
Abstract
Corneal blindness is the third leading cause of blindness in the world, and one of the main etiologies is dysfunction of the corneal endothelium. Current treatment of corneal endothelial disease is allogenic corneal transplantation, which is limited by the global shortage of donor corneas and immunological rejection. The corneal endothelium consists of a monolayer of cells derived from the neural crest and mesoderm. Its main function is to prevent corneal edema by tight junctions formed by zonular occludens-1 (ZO-1) and Na, K-ATPase pump function. The human umbilical cord (UC) is a rich source of mesenchymal stem cells (MSCs). UC-MSCs that have multi-lineage potential may be an accessible allogenic source. After inducing differentiation with medium containing glycogen synthase kinase (GSK) 3-β inhibitor, UC-MSCs formed polygonal corneal endothelial-like cells that functioned as tissue-engineered corneal endothelium (UTECE). Expressions of major corneal endothelial markers were confirmed by reverse transcription–polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR). Western blotting confirmed the expression of Na,K-ATPase and PITX2, the functional and developmental markers of corneal endothelial cells. Immunohistochemistry revealed the localization of Na,K-ATPase and ZO-1 in cell–cell junctions, suggesting the presence of tight junctions. In vitro functional analysis revealed that UTECE had significantly high pump function compared with UC-MSCs. Moreover, UTECE transplanted into a rabbit model of bullous keratopathy successfully maintained corneal thickness and transparency. Our findings suggest that UTECE may be used as a source of allogenic cells for the treatment of corneal endothelial disease.
Introduction
C
Currently, 70% of corneal diseases contributing to blindness are due to post surgical bullous keratopathy and corneal dystrophy [4]. Fuchs corneal endothelial dystrophy, the most common corneal dystrophy, specifically manifests in the endothelial cell layer. Since corneal endothelial cells do not proliferate in vivo, corneal transplantation is the only therapeutic treatment available for corneal endothelial dysfunction [5]. The major problem of corneal transplantation is the shortage of healthy donor corneas, with an estimated 12.7 million people waiting for a corneal transplantation worldwide [6]. Transplantation is also associated with complications such as graft failure, immunological rejection, and secondary glaucoma [7,8]. To address these issues, researchers in the field are exploring alternative therapeutic strategies.
We previously reported that skin-derived precursors (SKPs) could be induced to differentiate into functional corneal endothelial cells [9]. However, clinical application of SKPs may prove difficult when the source of cells is from an aged donor. Age-related changes such as decreased growth and differentiation capacity may be a problem, since an autologous source was intended due to the invasive nature of tissue procuration [10,11]. We therefore hypothesized that human umbilical cord mesenchymal stem cells (UC-MSCs) may be used as an alternative cell source for differentiation into corneal endothelial cells.
MSCs originate in the human embryo, and a subset of MSCs is reported to originate from the neural crest [12]. Compared with other sources of MSCs, UC-MSCs are a noninvasive source for allogenic use. MSCs in general are multipotent, with a low risk of teratoma formation compared with pluripotent stem cells. MSCs are also reported to have low immunogenicity with immunosuppressive properties [13]. UC-MSCs have multi-lineage differentiation potential into adipocytes, chondrocytes, osteocytes, neurons, and hepatocytes [14 –17]. Only two previous reports are related to the potential of umbilical MSCs to differentiate into the corneal endothelial phenotype. Joyce et al. showed that MSCs derived from human umbilical cord blood (hUCB-MSCs) could adhere to the denuded corneal endothelium and assume corneal endothelial cell-like phenotypes in an ex vivo culture model [18]. Shao et al. showed that transplantation of hUCB endothelial progenitor cells labeled with CD34 immunomagnetic nanoparticles can successfully repair corneal endothelium defects in a rabbit model [19]. However, none of these reports have tried to differentiate UC-MSCs into corneal endothelium in vitro.
The corneal endothelium originates from neural crest cells that migrate to the cornea during embryogenesis [20,21]. Expression of the homeobox gene, Pitx2 is required for ocular anterior segment development [22]. In this study, we first demonstrated how tissue-engineered corneal endothelium derived from UC-MSCs exhibit corneal endothelial gene and protein expression, including Pitx2. We then assessed whether UC-MSCs-derived tissue engineered corneal endothelium (UTECE) have Na,K-ATPase pump function equivalent to native corneal endothelium using the Ussing chamber [23]. Finally, we also show how UTECE maintains corneal transparency and thickness in a rabbit model of bullous keratopathy in vivo.
