitor, which can be incorporated into DNA and form suicide substrates for DNMTs [46,47]. Just like the SMs we described above, AZA has also been reported to enhance reprogramming efficiency [21,27,48]. Surprisingly, Mirakhori et al. [49] reported that HFFs could be converted into neural progenitor-like cells by only a short presence of AZA in suspension culture condition, although these cells could only differentiate into neurons and astrocytes, but not oligodendrocytes in vitro. Moreover, it has also been reported that AZA is more toxic for cells in monolayer condition than in suspension, indicating the need to optimize the proper condition to use this SM.
Ascorbic acid
Ascorbic acid (AA, also known as vitamin C, see Table 1, NO. 1.7) is usually known as an antioxidant. However, AA has also been reported as an epigenetic modifier, which can increase histone or DNA demethylation [50,51], and hence, it can regulate gene expression and improve the efficiency and quality of reprogramming of somatic cells [52,53]. More remarkably, a combination of AA and CHIR99021 has strong synergistic effects for iPSC generation, which gives at least 10-fold efficiency compared to individual treatment, although the mechanism is not clear. Interestingly, although AA has been more widely used for iPSC generation, there are few protocols that include AA for inducing NSCs/NPCs. Han et al. [45] used AA to reduce cell death, while converting MEFs to NSCs; however, later, they found AA was dispensable for TTF-to-NSC conversion [45].
Retinoic acid
Retinoic acid (RA, see Table 1, NO. 1.8) is a metabolite of vitamin A. It has been reported that RA can enhance reprogramming by erasing DNA methylation of somatic cells, especially when it is combined with ascorbate, forming complementary effects [54]. Moreover, some articles have shown that RA receptors or RA agonists are required for reprogramming [55,56], which also indicate the importance of RA. Interestingly, RA is also shown to enhance iPSC differentiation into neuronal progenitors [57]. Besides, RA has been reported as an essential component for Zhang's “M9” protocol, which can convert MEFs into NSCs [40].
Signaling Pathway Regulators
Transforming growth factor-β family inhibitors
Transforming growth factor (TGF)-β superfamily plays multiple and important roles during cell development processes and differentiation, from embryonic stem cells to somatic cells. Its ligands are divided into two distinct subclasses, one is TGF-β/activin/nodal branch (hereafter abbreviated as TGF-β) and another is bone morphogenetic protein (BMP)/growth and differentiation factor branch, (hereafter referred as BMP). Their receptors are classified as three types, which are TGFRI, (also named activin-like kinases (ALKs), TGFRII, and TGFRIII. By binding to their specific receptors, TGF-β superfamily members can finally activate different Smad proteins, which are known as transcriptional cofactors that modulate gene transcription (for more information, refer to Refs. 58,59). Due to the diverse and complicated effects of TGF-β superfamily during cell development, in this study, we mainly focus on the functions related to reprogramming or NSC induction.
It has been reported that TGF-β could induce epithelial–mesenchymal transition, an essential process during embryonic development [60,61], while inhibition of TGF-β could enhance the reverse transition, mesenchymal-epithelial transition (MET), and therefore enhance reprogramming [61,62]. Conversely, BMP could induce MET, which is opposite to TGF-β, and thus promote reprogramming [58,63]. Moreover, it has been found that some inhibitors of TGF-β could replace Sox2 in reprogramming [64]. Interestingly, regarding NSC/NPC induction, Lihui et al. [65] reported that a TGFβ inhibitor, A83-01, could switch the cell fate from iPSCs into NPCs in TF (OSKM)-induced HUC reprogramming, indicating the importance of TGF-β inhibitors for NSC/NPC induction.
A83-01
A83-01 (Table 1, NO. 1.9) is a strong inhibitor for ALK4, 5, and 7, and can weakly inhibit ALK1, 2, 3, and 6. It has been reported that A83-01 could enhance reprogramming induced with OSKM [62] and enable iPSC generation from only Oct4-transfected MEFs, when combined with AMI-5, a DNA methylation modulator [61]. Interestingly, like what we mentioned above, A83-01 is sufficient and indispensable to switch the cell fate to NPCs from iPSCs during OSKM-induced reprogramming of HUCs [65], suggesting its significant role for NSC/NPC generation. Moreover, Zhu et al. [38] reported a cocktail consisting of A83-01, CHIR99021, NaB, LPA, rolipram, and SP600125 could convert Oct4-transfected AHDFs into iNSCs. In Han's protocol [45], MEFs could be converted into NSCs by using A83-01 and other SMs, although it was dispensable for TTF conversion. Moreover, in another two MEF-to-NSC researches [30,40], the researchers reported A83-01 was essential for their protocol, that is, removal of A83-01 significantly reduced NSC generation efficiency, indicating an important role of TGFβ signaling in the suppression of neural lineage induction.
