
Abstract
Converging evidence suggest that levels of alpha-synuclein (aSyn) expression play a critical role in Parkinson's disease (PD). Several mutations of the SNCA gene, encoding for aSyn have been associated to either the familial or the sporadic forms of PD. Nonetheless, the mechanism underlying wild-type aSyn-mediated neurotoxicity in neuronal cells as well as its specific driving role in PD pathogenesis has yet to be fully clarified. In this view, the development of proper in vitro cellular systems is a crucial step. In this study, we present a novel human Tet-on human neural stem cell (hNSC) line, in which aSyn timing and level of expression can be tightly experimentally tuned. Induction of aSyn in self-renewing hNSCs leads to progressive formation of aSyn aggregates and impairs their proliferation and cell survival. Furthermore, aSyn induction during the neuronal differentiation process results in reduced neuronal differentiation and increased number of astrocytes and undifferentiated cells in culture. Finally, acute aSyn induction in hNSC-derived dopaminergic neuronal cultures results in cell toxicity. This novel conditional in vitro cell model system may be a valuable tool for dissecting of aSyn pathogenic effects in hNSCs and neurons and in developing new potential therapeutic strategies.
Introduction
L
Mutations of the SNCA gene, encoding for aSyn, are either associated to rare familial PD or represent risk factors for sporadic PD [3,4]. Furthermore, increased SCNA copy number variation has been shown to be causal for PD, thus suggesting that increased aSyn protein levels are sufficient to trigger the disease [5]. In PD patients, aggregation of aSyn in LBs has been shown, resulting in its mislocalization and loss of function, ultimately leading to the loss of dopaminergic neurons. Besides its role in mature neurons, in vivo aSyn overexpression has been shown to affect proliferative potential of mouse hippocampal neural stem cells (NSCs), resulting in a reduction of the pool of neural progenitors and decreased neuronal differentiation and maturation [6 –9]. Also, ectopic expression of aSyn was shown to affect the migration of NSCs in mouse subventricular zone [10].
Despite the association to PD has been known for decades, aSyn function in human neural stem cells (hNSCs) and neurons as well as its involvement in PD are still poorly understood. So far, the main limitations have been due to the inaccessibility of adequate in vitro cellular models. Indeed, most of the studies aimed at defining a clear role for aSyn in normal and PD neurons have been so far carried out using either non-human systems, including rodent culture [6,7,10], non-neuronal transformed cell lines [11], or noncentral nervous system (CNS)-derived neuronal-like cells [12,13].
More recently, the advent of human-induced pluripotent stem cells (hiPSCs)’ technology has widened opportunities to model human conditions, including the possibility to generate bona fide hNSCs and mature neurons and consequently matching the affected cells in neurodegenerative disorders [14]. hiPSC lines derived from PD patients carrying triplication of the SNCA locus have been reported [15]. These cells were proven to efficiently differentiate and mature into TH+ve dopaminergic neurons while showing a twofold increase in aSyn protein levels with respect to hiPSCs derived from unaffected relatives and recapitulating some aspects of the patient physiology. Interestingly, neuronal precursor cells derived from these PD hiPSCs have been reported to exhibit significant deficiencies in growth, viability, cellular energy metabolism, and stress resistance [16], thus indicating that aSyn may play important roles in hNSC physiology.
Similarly, wild-type (wt) aSyn overexpression in fetal cortex-derived hNSCs has been shown to impair cell growth and neuronal versus glial lineage commitment [17]. Finally, aSyn overexpression has been described to reduce the number of mouse secondary neurospheres formed and to affect NSC morphology and cell cycle progression, leading to their accelerated differentiation [10]. These studies suggest that there is a link between neurogenesis, aSyn, and neurodegenerative diseases [18].
In this study, we report the generation of a novel in vitro cell system in which levels and timing of human wt aSyn expression can be experimentally tightly controlled in hiPSC-derived long-term expandable NSCs. Following induction, a progressive increase of aSyn levels can be achieved, leading to the formation of aSyn cytoplasmic aggregates. This versatile system allows to investigate the effects of aSyn expression on hNSCs behavior, including self-renewal and differentiation programs.
Our results show that induction of aSyn leads to a reduction in hNSCs growth accompanied by an increased susceptibility to apoptosis. During neuronal differentiation process, aSyn induction affects neurogenic potential and induces an increase in the number of astrocytes and undifferentiated cells in culture. Furthermore, acute aSyn induction resulted in enhanced apoptotic cell death in hNSC-derived dopaminergic neurons.
This novel in vitro cell system may represent a valuable tool for studying aSyn-driven pathogenic-relevant mechanisms in hNSCs and their mature derivatives, and for screening potential therapeutics.
Materials and Methods
Cell cultures
AF22 cells (kindly donated by Prof. Austin Smith, Cambridge Stem Cell Institute, University of Cambridge, UK) were previously described [19]. Cells were routinely passaged every 2–3 days at a density of 2.5–3.5 × 104 cells/cm2 in hNSCs Self-renewal medium composed of Dulbecco's modified Eagle's medium/Nutrient Mixture F-12 (DMEM/F-12; Thermo Fisher Scientific), N2 supplement (1%; Thermo Fisher Scientific), B27 supplement (1%; Thermo Fisher Scientific), EGF (10 ng/mL; PeproTech), bFGF (10 ng/mL; PeproTech), and GlutaMAX (2 mM; Thermo Fisher Scientific). Briefly, before seeding the cells, plastic culture vessels were treated with 3 μg/mL Laminin (Thermo Fisher Scientific) at 37°C for 3–5 h. For passaging, cells were incubated for 1–2 min with StemPro Accutase (Thermo Fisher Scientific) and centrifuged at 260 g for 3 min. Pellet was resuspended in fresh medium and plated onto the laminin-coated vessels.
For general neuronal differentiation, cells were seeded on laminin-coated cell culture-grade plasticware at a density of 8 × 103 cells/cm2 in self-renewal medium. The following day, medium was shifted to self-renewal medium deprived of EGF and bFGF. After 3 additional days, medium was replaced with neuronal differentiation medium composed of a 1:1 mix of Neurobasal (Thermo Fisher Scientific), and DMEM/F-12, N2 supplement 1%, B27 supplement 1%, cAMP 300 ng/mL, and GlutaMAX. For aSyn induction during differentiation process, a medium supplemented with doxycycline was replaced every 24 h.
