
Abstract
While mesenchymal stem cells (MSCs) are promising candidates for inner ear hair cell regeneration, to date, there have been no convincing reports indicating whether MSCs can survive in the cochlea for more than a few weeks, as the high levels of potassium (K+) in the endolymph (EL) are thought to be toxic to transplanted stem cells. For conditioning the EL for MSC transplantation, we conducted this in vitro study to examine the effects of artificial EL with altered K+ concentration levels, in the range of 5–153.8 mM, on proliferation, apoptosis, and morphological changes in MSCs derived from various human tissues. Our findings demonstrate that altering the K+ concentration in artificial EL could significantly influence the survival of MSCs in vitro. We discovered that K+ concentrations of 55–130 mM in artificial EL could enhance the survival of MSCs in vitro. However, MSCs exhibited reduced proliferation regardless of K+ concentration.
Introduction
S
Fortunately, over the past few years, stem cell therapy has brought new hope for regenerating HCs and restoring hearing physiologically [5 –8]. In stem cell therapy, stem cells are transplanted into the endolymph (EL), which is a special extracellular solution characterized by a high potassium (K+) concentration of ∼153.8 mM [9,10] that fills the scala media of the cochlea; the HCs from the epithelial layer of the cochlea expose their stereocilia into this solution. Mesenchymal stem cells (MSCs), a type of stem cell that can be obtained from a patient's own tissue, are promising candidates for HC regeneration.
MSCs not only exhibit the capacity for self-renewal and the ability to differentiate into various specific cell types, such as osteoblasts, adipocytes, chondrocytes, myocytes, and neurons, but also have the advantages of autologous transplantation and immunosuppressive features. Such features overcome ethical issues and immune reactions inherent to other stem cell types, making MSCs more likely to become a therapeutic reality. Furthermore, in recent years, many studies demonstrated that MSCs derived from various tissues [11 –13] can differentiate into inner ear progenitor cells, HCs, and neurons both in vitro and in vivo. All of this evidence highlighted the potential of MSCs as a promising tool for inner ear hair regeneration in deaf animals.
The cochlea is located deep inside the temporal bone and is divided into three compartments: the scala tympani, scala media, and scala vestibule. The scala tympani and vestibule is the space between the bone and membranous labyrinth, and is filled with perilymph (PL), whereas the scala media is filled with EL inside the membranous labyrinth. Therefore, for HC replacement therapy, there are two routes for transplanting MSCs into the cochlea: intraperilymphatic transplantation and intraendolymphatic transplantation [14,15]. The HCs are located on the auditory epithelium (ie, organ of Corti), which sits upon the basilar membrane. However, when MSCs are transplanted into the PL (through the scala tympani or scala vestibule), they cannot easily cross the basilar membrane to migrate into the scala media and integrate into auditory epithelium.
Thus, it is more reasonable to transplant the MSCs directly into the scala media, which can help to target the auditory epithelium. Nevertheless, the EL in the scala media is a unique extracellular solution characterized by a high K+ concentration (153.8 mM), which is toxic to the transplanted cells [14,16]. Although Hildebrand et al. [17] found that partially differentiated mouse embryonic stem cells could survive in the EL after transplantation by the scala media, Lang et al. [18] speculated that it is the surgical trauma itself that disrupted the basilar membrane and caused the mixing of EL and PL, thereby diluting the K+ concentration of the EL to allow for cell survival.
Therefore, to determine whether K+ itself could play a key role in the survival or apoptosis of MSCs and to further ascertain the appropriate K+ concentration in the EL that allows MSCs to survive, we examined the effects of artificial EL with altered levels of K+ concentrations, in the range of 5–153.8 mM, on the in vitro proliferation, apoptosis, and morphological changes of MSCs derived from various human tissues. The results of this study demonstrate that when only changing the K+ concentrations in artificial EL, the survival of MSCs can be greatly influenced in vitro. Furthermore, we found that when K+ concentrations are moderately high, ranging from 55–130 mM, artificial EL can support the survival of MSCs of different origins in vitro, with glycogen particulates forming within the cytoplasm. However, artificial EL cannot support the growth of MSCs regardless of the K+ concentration.
Materials and Methods
Ethics statement
This study has obtained the approval of Ethics Committee of the Basic Medical Sciences, Chinese Academy of Medical Sciences (No. 003-2014). Tissues for sourcing various types of MSCs were collected from patients or donors after obtaining their informed consent. All methods were performed in accordance with the institutional regulations and ethical guidelines of Ethics Committee of the Basic Medical Sciences, Chinese Academy of Medical Sciences.
Preparation of human MSCs from adipose tissue, bone marrow, placenta, and umbilical cord
Isolation and culture of adipose-derived MSCs
Raw lipoaspirates collected from patients undergoing selective suction-assisted lipectomy were thoroughly washed with
Isolation and culture of bone marrow-derived MSCs
Human bone marrow aspirates were obtained from unrelated donors. Mononuclear cells were fractionated over a 1.077 g/mL Ficoll-Paque density gradient and depleted of hematopoietic cells using MACS CD45, GlyA, and CD34 micromagnetic beads (Miltenyi Biotec, Inc., Auburn, CA) [19,20]. The cells were washed twice and plated in T-75 tissue culture flasks at 1 × 106/mL. The expansion medium was the same as that used for adipose-derived MSCs (AMSCs). Once adherent cells were >70% confluent, they were detached with 0.125% trypsin and 0.01% EDTA and replated at a 1:3 dilution under the same culture conditions. Confluent cells (∼2 × 106) at the third passage were used for the experiments.