Repsox
Repsox (E616452 and 616452, see Table 1, NO. 1.10) is an ALK5 inhibitor and has been reported to improve reprogramming [66]. Repsox can replace Sox2 or both Sox2 and c-Myc in reprogramming by inducing the expression of another transcriptional factor, Nanog [64]. Interestingly, this role can be replaced by SB431542, but not E616451 (both are TGF-β inhibitors), indicating their difference in functions [64]. Significantly, Repsox can induce endogenous Sox2 expression and transform different somatic cells into NPCs under 5% O2 condition when combined with VPA and CHIR99021 only [31]. Moreover, Repsox can also be replaced by SB431542 in this case, again suggesting the importance of TGF-β inhibition for NPC induction.
SB431542
SB431542 (Table 1, NO. 1.11) inhibits ALK4, 5, and 7 and has also been reported to be implicated in the MET transition and to enhance reprogramming [7,61,67]. Actually, it has been shown that SB431542 could be a substitution for A83-01 [30] or Repsox [31,64] in some reprogramming protocols. Moreover, Wenlin et al. [68] reported that a combination of dorsomorphin, CHIR99021, SB431542, and human LIF could direct the specific conversion of primitive NSCs (pNSCs) from hESCs within 7 days, and especially, the combination of CHIR99021, SB431542, and human LIF is uniquely required for long-term self-renewal of pNSCs. When combined with Compound C, SB431542 could also greatly improve the differentiation of hiPSCs into NPCs [69]. Mirakhori et al. [29] reported that a cocktail consisting of SB431542, LDN193189, CHIR99021, purmorphamine, VPA, and LIF could convert HFFs into NPCs in combination with SOX2 protein transduction.
LDN193189
LDN193189 (Table 1, NO. 1.12) is a potent inhibitor of BMP receptors ALK2, 3, and 6, and a weak inhibitor of ALK4, 5, and 7. Interestingly, although BMPs have been known to maintain mESC properties in the presence of LIF [70] and to induce MET (which is helpful for reprogramming), it has also been reported that BMPs could facilitate mesodermal and trophectodermal differentiation [71,72], indicating that the inhibition of BMPs could be an enhancer of neural induction. In fact, LDN193189 has been used in some NSC/NPC induction protocols [29,40,73,74]. Lee et al. [73] reported that a cocktail including Noggin (a protein that is a BMP antagonist), LDN193189, SB431542, and CHIR99021 could convert human blood cells into NPCs when combined with OCT4, and enhance NPC proliferation. Moreover, a combination of Noggin, LDN193189, and SB431542 has been reported to efficiently generate iNSCs from human adipose-derived stem cells [74]. These studies suggest that BMP inhibitors are helpful for NSC/NPC induction. Furthermore, removal of LDN193189 could lead to significant reduction of reprogramming efficiency [40]. It is also worthy to note that, when a BMP inhibitor has been used in NSC/NPC induction protocol, it is usually combined with a TGF-β inhibitor to dually inhibit Smad signaling, which can facilitate neural transition [29,40,73,75].
GSK-3 β inhibitors
GSK-3 β is an isoform of GSK-3, which has been found to have multiple functions and modulates various signal pathways, such as wnt/β-catenin, FGF/PI3K, and SHH/Gli [76,77]. Unlike most protein kinases, GSK-3 is active in resting cells and needs to be inhibited for pathway to function. Moreover, the activation of these three pathways has been reported to facilitate reprogramming in different articles [40,78,79], suggesting that the inhibition of GSK-3(β) may be very useful for reprogramming. Indeed, some researchers have shown an enhanced reprogramming by inhibiting GSK-3β, especially when it is combined with the inhibition of the MAPK/ERK pathway [80 –82]. Furthermore, due to its effective role for reprogramming and various functions in neural development [77,83] (for example, inhibition of GSK3β can enhance proliferation for both NPCs [84] and induced NPCs [73]), GSK-3β inhibitors have been widely used in NSC/NPC induction.