For dopaminergic neuronal differentiation, self-renewing cultures were maintained in the presence of 200 ng/mL of both FGF8 and SHH for 2–3 weeks to specify a ventral midbrain dopaminergic fate, before differentiating them as in the general neuronal differentiation. Seven hundred and fifty nanograms per milliliter of doxycycline were added at 21 days in vitro (DIV) every 24 h for 4 DIV before fixing the cells for immunocytochemistry.
Lentiviral particle preparation and AF22 cell infection
Lentiviral particles carrying the pLVX-TetOne-Puro-human aSyn vector (kindly donated by Dr. Tilo Kunath, University of Edinburgh, UK) were prepared. This vector, based on the pLVX-TetOn-Puro plasmid (Clontech) is specifically designed to carry on a single vector the complementary DNA (cDNA) for a tetracycline transactivator (Tet-on 3G) and a tetracycline responsive promoter (TREGS promoter) containing seven tetracycline-responsive elements (TRE) controlling the expression of the cDNA for human aSyn. The vector also carries a puromycin resistance cassette for selection of infected cells.
For lentiviral particle preparation, 6 × 104 HEK 293T cells/cm2 were seeded in DMEM containing 10% fetal bovine serum (FBS) and left undisturbed overnight. The following day, cells were transfected by CaPO4 method with 20 μg of pLVX-TetOne-Puro-human aSyn, 15 μg of psPAX2 vector (kindly provided by Prof. M. Pizzato, University of Trento, Italy), and 5 μg of VSV-G. Two days after transfection, the supernatant was collected and concentrated using the Lenti-X concentrator reagent (Clontech) following the manufacturer's recommendation.
For AF22 cell infection, 10 μL of concentrated lentiviral particles were used to infect 2 × 104 cells/cm2. After 8 h, the medium was completely changed, and 72 h later, positive selection of the transduced cells was started with 0.3 μg/mL puromycin (Thermo Fisher Scientific) until the noninfected control cells died completely. Cultures were subsequently selected with higher doses of puromycin as discussed in the Results section.
Cell growth assay
For growth assay, 2 × 103 cells/cm2 were seeded onto laminin-coated 24-well plates. aSyn induction was achieved by treatment with 750 ng/mL doxycycline added to the cultures and the medium was renewed every day. Cells were fixed at specific time points by using 4% paraformaldehyde for 15 min at room temperature (RT) and then nuclei were stained with Hoechst 33258 (Thermo Fisher Scientific). Cell counting was performed using Operetta High-Content Imaging System (PerkinElmer). Images were collected with a 10× long working distance objective considering technical quadruplicate for each time point and the cell number was determined using the software Harmony 4.1 (PerkinElmer) by the segmentation of the nuclear region.
Immunocytochemistry and evaluation of aSyn aggregates
For immunofluorescence assay, cells were fixed with 4% paraformaldehyde for 15 min at RT, then permeabilized with 0.5% Triton X-100 in phosphate-buffered saline (PBS) for 15 min at RT and blocked with a blocking solution (5% FBS, 0.3% Triton X-100 in PBS) for 2 h at RT. Cultures were then incubated over-night at 4°C with specific primary antibodies (Supplementary Table S1; Supplementary Data are available online at
For evaluation of aSyn aggregation, cells were treated with 750 ng/mL of doxycycline for 12 days. Cultures were fixed with 4% paraformaldehyde for 15 min at RT and stained with anti-aSyn antibody. Nuclei and cytoplasm were counterstained using Hoechst 33258 and CellMask Deep Red (Molecular Probes). Cells were analyzed using the Operetta High-Content Imaging System. Images were collected using a 20× long WD objective considering technical quadruplicate and analyzed with Harmony 4.1 software. Briefly, cells were identified by the segmentation of the nuclear region based on the Hoechst 33258 signal and cytoplasm region of interest (ROI) was defined based on the CellMask signal. aSyn aggregates were detected as fluorescent spots inside the ROI. Number of objects, area, and intensity of the signal were calculated for each well and normalized on the number of aSynlow/Med- and aSynhigh-intensity aSyn immunoreactive cells.
For the quantitative evaluation of the effect of aSyn induction on mitochondrial number, cells were treated with 750 ng/mL of doxycycline for 5 days, before fixing with 4% paraformaldehyde for 15 min at RT and stained with Mitotracker (Molecular Probes) to image mitochondria as recommended by the manufacturer. Nuclei and cytoplasm were counterstained using Hoechst 33258 and CellMask Deep Red (Molecular Probes). Cells were analyzed using the Operetta High-Content Imaging System. Images were collected and analyzed as described above. Briefly, cells were identified by the segmentation of the nuclear region based on the Hoechst 33258 signal and cytoplasm. ROI was defined based on the CellMask signal. Mitochondria were detected as fluorescent spots inside the ROI. Area and number of objects were calculated for each well and normalized on the number of cells.
Western blot analysis
Cells were lysed using the sodium dodecyl sulfate (SDS) sample buffer (62.5 mM Tris-HCl pH 6.8, 2% SDS, 10% Glycerol, 50 mM DTT) and boiled for 5 min at 95°C before loading into a 15% polyacrylamide gel run at 15 mA. After transfer on PVDF membrane at 100 V constant for 2 h, proteins were incubated in 0.4% paraformaldehyde in PBS solution for 30 min at RT as previously reported [20] and then in blocking solution (10% milk) in TBS-T for 1 h. Membranes were further incubated O/N at 4°C in agitation with primary antibody. After the washing step with TBS-T, membranes were incubated with the secondary antibody for 2 h. Both primary and secondary antibodies were prepared in TBS-T supplemented with 5% non-fat milk diluted in TBS-T. Signal was detected using Clarity ECL reagents (Bio-Rad) in a dark chamber Uvitec Alliance (Uvitec) and the manufacturer software to acquire and analyze the data.
Statistical analysis
Statistical significance was assayed using a two-sided unpaired Student's t-test or two-way analysis of variance with Bonferroni post hoc test using the GraphPad Prism software. Data distribution was assessed for normality using the Shapiro–Wilk test. P < 0.05 was considered statistically significant.