Materials and Methods
Cell culture
This study was approved by the Ethics Committee of Keio University School of Medicine, Tokyo, Japan (Approval Number: 20150209). Human mesenchymal stem cells derived from umbilical cord (UC-MSCs) were supplied by the Department of Cell Processing and Transfusion, Institute of Medical Science, University of Tokyo, Japan. The methods for UC-MSCs isolation are described in previous reports [24
–26]. Cells were fed with α-modified Eagle's medium (α-MEM; Invitrogen, Thermo Fisher Scientific, Carlsbad, CA,
Human corneal endothelial cell line (B4G12) immortalized by SV40 large T- and small T-antigens, were purchased from DSMZ (Braunschweig, Germany,
Flow cytometry analysis
Standard flow cytometry (FCM) techniques were used to determine cell surface epitope profiles (CD19, CD29, CD34, CD44, CD45, CD49d, CD73, CD90, CD105, CD106, CD146, CD166, CD271, Stro-1, and HLA-DR). Briefly, adherent cells were incubated for 30 min at room temperature with the following markers: mesenchymal cell marker antibody panel kit (anti-Stro-1, anti-CD90, anti-CD105, anti-CD106, anti-CD146, anti-CD166, anti-CD44, plus leukocytes markers; anti-CD19 and anti-CD45: Cat. No. SC017; R&D Systems, Minneapolis, MN,
Adipogenic, osteogenic, and chondrogenic differentiation
UC-MSCs were plated at a density of 2 × 105 cells in 35 mm dishes and incubated at 37°C in a humidified atmosphere of 5% CO2. Cells were fed every 2 days by completely replacing with α-modified Eagle's medium until the cultures reached confluence (3–5 days). At 100% confluence, we performed adipogenic differentiation by treating UC-MSCs with hMSC Adipogenic BulletKit (Cat. No. PT-3004; Lonza, Basel, Switzerland,
To investigate the osteogenic differentiation capacity of UC-MSCs, confluent UC-MSCs in 35 mm dishes were treated with hMSC Osteogenic BulletKit (Cat. No. PT-3002; Lonza). The medium was changed every 2–3 days. After 2–3 weeks, the cells were stained with Alizarin Red S (Cat. No. 01303-52; Nacalai tesque, Kyoto, Japan,
A pellet culture system was used to evaluate the capacity of UC-MSCs to differentiate to chondrocytes. In brief, 2.5 × 105 pelleted UC-MSCs were resuspended and cultured in hMSC Chondro BulletKit (Cat. No. PT-3003; Lonza) in 15-mL conical tubes with medium changes every 3 days. After 3–4 weeks in culture, the pellet was fixed with 4% formaldehyde, sliced, and stained with 0.05% Toluidine blue (Cat. No. 89640-5G; Sigma-Aldrich), one percent Alcian blue (CAS#33864-99-2), and Nuclear Fast Red (C.I. 60760; Merck, Darmstadt, Germany,
Sphere formation
UC-MSCs were plated at a density of 3 × 106 cells/dish in EZSPHERE™ 3D culture plates (Cat. No. 4020-900; AGC techno glass, Shizuoka, Japan,
Corneal endothelial differentiation
To induce endothelial cell differentiation, we modified our previous protocol [9,27]. Briefly, cell spheres were dissociated into single cells in Accumax (Innovative Cell Technologies), and then suspended at a cell density of 2 × 105 cells/cm2 to 35 mm dish with FNC Coating Mix (Cat. No. 0407; Athena Environmental Sciences, Inc., Baltimore, MD,
Immunocytochemistry
Immunocytochemical analyses of cultured cells were performed as described previously [27,28]. In brief, the cells were fixed at room temperature for 10 min in 4% formaldehyde in PBS. After three-time PBS washes, the specimens were incubated for 30 min in Morphosave (Cat. No. 250-010; Ventana Medical Systems, Inc., Tucson, AZ,
Reverse transcription–polymerase chain reaction analysis and quantitative reverse transcription–polymerase chain reaction analysis
Total RNA was purified with the use of RNeasy kit (Qiagen, Hilden, Germany,
Western blot analysis
Western blot analyses were performed as described previously [9,27]. UTECE were washed with PBS twice, and dissolved in a lysis buffer (M-PER; Thermo Fisher Scientific, Inc.) with a protein inhibitor cocktail (Thermo Fisher Scientific, Inc.). Western blot analysis was performed by using a standard protocol with primary antibodies for β-actin (Abcam, Cambridge, United Kingdom,
Measurement of Na,K-ATPase pump function
The pump function of confluent monolayers of various types of cells was measured with the Ussing chamber as described previously [9,23,27]. As described above (cell culture, sphere formation, and corneal endothelial differentiation section), B4G12, UC-MSC, and UTECE cells were cultured until confluent. All media were changed to the MEM supplemented with CaCl2 (1 mM), sodium pyruvate (1 mM), MEM amino acid (1 × ), and MEM essential vitamin mixture (1 × ) after cells became confluent, and cells were cultured additional 48 h.
The cells cultured on Snapwell inserts were placed in an Ussing chamber (EM-CSYS-2; Physiologic Instruments, San Diego, CA,
After continuously monitoring for at least 10 min until SCC reached a steady state, the cells on the Snapwell membrane were loaded with 1 mV currents through electrodes thrice, and the SCC change was recorded. When the SCC had reached a steady state, the specific Na,K-ATPase inhibitor ouabain (final concentration, 10 mM; Sigma-Aldrich) was added to the chamber, and the SCC was measured again. The arrow represents SCC changes before and after final ouabain addition (Fig. 4A). Trans-endothelial resistance (TER) was calculated from the average SCC change and loaded voltage according to Ohm's law. SCC was continually recorded during this procedure. The SCC attributable to Na,K-ATPase activity was calculated as the difference in SCC measured before and after the addition of ouabain. The arrow represents SCC changes before and after final ouabain addition (Fig. 3A). Finally, the SCC attributable to Na,K-ATPase activity was calculated as the potential difference (PD) by multiplying TER.
Animals
All animals were handled in full accordance with the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. This study was approved by the Committee for Tokyo Dental College Ichikawa General Hospital (Approval Number: 297605).