Isolation and culture of placenta-derived MSCs
Human placental tissues obtained from unrelated donors were washed thrice with
Isolation and culture of umbilical cord-derived MSCs
Human umbilical cords were obtained from healthy donors after routine deliveries. The umbilical cords were washed thrice with
All experiments were performed with cells from the third passage.
Immunophenotypic analysis
For immunophenotypic analysis of MSCs, cells at passage 3 were incubated for 30 min on ice in the dark with the following fluorochrome-conjugated antibodies: CD29, CD90, CD105, CD44, CD73, CD31, CD34, CD45, and HLA-DR (BD Biosciences, San Diego, CA). Cells were then fixed with 4% paraformaldehyde, kept at 4°C, and analyzed within 24 h using a fluorescence-activated cell sorter (FACS Vantage SE; BD Biosciences).
Osteogenic and adipogenic differentiation
To identify the MSC capacity for multilineage differentiation, MSCs were cultured under differentiation conditions. Culture-expanded cells at 2 × 104/cm2 were induced in the following osteogenic medium for 2 to 3 weeks: DMEM supplemented with 10% FCS, 10 mM β-glycerophosphate, 10−8 M dexamethasone, and 0.2 mM ascorbic acid (all from Sigma). Cells were then stained with the alkaline phosphatase (ALP) staining kit (Beyotime, Haimen, China) to reveal osteogenic differentiation.
To assess the adipogenic differentiation ability, culture-expanded cells at 2 × 104/cm2 were induced for 3 weeks in DMEM supplemented with 10% FCS, 0.5 μM hydrocortisone, 0.5 μM isobutylmethylxanthine, and 50 μg/mL indomethacin (all from Sigma). At the end of the culture period, cells were fixed in 10% formalin for 10 min and stained with fresh Oil Red O solution (Sigma) to visualize lipid droplets in the induced cells.
Preparation of artificial EL with altered potassium concentrations
The composition of the artificial EL medium (pH 7.4, osmolality 300 mOsm/kg H2O) was as follows (in mM): KCl, 126; NaCl, 1; KHCO3, 25; MgCl2, 0.025; CaCl2, 0.025; K2HPO4, 1.4; and mannitol, 25 [9,10]. To study the influence of potassium on MSCs, the K+ concentration of the artificial EL medium was altered for the treatment groups in our study as follows: 130, 105, 80, 55, 30, and 5 mM (pH 7.4). In the K+ concentrations of 153.8–30 mM, we modified KCl alone and the 25 mM KHCO3 was preserved, while in 5 mM K+, we leave the KCl alone by 5 mM. The osmolality was kept constant with 300 mOsm/kg H2O by adding mannitol [10]. Natural EL was composed of 153.8 mM K+, 1.3 mM Na+, 132 mM Cl−, 31 mM HCO3 −, and 0.023 mM Ca2+ (pH 7.4) [9,10,21,22].
Apoptosis assay
MSCs seeded in 60-mm plastic tissue culture dishes were exposed to respective test media for 24 h. Cells undergoing apoptosis were detected using an Annexin V-FITC Apoptosis Detection Kit (Keygen, Nanjing, China). The MSCs were digested with 0.25% trypsin for 1 min at 37°C, resuspended at a concentration of 5 × 105 cells per mL, and centrifuged at 2,000 rpm for 5 min. Next, the cells were collected in phosphate-buffered saline (PBS) at 4°C, washed twice, and resuspended in 500 mL binding buffer (Keygen). After 5 μL Annexin V-FITC (Keygen) was added and mixed well, 5 μL propidium iodide (Keygen) was added and mixed well. The reaction was performed at 20°C for 5–15 min in the dark, and flow cytometry (FACSort; Becton Dickinson, San Jose, CA) was performed. All experiments were repeated thrice.
Cell proliferation assay
MSCs were seeded in 96-well cell culture plates (2,000 cells per well) and incubated for 8 h with the expansion medium. Once the MSCs were attached to the 96-well plate, the expansion medium was removed and cell lines were incubated with respective test media (control: DMEM/F-12; group E: 153.8 mM artificial EL; group E1: 130 mM K+ artificial EL; group E2: 105 mM K+ artificial EL; group E3: 80 mM K+ artificial EL; group E4: 55 mM K+ artificial EL; group E5: 30 mM K+ artificial EL; and group E6: 5 mM K+ artificial EL). Cells were cultured in 96-well plates for 24 h and incubated with 20 μL per well 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethyl phenyl)-2-(4- sulfophenyl)-2H-tetrazolium inner salt (MTS) reagent (CellTiter 96® AQueous One Solution Reagent, Catalogue No. G3581; Promega, Madison, WI) at 37°C for 3 h. Absorbance was measured using a spectrophotometer at a wavelength of 490 nm. Background absorbance was measured in a control well containing only DMEM/F-12 and MTS reagent. All experiments were repeated thrice.
Caspase-3 colorimetric protease assay
The enzyme activity of caspase-3 was measured in MSC lysates with a colorimetric caspase-3 assay kit (CaspACE™ Assay System, Colorimetric, Catalogue No. G7220; Promega). MSCs seeded in six-well plates were exposed to respective test media for 24 h. Cells were lysed by freeze-thaw and then incubated on ice for 15 min. Cells were centrifuged at 15,000g for 20 min at 4°C, and the supernatant fraction was collected. The 100 μL reaction mixture contained 54 μL cell lysate, 2 μL dimethyl sulfoxide, 10 μL dithiothreitol (100 mM), and 2 μL DEVD-pNA substrate (10 mM stock) in 32 μL assay buffer. The assay was performed in a 96-well plate. The mixture was incubated for 3 h at 37°C, and the absorbance was measured at 405 nm. All experiments were repeated thrice.