Results
Modulation of wt aSyn expression in AF22 Tet-On aSyn cells
To generate a hNSC line for the inducible expression of wt aSyn, hiPSC-derived AF22 cells were infected with lentiviral particles carrying the pLVX-TetOn-Puro-human αSyn vector. Following infection, cultures were exposed to 1 μg/mL puromycin, chosen as optimal puromycin dose that allows for effective selection without altering the normal self-renewal potential of the resistant cells (Supplementary Fig. S1A). AF22 Tet-On aSyn cells selected with this puromycin dose exhibited low aSyn basal expression levels that increased 7.7-fold following treatment with doxycycline for 72 h (Supplementary Fig. S1B). Based on our previous experience with Tet-On systems, the abovementioned induction experiments were performed using a 750 ng/mL dose of doxycycline.
To test if AF22 Tet-On aSyn cells show a dose-dependent level of induction and to test at which dose of doxycycline it reached the maximum level of induction, we treated the cultures for 72 h with different concentrations of doxycycline (0, 100, 250, 500, 750, and 1,000 ng/mL). A clear dose-response relationship in the induction of aSyn expression levels was found, reaching a maximum plateau of eightfold induction already at 750 ng/mL doxycycline treatment (Supplementary Fig. S2A). Higher doses of doxycycline did not lead to a significant increase of aSyn induction levels. Based on these results, we confirmed 750 ng/mL doxycycline as the optimal dose for further analyses reported in this study.
Short-term time-course analysis of doxycycline induction on AF22 Tet-On aSyn cells (Fig. 1A) showed a progressive increase of aSyn expression levels along with time of doxycycline treatment (2.2-, 5.3-, and 7.9-fold induction at 24, 48, and 72 h of doxycycline treatment, respectively) (Fig. 1B). Immunofluorescence assay performed on these cultures showed that not all of the aSyn immunoreactive cells exhibited the same levels of expression, with even a fraction of the cells that barely showed any transgene expression (Fig. 1C). Quantitative analysis revealed that 72.21% ± 16.64% of cells in culture were aSyn+ve, the remaining showing very low or undetectable aSyn immunoreactivity (Fig. 1D). Furthermore, the pool of aSyn+ve cells can be divided in 30.3% ± 8.21% cells exhibiting high aSyn immunoreactivity (aSynhigh cells) and 69.67% ± 8.05% cells exhibiting low/medium aSyn immunoreactivity (aSynlow/Med cells) (Fig. 1E).
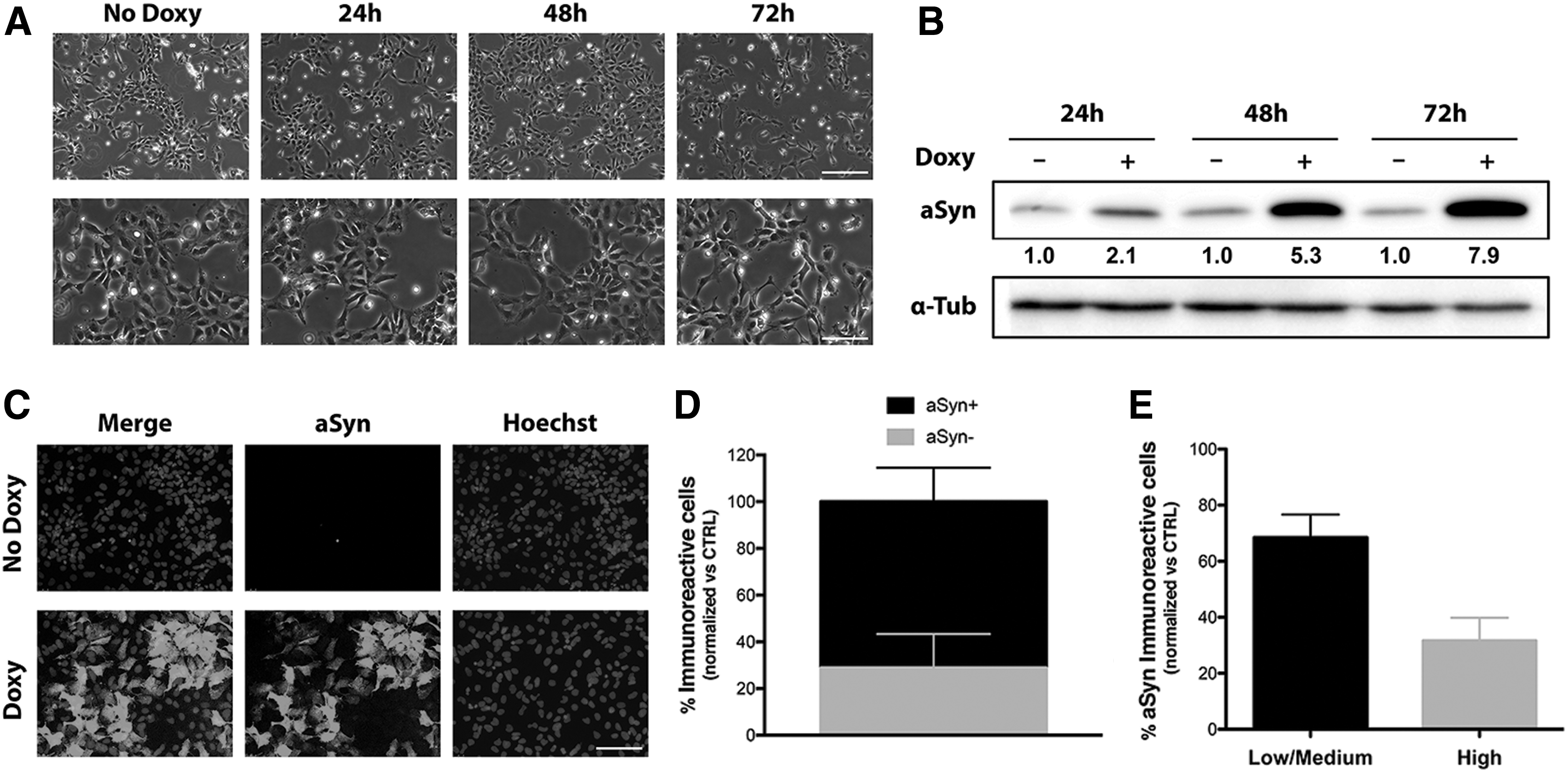
Modulation of wt aSyn expression in AF22 Tet-On aSyn cells.
