
Abstract
The stem cell compartment of the hematopoietic system constitutes one of the most radiosensitive tissues of the body and leukemias represent one of the most frequent radiogenic cancers with short latency periods. As such, leukemias may pose a particular threat to astronauts during prolonged space missions. Control of hematopoiesis is tightly governed by a specialized bone marrow (BM) microenvironment/niche. As such, any environmental insult that damages cells of this niche would be expected to produce pronounced effects on the types and functionality of hematopoietic/immune cells generated. We recently reported that direct exposure of human hematopoietic stem cells (HSC) to simulated solar energetic particle (SEP) and galactic cosmic ray (GCR) radiation dramatically altered the differentiative potential of these cells, and that simulated GCR exposures can directly induce DNA damage and mutations within human HSC, which led to leukemic transformation when these cells repopulated murine recipients. In this study, we performed the first in-depth examination to define changes that occur in mesenchymal stem cells present in the human BM niche following exposure to accelerated protons and iron ions and assess the impact these changes have upon human hematopoiesis. Our data provide compelling evidence that simulated SEP/GCR exposures can also contribute to defective hematopoiesis/immunity through so-called “biological bystander effects” by damaging the stromal cells that comprise the human marrow microenvironment, thereby altering their ability to support normal hematopoiesis.
Introduction
A
Considering the various malignancies that could potentially be induced/arise as a result of exposure to GCR and SEP radiation, leukemias represent one of the most frequent radiogenic cancers [12]. Moreover, in contrast to many solid tumors, leukemias have a short enough latency period [12] to pose a particular threat to astronaut health and mission success during prolonged missions. Compounding the carcinogenic risks that could arise from chronic SEP/GCR exposures are numerous studies collectively demonstrating that extended spaceflight conditions deleteriously affect the immune system at multiple levels [13 –29] and impair astronauts' ability to respond to infection or immune challenge [30 –36]. Collectively, these findings illustrate an important need to use appropriate human hematopoietic experimental models to precisely identify SEP/GCR radiation-induced effects, namely to better understand the genomic and epigenomic alterations responsible for low or high linear energy transfer (LET) charged particle-induced carcinogenesis; identify appropriate molecular targets for effective countermeasure development; and provide more refined datasets for NASA's risk estimation modeling efforts.
Leukemias are hematological malignancies that arise when hematopoietic stem/progenitor cells (HSC/HPC) suffer genetic damage that triggers subsequent malignant transformation, arresting them at a specific stage of lineage commitment and precluding their ability to complete differentiation, to yield mature/functional blood/immune cells. All mature hematopoietic cells present in the circulation at any given time are generated from HSC through the process of hematopoiesis, which occurs in a unique microenvironment/niche in the bone marrow (BM). This niche both maintains the primitive HSC and when required, induces their differentiation to specific lineages as the need arises for those particular cell types [37 –41]. The tightly regulated process of hematopoiesis involves interactions between HSC, extracellular factors, and the stromal cells that comprise the hematopoietic niche. These interactions have been the focus of intense investigation in recent years, and have been defined in terms of cells, extracellular matrix, soluble factors, and signaling pathways [38 –43].
With the exception of macrophages, all of the cells that comprise the BM niche are derived from mesenchymal stem cells (MSC) [44]. MSC thus play a highly critical role in the control/regulation of hematopoiesis. In addition, recent studies have suggested that MSC may play a key role in suppressing the growth/expansion of hematopoietic clones that have undergone leukemic transformation, functioning as sentinels against hematologic malignancies [45,46]. As such, any insult that damages MSC would be expected to produce pronounced downstream effects on the types and functionality of hematopoietic/immune cells generated, as well as increase the risk of leukemogenesis. However, little is known about the effects of space radiation on human MSC stability/function, what changes occur in the BM niche as a result of spaceflight, and how these changes may impact upon hematopoiesis, immune function, and astronaut risk of leukemia.
We recently reported that simulated SEP/GCR radiation dramatically alters the functionality and differentiative potential of human HSC and acute/high dose-rate mission-equivalent GCR exposures can directly induce DNA damage and mutations within human HSC, which led to leukemic transformation when irradiated cells from two donors were used to repopulate immunodeficient mice [47]. In this study, we performed the first in-depth examination of the effects of mission-relevant doses of SEP/GCR radiation on human MSC, which are likewise essential for normal hematopoiesis. Our data provide compelling evidence that SEP/GCR irradiation of MSC can contribute to defective HSC hematopoiesis/immunity through so-called “biological bystander effects,” by altering/damaging stromal cells that comprise the human BM microenvironment.
Materials and Methods
Isolation and culture of MSC from human BM
Adult human BM mononuclear cells from five different healthy donors (aged 18–45) were purchased from AllCells, LLC (Alameda, CA) or Stem Cell Technologies, Inc. (Vancouver, Canada). Cells received from both commercial sources were of similar viability (>90% viable). Upon thawing, the cells were subjected to magnetic sorting (MiniMacs; Miltenyi Biotec, Bergisch Gladbach, Germany) to obtain the Stro-1+ population, which is highly enriched for MSC with high proliferative and differentiative capacity [48 –55]. Following isolation, Stro-1+ MSC were expanded in culture and confirmed to meet the criteria of MSC by flow cytometric analysis and differentiation capability into bone, cartilage, and adipocytes, as we have previously described [49,50,56 –60]. Briefly, Stro-1+ MSC were maintained in gelatin-coated Falcon® tissue culture flasks (VWR, Radnor, PA) using MSC growth media (MSCGM; Lonza, Walkersville, MD) in a humidified 37°C incubator at 95% air/5% CO2 mixture. Once cells had reached passage 3–4 and were 50%–60% confluent in nonvented T25 flasks, the flasks were filled completely with MSCGM (Lonza), and the lids closed tightly and sealed with Parafilm® (SigmaAldrich, St. Louis, MO). The flasks were then carefully packed in an insulated box and shipped through FedEx First Overnight® for first morning delivery to the Brookhaven National Laboratory (BNL) Biomedical building at BNL (Upton, NY) early the following morning.
Thawing and shipping of human BM-derived HSC
Frozen human CD34+ cells isolated from healthy adult donors of typical astronaut age (30–55 years) were purchased from AllCells, LLC, or Stem Cell Technologies, Inc. and used for these experiments. Cells received from both commercial sources were of similar purity (>90% CD34+ cells) and viability (>90% viable). On the day before the scheduled irradiations at the NASA Space Radiation Laboratory (NSRL) at BNL, human CD34+ cells were thawed using procedures to maximize recovery and viability. In brief, the thawing medium consisting of RPMI (Gibco™; ThermoFisher Scientific, Grand Island, NY) containing 30% fetal bovine serum (Lonza), and supplemented with 20 U/mL heparin, penicillin/streptomycin (100 U/mL), and
Exposure of human HSC to simulated SEP and GCR radiation
Upon receipt at BNL, the HSC cryovials were maintained in a sterile 37°C incubator in the BNL Medical building and transferred to the NSRL cell culture facility ∼1 h before the scheduled irradiation time. After the NSRL physicists completed calibration and dosimetry of the NSRL proton and 56Fe ion beams, the HSC cryovials were placed upright in custom foam holders and irradiated at room temperature with 1 Gy of 50 MeV protons (LET = 1.26 keV/μm) at a dose rate of 50–100 cGy/min. Doses were measured by the NSRL physicists at the approximate cell position within the flasks using an NIST-traceable tissue-equivalent ion chamber (EG&G model IC-17) used to calibrate a series of custom parallel-plate beamline ionization chambers to control beam delivery. The NSRL ion source was then immediately switched to deliver iron ions, tuning/dosimetry was again completed, and 15 min following the completion of the proton irradiations, additional HSC cryovials were irradiated with 20 cGy 56Fe ions (LET = 151.4 keV/μm) at a dose rate of 20–40 cGy/min. Some samples received sequential doses of protons and 56Fe ions (dual ion exposures) to better simulate a more typical deep space mixed-field exposure scenario. After irradiation, cells were immediately returned to a 37°C incubator. Another group of samples from each donor was irradiated with 1 Gy of 137Cs γ-irradiation (LET = 0.91 keV/μm) at a dose rate of 125 cGy/min using a J.L. Shepherd and Associates Mark I Model 68A located in the BNL Biomedical building. A final group of samples from each donor was also transported to the NSRL and the gamma source to serve as unirradiated (sham irradiated) controls. At the completion of all irradiations, the cryovials were securely packaged up for overnight return shipping back to Wake Forest Institute for Regenerative Medicine (WFIRM) through FedEx First Overnight service.
Upon receipt at WFIRM, cells in each cryovial were washed with 10 mL of fresh QBSF-60 serum-free media (Quality Biologicals, Inc.), collected by centrifugation at 300 g for 10 min, counted with a hemocytometer, and their viability assessed by Trypan Blue exclusion according to standard methods. To assess the in vitro functionality/colony-forming potential of HSC following exposure to each irradiation scheme, we performed Hematopoietic/Hemotoxicity Assay through Luminescence Output (HALO) assays, as detailed below.