Transmission electron microscopy
After exposure to respective test media for 24 h, MSCs were fixed at room temperature in 2.5% glutaraldehyde in 0.1 M PBS for 2 h. The cells were washed overnight at 4°C in the same buffer and post-fixed with 1% OsO4 buffered with 0.1 M PBS for 2 h. The pellets were dehydrated in a graded series of ethanol and embedded in a mixture of Epon-812 araldite. Ultrathin (60–70 nm) sections were collected on copper grids, double stained with uranyl acetate and lead citrate, and examined by transmission electron microscopy (TEM) (JEOLJEM-1010; JEOL Ltd., Japan; Gatan CCD camera).
Detection of cytokines secreted from MSCs
After 1 × 106 cells per well were plated in six-well plates and incubated with respective test media for 24 h, cell culture supernatants were assayed using interleukin (IL)-2, IL-6, IL-10, and tumor necrosis factor (TNF)-α ELISA Kits (Neobioscience, Shenzhen, China) according to the manufacturer's protocol. The absorbance of each well at 450 nm was measured.
Statistical analysis
All data are expressed as the mean ± standard error. GraphPad Prism 6.05 software was used for analysis. Statistically significant differences between groups were assessed by a one-way or two-way analysis of variance (ANOVA), followed by Tukey's multiple comparisons test. P-value <0.05 was considered statistically significant.
Data availability
The datasets generated and/or analyzed during this study are available from the corresponding author on request.
Results
Identification and characterization of MSCs of various origins
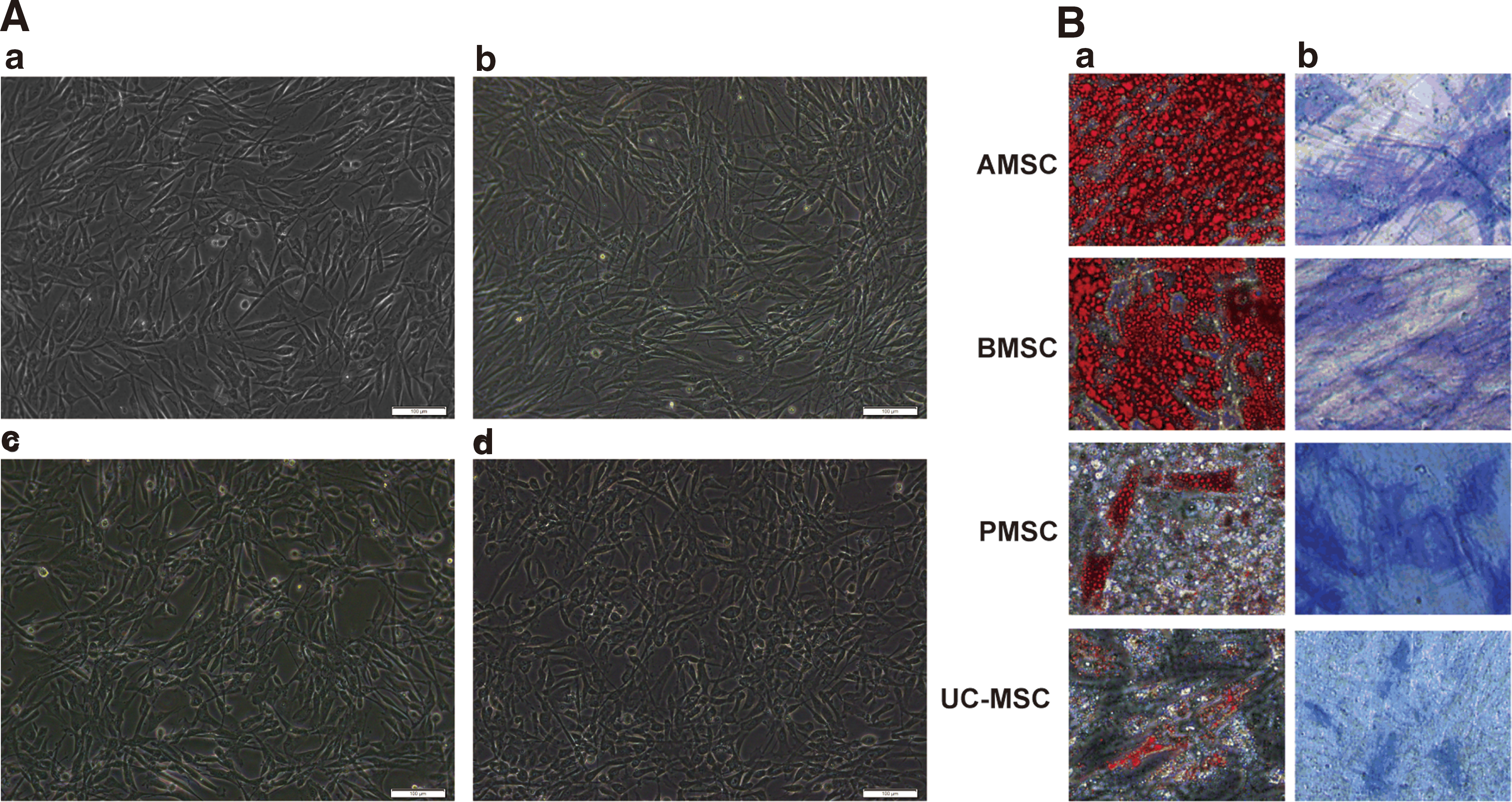
All adherent cultured MSCs from human AMSCs, bone marrow (BMSCs), placenta (PMSCs), and umbilical cord (UC-MSCs) displayed fibroblast-like morphologies in normal MSC culture medium when observed under light microscope, as shown in Fig. 1. To characterize the phenotypes of the MSCs, we investigated their immunophenotypes using flow cytometry (Table 1). The results showed that all cells were positive for CD29, CD90, CD105, CD44, and CD73 and negative for CD31, CD34, CD45, and HLA-DR (Table 1). Furthermore, under adipogenic and osteogenic differentiation conditions, all cells could be differentiated into adipocytes and osteoblasts, which were separately confirmed by Oil Red O and ALP staining (Fig. 1). These features imply that the cells cultured in this research are all MSCs that exhibit the typical characteristics of MSCs [23,24].