Longer time-course analysis of doxycycline induction on AF22 Tet-On aSyn cells (Supplementary Fig. S2B) confirmed the progressive increase of transgene expression levels from 48 to 72 h of doxycycline induction and a strong immunoreactive band at 12 days of induction (2.0-, 7.4-, and 13.4-fold induction at 48 and 72 h and 12 days of doxycycline treatment, respectively) (Supplementary Fig. S2C). On the whole, these results indicate that aSyn induction in AF22 Tet-On aSyn cells is maintained long-term and progressive in aSyn accumulation.
wt aSyn overexpression affects cell division and cell viability in AF22 Tet-On aSyn cells
To test if aSyn induction in the cultures results in phenotypic abnormalities on hNSCs growth, we performed a cell growth assay on AF22 Tet-On aSyn cells. Since aSyn has been described to influence mitochondrial activity, we decided to avoid mitochondrial activity-based growth assays. We thus performed an automated high-throughput screening (HTS) cell count of stained cell nuclei on cultures fixed at defined time points.
aSyn induction produced a slight reduction in cell growth already after 72 h and this effect was more marked at later time points (Figs. 1A and 2A), where a strong impairment in the growth occurred (% reduction in cell number: 34.12 and 44.74 at 5 and 7 days, respectively) (Fig. 2A). These data could be interpreted by possible effects elicited by aSyn either on (1) cell division, (2) cell death, (3) change of fate by induction of differentiation, or (4) a combination of the abovementioned effects.
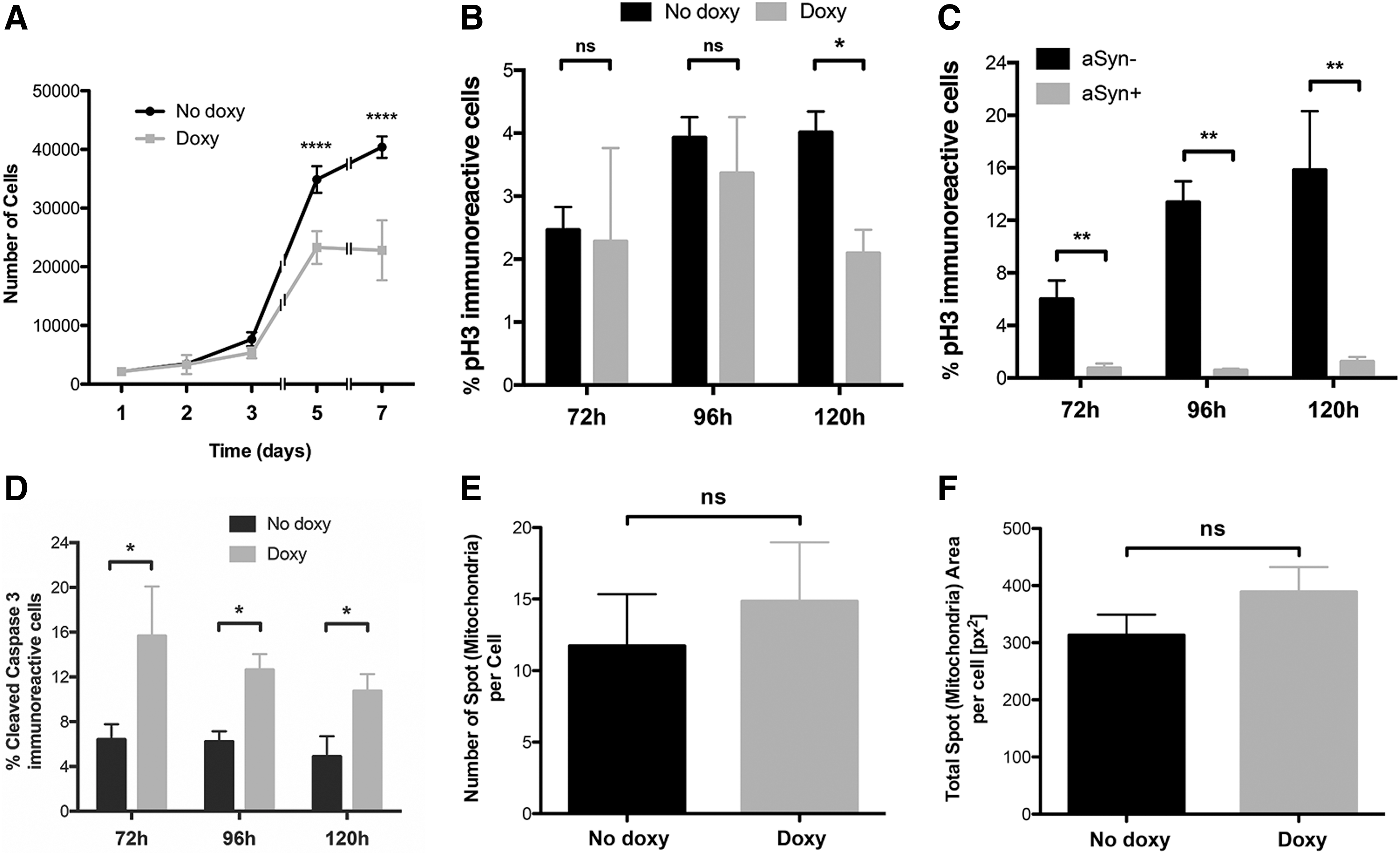
wt aSyn expression affects proliferation and cell viability of AF22 Tet-On aSyn cells.
To dissect out which of these possibilities was prominent, we performed specific assays. Analysis of phospho-Histone H3+ve cells present in the cultures at different time points (Supplementary Fig. S3) showed a reduction, although not statistically significant, of phospho-Histone H3+ve cells occurring both at 72 and 96 h of induction (Fig. 2B). A statistically significant 47.16% reduction was appreciated at 120 h (Fig. 2B). Furthermore, a stronger statistically significant reduction of phospho-Histone H3+ve cells in aSyn+ve cells compared with aSyn−ve cells was visible, at all the time points considered (% of reduction of phospho-Histone H3+ve cells in aSyn+ve cells in culture: 82.3, 96.79, and 91.54 at 3, 4, and 5 days, respectively) (Fig. 2C). These results indicate that aSyn induction affects cell division in AF22 Tet-On aSyn cells.