CHIR99021
CHIR99021 (Table 1, NO. 1.13) is the most selective GSK-3β inhibitor that is widely used in cell reprogramming researches. It has been shown that CHIR99021 could improve reprogramming efficiency and completion when combined with an MAPK/ERK inhibitor, PD0325901 [81,82]. Interestingly, CHIR99021 also has strong synergistic effects with AA during reprogramming, which can enhance the efficiency by more than 10-fold compared to individual treatments, although the mechanism needs to be clarified [85]. Furthermore, CHIR99021 is also a critical component for some NSC/NPC induction protocols. Cheng et al. [31] combined CHIR99021 with VPA and Repsox to convert MEFs, TTFs, and HUCs into NPCs under hypoxia. Moreover, they proved the importance of the three molecules by individually removing them and mentioned that further supplement with any other reprogramming-promoting compound showed no significant improvement. Also, CHIR99021 has been proved indispensable in the 9-molecule protocol, which could generate NSCs from MEFs [40]. Zhu et al. [38] used CHIR99021 and five other SMs to formulate a cocktail that could efficiently improve NSC generation from AHDFs. Also, when combined with three SMAD inhibitors [73], CHIR99021 could facilitate NPC reprogramming from human blood cells. It has been mentioned that this SMAD+GSK-3 inhibition can also enhance NPC proliferation, but with the expense of differentiation [68,73].
MAPK signaling pathway inhibitors
Mitogen-activated protein kinase (MAPK) pathway is critical for multiple biological processes, such as, cell differentiation, proliferation, and apoptosis [86,87]. To date, four distinct MAPKs have been found, including (1) extracellular signal-regulated kinase1/2 (ERK1/2); (2) c-Jun N-terminal kinase (JNK); (3) ERK5; and (4) p38 MAPK (p38), which function in different cascades [87]. Importantly, inhibition of ERK1/2 or JNK has both been shown to improve reprogramming [38,45,81,82,88], although inhibition of ERK1/2 is more widely used than JNK.
PD0325901
PD0325901 (Table 1, NO. 1.14) is a potent inhibitor of ERK1 and ERK2, which has been shown to enhance reprogramming by about 200-fold when combined with SB431542 and Thiazovivin [88]. PD0325901 together with CHIR99021 are termed “2i” (two inhibitors), which is a special set for reprogramming in iPSC generation and can significantly improve its efficiency [81,82]. Furthermore, PD0325901 is an essential component for Han's protocol, which can convert MEFs or TTFs to NSCs [45]. In another MEF-to-NSC article, however, PD0325901 has been shown to reduce the efficiency by about ninefold due to its incompatible effects with basic fibroblast growth factor [40], indicating the complicated mechanisms of reprogramming by the Erk1/2 pathway. It is also worthy to know that MEK is important for somatic cell survival [89], suggesting that the timing of MEK inhibitor treatment, for example, PD0325901, should be optimized.
SP600125
It has been reported that JNK1 and 2 can suppress Klf4 activity and therefore restrain reprogramming [90]. Reversely, the inhibition of JNKs by SP600125 (a JNK inhibitor, see Table 1, NO. 1.15) can increase ESC self-renewal and may be useful for reprogramming [90]. Indeed, a combination of A83-01, CHIR99021, NaB, rolipram, SP600125, and LPA, and an ectopic expression of OCT4 can convert AHDFs into NSCs [38].
Sonic hedgehog pathway agonists
The sonic hedgehog (SHH) pathway plays essential roles during development by regulating patterning and cell fate specification. It has been reported that the activation of SHH pathway can induce Bmi1, Sox2, and N-Myc expression, which are important TFs for reprogramming [91 –93]. Significantly, Moon et al. [94] converted MEFs into NSC-like cells by only using an SHH agonist, purmorphamine or oxysterol. Moreover, MEFs could be reprogrammed into iPSCs by combining any of these agonists with OCT4 or Nanog alone [94,95]. Furthermore, the SHH pathway has also been shown to regulate proliferation and differentiation of NSCs/NPCs [96,97]. Besides, agonists of the SHH pathway have been used in the induction research on NSCs/NPCs. Purmorphamine (Table 1, NO. 1.16) is a crucial component of Zheng's MEF-to-NSC protocol [30], in which the removal of Purmorphamine leads to significant reduction of the reprogramming efficiency. Yet, Purmorphamine can be replaced by another SHH agonist, 25-hydroxycholesterol (Table 1, NO. 1.17), indicating the importance of SHH signaling activation for reprogramming. Moreover, Purmorphamine is also included in another protocol, by which Mirakhori et al. [29] have generated NPCs from HFFs. Another SHH signaling agonist, Hh-Ag 1.5 (Table 1, NO. 1.18), is an extremely important molecule in Zhang's “M9” protocol, which could generate NSCs from MEFs; omitting Hh-Ag 1.5 would mostly reduce the induction when compared to other SMs [40].