Moderately high K+ concentrations in artificial EL inhibit apoptosis in MSCs
With the goal of elucidating the effects of artificial EL with different K+ concentrations on the survival of MSCs derived from various tissues, we measured the rates of apoptosis in all MSCs by flow cytometry using Annexin V-FITC/PI staining after 24 h of cell culture. Populations of apoptotic cells, including early apoptotic (FITC+/PI−) and late apoptotic (FITC+/PI+) cells, were quantitated using this method (Fig. 2). To compare differences in the apoptotic rates between groups, all data were subjected to a one-way ANOVA followed by Tukey's tests (Fig. 3). Because of the existence of the blood–labyrinthine barrier, an important structure between the blood and inner ear fluids, the microenvironment and composition of EL are quite different from that of blood [25 –29]. In this research, therefore, we used cells cultured in MSC culture medium, DMEM/F-12, as our control group.
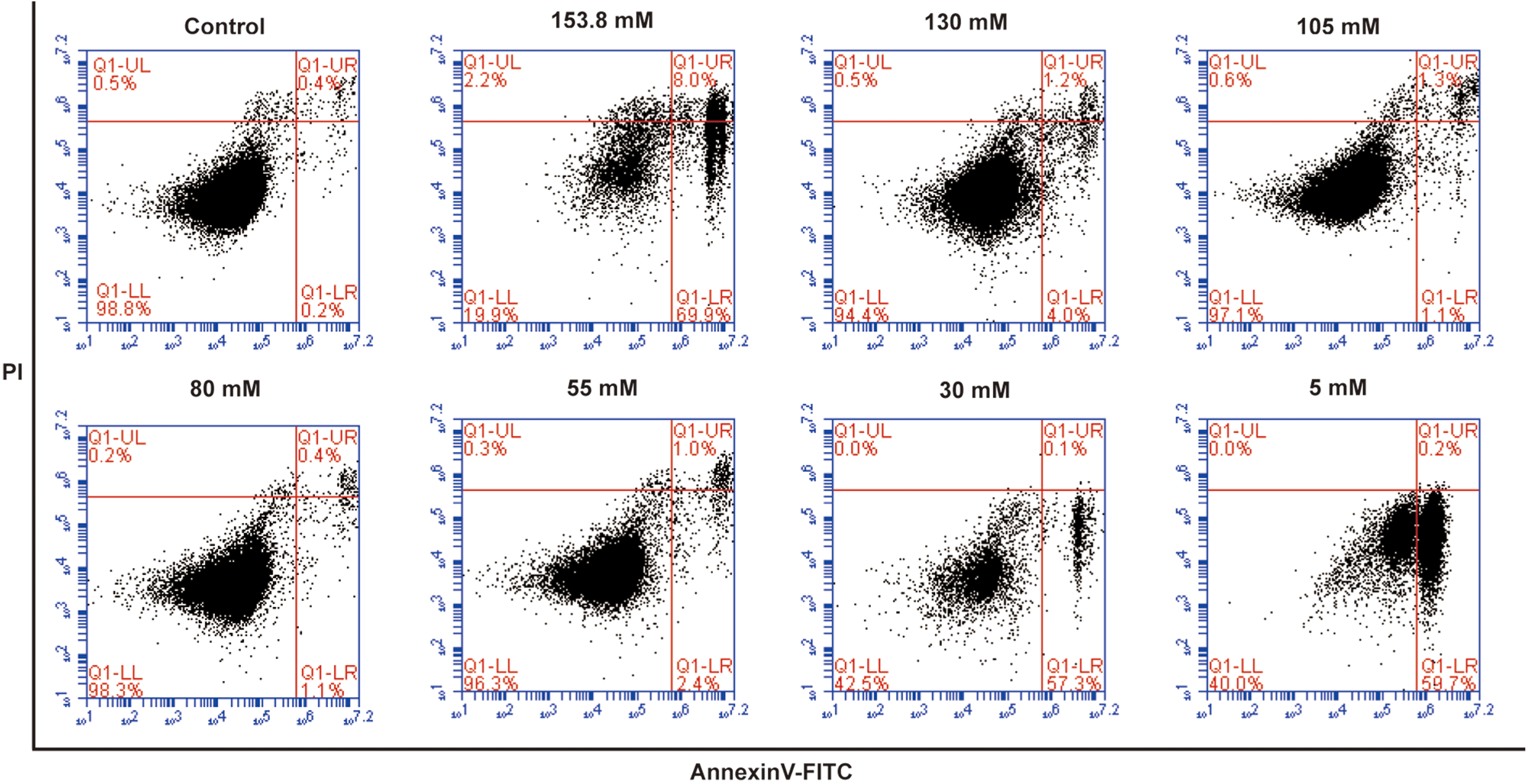
Flow cytometric analysis of apoptosis in AMSCs cultured in artificial EL with different potassium concentrations, stained with Annexin V-FITC and PI (control group: DMEM/F-12). Viable cells are FITC−/PI−, early apoptotic cells are FITC+/PI−, late apoptotic cells are FITC+/PI+, and necrotic cells are FITC−/PI+. Each dot plot represents 1 of 3 independent experiments. EL, endolymph. Color images available online at