Transplantation of UTECE sheets into a rabbit model
Transplantation study performed as described previously [9,27], type I atelo collagen sheets (CM6; Koken) were used as carriers. Sphere-derived umbilical cord mesenchymal stem cells (UCSp) were plated and cultured on type I atelocollagen sheets and then induced to UTECE, and the collagen sheets with UTECE were cultured for 1 week. Before transplantation, cell densities of UTECE sheets were manually counted by microscope. Japanese white rabbits (female, 2.5 kg body weight; Shiraishi Experimental Animal Breeding Farm) were anesthetized intravenously with a mixture of medetomidine hydrochloricde (0.5 mg/kg; Domitor; Meiji Seika Kaisha, Tokyo, Japan.
Corneal buttons were prepared by 8.0-mm Barron donor cornea punch (Barron Precision Instruments) from the center of donor corneas purchased from Funakoshi Co., Ltd., and Descemet's membrane along with the entire endothelium was stripped from the corneal buttons. PKH26 (Cell staining kit; Sigma-Aldrich)-labeled UTECE sheets with carrier collagen sheets were harvested by same size punch and immediately placed on the stromal bed. Within 5 min, the sheets produced stable attachment to the stroma. The recipient central cornea was incised and opened by Hassburg-Barron Vacuum Trephine (Barron Precision Instruments, Grand Blanc, MI,
Six UTECE sheets with a cell density of 2034.65 ± 292 cells per mm2, six grafts without corneal endothelium, six UC-MSC with a cell density of 2164.33 ± 429 cells per mm2, and three normal corneas were transplanted to rabbit corneas by penetrating keratoplasty. Antibiotics (0.3% ofloxacin) and steroids (0.1% betamethasone) were applied topically three times a day.
After transplantation, eyes were carefully observed by slit-lamp microscopy, and serial photographs were obtained. The corneal edema was classified as described by Mimura: 0, completely transparent cornea; 1, minimal corneal opacity, but iris vessels easily visible; 2, moderate corneal opacity, iris vessels still visible; 3, moderate corneal opacity, only pupil margin visible; 4, complete corneal opacity, pupil not visible [29]. Central corneal thickness was measured with an ultrasound pachymeter (Tomey, Nagoya, Japan,
Finally, 8 days after surgery, rabbits were sacrificed (above-described anesthesia, 20 mg/kg thiopental sodium (Ravonal, Tanabe Seiyaku Co., Osaka, Japan,
Histological examination of rabbit eyes after transplantation of UTECE sheets
The enucleated eyes were excised and fixed in 4% paraformaldehyde for 18 h. The corneal paraffin sections were used for Hematoxylin and Eosin (HE) staining. Immunohistochemistry (IHC) staining performed automatically by BOND-MAX™ (IHC staining system, Leica Microsystems GmbH, Leica, Wetzlar, Germany,
Statistical analysis
Data are presented as means ± SD and were compared by Student's t-test or multiple t-test with Bonferroni correction after ANOVA with the use of Excel 2007 software (Microsoft corporation, Redmond, WA,
Results
UC-MSCs express MSC biomarkers
UC-MSCs were plastic-adherent cells, spindle-shaped. We confirmed UC-MSCs had the differentiation capacity of adipocytes, chondrocytes, and osteoblasts. The potential for UC-MSCs to undergo osteogenic differentiation, as demonstrated by Alizarin red S staining (Fig. 1A). An accumulation of oil red O–stained lipid drops were observed in UC-MSCs cultured in adipocyte differentiation medium (Fig. 1B). In chondrogenic differentiation, the pellet culture system was applied and elastic firm pellets with diameters of approximately 1.5 mm were observed. Furthermore, toluidine blue staining revealed the presence of extracellular matrix formation in histological sections obtained of cells grown in chondrogenic induction medium (Fig. 1C). Moreover, Alcian blue staining revealed the presence of proteoglycan and extracellular matrix (Fig. 1D). UC-MSCs are positive for CD44, CD166, CD29, CD105, CD73, CD90, CD146, and CD49d and negative for CD45, CD34, CD106, HLA–DR, Stro-1, CD271, and CD19 (Fig. 1E). Figure 1E shows that UC-MSCs were a heterogeneous cell group, where some negative cells were remaining in the population.