Exposure of human MSC to simulated SEP and GCR radiation
Upon receipt at BNL, the MSC flasks were inspected and maintained in a sterile 37°C incubator in the BNL Biomedical building before being transferred to the NSRL cell culture facility ∼1 h before the scheduled irradiation time. After NSRL physicists calibrated and confirmed the dosimetry of the NSRL 50 MeV proton and 1 GeV/n 56Fe ion beams, the MSC flasks were placed upright in custom foam holders with the cell monolayer oriented closest to the beam and irradiated at room temperature with 100 cGy of 50 MeV protons (LET = 1.22 keV/μm) at a dose rate of ∼50–100 cGy/min. Doses were measured by the NSRL physicists at the approximate cell position within the flasks using an NIST-traceable tissue-equivalent ion chamber (EG&G model IC-17) used to calibrate a series of custom parallel-plate beamline ionization chambers to control beam delivery. The NSRL ion source was then immediately switched to deliver 56Fe ions, tuning/dosimetry was again completed, and 15 min following the completion of the proton irradiations, additional MSC flasks were irradiated with 20 cGy 56Fe ions (LET = 151.4 keV/μm) at a dose rate of ∼20–40 cGy/min. Some samples received sequential doses of protons and 56Fe ions (dual ion exposures) to better simulate a more typical deep space mixed-field exposure scenario. After irradiation, cells were immediately returned to a 37°C incubator. Another group of samples from each donor was irradiated with 100 cGy of 137Cs γ-rays (LET = 0.91 keV/μm) at a dose rate of 125 cGy/min using a J.L. Shepherd and Associates Mark I Model 68A cabinet irradiator located in the BNL Biomedical building. A final group of samples from each donor was also transported to the NSRL and the gamma source to serve as unirradiated (sham irradiated) controls. At the completion of all irradiations (and deactivation of NSRL-exposed samples), the sealed flasks were securely packaged up for overnight return shipping back to WFIRM through FedEx First Overnight service.
Cytokine arrays
We collected MSC culture supernatants from each donor 48 h following exposure to each type of radiation scheme, using MSC culture supernatant flown to BNL and not exposed to radiation as donor-specific controls. These supernatants were analyzed using a luminescence-based Human Cytokine Antibody Array (RayBio®, Norcross, GA) that allows simultaneous quantitation of expression levels of 42 cytokines involved in hematopoiesis/inflammation/immune function, according to the manufacturer's instructions. Uncompressed TIFF images of luminescence intensities for each blot were captured using a LAS3000 Luminescent Image Analyzer (FujiFilm, Valhalla, NY). We then used ImageJ software (
RNA isolation and analysis
RNA was isolated from MSC aliquots at 48 h postirradiation, and from another series of aliquots after the MSC had undergone five passages in tissue culture. RNA isolation was performed using a commercial kit (RNeasy Mini) according to the manufacturer's instructions (Qiagen, Germantown, MD). To ensure sufficient RNA was available for analysis, we subjected the samples to global, unbiased mRNA amplification using the QuantiTect Whole Transcriptome Kit according to the manufacturer's instructions (Qiagen). All samples were then quantitated and subjected to quality control checks on a 2100 Bioanalyzer (Agilent, Santa Clara, CA). RNA samples from the various MSC populations were then labeled with a commercial kit (TargetAmp™-Nano Labeling Kit; Lucigen, Madison, WI) and analyzed by microarray (Illumina® Expression BeadChip®, San Diego, CA). Following hybridization, washing, and scanning, data were extracted from scanned images using Genome Studio Software (Gene Expression module; Illumina, Inc.) and processed for upload to gene expression analysis software.
Statistical analysis
Following normalization (quantile) and log transformation of the raw data, unsupervised hierarchical clustering and analysis of variance (ANOVA) were performed, using Qlucore Omics Explorer (Qlucore, Lund, Sweden), to generate principal component analysis (PCA) plots and heat maps. Individual pair-wise comparisons were performed using Student's t-test (fold change ≥1.5; P ≤ 0.05) and groups of differentially expressed genes were compared between groups using GeneSifter® Analysis Edition (Perkin Elmer).
Coculture of MSC with HSC and assessment of hematopoietic potential
MSC exposed to each irradiation scheme as well as same-donor, sham-irradiated controls were used as “feeder” layers in long-term cultures with human HSC, using an allogeneic coculture system we previously described in detail [50,58,60]. In brief, upon return from BNL, MSC from each irradiation group were overlaid with human HSC (CD34+ BM-derived cells from healthy adult donors, purchased from AllCells, LLC, or Stem Cell Technologies) that were either unirradiated or had been exposed to the identical irradiation scheme as their respective MSC feeder layer. To allow maintenance of the HSC, MSCGM in the cocultures was supplemented with SCF (100 ng/mL), Fms-related tyrosine kinase 3 ligand (FLT3-L; 100 ng/mL), basic fibroblast growth factor (5 ng/mL), and leukemia inhibitory factor (10 U/mL). All cytokines were purchased from Peprotech (Rocky Hill, NJ). On day 3, 2.5 mL of fresh cytokine-supplemented media was added to each flask to replenish the cytokines. After a total of 7 days of coculture, nonadherent hematopoietic cells were harvested from the flasks and viable cells enumerated using Trypan Blue exclusion.
HALO® assay
To assess the in vitro functionality/colony-forming potential of HSC present in each coculture, we performed a highly sensitive, high-throughput ATP bioluminescence proliferation assay for lympho-HSC and HPC (HALO) [61], according to the manufacturer's instructions (HemoGenix®, Inc., Colorado Springs, CO). The HALO-96 Human Stem/Progenitor Cell Assay is a high-throughput, semiautomated assay that can simultaneously detect and quantitate the following seven different stem/progenitor cell populations at varying stages of differentiation: high proliferative potential colony-forming cell (HPP-CFC), colony-forming unit granulocyte, erythrocyte, monocyte, megakaryocyte (CFU-GEMM), colony-forming unit granulocyte, monocyte (CFU-GM), blast-forming unit erythroid (BFU-E), megakaryocyte colony-forming cell (CFC-Mk), T-lymphoid colony-forming cell (CFC-T), and B-lymphoid colony-forming cell (CFC-B). Instead of requiring colonies of cells to be identified visually and enumerated, HALO relies upon advanced, ATP-based, luciferin/luciferase bioluminescence readout. The assay is highly reproducible, eliminates the inherent subjectivity of visually scored methylcellulose assays, and is extremely sensitive, requiring roughly two orders of magnitude fewer input cells.
To perform the HALO assay, to a HALO master mix tube containing 0.8 mL of the appropriate growth factor combination (using manufacturer-provided premade master mixes with optimized cytokines/concentrations for each specific colony type to be assayed), ∼200 μL of CD34+ cell suspension was added for a final cell concentration of 10,000 cells/mL. The contents of each tube were mixed by vortexing, and the tube was allowed to settle for a minute. One hundred microliters of the master mix containing cells was dispensed into individual wells of 96-well culture plates in 6–8 replicates, such that ∼1,000 cells were deposited into each well. The culture plates were placed in a sterile 37°C incubator supplied with atmosphere of 95% air/5% CO2 mixture. After 6–7 days of culture, 100 μL of the ATP Enumeration Reagent [prepared fresh according to the manufacturer's instructions (HemoGenix, Inc.)] was added to each well using a multichannel pipette, and the contents of each well were mixed well by repeated pipetting. The plate was then allowed to incubate at room temperature in the dark for 15 min, after which the bioluminescence of each well was measured with a Veritas microplate luminometer (Turner BioSystems, Sunnyvale, CA) and the data exported directly into a Microsoft Excel spreadsheet (Microsoft Corp.) for analysis.
Statistical analysis for HALO assays
All data are presented as the mean ± standard error of the mean (SEM), and were analyzed with Prism 6 (GraphPad Software, Inc., La Jolla, CA). Statistical significance of differences observed between the various experimental conditions was determined using two-way ANOVA followed by the Bonferonni-Šidák correction for multiple comparisons. For all analyses, P ≤ 0.05 was considered to be statistically significant.
Results
SEP/GCR irradiation alters hematopoiesis-supporting ability of human MSC
We recently reported that SEP/GCR radiation exerts marked effects directly on both primitive and more differentiated hematopoietic cells [47]. Recent studies have provided evidence that the genomic instability induced by ionizing radiation can also lead to alterations within cells in the vicinity of the irradiated cells through so-called bystander effects, even if exposures occurred up to 35 cell doublings in the past [62,63]. Given the close proximity and intimate relationship between the HSC and MSC of the marrow microenvironment/niche, we hypothesized that exposure of MSC to SEP/GCR would induce alterations in these critical microenvironmental cells, which could then result in bystander damage to nearby HSC.
To begin testing whether SEP/GCR exposures of cells comprising the hematopoietic niche of the human BM would produce negative effects on human hematopoiesis, we made use of an allogeneic MSC-based coculture system we developed and previously described in detail [50,58,60]. This system is based on the use of a “feeder” cell layer comprising the Stro-1+ MSC population from human BM to support the maintenance and differentiation of human HSC (CD34+ BM-derived cells from healthy adult donors, purchased from AllCells, LLC, or Stem Cell Technologies). We chose this particular MSC population because we have a great deal of experience with these cells; know from prior work that these cells support long-term hematopoiesis; and have confirmed the multipotentiality of this population both in vitro and in vivo using the fetal sheep model system [49,56,57,64 –67]; and because this population is known to be highly enriched for MSC with high proliferative and differentiative capacity [48 –55,57,68].