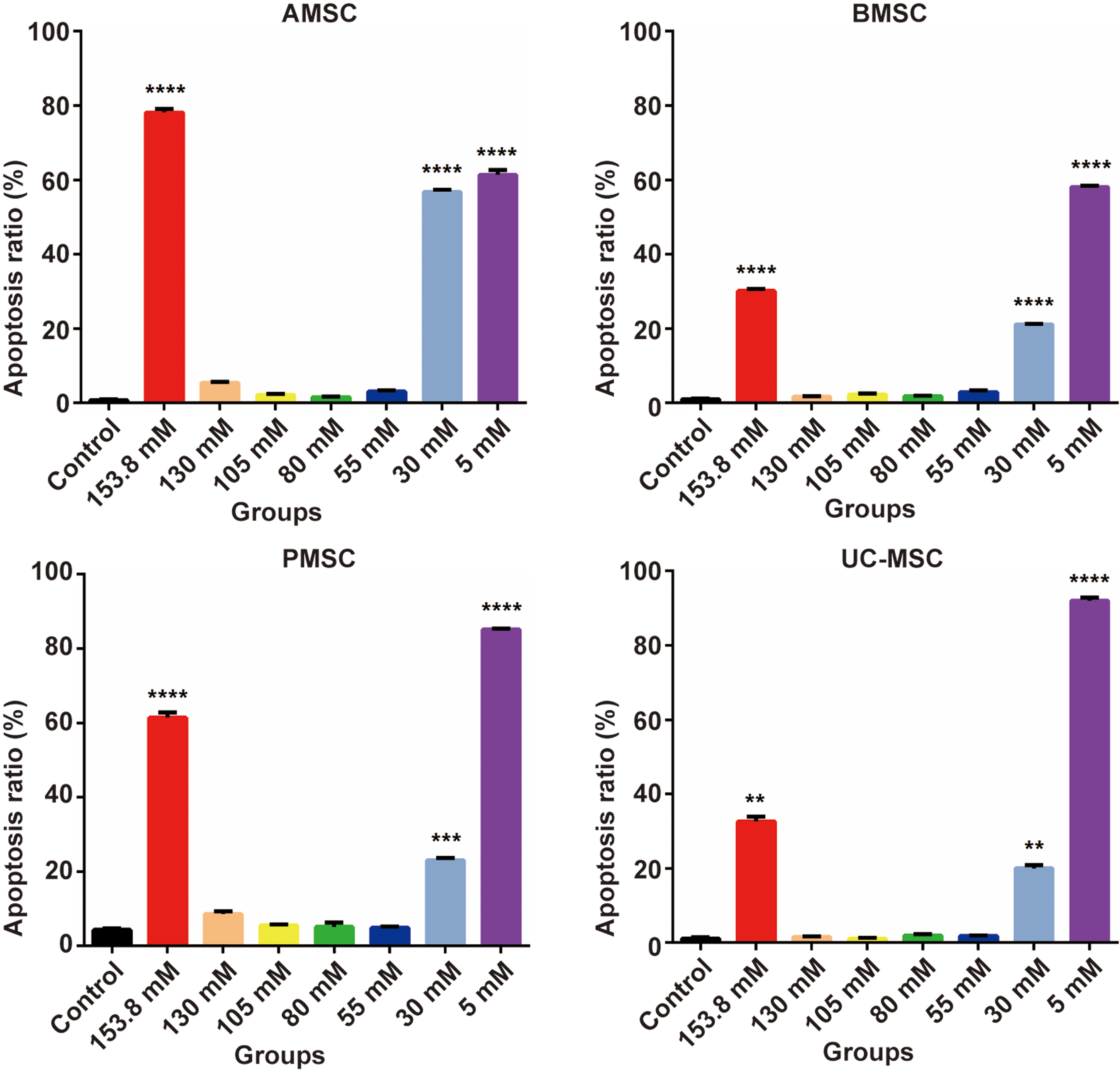
Comparison of total apoptotic rates among groups (n = 3). MSCs were cultured in artificial EL with different K+ concentrations for 24 h. The results are expressed as the mean ± standard deviation of three independent experiments. **P < 0.01, ***P < 0.001, ****P < 0.0001 vs. control group. Color images available online at
As depicted in Fig. 2, in artificial EL with K+ concentrations of 153.8, 30, or 5 mM, the percentages of apoptotic AMSCs were 78.17 ± 0.93%, 56.77 ± 0.57%, and 61.40 ± 1.32%, respectively; these values were significantly higher than the percentages in the 130, 105, 80, and 55 mM K+ groups (P < 0.0001). In contrast, cells exposed to 130, 105, 80, and 55 mM K+ exhibited slightly higher apoptotic rates than those of the control group (P < 0.05 or P < 0.01) (Fig. 3). Rates of apoptosis in treated BMSCs, PMSCs, and UC-MSCs were similar to those seen in AMSCs, with significantly increased apoptosis in the 153.8 mM, 30, and 5 mM K+ artificial EL groups and only slight increases (P < 0.05 or P < 0.01), no significant change, or even inhibition of apoptosis (P > 0.05) in the 130, 105, 80, and 55 mM K+ groups (Fig. 3). The proportion of necrotic cells (FITC−/PI+) was below 10% in all groups after 24 h of culture (Fig. 2).