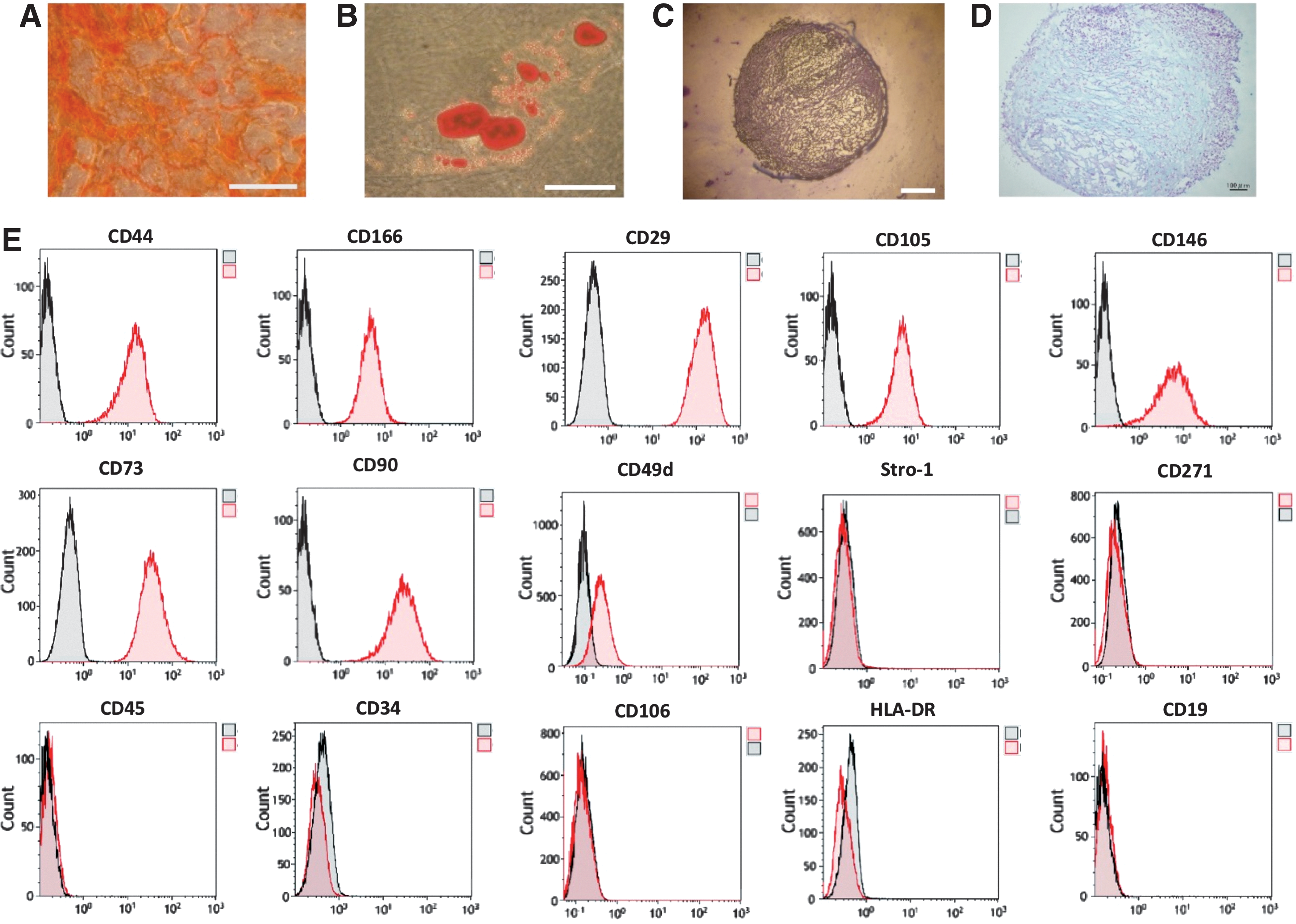
Characterization and differentiation into osteocyte, adipocyte, and chondrocytes of s (UC-MSCs).
UCSp and immunofluorescence staining of UCSp
When UC-MSCs were suspended in medium supplemented with EGF, bFGF, 2% MACS NeuroBrew-21 and GlutaMAX™ (Life Technologies) in EZSPHERE™ (AGC techno glass), UC-MSCs began to aggregate and form clusters defined as UCSp within 2–4 days (Supplementary Fig. S1A). The average diameter of spheres was 113.6 ± 12.7 μm and consisted of 3.9 ± 0.63 × 103 cells with more than 90% viable cells as determined by Trypan blue staining after dissociation. Expression of human neural crest markers ITGA4 (integrin subunit alpha4), SOX9 (SRY-box9), SOX10 (SRY-box10), SNAIL2 (snail family transcriptional repressor2), and TFAP2 (transcription factor AP-2 alpha) were shown by reverse transcription–polymerase chain reaction (RT-PCR) (Supplementary Fig. S1B). The expression of functional markers of corneal endothelial cells, ZO-1 (green), ATP1A (green), CDH2 (green), and neural crest markers SOX10 (green) were examined in spheres by immunostaining (Supplementary Fig. S1C). Nuclei were labeled with DAPI (blue). The results show that all of ZO-1, Na,K-ATPase, CDH2, and SOX10 were localized in the sphere.
Functional TECE induced from UCSp
We further used this protocol to differentiate UCSp into functional TECE. UCSp were cultivated and expanded similar to human SKPs reported previously [9]. UCSp could be maintained in sphere culture for at least a week. Immunohistochemical analysis confirmed positive staining of ZO-1 and ATP1A1 in UC-MSCs derived tissue engineered corneal endothelium UTECE (Fig. 2, middle panel), which were similar to control human corneal endothelial cell line (B4G12) (Fig. 2, lower panel). Human corneal endothelial markers ATP1A1 (P < 0.01), PITX2 (P < 0.01), SLC4A4 (P < 0.001), and COL4A2 (P < 0.05) were significantly upregulated after induction as shown by quantitative RT-PCR (Fig. 3B). Western blot analysis also showed that UTECE express significantly higher protein levels of ATP1A1 (P < 0.01) and PITX2 (P < 0.01) compared to before induction UC-MSCs (Fig. 3D).

Morphology of UC-MSC-derived TECE. Comparison of in vitro morphology, immunohistochemistry of ZO-1 (red) and Na,K-ATPase (green) in UC-MSC, UTECE, and B4G12. Original UC-MSC (top panel) showed spindle morphology in vitro. UTECE (middle panel) was similar to control human corneal endothelial cell line (B4G12) (bottom panel). Nuclei were labeled with DAPI (blue). Scale bars = 50 μm. UTECE, umbilical cord mesenchymal stem cells-derived tissue-engineered corneal endothelium.