Furthermore, immunofluorescent analysis for NSC markers (Nestin and Sox2), neurons (β3-Tubulin), and astrocytes (GFAP) indicated that more than 97% of the cells retain their normal NSC identity without any significant induction of neuronal or glial cells (not shown), thus indicating that aSyn overexpression in self-renewing conditions does not force differentiation.
To test if aSyn induction could also impact on cell survival, we performed an immunofluorescent analysis for cleaved Caspase-3 at different time points following doxycycline treatment (Supplementary Fig. S4). aSyn induction led to a marked increase in the number of apoptotic cells in culture (fold increase of cleaved Caspase-3+ve cells in culture: 3.31, 2.03, and 2.19 at 3, 4, and 5 days, respectively) (Fig. 2D). Mitochondrial impairments have been explored in many aSyn assays both in vivo and in vitro. Besides impaired mitochondrial function, aSyn has been demonstrated to induce severe mitochondrial fragmentation in a number of cell culture studies. We thus explored by automated analysis, mitochondria number in basal and doxycycline-treated (5 days) cells in self-renewal condition (Fig. 2E, F and Supplementary Fig. S5). We did not find any significant variation on this parameter and also on mitochondrial size (not shown) in relation to aSyn induction.
On the whole, these results indicate that aSyn overexpression in hNSCs leads to a reduced cell growth by a double action, both by affecting proliferation capability and by enhancing cell death occurrence. Importantly, further analysis is required to dissect the contribution of mitochondrial dysfunction to these events.
Conditional overexpression of wt aSyn leads to the formation of intracellular aggregates in AF22 Tet-On aSyn cells
aSyn has been shown to generate aggregates inside of the cells and that these may contribute to cellular dysfunctions. Immunofluorescence analysis for aSyn in AF22 Tet-On aSyn cells following 12 days of doxycycline treatment showed that a fraction of aSyn+ve cells exhibited punctate immunoreactive dots in the cytoplasm (arrows in Fig. 3A). aSyn aggregates were present both in aSynhigh cells and aSynlow/Med cells, although the former presented higher number of aggregates with respect to the latter (number of aggregates: 0.48 ± 0.19 and 3.14 ± 0.32 in aSynlow/Med and aSynhigh cells, respectively) (Fig. 3B). Also, aggregates in aSynhigh cells exhibited a 5.8-fold increase in the immunoreactive signal with respect to the ones present in aSynmod/low cells (Fig. 3C). Furthermore, aSynhigh cells showed a 3.1-fold increase in the parameter of area of aggregated spots with respect to aSynmod/low cells (Fig. 3D).
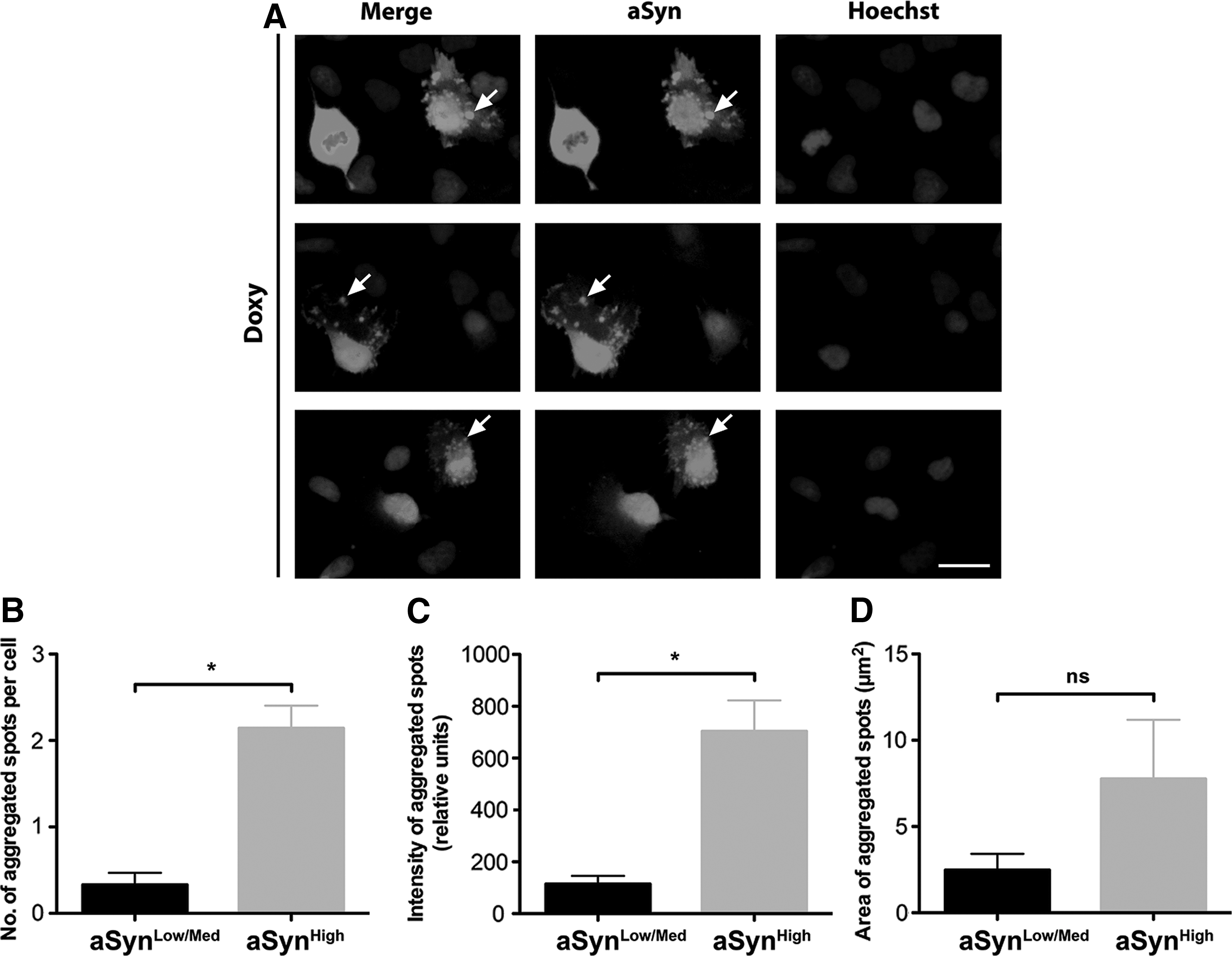
Conditional overexpression of wt aSyn leads to the formation of intracellular aSyn+ve aggregates in AF22 Tet-On aSyn cells.