Artificial EL with altered K+ concentrations suppresses MSC proliferation
We next assessed cell growth by means of an MTS assay performed at different time intervals (24, 48, 72, or 120 h), as shown in Fig. 4. We performed a two-way ANOVA across all groups to determine significant differences. Generally, MSCs from different tissues exhibited similar trends compared with those in the control groups, and all cells in artificial EL ceased to proliferate with prolonged culture time, regardless of the potassium concentration (P < 0.0001). Nevertheless, cells exposed to 130, 105, 80, and 55 mM K+ in artificial EL exhibited better survival after 24 and 48 h of cell culture than those exposed to 153.8 mM K+ in artificial EL (P < 0.05). In addition, there were no significant differences between the 130, 105, 80, and 55 mM K+ groups (P > 0.05) or between the 30 and 5 mM K+ groups (P > 0.05).

Proliferation analyses of MSCs cultured in artificial EL with different potassium concentrations. Cells were incubated with MTS reagent for 3 h, and changes in proliferation were studied at different time intervals (24, 48, 72, and 120 h). The data are represented as the mean ± SEM optical density at 490 nm. All artificial EL groups with altered potassium concentrations ceased to proliferate and exhibited significant differences compared with the control group (P < 0.0001). SEM, standard error of the mean. Color images available online at
Caspase-3 might contribute to MSC apoptosis in artificial EL with very high or low K+ concentrations
To estimate the contribution of caspase-3 to the apoptosis of MSCs, we investigated caspase-3 activity after cells were treated with artificial EL with altered K+ concentrations for 24 h. Interestingly, treated MSCs from the four different tissues exhibited similar caspase-3 activity. As shown in Fig. 5, colorimetric assays revealed that caspase-3 activity was markedly higher in the 153.8, 30, and 5 mM K+ artificial EL groups (P < 0.01 or P < 0.001) than in the control group. In contrast, no increase in caspase-3 activity was observed in the 130, 105, 80, or 55 mM K+ groups (P > 0.05).

Caspase-3 activity in MSCs cultured in artificial EL with different K+ concentrations for 24 h. Data are represented as the mean ± SEM of three individual experiments. **P < 0.01, ****P < 0.0001 vs. control group.
MSCs cultured in artificial EL with moderately high K+ concentrations survived and formed glycogen particulates
After being cultured in artificial EL with altered K+ concentrations for 24 h, the MSCs from the four different tissues were observed under a light microscope (Fig. 6). In contrast to the control group, cell morphology in the 153.8 and 30 mM K+ artificial EL groups changed from fibroblast like to round, while in the 5 mM K+ artificial EL group, cells became shrunken and lost the normal cell skeleton (Fig. 6). In the 130, 105, 80, and 55 mM K+ groups, several soap bubble-like vacuoles appeared within the MSCs, although they still displayed fibroblast-like cell morphology (Fig. 6).