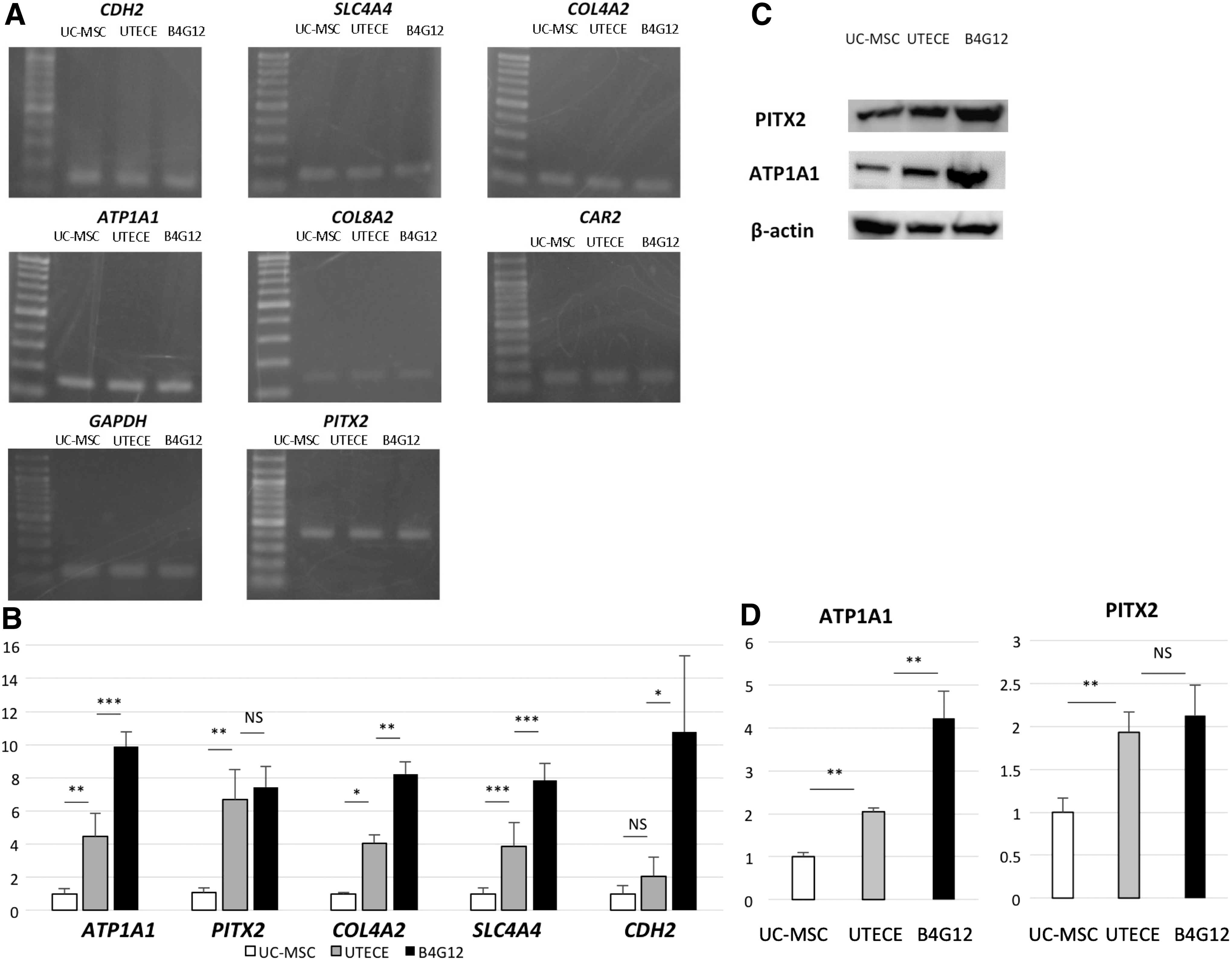
The expression of corneal endothelium markers by RT-PCR and western blots.
In vitro functional analysis of Na,K-ATPase of UC-MSCs-derived corneal endothelial cells revealed relatively high pump function
The Na,K-ATPase pump is one of the most important functions of the corneal endothelium. We therefore assessed whether UC-MSCs derived tissue engineered corneal endothelium UTECE has pump function comparable to human corneal endothelial cell line (B4G12). Figure 4A shows Na,K-ATPase pump function measured by Ussing chamber in cultured B4G12, UC-MSCs, and UTECE. Figure 4B shows the ouabain dependent SCC, which was calculated before and after addition of ouabain. SCC of UC-MSC (173.90 ± 31.39 μA/cm2) was larger than B4G12 cells (86.54 ± 16.79 μA/cm2) (P < 0.01) and SCC of UTECE (233.29 ± 21.44 μA/cm2) was larger than UC-MSC cells (P < 0.05). The ouabain-dependent PD in each cell type attributed to Na, K pump function is shown in Fig. 4C, which basically correlates with Fig. 4B. The ouabain-dependent PD of UC-MSC (11.79 ± 3.68 mV/cm2) was larger than B4G12 cells (3.96 ± 0.64 mV/cm2) (P < 0.01) and the ouabain-dependent PD of UTECE (17.25 ± 2.95 mV/cm2) was larger than UC-MSC (P < 0.05). TER revealed no significant changes among UC-MSCs (13.77 ± 2.71 Ω/cm2) and UTECE (12.43 ± 2.88 Ω/cm2) (P = 0.26) in Fig. 4D.