ROCK inhibitors
Rho-associated protein kinase (ROCK) has multiple functions, including the regulation of cellular contraction, motility, morphology, polarity, cell division, and gene expression [46]. It has been reported that inhibition of ROCK can enhance viability of iPSCs and NPCs [88,98,99], therefore improving reprogramming efficiency. For instance, Thiazovivin (Table 1, NO. 1.19) has been shown to significantly enhance the efficiency of MEF-to-NSC conversion in Zheng's protocol [30]. Also, this effect could be replaced by another ROCK inhibitor, Y-27632 (Table 1, NO. 1.20). However, it is also worthy to note that inhibition of ROCK may lead to cellular differentiation, which may affect NSC/NPC generation [100].
Others
SMER28
SMER28 (Table 1, NO. 1.21) is an autophagy modulator, which has been shown essential for the “M9” protocol [40]. Removal of SMER28 significantly reduced the MEF-to-NSC conversion [40].
Rolipram
Rolipram (Table 1, NO. 1.22) is a selective inhibitor of phosphodiesterase 4 (PDE4), which can prevent cyclic adenosine monophosphate (cAMP) hydrolyzation and inhibit the production of reactive oxygen species [101,102], thereby increasing cAMP levels within cells and protecting cells. Zhu et al. included rolipram in their protocol, which generated NSCs from AHDFs [38].
Lysophosphatidic acid
Lysophosphatidic acid (LPA; a phospholipid derivative, see Table 1, NO. 1.23) has also been used in Zhu's protocol to convert AHDFs into NSCs when combined with A83-01, CHIR99021, NaB, rolipram, SP600125 and OCT4 [38]. However, it has been reported that LPA can activate Rho/Rock signaling [103], which contrasts with Rock inhibitors that we have introduced earlier. It still needs to clarify how LPA works for NSC/NPC conversion.
Conclusions and Perspectives
It has been known for a long time that cell fate can be reversible. Based on this, generation of iPSCs from different cell types has been achieved in these years. However, since iNSCs/iNPCs hold more opportunities for clinical applications to treat neurological diseases than iPSCs, the generation of iNSCs/iNPCs from somatic cells has attracted more and more interests.
SMs can not only improve reprogramming efficiency, replace TFs, but also can reprogram cells in the absence of genetic material. Reprogramming by SMs is safe and desirable for clinical applications. It is assumed that the SMs, particularly epigenetic modifiers alter the prevailing genetic imprinting to generate a flexible and naïve environment favoring the NSC/NPC generation together with the signal pathway modulators [104]. However, there are several problems associated with this approach. First, there is hardly any protocol that can be reproducible among different laboratories without substantial modifications, even using the same type of cells for reprogramming. Second, the published protocols and recipes may only work with specific cells in a particular laboratory and cell status may largely determine what SMs are required during reprogramming and induction. Third, a significant drawback of some SMs are their multiple targeted activities. Thus, their effects may unexpectedly make the reprogramming process more complicated. Besides, the proper dosage of an SM, another important factor of effects, may be variable among different cell types, indicating that it is important, but maybe also difficult, to optimize the concentration of each SM in a specific protocol. Therefore, a detailed study is required to be carried out in terms of understanding the clear mechanism of these molecules, to develop a universal protocol for generating iNSCs from any type of somatic cell for clinical applications.
To date, although many groups have generated NSCs/NPCs from somatic cells by using SMs alone, most of the host cells were from mouse, not human (Table 2), which indicates that the reprogramming barriers of different species/cells may be different. Fortunately, the cellular mechanisms between mouse and human are usually similar, indicating that a good understanding of reprogramming mechanisms of mouse cell can help reprogramming human cells. In addition, it is also essential to understand the functions of SMs, which are used for NSC/NPC reprogramming. Therefore, understanding the reprogramming mechanism and the functions of individual SMs will avoid the try-and-error tedious testing and increase the chances of successful reprogramming to NSCs. By this review, we have also introduced different combinations of these SMs, which can help generate NSCs/NPCs from different types of cells (Table 2). Taken together, this review may help to understand the roles of some SMs in NSC/NPC reprogramming and the importance of some pathways for NSC/NPC generation (Fig. 1). With a better understanding of the reprogramming mechanisms and the common features of these SMs, it will be easier to find more efficient SMs and to generate human NSCs/NPCs, and finally, to use these cells for clinical treatments.