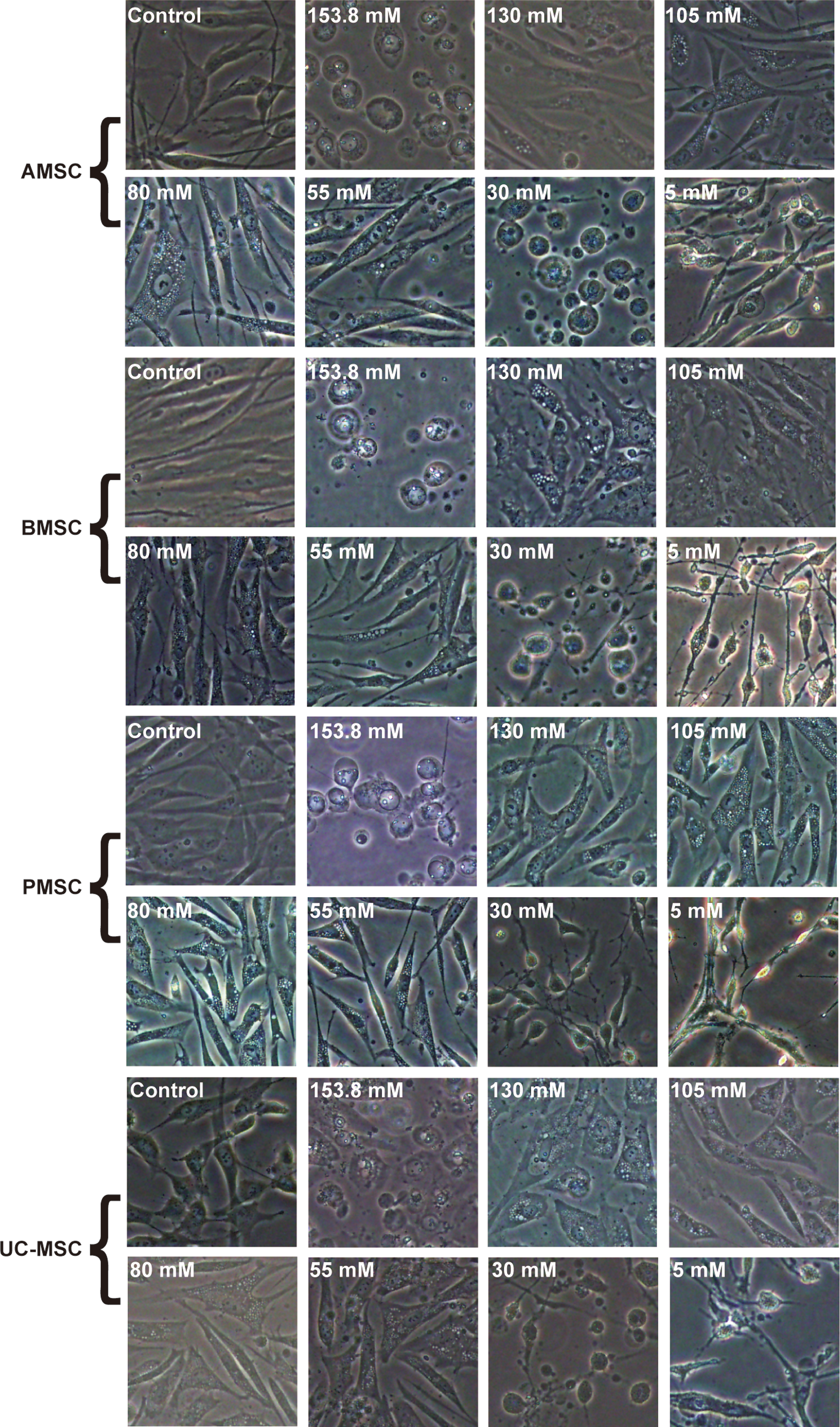
Morphology of MSCs from various tissues under light microscope after culture in artificial EL with altered K+ concentrations for 24 h. Scale bar = 100 μm. Color images available online at
Since MSCs originating from different tissues showed similar characteristics under light microscopy, we selected one of the cultured MSC types, AMSCs, to investigate fine morphological differences using TEM. TEM results indicated that apoptosis occurred in the 153.8, 30, and 5 mM K+ artificial EL groups after 24 h of cell culture with condensation and aggregation of nuclear chromatin, which did not occur in the control group (Fig. 7). In contrast, very few apoptotic cells were observed in the 130, 105, 80, and 55 mM K+ groups; instead, many glycogen particulates manifested within the cytoplasm of the MSCs (Fig. 8). These particulates were not observed in the control group, which is consistent with the ultrastructural features of glycogen particulates reported in previous studies [30,31].

Transmission electron microscopy images of AMSCs after culture in artificial EL with K+ concentrations of 153.8 mM, 30 mM, or 5 mM for 24 h. Scale bar = 0.5 μm. Red arrow indicates condensed and aggregated nuclear chromatin. Color images available online at
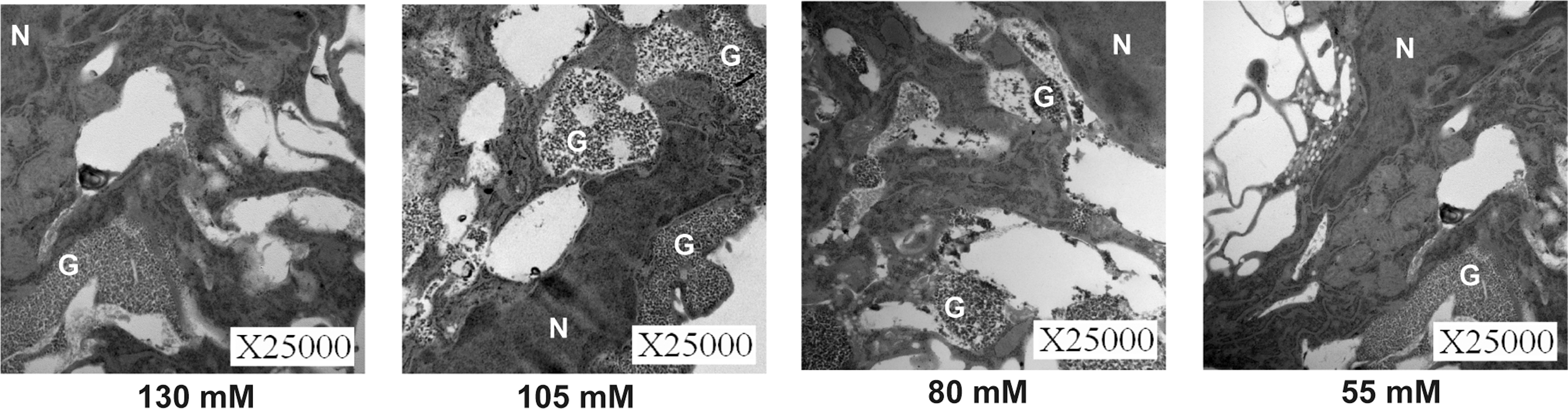
Transmission electron microscopy images of AMSCs after culture in artificial EL with K+ concentrations of 130 mM, 105 mM, 80 mM, or 55 mM for 24 h. Scale bar = 0.5 μm. G, glycogen particulates; N, nucleus.
No significant differences in IL-2, IL-6, IL-10, or TNF-α expression
To elucidate whether any of these results were related to inflammation-related reactions, we investigated the levels of IL-2, IL-6, IL-10, and TNF-α in the cell culture supernatants of the 153.8, 130, 80, and 30 mM K+ groups. Analysis revealed no significant differences across the artificial EL groups with various K+ concentrations, regardless of MSC type (P > 0.05; Supplementary Fig. S1; Supplementary Data are available online at
Discussion
Intraendolymphatic transplantation is a reasonable inner ear transplantation route, because the delivery efficiency (ie, introduce the transplanted cells to the auditory epithelium) is much better than intraperilymphatic route. However, high K+ concentration (153.8 mM) in EL is toxic to the transplanted cells, which makes a huge obstacle for the successful application of intraendolymphatic transplantation route [14,16].