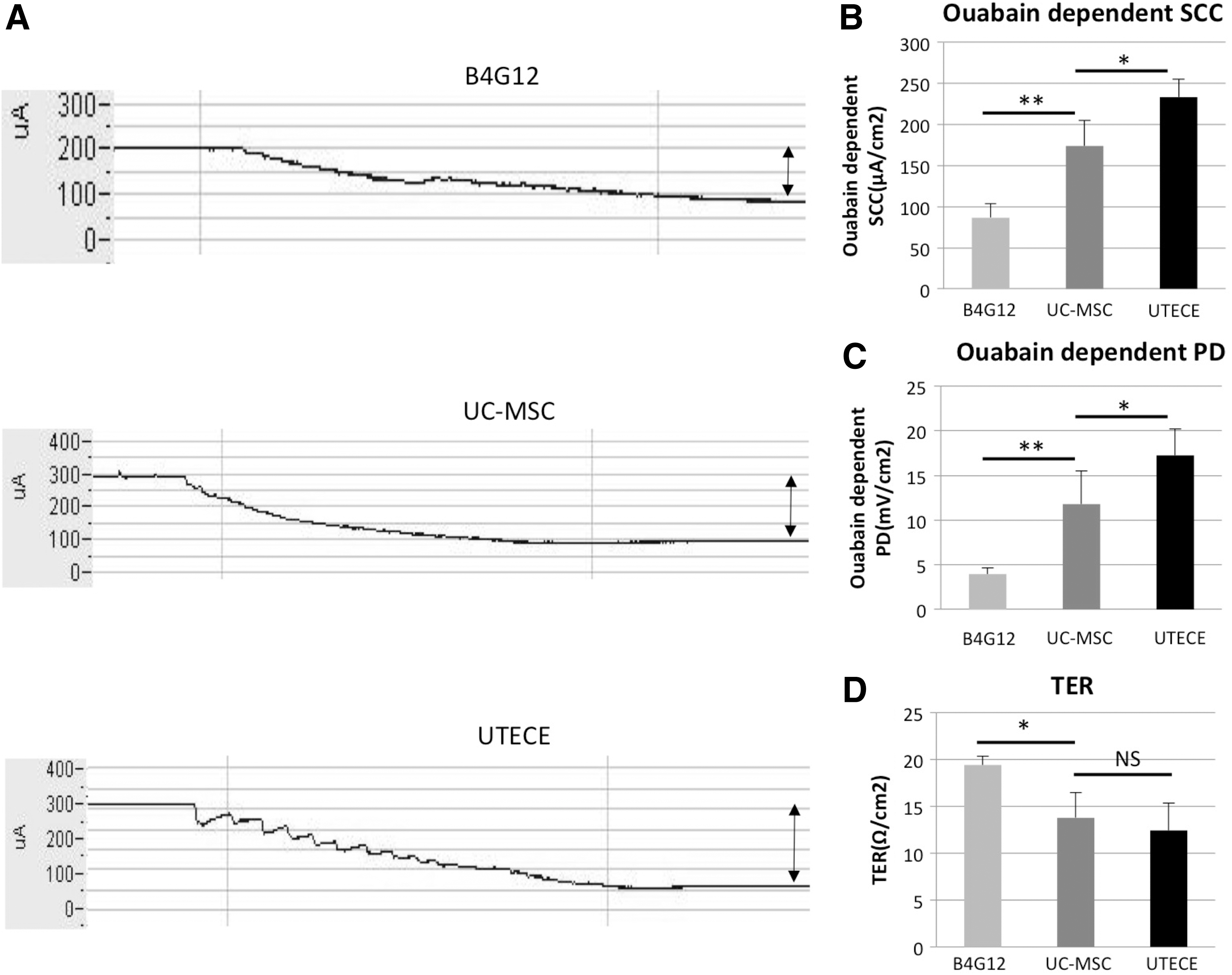
Measurement of pump function by the Ussing chamber system.
In vivo transplantation of UC-MSC-derived corneal endothelium recovered corneal transparency in a rabbit model of bullous keratopathy
Figure 5A shows slit lamp photographs of rabbit eyes 1, 2, 5, and 8 days after transplantation. Corneas transplanted with UTECE were less edematous and maintained transparency (Fig. 5A: lower panel), whereas eyes grafted without corneal endothelium suffered severe corneal edema and thickened stroma (Fig. 5A: upper panel). Figure 5B shows corneal thickness change in each group. During 8 days after surgery, corneal thickness of eyes transplanted with grafts lacking corneal endothelium increased over 1,000 μm (1139.97 ± 137.92 μm) and showed severe corneal cloudiness consistent with bullous keratopathy, whereas UTECE-transplanted eyes maintained significantly lower corneal thickness (566.39 ± 118.01 μm) (P < 0.001). Figure 5C shows changes in intraocular pressure (IOP) during the observation period. There was no significant difference among all group 8 days after transplantation. Average IOP of UTECE-transplanted eyes (19.7 ± 9.0 mmHg) was not significantly different from eyes transplanted with grafts lacking corneal endothelium (18.9 ± 5.7 mmHg) and UC-MSC-transplanted eyes (17.5 ± 5.1 mmHg). Figure 5C indicates that the difference in corneal thickness was due to UTECE pump function, and not IOP. There was no difference between UC-MSC and UTECE (P = 0.95) using Mimura's corneal opacity score [29].