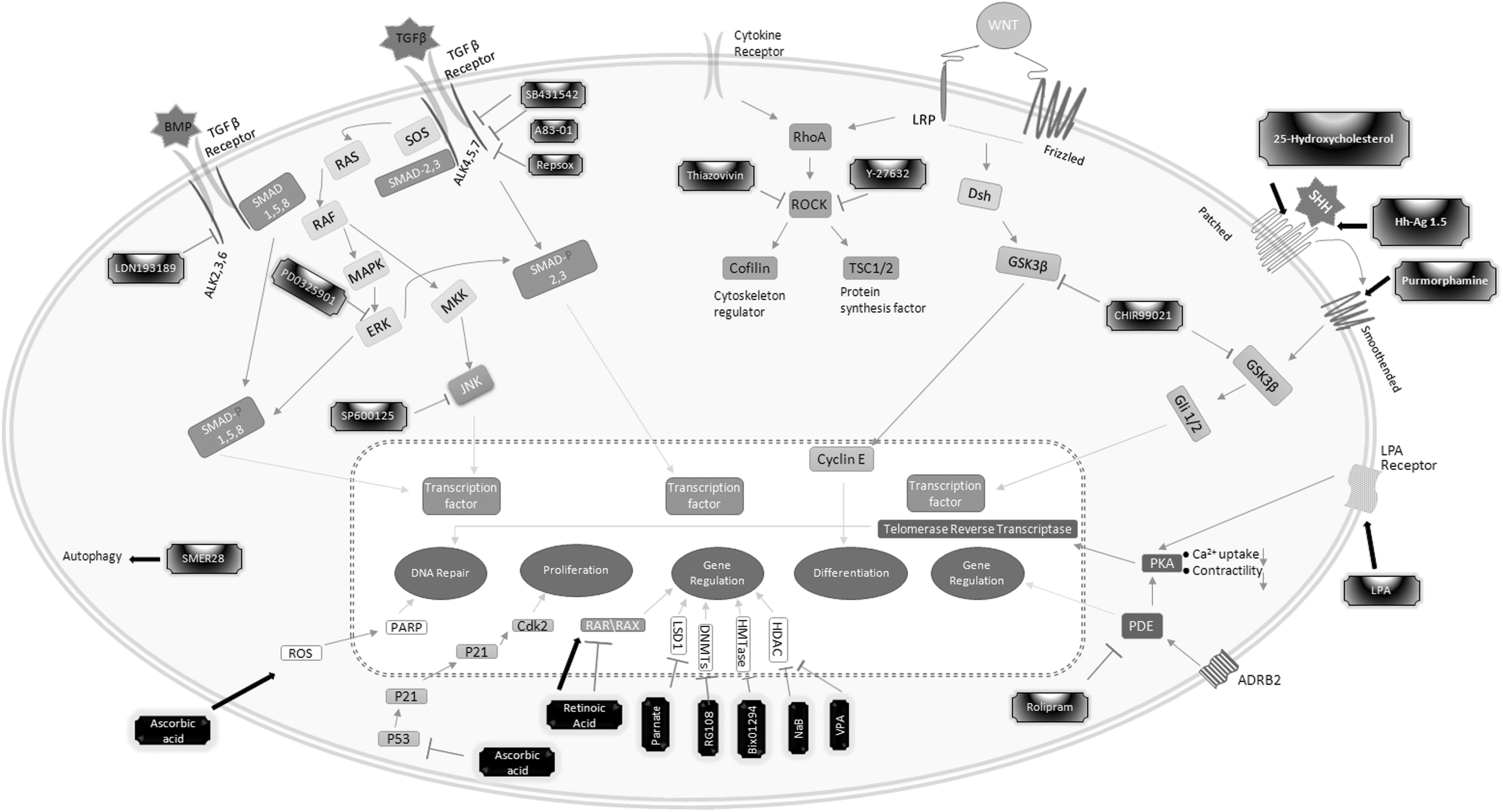
Small molecules reprogramming targets on somatic cells to induce NSC/NPC reprogramming. Black arrow represents modulating effect; T represents interfering effect. NPC, neural progenitor cell; NSC, neural stem cell.
Footnotes
Acknowledgments
This work was supported by D&R Pharmaceutics China, UniSA president scholarship to Nimshitha Pavathuparambil Abdul Manaph, and NHMRC fellowship to Xin-Fu Zhou.
Author Disclosure Statement
No competing financial interests exist.