To exclude that the aSyn+ve spots we detect in our cultures might be the result of nonspecific apoptosis-related structures instead of aggregation process, we performed a double immunofluorescent staining for Cleaved Caspase-3 (Supplementary Fig. S6). The presence of aSyn+ve spots in Cleaved Caspase-3−ve cells (Supplementary Fig. S6) excluded this possibility, nevertheless, additional biochemical analyses will be required to deeper investigate the properties of these structures.
These results indicate that aSyn overexpression produces aggregation on long-term induced AF22 Tet-On aSyn cells and that the occurrence of aggregation is dependent on the level of aSyn overexpression, with aSynhigh cells producing more aggregated spots with greater intensity with respect to aSynmod/low cells.
aSyn overexpression impairs neuronal differentiation in AF22 Tet-On aSyn cells
aSyn overexpression has been indicated as a main player in neuronal dysfunction. To test if aSyn induction in AF22 Tet-On aSyn cells could induce defects in the neuronal differentiation potential of the cultures, AF22 Tet-On aSyn cells (CTRL and cells maintained in doxycycline for the entire differentiation procedure) were exposed to a 2-week neuronal differentiation protocol. In these conditions, cells started to show morphological changes indicative of their progressive neuronal maturation. Immunofluorescence analysis on 14-day induced cultures showed that 73.25% ± 1.97% of the cells in culture were aSyn+ve (not shown). As expected, at this stage, not induced cultures were mainly composed of β3-tubulin+ve neurons (% of β3-tubulin+ve cells: 87.52 ± 6.09) (Supplementary Fig. S7A) with only a fraction of the cells in the culture positive for Map2, a marker for mature neurons (% of Map2+ve cells: 21.14 ± 4.35) (Supplementary Fig. S7B). On the contrary, doxycycline-treated cultures showed a 51.85% and 38.45% reduction in the overall number of β3-tubulin+ve cells and Map2+ve cells, respectively (Fig. 4B). These results indicate that aSyn overexpression partially affects neuronal differentiation capability of hNSCs.
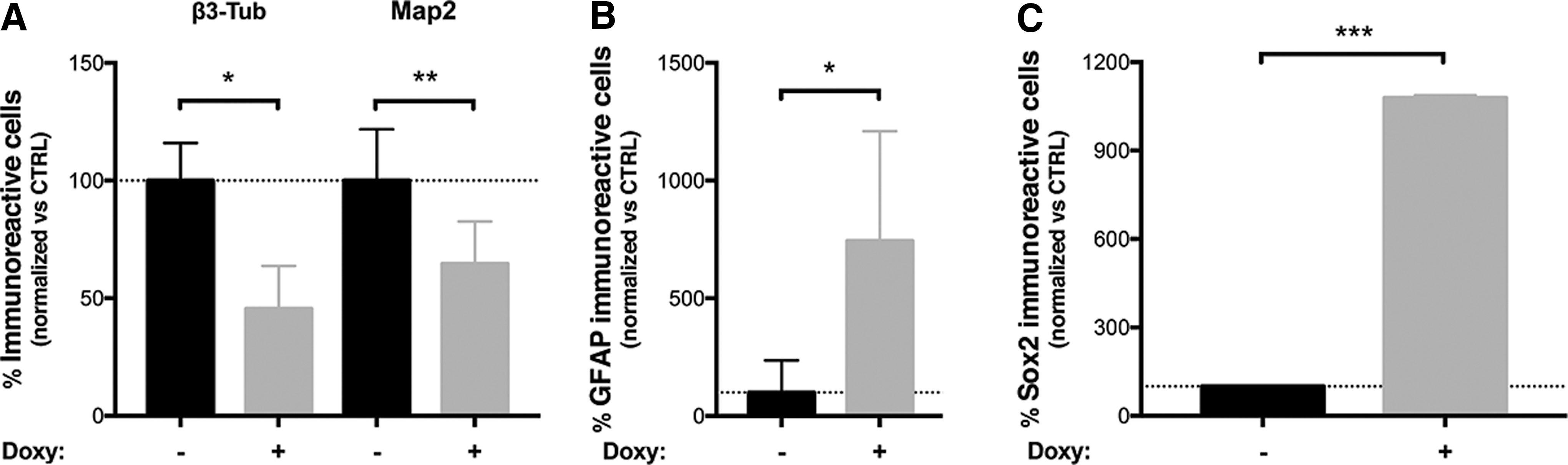
aSyn expression impairs neuronal differentiation of AF22 Tet-On aSyn cells.
We next asked if the reduction in the number of neuronal cells is mainly due to (1) induced competence to differentiate toward non-neuronal fates (ie, shift from neurogenic versus gliogenic fate) or (2) to an impaired competence of the cells to start the neuronal differentiation process. In this respect, we found that aSyn overexpression induced a 7.61-fold increase in the number of GFAP+ve cells (% of GFAP+ve cells: 0.46 ± 0.23 and 3.57 ± 0.61 in not induced and doxycycline-induced cells, respectively) (Fig. 4B and Supplementary Fig. S7C) in the cultures, thus indicating a possible enhanced propensity of the cells to differentiate toward the astrocytic lineage. Furthermore, Sox2 immunofluorescence showed the presence of large clusters of Sox2+ve cells in the doxycycline-treated culture with a 12-fold increase in the number of Sox2+ve cells, thus indicating that aSyn overexpression increased the percentage of the cell's refractory to undergo neuronal differentiation process (Fig. 4B and Supplementary Fig. S8).
On the whole, these results indicate that there is a combined effect of aSyn overexpression in disturbing hNSC neuronal or neurogenic process.
aSyn overexpression affects cell viability in AF22 Tet-On aSyn cell-derived dopaminergic neurons
AF22 cells have the competence to differentiate toward dopaminergic neuronal fate in defined in vitro conditions [19]. Thus, we next analyzed the possible effects elicited by aSyn acute induction in long-term differentiated hNSC-derived dopaminergic neuronal cultures. To this aim, self-renewing AF22 Tet-On aSyn cells were patterned for 3 weeks with FGF8/SHH and then induced to differentiate to dopaminergic neurons for 3 weeks as previously reported [19,21]. After 3 weeks of dopaminergic neuronal maturation, cultures were mainly composed of neurons (% of β3-tubulin+ve cells: 86.53 ± 7.17), most of which were dopaminergic neurons positive for Nurr-1 (% of Nurr-1+ve/β3-tubulin+ve neurons: 74.43% ± 8.26%) (Supplementary Fig. S9A) and tyrosine hydroxylase (TH) (Supplementary Fig. S9B).