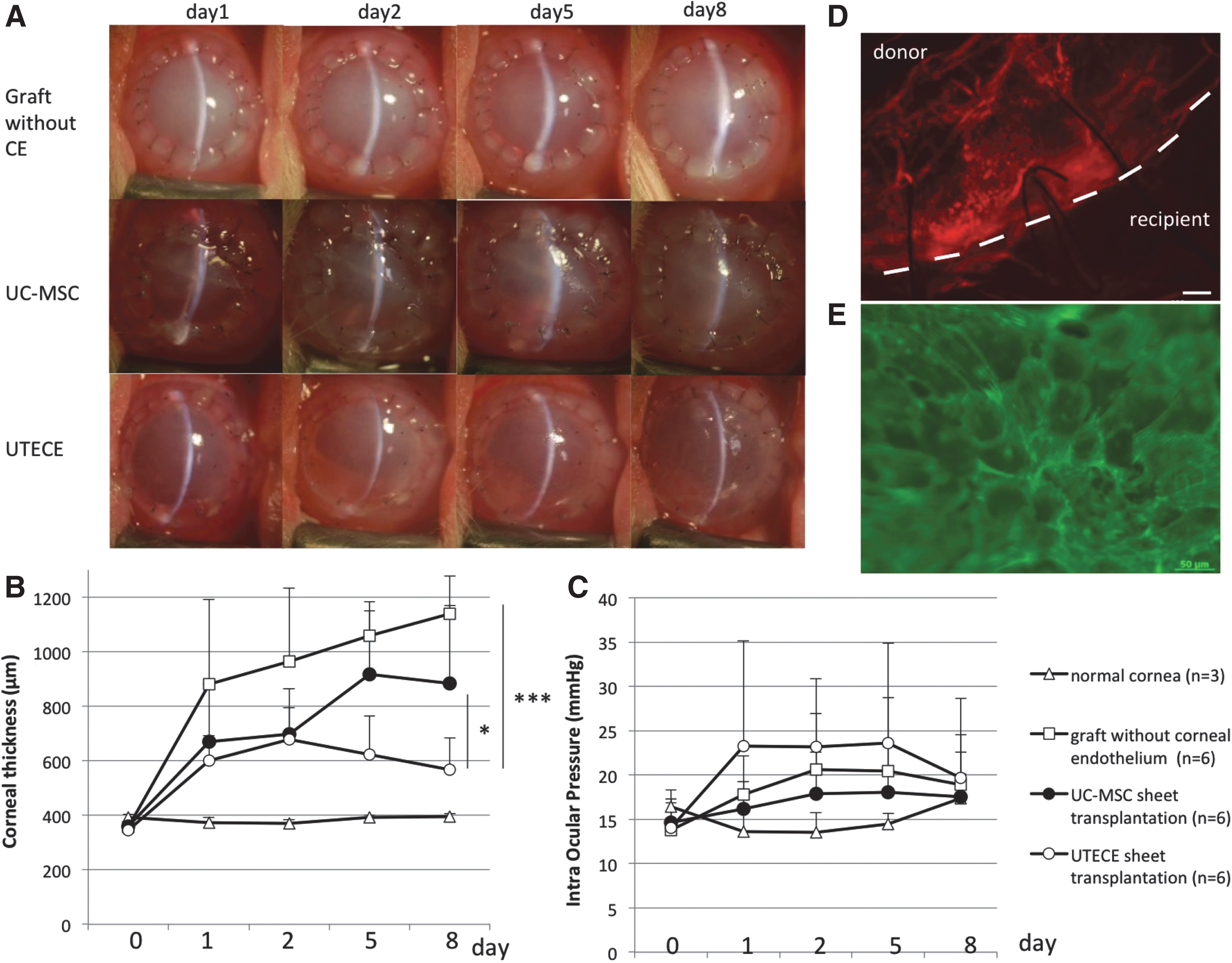
In vivo transplantation of UTECE to rabbit cornea.
Immunocytochemistry and histological examination after transplantation
Figure 5D and E shows whole mount photographs of UTECE-transplanted corneas. PKH derived florescence was detected only in the transplanted site inside the host-graft junction at 8 days after transplantation (Fig. 5D). Figure 5E shows UTECE stained with phalloidin arranged as polygonal cells similar to the endothelial phenotype. Hematoxylin and Eosin (HE) staining also revealed that confluent UC-MSCs in the UTECE group were attached firmly to the type I atelocollagen sheet (Supplementary Fig. S2A). No inflammatory cells or neovascularization were observed (Supplementary Fig. S2B). Grafts stripped of endothelium had no corneal endothelial cells and the Descemet's membrane was also missing. Eyes transplanted with grafts without corneal endothelium showed marked corneal stromal edema. The normal cornea expressed corneal endothelial monolayers.
Discussion
Currently, the precise molecular pathways that govern development and differentiation of corneal endothelial cells are still not clear. Several groups have reported the in vitro and in vivo differentiation of corneal endothelium from different stem cell sources. Zhao et al. generated a renewable supply of corneal endothelial cells from human pluripotent stem cells [30]. Other stem cell sources such as ES cells [31 –33], iPS cells [34], umbilical cord blood cells [18], and umbilical cord blood endothelial cells [19] also seem promising. Our group previously reported derivation of functional tissue-engineered corneal endothelium from cornea-derived precursors [27,28] and SKPs [9]. These cell sources are similar to MSCs in that they are multipotent precursors [14]. MSCs share a developmentally common origin with corneal endothelium, both of which come from the neural crest [12]. Applying our previous protocol using Wnt/β catenin signaling, we successfully differentiated functional corneal endothelial cells from UC-MSCs. It is worth noting that MSCs themselves are weakly immunogenic and show immunomodulatory properties. [35 –37]. In addition, MSCs have advantages for the ability to “home” to areas of corneal endothelial injury [18]. UC-MSCs can be obtained from residual tissue during child birth. Since it is not necessary to perform transgenic procedures, ethical issues such as using embryonic stem cells should not be a problem. Therefore, UC-MSCs may prove to be an accessible allogenic source for potential use in stem cell therapy of the corneal endothelium.