Since we wished to compare both the direct effects of SEP/GCR radiation on HSC and the indirect/biological bystander effects mediated through SEP/GCR irradiation of MSC, and determine whether these direct/indirect effects might act in an additive or synergistic manner, we exposed both HSC and MSC (n = 5 different healthy human donors) to one of the 5 following irradiation schemes: (i) sham irradiation (cells flown to BNL and returned to WFIRM without irradiation); (ii) 100 cGy of 137Cs γ-irradiation; (iii) 100 cGy of 50 MeV protons; (iv) 20 cGy of 56Fe ions; and (v) sequential protons + 56Fe ions (ie, schemes 3 and 4 separated by 15 min), and then set up HSC + MSC cocultures under four different combinations for each irradiation scheme: (i) Unirradiated HSC plated over unirradiated MSC (ii) Unirradiated HSC plated over irradiated MSC (iii) Irradiated HSC plated over unirradiated MSC (iv) Irradiated HSC plated over irradiated MSC
Before discussing the results of the cocultures, it is important to note that the exposure of MSC to these low doses of radiation did not result in any marked changes in morphology or cell number, although absolute cell number is rather hard to quantify, as Stro-1+ MSC proliferate extensively in culture. However, the fact that we were able to propagate the MSC for five passages after radiation, and their doubling time did not differ significantly from that of their unirradiated counterparts, supports the conclusion that their viability/potency was not altered.
After 7 days of coculture, nonadherent hematopoietic cells were collected from the cultures and examined for their in vitro ability to form clonogenic progenitor colonies using the sensitive luminescence-based HALO assay system (Hemogenix). The HALO assay simultaneously detects and quantitates the following seven stem/progenitor cell populations at varying stages of differentiation: HPP-CFC, CFU-GEMM/Mix, CFU-GM, BFU-E, CFC-Mk, CFC-T, and CFC-B. In contrast to standard methylcellulose assays, which require colonies of cells to be visually identified and enumerated, the HALO assay relies upon direct ATP-based, luciferin/luciferase bioluminescence readouts, making the assay highly reproducible and extremely sensitive, and allows analyses to be performed with ∼2-logs fewer input cells than methylcellulose assays. To ensure the data generated were rigorous and reproducible, each sample was analyzed in sextuplicate in the HALO assay, and the mean and SEM calculated using Microsoft Excel. Statistical analyses were then performed using Prism 6 (GraphPad, Inc. Software), as detailed in “Materials and Methods” section, taking a P value of <0.05 to be statistically significant.
A dose of 20 cGy of 1 GeV/n 56Fe ions was chosen as it is the approximate total dose delivered by a mean of one iron ion traversal/MSC cell nucleus (22.73 cGy for a 100 μm2 nuclear surface area). It is also the approximate annual dose contribution from GCR ions in deep space [69] and threshold dose identified for 56Fe ion-induced in vitro neoplastic transformation [70]—although it should be noted that doses as low as 5 cGy of 1 GeV/n iron ions delivered at NSRL were shown to significantly increase primary fibroblast anchorage-independent growth frequencies [71]. A proton dose of 100 cGy was chosen as a potential dose to the hematopoietic blood-forming organs of astronauts during an intense solar particle event such as the August 1972 or October 1989 events [72,73]. We likewise utilized the same doses, ion sequence, and 15-min interval for our proton/iron ion dual beam (mixed-field) exposures previously shown to maximize cellular transformation frequencies [71,74,75]. Aspects of the charged particle microdosimetry of the numbers of particle traversals per cell nucleus for each irradiation scheme are also important to consider for these exposures. Assuming an estimated surface area of 100 μm2 for MSC nuclei, the numbers of protons and iron ions per nucleus can be calculated from the respective ion beam fluences (∼5.3 × 106 and 4.4 × 104 particles/cm2 · cGy): for the 50 MeV proton irradiations, the MSC nuclei ∼5.3 protons/cGy with the 100 cGy dose equating to a mean of 530 protons/nucleus. For the 1 GeV/n 56Fe ion irradiations, the MSC nuclei receive only 0.044 ions/cGy with the 20 cGy dose equating to a mean of only 0.88 56Fe ions/nucleus, with the number of particle traversals/nucleus following a Poisson distribution (ie, ∼41% of MSC nuclei do not receive iron ion traversals). While some MSC nuclei may not have been directly hit by an iron ion track directly, all cells within the culture are uniformly irradiated with low LET delta-ray electrons generated in the iron ion's penumbra [76]. With the LET of 137Cs γ-ray-induced photoelectrons being very similar to 50 MeV protons (0.91 vs. 1.22 keV/μm in H2O, respectively), MSC nuclei experience similar numbers of photoelectron traversals during gamma irradiation [77].
As can be seen in Fig. 1, gamma and proton irradiation produced similar effects on the most primitive HSC, both directly (irradiation of HSC reduced HPP-CFC output by ∼50%) and indirectly (irradiation of MSC reduced HPP-CFC output by 30%–40%). Importantly, the irradiation of both HSC and MSC produced an additive negative effect on the most primitive HSC, reducing their ability to give rise to HPP-CFC by ∼75%. The results with 56Fe ion irradiations were far more unexpected, as exposures of HSC or MSC cultures alone resulted in a reduction in HPP-CFC output of 50% and 30%, respectively, yet coculture of HSC and MSC that had both been irradiated with 20 cGy of 56Fe ions somehow restored their HPP-CFC potential to normal values. Perhaps the most remarkable results came from the sequential irradiations with proton and 56Fe ions, however. While treating the HSC with protons followed 15 min later by 56Fe ions reduced the HPP-CFC output by 70%, treating the MSC with this sequential dosing regimen actually increased the HPP-CFC output of the coculture by roughly 20% (although this difference did not reach statistical significance). Unexpectedly, given the seemingly beneficial effect of exposing MSC to sequential proton and 56Fe ion irradiation, coculturing HSC and MSC that had both been sequentially exposed to these ions resulted in an 80% reduction in HPP-CFC output.
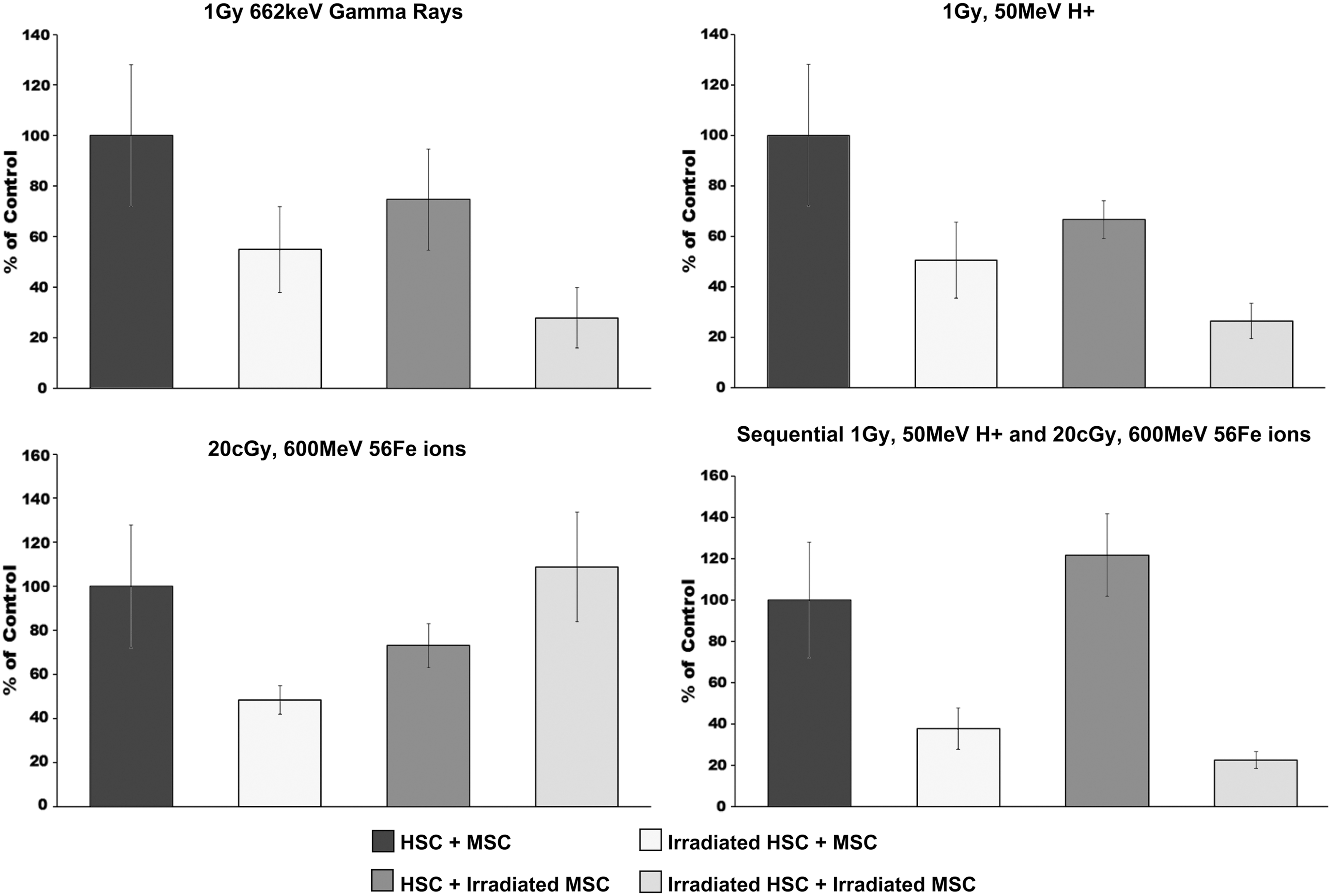
HALO assay was used to quantify the HPP-CFC output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these similar radiation species. Experimental groups were as follows: HSC ± MSC: unirradiated HSC plated over unirradiated MSC. Irradiated HSC ± MSC: HSC exposed to each indicated radiation species plated over unirradiated MSC. HSC ± irradiated MSC: unirradiated HSC plated over MSC exposed to each indicated radiation species. Irradiated HSC ± irradiated MSC: HSC exposed to each indicated radiation species plated over MSC exposed to the same radiation species. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. HPP-CFC, high proliferative potential colony-forming cell; HSC, hematopoietic stem cells; MSC, mesenchymal stem cells; SEM, standard error of the mean; HALO, Hematopoietic/Hemotoxicity Assay by Luminescence Output.