At this stage, acute aSyn overexpression was achieved by treating the cultures for 4 days with 750 ng/mL doxycycline. aSyn overexpression produced an acute toxic effect resulting in the degeneration of the neuronal cells in culture (% reduction of the total number of neurons in aSyn-induced cultures: 24.72 ± 6.13) (Supplementary Fig. S9A) and by a 1.94-fold increase in the number of cleaved Caspase-3+ve cells (Fig. 5A and Supplementary Fig. S9C). Quantitative analysis showed that the fraction of Nurr-1+ve/β3-tubulin+ve neurons was significantly decreased following aSyn induction (% reduction of the number of Nurr-1+ve/β3-tubulin+ve neurons in aSyn-induced cultures: 31.72 ± 4.36) (Fig. 5B) with only a minor effect on the Nurr-1−ve/β3-tubulin+ve neurons, that were not significantly affected by acute aSyn overexpression (Fig. 5C). Thus, aSyn overexpression impairs the survival of hNSC-derived neurons, with dopaminergic neurons being differentially affected with respect to nondopaminergic neuronal subtypes.
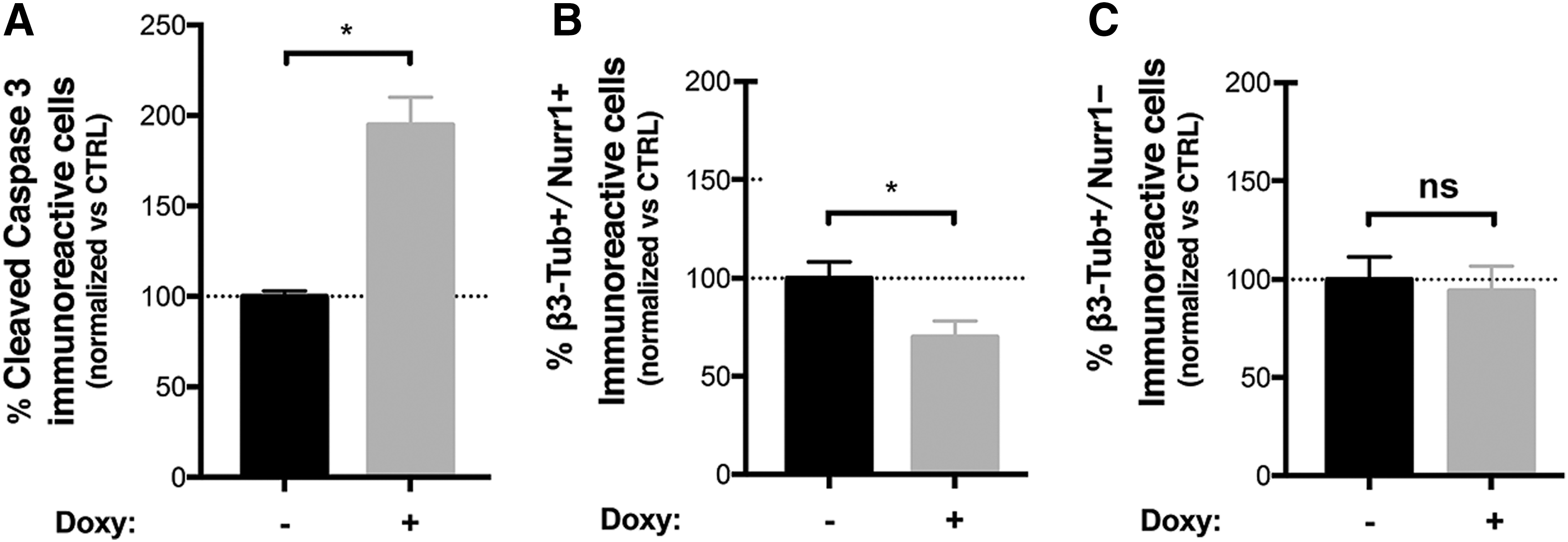
aSyn overexpression affects cell viability in AF22 Tet-On aSyn cell-derived dopaminergic neurons.
Discussion
Although aSyn is considered of relevance for PD pathogenic process, the molecular mechanism triggering PD starting from aSyn homeostasis alteration is still a matter of debate. In this study, we report the generation of a novel cellular model based on the overexpression of wt human aSyn in NSCs. The system is characterized by the controlled expression of wt human aSyn by means of a Tet-On inducible mechanism and represents a valuable tool to study the effects of aSyn overexpression in hNSCs and neurons.
The AF22 Tet-On aSyn cell model developed in this study has several features that make it attractive for studying the biological and molecular effects elicited by aSyn in different developmental paradigms. These include the hNSC nature of the parental AF22 cells. Originally, described by Falk et al. [19], these cells have been obtained from normal hiPSCs and show features that make them ideal as parental cells to be engineered. Indeed, they are homogeneously composed of self-renewing NSCs characterized by genomic stability and high amenability to genetic manipulation. Importantly, these cells maintain a stable high neurogenic capability along long-term in vitro expansion and the competence to respond to specific patterning cues that allows generating defined neuronal subtype populations, including dopaminergic neurons.
The inducible nature of aSyn expression coupled to the NSC system opens to the possibility to study both acute and chronic aSyn-mediated effects in defined relevant cell populations, that is, neural progenitors and mature neurons. Also, this is instrumental to sort out specific effects in several processes, including self-renewing, lineage commitment, and neuronal maturation and/or maintenance.
aSyn inducible systems have been reported from different immature parental cell lines, mainly PC12 cells and human neuroblastoma lines [12,13,22 –24]. Nonetheless, these systems have some intrinsic limitations that are overcome in AF22 Tet-On aSyn cell model. Indeed, PC12 cells are of rodent origin and species-specific differences in aSyn sequence and roles have been reported [25 –27]. Additionally, both PC12 cells and neuroblastoma lines are transformed and share a non-CNS origin. Noteworthy, their neurogenic potential is limited in terms of efficiency and quality of neuronal-like cells that can be obtained following their differentiation, thus limiting their physiological relevance for studies aimed at dissecting aSyn roles in human CNS neurons.