UTECE expressed a series of major corneal endothelial markers by RT-PCR. Of particular note was the upregulation of Pitx2 mRNA and protein after endothelial induction measured by qPCR and western blots. Pitx2 plays a crucial role in ocular development [22], along with signaling crosstalk of Wnt/β catenin. Our culture protocol shows that UTECE derivation mimics this developmental stage. However, long-term observations are necessary to confirm that corneal clarity is not compromised. We further demonstrated that UTECE functioned in an in vivo model of bullous keratopathy in rabbits to recover corneal transparency. As far as we know, this is the first report on the efficacy of human UC-MSC-derived corneal endothelium in vivo.
Our data show that UTECE had sufficient pump function after transplantation into the eye. Furthermore, despite the fact that this was xenograft transplantation from human to rabbit, examinations showed no signs of immunological rejection, or neovascularization during the observation period. Secretion of trophic factors may be another mechanism by which UTECE confers regenerative effects. Two previous reports demonstrated unilateral intrastromal injection of human UC-MSCs to treat lumican null mice that manifest thin and hazy corneas due to null expression of lumican, reduced expression of keratocan, and disrupted corneal stromal extracellular matrix structure [38]. Intrastromal injection of UCMSCs in mucopolysaccharidosis VII mice resulted in the restoration of the dendritic and hexagonal morphology of host keratocytes and endothelial cells, respectively [39].
It is worth mentioning that these corneas had almost no immune rejection after UTECE transplantation. This may be due to three reasons. First, UC-MSCs do not elicit acute rejection, and are suitable for allogeneic cell-based therapy. UC-MSCs lack HLA-DR, CD80, and CD86, which are costimulatory molecules inducing T cell activation and survival [35–37]. The immunosuppressive effect of UC-MSCs is mediated by soluble factors and cell-to-cell contacts. PGE2, galectin-1, and HLA-G5 are released from MSCs and serve as effective factors of immunosuppression [40]. Second, the anterior chamber is characterized with anterior chamber-associated immune deviation (ACAID), permitting the long-term acceptance of a graft [41,42]. Third, we used serum-free medium to culture UCSp and UTECE, avoiding many immunogenic factors. Taken together, UTECEs have weak immunogenicity with no need for large amounts of immunosuppressive drugs.
One limitation of our study is the use of a rabbit model of bullous keratopathy for in vivo experiments. We used this model due to the established surgical technique and evaluation protocol used in a previous study [27]. The rabbit has a high inbuilt capacity for corneal endothelial proliferation in vivo [43], and therefore, long-term observation is not possible. However, the objective of this data was to show in vivo function using corneal thickness as a parameter, and to this end, we were successful in showing pump function in vivo.
Some investigators consider sheet transplantation of HCECs as the optimal method for endothelial transplantation, as it retains the polarity of the transplanted endothelial cells [44]. Shao et al. showed that anterior chamber injection of immuno-magnetic infused hUCB endothelial progenitor cells to repair corneal endothelium defect successfully in rabbit model [19]. However, the viability, identity, and barrier function of nanoparticle-endocytosing transplanted cells, and the long-term safety of these particles still need to be confirmed. There are reports of teratogenicity from the use of magnetic nanoparticles in animal models [45].
Another limitation of our study is that the mechanism of transplanted UTECE has not been fully elucidated. UTECE decreased corneal opacity due to bullous keratopathy and restored early corneal endothelial function in our rabbit model. UTECE improved early repair through tight engraftment on the atelocollagen sheet, and differentiation to corneal endothelial like cells. We found that the size of UTECE after 8-days transplantation were larger than B4G12 by microscopy. These observations suggest that our protocol may also aid in the repair of dysfunctional corneal endothelial cells through paracrine factors released by MSCs.
UC-MSCs are promising as a possible versatile tool for regenerative medicine and immunotherapy [13]. Recently clinical trials have been conducted using UC-MSCs for neurogenic disorders (spinocerebellar ataxia and multiple system atrophy of the cerebellar type) [46] and liver disorders [47,48]. However, further optimization of isolation and culture techniques of UC-MSCs are required before clinical use. The cornea is an ideal tissue to apply cell injection therapy due to the small number of cells required. Furthermore, the transparent nature of the cornea will allow direct visualization of transplanted cells, and quick intervention in the case of unexpected events.
Conclusion
We successfully induced functional corneal endothelial-like cells from UC-MSCs that showed sufficient pump function in a rabbit model of corneal endothelial dysfunction. Our findings suggest that UC-MSCs may be used as a source of allogenic cells for the treatment of corneal endothelial disease.
Footnotes
Acknowledgments
We thank Dr. Tokiko Nagamura-Inoue from the Institute of Medical Science, University of Tokyo, for suppling mesenchymal stem cells derived from human umbilical cord. We thank Tomomi Sekiguchi, Miyuki Yasuda, and Hiroko Niwano for technical assistance and Dr. Satoru Yoshida for general support. We thank Shigeru Saito from Beckman Coulter for technical assistance.
Author Disclosure Statement
This work was supported by grants from the ROHTO Pharmaceutical Co., Ltd. and the Ministry of Education, Culture, Sports, Science and Technology of Japan (MEXT). (Grant-in-Aid for Young Scientists(B) 050451155). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.