As can be seen in Fig. 2, very similar results were obtained when we examined the next most primitive colony type assayable with the HALO system, the CFU-GEMM (CFU-Mix). As we observed with the HPP-CFC, γ-rays and protons produced very similar effects, with exposure of the HSC resulting in ∼75% reduction in CFU-GEMM and exposure of the MSC resulting in a 20%–30% reduction, but combined exposures of both HSC and MSC reducing CFU-GEMM output by roughly 85%. Exposure of HSC or MSC alone to 56Fe ions reduced the CFU-GEMM output by 30%, yet, just as was seen with the HPP-CFC, coculture of HSC and MSC that had each been exposed to 56Fe ion irradiation restored the CFU-GEMM potential. Exposure of the HSC and MSC to sequential protons and 56Fe ions yielded similar results to those seen with protons alone, namely an additive effect of the two ion species, such that sequential treatment of both MSC and HSC with protons followed by 56Fe ions resulted in an 80%–90% reduction in CFU-GEMM output of the cocultures.

HALO assay was used to quantify the CFU-GEMM output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these same radiation species. Experimental groups were as described in detail in Fig. 1. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. CFU-GEMM, colony-forming unit granulocyte, erythrocyte, monocyte, megakaryocyte.
Figure 3 shows the impact of γ-ray, proton, 56Fe ion, or sequential proton + 56Fe ion irradiation on the more mature myeloid (CFU-GM) output of the HSC + MSC cocultures. As was seen with the more primitive colony types, gamma and proton irradiation of the HSC themselves produced fairly similar results, reducing CFU-GM colony output by ∼60%. Irradiating MSC with γ-rays or protons, however, produced very different results. Gamma irradiation of the MSC had no impact on the CFU-GM output of the cocultures, and coculturing γ-irradiated HSC with γ-irradiated MSC produced a similar number of CFU-GM as culturing γ-irradiated HSC over unirradiated MSC. In contrast, proton irradiation of the MSC alone reduced the CFU-GM output of the cocultures by ∼30%, and coculturing proton-irradiated HSC over proton-irradiated MSC produced an additive (negative) impact on CFU-GM output, reducing CFU-GM numbers by ∼80%.

HALO assay was used to quantify the CFU-GM output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these same radiation species. Experimental groups were as in Fig. 1. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. CFU-GM, colony-forming unit granulocyte, monocyte.
Looking at the platelet (megakaryocyte) potential of the cocultures following exposure to each radiation type, we found (Fig. 4) that gamma or proton irradiation of the HSC resulted in a 60% decrease in CFC-Mk colonies. Gamma irradiation of the MSC alone had no real effect upon CFC-Mk output of the cocultures, while proton irradiation of the MSC decreased the CFC-Mk output by 30%. Curiously, irradiating both HSC and MSC with γ-rays resulted in a further decrease in CFC-Mk over what was seen after irradiating the HSC alone (total reduction in CFC-Mk of 70%). In contrast, irradiating just the MSC with protons reduced CFC-Mk output by about 40%; however, combined irradiation of both HSC and MSC produced no further decrease in CFC-Mk over what was seen with irradiation of the HSC alone. Iron ion irradiation had no significant effect upon CFC-Mk output, irrespective of whether the HSC, the MSC, or both were irradiated. Exposing HSC to sequential proton + 56Fe ions reduced CFC-MK output by about the same amount as was seen with protons alone (60% reduction), while sequential irradiation of the MSC increased CFC-Mk output to 130% of what was seen with no irradiation. Surprisingly, however, exposing both HSC and MSC to protons followed by 56Fe ions resulted in a nearly 90% reduction in CFC-Mk potential, again stressing the potentially deleterious effects of exposing both HSC and MSC to combined SEP/GCR, which could not be predicted based on studies with a single ion or a single cell type.
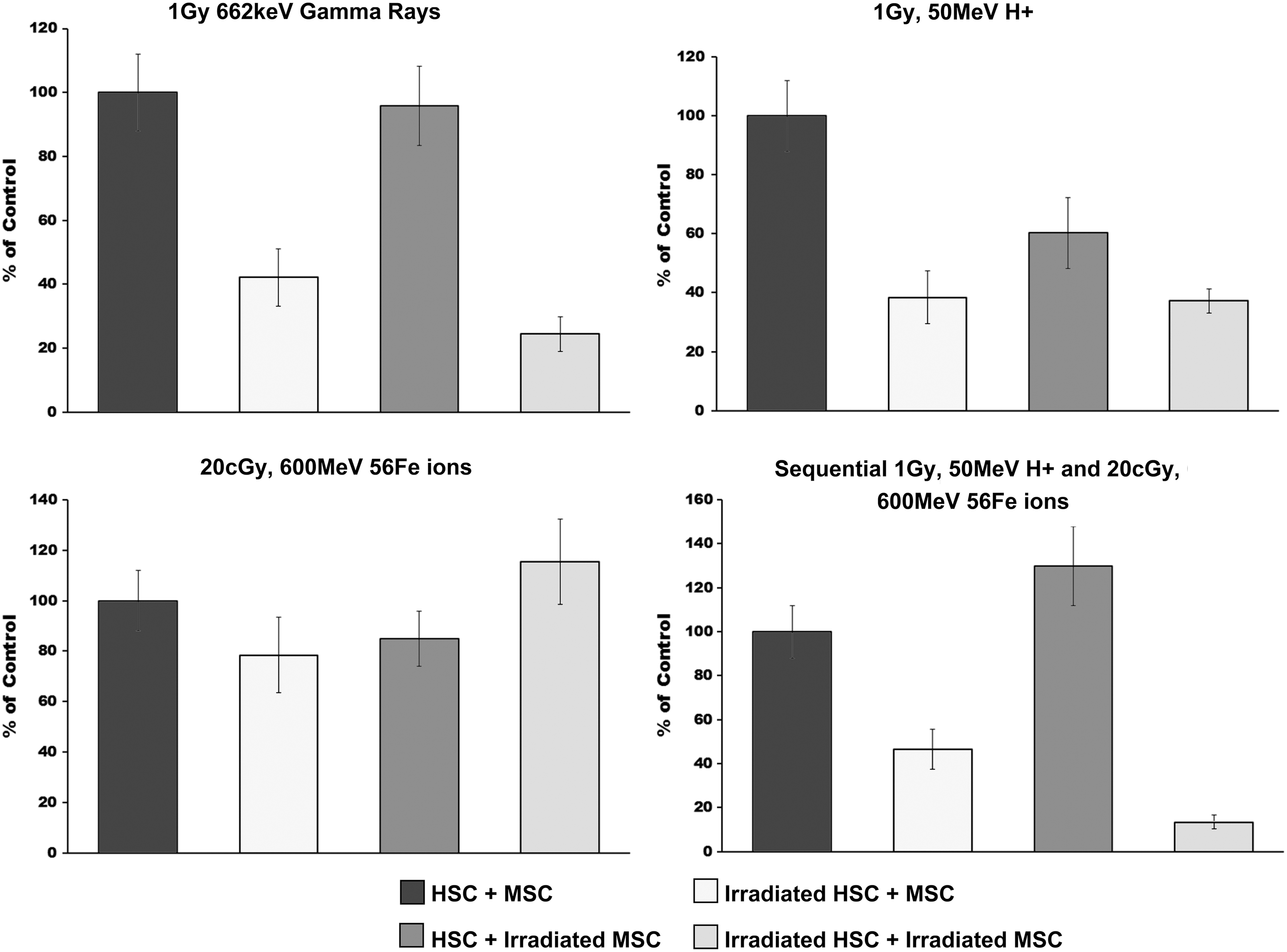
HALO assay was used to quantify the CFC-Mk output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these same radiation species. Experimental groups were as in Fig. 1. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. CFC-Mk, megakaryocyte colony-forming cell.