AF22 Tet-On aSyn cells show a clear dose–response transgene expression with a robust aSyn induction up to seven- to eightfold; also, induction can be maintained long term leading to a progressive increase in aSyn levels. When we tried to induce aSyn aggregation in AF22 Tet-On aSyn cells, we found that this process requires a prolonged induction period allowing to aSyn levels to increase progressively. The appearance of aSyn aggregates in AF22 Tet-On aSyn cells occurs after 12 days of induction. Other studies performed on PC12/TetOn aSyn inducible systems failed to observe wt aSyn aggregates in proliferating cells [22]. This could be due to the different origins of the parental cells or to the fact that levels of aSyn induction are lower or due to the quite reduced time of induction with respect to our study. Also, several reports have shown different oligomeric aSyn forms and prion-like transmission of aSyn in vitro and in vivo. Further investigation is required to dissect these aspects in our cellular system, both in self-renewal and differentiating conditions.
It is interesting to note that wt aSyn overexpression induces phenotypic defects in hNSCs in the self-renewing state. Other studies have reported that overexpression of aSyn in the proliferating state fails to induce any cell death in proliferating neural-like cells, despite the prominent accumulation of aSyn aggregates [12]. The factors accounting for the differential death effects could include differences in clearance mechanisms, or involvement of cell cycle molecules or other proteins differentially expressed in the two states.
An increasing number of studies reveal that aSyn may play an important role in neurogenesis. When the SNCA gene is differentially expressed or bears mutations, the in vivo NSC pool is negatively regulated and both neurogenesis and survival of newly generated neurons are decreased [9]. These studies suggest that a link might exist among neurogenesis, aSyn, and neurodegenerative diseases [18].
In vitro, aSyn overexpression has been described to reduce the number of mouse secondary neurospheres formed and to affect NSC morphology and cell cycle progression, leading to their accelerated differentiation [10]. hiPSC-derived neuronal precursor cells from a PD patient carrying a genomic triplication of the SNCA gene showed substantial impairments in growth, viability, cellular energy metabolism, and stress resistance. These effects were exacerbated when the cultures were challenged by starvation or toxic stimuli [16]. Also, overexpression of wt aSyn in expanded populations of progenitors derived from the human fetal cortex showed a slight effect on cell growth and a progressive impairment of lineage commitment competence [17]. Similarly, studies on human embryonic stem cell (hESC)-derived neural progenitors overexpressing wt aSyn and on neural progenitors obtained from hiPSCs from a PD patient with a SCNA locus triplication, showed an increased cell death and reduced neurogenic capacity compared with control cultures [17,28].
Our results confirm that overexpression of human wt aSyn impairs the process of differentiation of hNSCs into neuronal cells. In particular, we have seen a marked reduction on the efficiency of the cells to generate neurons when exposed to neuronal differentiative cues with a parallel increase in the number of astrocytes and undifferentiated cells.
Further studies are required to dissect the molecular mechanisms triggering these specific defects. PSC-derived long-term expandable hNSCs, including the AF 22 cells, are highly responsive to efficiently undergo neuronal differentiation when exposed to the neuronal differentiation protocol employed in this study[19,21]. Additional investigation is required to define if this defect is maintained when exposing the cultures to other proneuronal differentiation conditions. Furthermore, exposure to non-neuronal (ie, gliogenic) differentiation cues could help to understand if the observed refractoriness to exit self-renewal is specific for the transition toward the neuronal lineage or is a more general aSyn-mediated effect. Finally, we cannot exclude aSyn-selective toxicity toward specific cell types might contribute to these alterations. Besides this, we have in this study reported an aSyn-driven effect on hNSCs neurogenic process that results in increased number of astrocytes found in neuronally differentiating cultures.
Other studies have reported that wt aSyn overexpression in human neural progenitors derived from fetal cortex preserved the neurogenic competence of the cells following long-term expansion [17]. To this respect, we can speculate that this discrepancy might be related to the different nature and identity between our and the abovementioned cell system. Indeed, differently from fetal brain-derived cell systems that are representative of late developmental neural stages and in which the neurogenic competence quickly declines with in vitro passages, the PSC-derived hNSCs we employed are representative of earlier developmental neural stages and are extremely stable also following extensive long-term expansion [29].
Finally, we have observed an aSyn-mediated acute toxicity in hNSC-derived neurons, being dopaminergic neurons selectively affected. These results are in agreement with a previous study reporting acute aSyn toxicity in hESC-derived neuronal cultures [17]. Those authors showed that hESC-derived neuronal cultures are highly vulnerable to expression of both wt aSyn or mutant aSyn forms, with dopaminergic neurons exhibiting higher toxic susceptibility with respect to nondopaminergic (GABAergic) neurons. It is yet unclear the reason of this neuronal subtype's selective cytotoxicity. The factors accounting for this differential death effects in different neuronal subtypes are unknown but could include differences in clearance mechanisms or other proteins differentially expressed in the two states. Interestingly, aSyn overexpression has been shown to directly affect TH expression, suggesting possible direct TH effects [30]. Regardless of the exact reason, this fact further validates the current model as one in which toxic effects occur preferentially in dopaminergic neurons.
In conclusion, we have developed a cell system for controlled expression of wt aSyn in hNSCs that exhibit defined aSyn-driven phenotypes both in self-renewal and differentiating/differentiated stages. This novel inducible model may prove valuable in the deciphering of aSyn-mediated pathogenic effects and in the assessment and screening of potential therapeutic strategies.
Footnotes
Acknowledgments
The authors wish to thank Prof. Austin Smith (Cambridge Stem Cell Institute, University of Cambridge) for providing the AF22 cell line, Dr. Tilo Kunath (MRC Center for Regenerative Medicine, University of Edinburgh, UK) for the pLVX-TetOn-Puro-human aSyn vector, and Prof. Massimo Pizzato for the psPax2 vector. They also thank the personnel of Cibio HTS Core Facility and of Advanced Imaging Core Facility. This work was supported by the intramural funding from the University of Trento.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.