With respect to the lymphoid lineages (Figs. 5 and 6), gamma irradiation of the HSC reduced CFC-T (Fig. 5) and CFC-B (Fig. 6) output by 60%; irradiation of the MSC had no real impact on CFC-T output, but negatively impacted B lineage potential, dropping CFC-B output by ∼35%. Coculturing HSC and MSC that had both been exposed to γ-rays, however, reduced both CFC-T and CFC-B output by about 70%. Proton irradiation of either HSC or MSC alone exerted a pronounced effect on CFC-T and CFC-B output, but the effect on CFC-B was the most pronounced, with proton irradiation of the HSC dropping CFC-B output by nearly 80%, and irradiation of the MSC dropping CFC-B potential by roughly 60%. Coculturing proton-irradiated HSC and MSC, however, had no further effect upon CFC-B over what was seen with irradiating the HSC alone, but further reduced the output of CFC-T over what was seen with irradiated HSC alone (70% reduction with irradiated HSC + irradiated MSC vs. 60% reduction with irradiated HSC alone, respectively). 56Fe ion irradiation of either the HSC or MSC alone negatively impacted both CFC-T and CFC-B output, but the effects on CFC-B were the most pronounced [55% (HSC irradiation) and 40% (MSC irradiation)]. Counter intuitively, irradiation of both HSC and MSC, followed by coculture of these two irradiated cell types, restored both CFC-T and CFC-B output to control (sham irradiated) levels. In similarity to what was seen with 56Fe ions alone, sequential treatment of HSC or MSC alone with protons followed by 56Fe ions had the most pronounced effect on the B cell lineage, dropping CFC-B output by 50% (HSC alone) and 40% (MSC alone). As with the other lineages, however, if both HSC and MSC were treated with the sequential irradiation scheme, CFC-T and CFC-B output were both reduced by 80%, again highlighting the marked differences in the effect seen when both cell types are irradiated and sequential SEP + GCR radiations are used, compared to the effects when only a single cell type is irradiated with SEP or GCR radiation alone. The ability of SEP, GCR, and particularly SEP + GCR radiation to negatively impact upon the ability of HSC to differentiate along the lymphoid (T and B cell) lineages raises the troubling possibility that astronauts may be doubly at risk both due to direct genetic damage/carcinogenesis as a result of HSC exposure to space radiation and as a result of space radiation-induced deficiencies in immune cell formation/function, which could preclude recognition and removal of any potential malignant hematopoietic clones.

HALO assay was used to quantify the CFC-T output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these similar radiation species. Experimental groups were as in Fig. 1. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. CFC-T, T-lymphoid colony-forming cell.
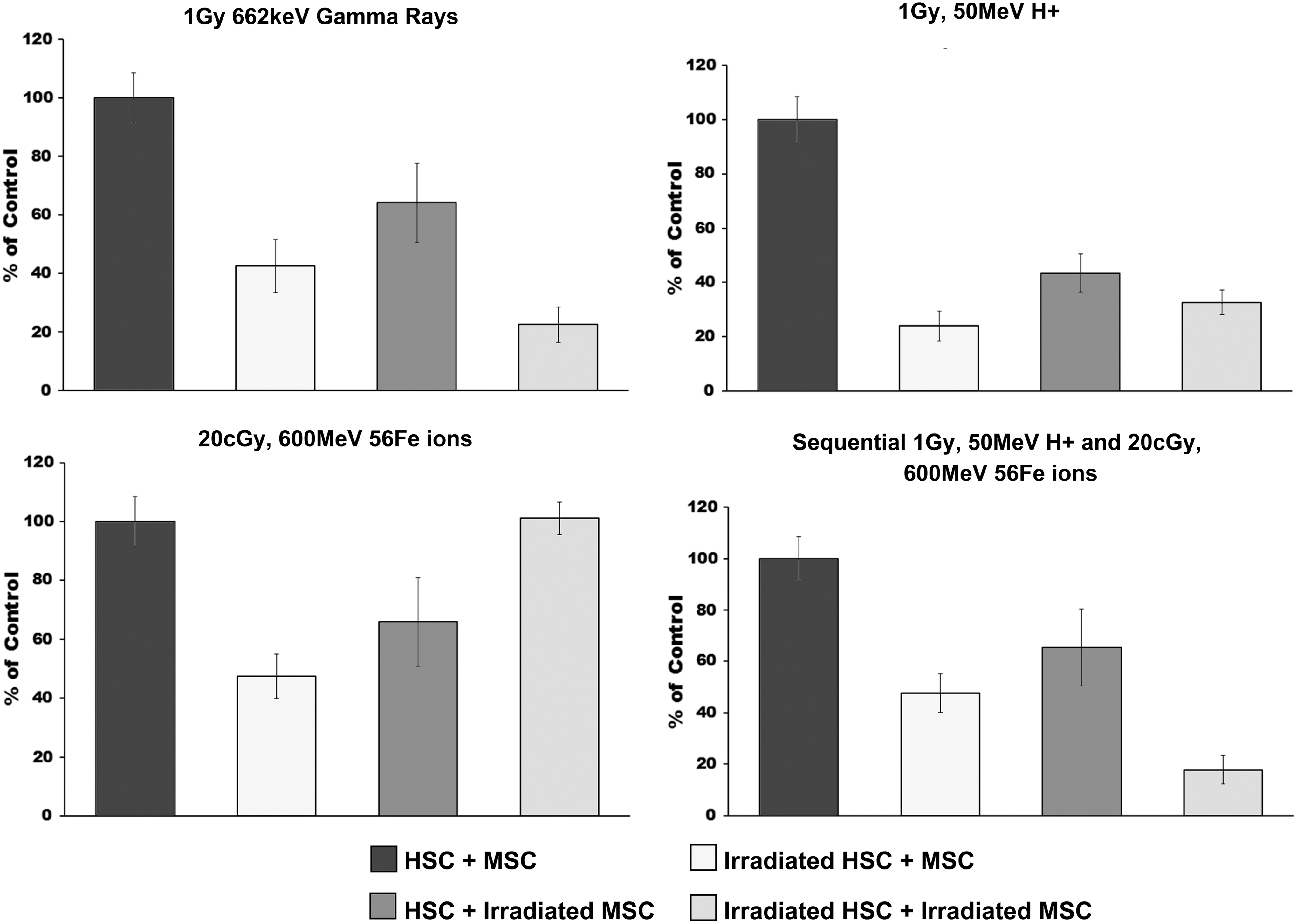
HALO assay was used to quantify the CFC-B output of human HSC following exposure to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation and subsequent culture over unirradiated human MSC, or MSC exposed to each of these same radiation species. Experimental groups were as in Fig. 1. All samples were run in replicates of 6–8 in the HALO assays, and experiments were repeated with HSC and MSC from a minimum of four different human donors. All data are presented as the mean ± SEM, and were analyzed with Prism 6 (GraphPad Software, Inc.), as detailed in “Materials and Methods” section. For all analyses, P ≤ 0.05 was considered to be statistically significant. CFC-B, B-lymphoid colony-forming cell.
Defining the effects of SEP and GCR radiation on the human BM microenvironment/niche
To begin delineating the mechanism whereby the exposure of MSC to each form of radiation might alter the ability of the MSC to support normal/functional hematopoiesis, we isolated RNA from MSC before and 48 h following exposure to 137Cs γ-rays, 50 MeV protons, 1 GeV/n 56Fe ions, or sequential proton + iron ion radiation. We also collected RNA after the MSC had been passaged five times in culture, to enable us to determine whether long-term/stable alterations in gene expression took place as a result of exposure to these various radiation species. All samples were quantitated, subjected to Bioanalyzer (Agilent) quality control checks, and were then analyzed by Illumina microarrays to compile a comprehensive picture of the significant changes each type of radiation, delivered either singly or in combination, produced at the transcriptional level within these critical BM niche cells.
Analyses by heat map (Fig. 7) and PCA (Fig. 8) revealed very tight clustering of transcriptomic alterations associated with each radiation type, showing that significant differences exist between not only irradiated cells and their respective preirradiation control but also between the same donor-derived cells exposed to γ-rays, protons, or iron ions. Setting our analysis to only consider genes whose differential expression is significant at P ≤ 0.05, we observed differential expression in as many as 287 genes, depending which radiation conditions were being compared.

Heat map of differentially expressed genes in MSC following irradiation. MSC from multiple human donors were exposed to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation at NSRL. Upon return to WFIRM (24 h postirradiation), RNA was isolated and gene expression examined by Illumina microarray. Heat map analysis of the resultant gene expression data showed tight hierarchical clustering based on the type of radiation to which the MSC were exposed. Only genes whose differential expression was significant at P ≤ 0.05 were considered in the presented analysis. Unfortunately, the RNA we obtained from the third biological replicate of MSC exposed to protons was of insufficient quality to proceed with microarray analysis. As such, n = 2 for the data for this irradiation scheme. NSRL, NASA Space Radiation Laboratory; WFIRM, Wake Forest Institute for Regenerative Medicine.
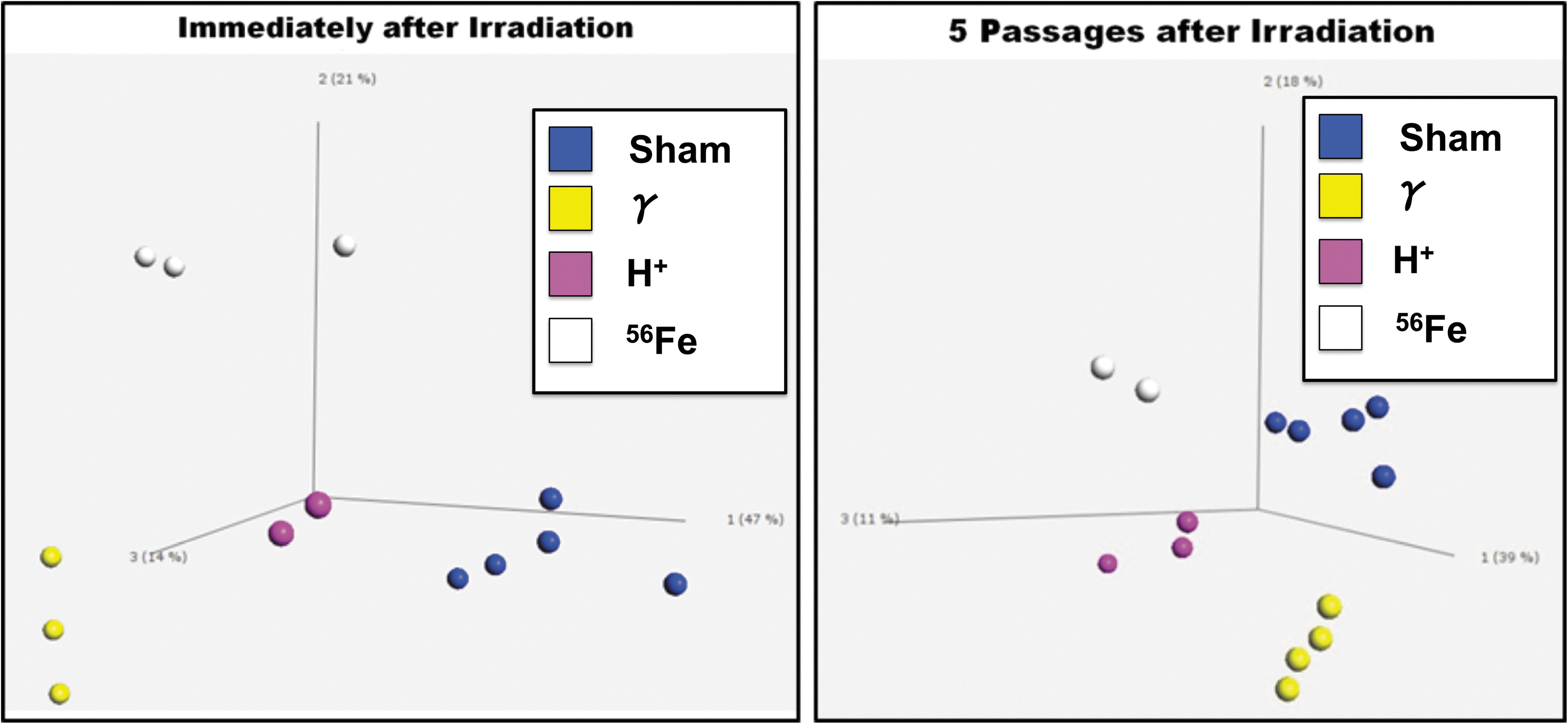
PCA analysis on MSC following irradiation. MSC from multiple human donors were exposed to: (i) sham, (ii) gamma, (iii) proton, (iv) 56Fe ion, or (v) sequential proton + 56Fe ion irradiation at NSRL. Upon return to WFIRM (24 h postirradiation), and after five passages in culture, RNA was isolated from an aliquot of MSC from each experimental group and gene expression examined by Illumina microarray. PCA of the resultant gene expression data showed tight clustering based on the type of radiation to which the MSC were exposed, and this clustering persisted after five culture passages, suggesting the differential changes in gene expression induced by exposure to each radiation type were fairly long lasting. Only genes whose differential expression was significant at P ≤ 0.05 were considered in the presented analysis. PCA, principal component analysis.
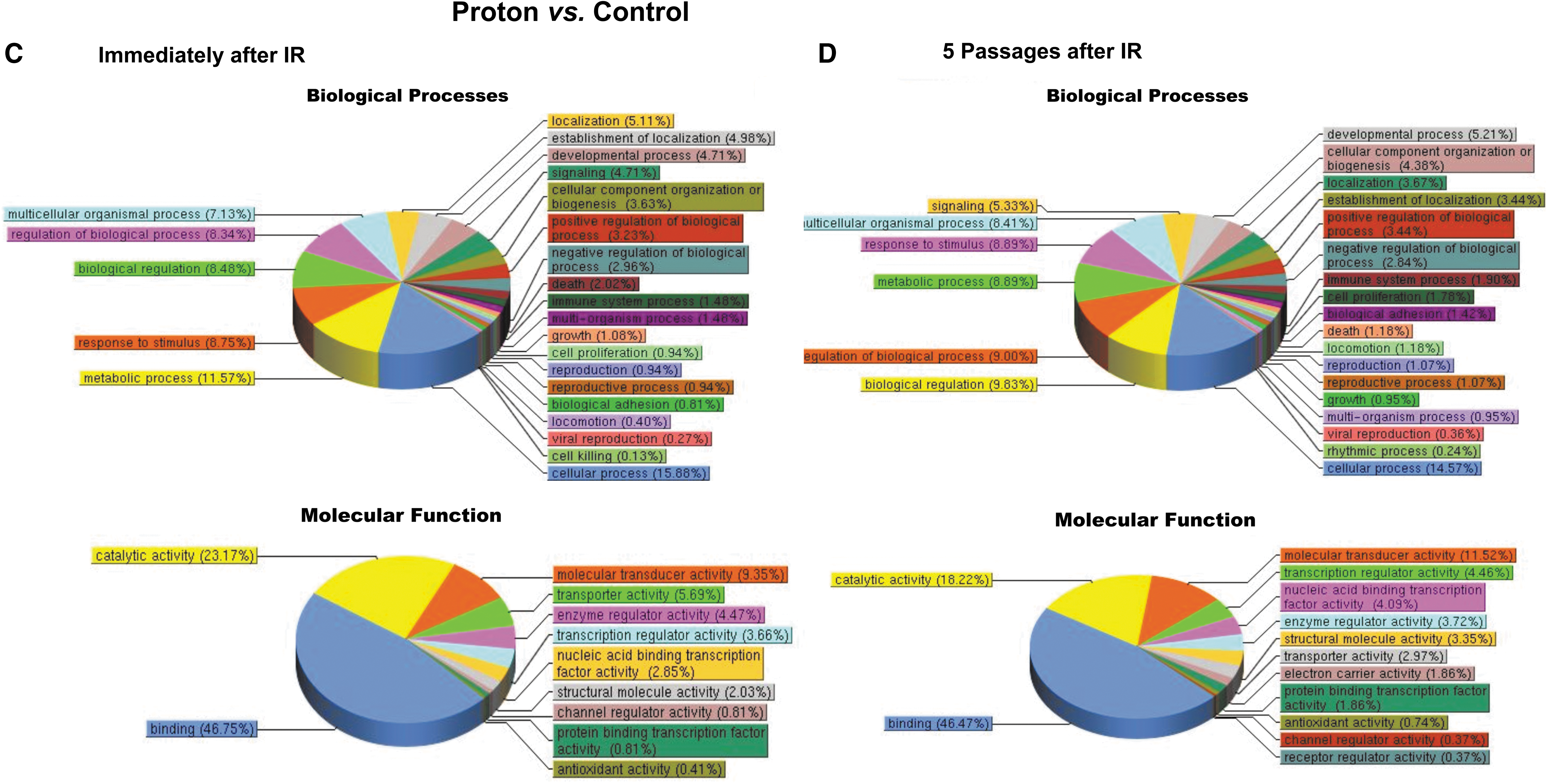
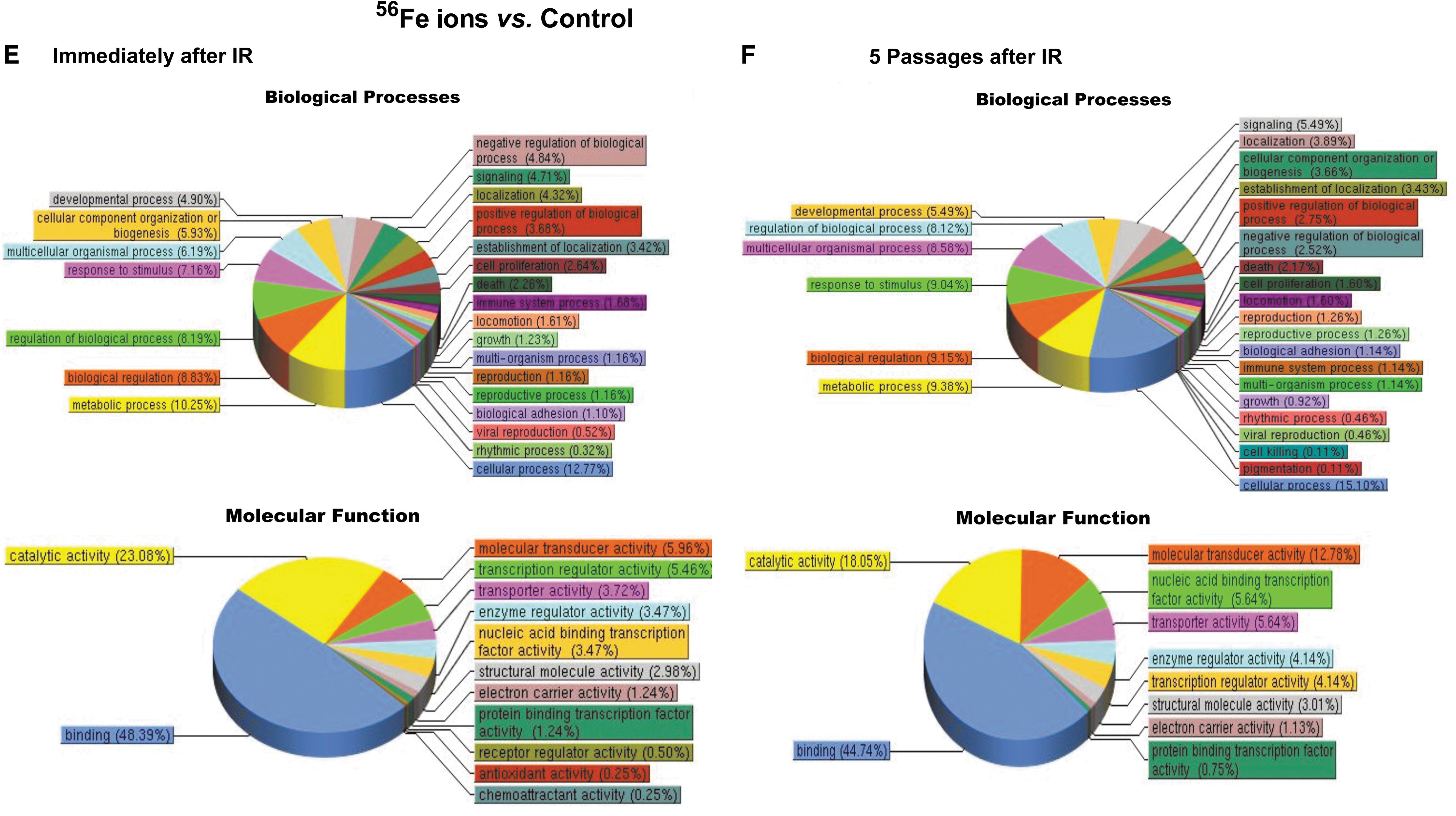
Pathway analysis results (Fig. 9) showed that at 24 h postirradiation, MSC gene expression profiles were markedly altered by exposure to each radiation type, with the proton and 56Fe ion irradiations significantly affecting a greater number of cellular pathways than γ-irradiation. Specifically, exposure to either protons or 56Fe ions led to marked alterations in multiple classes of genes that were largely unaffected by gamma radiation, such as genes associated with developmental processes, positive and negative regulation of biological processes, response to stimulus, growth, viral reproduction, cell killing, molecular transducer activity, nucleic acid-binding transcription factor activity, protein-binding transcription factor activity, channel regulator activity, transporter activity, rhythmic processes, biological regulation, cell proliferation, and antioxidant activity. Moreover, it is particularly interesting to note that, while gene expression changes following exposure of human MSC to 100 cGy of 137Cs γ-irradiation largely return to normal preirradiation levels following five passages in culture (Fig. 9A, B), the changes induced by exposure to either 100 cGy of 50 MeV protons (SEP; Fig. 9C, D) or 20 cGy of 1 GeV/n 56Fe ions (GCR; Fig. 9E, F) persisted in the MSC even after five culture passages. These results thus suggest that (as-yet unidentified) epigenetic changes induced by SEP/GCR radiation are likely perpetuated following cell division, and are passed on to the clonal daughter cells. As such, exposure to space radiation may have the potential to result in long-term alterations to the BM microenvironment and its ability to support normal hematopoiesis.
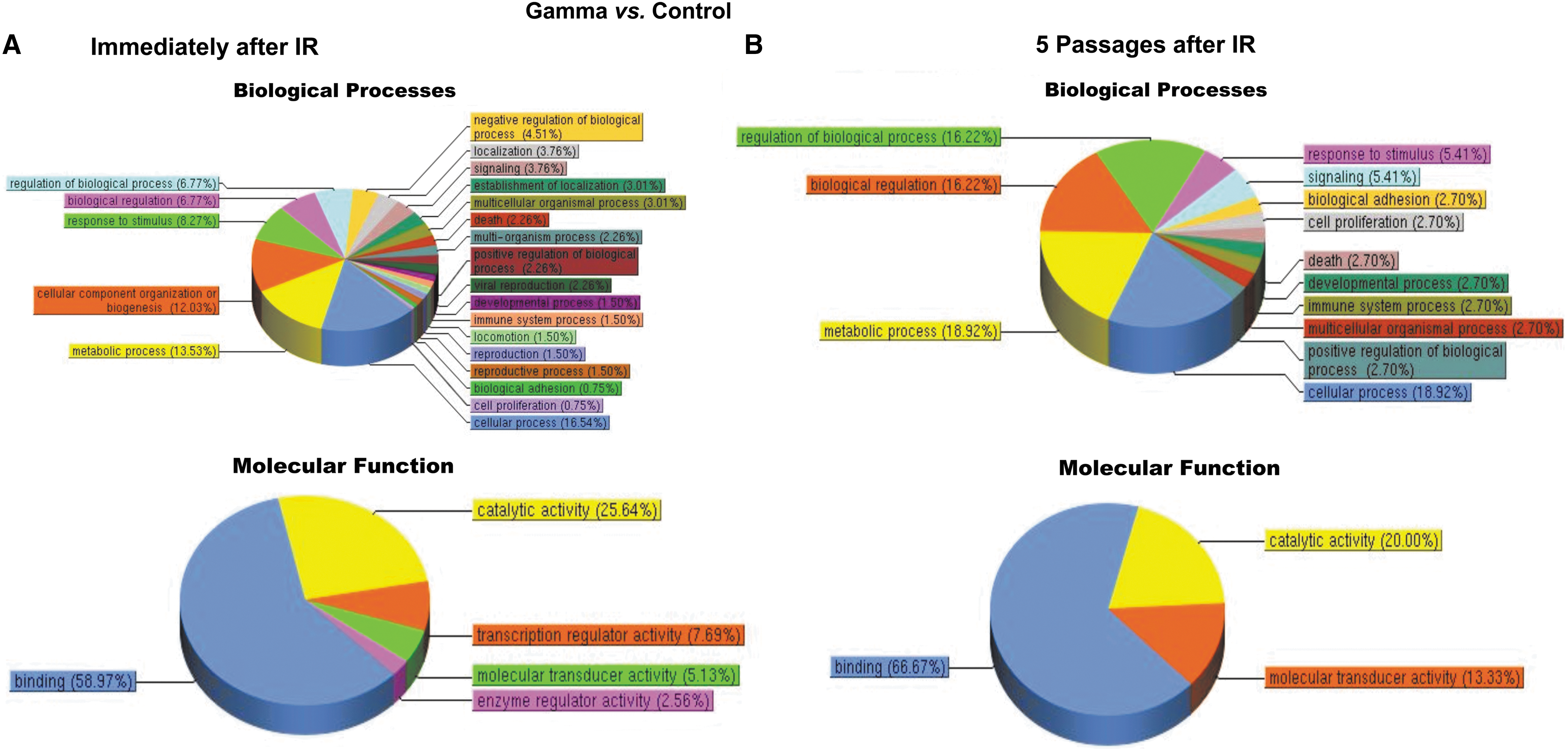
Pathway analysis of the differentially expressed genes in human MSC at 24 h postirradiation and after five passages in culture. (
Since changes in RNA expression do not necessarily translate to altered levels of their respective protein products [78 –81], we also collected culture supernatants from MSC cultures from each donor at 48 h following exposure to each type of radiation, using the culture supernatant from the MSC that were flown to BNL and not exposed to radiation as donor-specific controls. These supernatants were analyzed using a luminescence-based RayBio Human Cytokine Antibody Array, which allows simultaneous quantitation of the expression levels of 42 cytokines/chemokines involved in hematopoiesis/inflammation/immune function. The map of this array and the luminescence image of this assay are shown in Fig. 10A and B, respectively. We used ImageJ software to quantify the intensity of each individual spot on each array, and used Microsoft Excel to normalize the data with respect to the basal levels of expression of each individual cytokine seen for each donor in the control sample that was flown to BNL but not irradiated. These values were then compared to those obtained following exposure to γ-rays, protons, or 56Fe ions to determine the impact each form of radiation had on MSC production of cytokines/growth factors known to be involved in hematopoiesis support. As can be seen in Fig. 11, exposure to gamma, proton, or iron ion irradiation markedly altered the expression of the majority of cytokines present on this focused array (n = 3 different human donors), providing multiple mechanistic insights for the pronounced effects we observed on hematopoietic cell function and potential in the HALO assays presented in Figs. 1 –6.

Supernatants were collected from human MSC at 24 h following exposure to each radiation species and were then analyzed by a multiplexed, membrane-based array to probe expression of 42 cytokines/chemokines involved in hematopoiesis/inflammation/immune function. A map of this array appears in
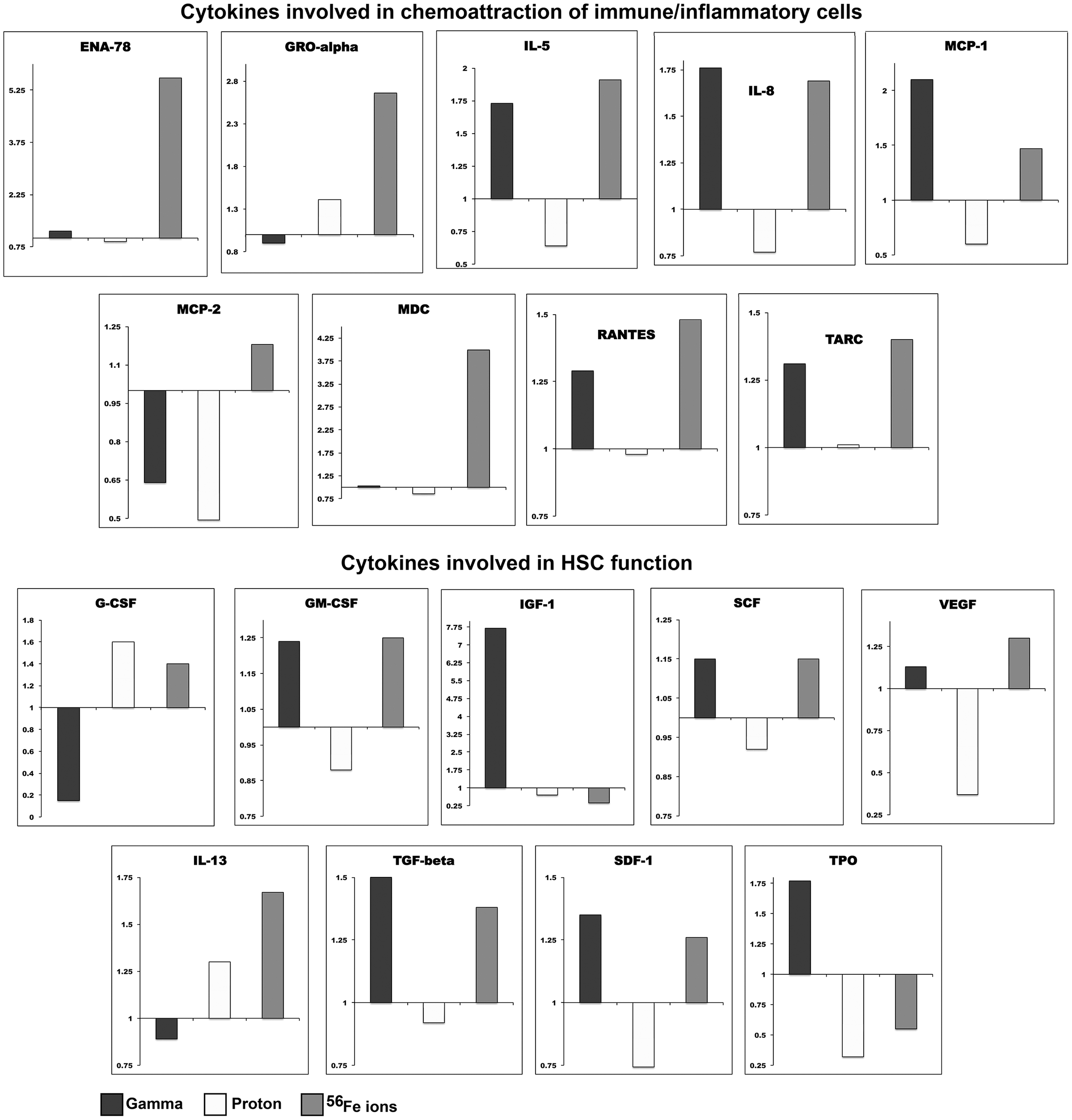
ImageJ software was used to quantify the intensity of each individual spot on each array, and Microsoft Excel was used to normalize the data with respect to the basal levels of expression of each individual cytokine seen for each donor in the control sample that was flown to Brookhaven National Laboratory, but not irradiated. These values were then compared to those obtained following exposure to γ-rays, protons, or 56Fe ions to determine the impact each form of radiation had on MSC production of cytokines/growth factors known to be involved in hematopoiesis support. Data presented are the mean ± standard error of the mean of the results obtained with MSC from three different human donors.
It is particularly interesting to note that the effects of cesium-137 gamma radiation on MSC are often dramatically different from those observed following exposure of these cells to either the same 100 cGy dose of 50 MeV protons (which are similarly low LET) or 20 cGy of 1 GeV/n 56Fe ions, highlighting the necessity of performing studies with human cells and radiation species/doses that accurately mimic conditions to which astronauts would be exposed during deep space missions, to accurately assess the risks of such exposures. Equally intriguing are the pronounced differences in the effects of low LET protons compared to high LET 56Fe ions, with these two forms of radiation often exerting opposing effects on the same cytokines. Iron ion irradiation tended to upregulate many cytokines involved in the support/maintenance of hematopoiesis and immune cell development (in agreement with our HALO data), while proton irradiation downregulated many of these same cytokines. The downregulation of Interleukins 2, 3, 5, 6, 7, and 8 by proton irradiation is particularly noteworthy, as these cytokines are essential for the induction and differentiation of mature immune effector cells from HSC, and for proper functionality of the resultant immune cells. The ability of proton irradiation to negatively regulate the expression of these cytokines lends further support to the notion that astronauts may be doubly at risk of cancer, both due to direct genetic damage/carcinogenesis as a result of HSC exposure to space radiation and as a result of space radiation-induced deficiencies in immune cell formation/function, precluding recognition and removal of any malignant hematopoietic clones that do form.
Discussion
The results of this study build upon our prior work showing that the most primitive HSC/HPC can not only be negatively affected by direct exposure to modeled SEP and GCR radiations [47] but also indirectly by “biological bystander effects” that occur when unirradiated HSC are placed in close proximity to a marrow microenvironment that has been exposed to SEP/GCR radiation. The bystander effect that we refer to in this study should not be confused with the more traditional “radiation-induced bystander effect” phenomenon in radiobiology, in which unirradiated cells neighboring directly irradiated ones exhibit similar stress/DNA damage responses, but this remains an important concept in our low dose studies given that the MSC received ∼0.88 56Fe ion particle traversals/cell for the 20 cGy dose (although all MSC nuclei were directly irradiated from delta-ray electrons induced by the primary Fe ion traversals) [82 –84]. Perhaps most importantly, our results show that these direct and indirect effects of modeled SEP/GCR irradiation are often additive, combining to produce a pronounced decrease in the output of hematopoietic colonies of various lineages. Moreover, the exposure of both HSC and MSC to sequential proton and 56Fe ion irradiation (as would occur in deep space, given the differing fluences of these radiation species) produced much more deleterious effects than exposure to either ion alone, combining in effectiveness to reduce the output of the most primitive HPP-CFC colonies by ∼80%, as well as other colony types, to varying degrees.
In this study, we employed Stro-1+ MSC as a paradigmatic cell of the HSC niche. Control of hematopoiesis within this supportive/regulatory microenvironment of the BM involves interactions between the HSC and MSC, extracellular factors, and other BM stromal cells, which collectively direct commitment to one of the various hematopoietic lineages, and ultimately lead to the full maturation of the respective precursor [37 –41,85 –87]. This specialized niche has been the focus of intense investigation in recent years, and has been defined in terms of cells, extracellular matrix, soluble factors, and signaling pathways [37 –42,85 –87]. While the stromal microenvironment of the marrow comprised multiple cell populations, with the exception of macrophages, all are derived from MSC [43,88 –93]. MSC thus play a critical role in the control/regulation of hematopoiesis, since they give rise to all the cells that comprise the hematopoietic microenvironment/niche. For this reason, we feel they represent a highly biologically relevant model system for studying the effects of SEP/GCR radiation on the marrow niche, and the impact upon hematopoiesis. It is also important to note that MSC have been shown to play a critical role in suppressing the growth/expansion of hematopoietic clones that have undergone leukemic transformation, functioning as sentinels against hematologic malignancies [94 –96]. As such, any environmental insult that damages MSC would be expected to not only produce pronounced effects on the types and functionality of hematopoietic/immune cells generated, as we show in these studies, but perhaps also impact upon the risk of leukemogenesis as a result of exposure to SEP/GCR radiation.
Also, of particular note are our findings on the transcriptional alterations that modeled SEP and GCR irradiation induce in human MSC. It is perhaps not surprising that exposure to these unique radiation species produces marked alterations in gene expression and in the secretion of hematopoiesis-related cytokines/chemokines. It is, however, intriguing and unexpected that the transcriptional changes induced by protons and 56Fe ions persist with culture expansion, and are still present five passages after exposure. This is in contrast to the results obtained with γ-rays, and suggests that species of radiation found in deep space may have the ability to exert far more lasting effects on human cell function than would be predicted with studies using standard terrestrial radiation sources.
These findings are critical for NASA to consider as they update and revise their radiation risk assessments with respect to hematological malignancies during prolonged deep space missions. Compounding the risks of hematological malignancies are numerous studies collectively demonstrating that extended spaceflight conditions, in particular, microgravity, deleteriously affect the immune system at multiple levels [13 –29] and impair astronauts' ability to respond to infection or immune challenge [30 –36]. As such, astronauts could potentially be doubly at risk, due not only to the possibility of radiation-induced carcinogenesis but also a reduction in the ability of their immune system to recognize and eliminate cells that have undergone transformation. This finding that the output of CFC-T and CFC-B is reduced when the microenvironment is exposed to SEP and GCR radiation would be expected to further add to this potential risk.
Footnotes
Acknowledgments
This work was supported by grant no. NNX13AB67G from the National Aeronautics and Space Administration (NASA) Human Research Program and grant no. NNX17AE49G from the NASA Space Biology Program.
Author Disclosure Statement
No competing financial interests exist.