Nevertheless, in previous studies, moderately high extracellular K+ concentration was reported to protect various types of cells from caspase activation, chromatin condensation, and DNA fragmentation [32,33]. Park et al. [16] found that most of the HeLa cells died within 6 h when cultured in artificial EL, while HeLa cell survival was greatly enhanced when cultured in a 50:50 mixture of artificial EL and PL. In that study, the K+ concentration of the 50:50 mixture of artificial EL and PL was 84 mM and the theoretical K+ equilibrium potential was −12 mV according to the Nernst equation [34]. With regard to HCs, Mazurek et al. [35] found that high K+ (70 mM) could protect HCs from ischemia-induced degeneration. In this study, we further confirmed this protective effect of K+ on cell survival and found that moderately high K+ concentrations of 55–130 mM in artificial EL can enhance the survival of MSCs in vitro.
In an in vivo study, Lee et al. [36] found that when conditioning the scala media (ie, flushing the EL with artificial PL), some transplanted embryonic stem cells could survive for as long as 7 days. During this time window (days 1–day 7), K+ levels gradually recovered (at least partially) following the conditioning procedure, and some of the transplanted cells, although few, could still survive. This could also partially support our result that a moderately high K+ concentration of the EL might not be as toxic as the normal artificial EL. Our study contributes to this line of research by providing evidence that MSCs can survive for at least 1 day with K+ concentrations as high as 55–130 mM in artificial EL. Since this was an in vitro experiment, the conditions are less complex than those in the in vivo environment of the cochlea; we reasoned that the duration of 24 h in vitro would be long to determine the most appropriate K+ concentrations for MSC survival, which provides confidence for follow-up studies in vivo.
Although the precise mechanism remains unclear, two lines of evidence might explain this protective effect of moderately high K+ in artificial EL. First, it is considered that the depolarization caused by high extracellular K+ concentrations can result in Ca2+ influx through Ca2+ channels, and this Ca2+ overload could in turn lead to the apoptosis of cells [37 –40]. However, the plasma membrane calcium ATPase (PMCA), which could pump out excess Ca2+, might contribute to the protective effect of extracellular high K+ and prevent cells from apoptosis [41,42]. Mazurek et al. [35] found that when blocking PMCA, the protective effect of high K+ was completely lost. Based on these results, we hypothesize that the protective effect of a moderately high K+ concentration on the survival of MSCs might contribute to the low concentration of Ca2+ (0.025 mM) in artificial EL, which would prevent the Ca2+ overloading effect. Second, the efflux of K+ can stimulate the cells to release the inflammatory factor IL-1β and then induce cell apoptosis [43 –45]. Conversely, under a high K+ concentration, the K+ efflux is attenuated, which might also be responsible for the enhanced cell survival.
In this study, after 24 h of treatment, although there was very little apoptosis and no increase in caspase-3 activity in 55–130 Mm K+ concentration groups, the 153.8, 30, and 5 mM K+ groups experienced significant increases in apoptosis rates and marked increases in caspase-3 activity. We infer from these results that the EL is toxic to MSCs if the K+ concentration is too high or too low. Our study and a previous study by Park et al. [16] both confirmed that the normal artificial EL (K+ concentration: 153.8 mM) is toxic to cells, but the underlying mechanisms remain unknown.
We speculate that one possible reason for the low survival of MSCs in the normal artificial EL is the overload of K+ ions. This speculation can be partially supported by studies on a K+ channel gene expressed in HCs, KCNQ4, which is responsible for the autosomal dominant nonsyndromic hearing loss 2, and mutation of this gene might result in a K+ overload in HCs, leading to cell degeneration [46]. In our study, the influx or efflux of K+ ions largely depends on the relationship between the resting potential and theoretical K+ equilibrium. Under a K+ concentration of 153.8 mM, according to the Nernst equation, the theoretical K+ equilibrium potential is +3.4 mV [34]. However, the resting membrane potential would be unpredictable because other conductances also have contributions.
On the one hand, when the resting potential is lower than the K+ equilibrium, the K+ ions might be overloaded, leading to the low survival of MSCs. On the other hand, when the resting potential is higher than the K+ equilibrium, the K+ ions might efflux and there would be other mechanisms leading to the low survival of MSCs, which require further studies to elucidate. In this study, when altering K+ concentration to 5 mM or 30 Mm, the survival of MSCs was also quite low. The reason for this phenomenon is currently unclear. Although cells could survive well in normal extracellular fluids with a low K+ concentration (4 mM) [34], this is quite different from the case of the low K+ concentration in the artificial EL, considering the difference in the other ion concentrations between normal extracellular fluids and artificial EL, such as Na+. Therefore, further investigations are needed to best understand the mechanisms contributing to the observed results.
The endocochlear potential (EP) in the vestibule is independent of that in the cochlear region, with the latter being much higher [47]. Because the presence of a high K+ concentration has been reported to be important by providing the major charge carrier for sensory transduction [48], the difference in EP between the vestibule and cochlea is mainly determined by the differences of K+ cycling between them. In the cochlea, the EP is generated by the stria vascularis (including the intermediate cells and strial marginal cells), whereas the EP in the vestibule is generated by the K+ secretion of vestibular dark cells. The strial marginal cells and vestibular dark cells both secrete K+ into the cochlear and vestibular EL through the K+ channel KCNQ1/KCNE1, which results in a membrane potential between 0 and +10 mV [49,50].
However, the EP is mainly determined by the K+ channel KCNJ10 in the intermediate cells of the stria vascularis, which results in the high EP in the cochlea (+80 mV) [51,52]. Therefore, high K+ is the major cation in the EL as well as the main charge carrier for sensory transduction, while the K+ channel in the intermediate cells of the cochlea is largely responsible for generating the EP. Furthermore, the K+ channels KCNQ1/KCNE1 and KCNJ10 show good promise as excellent pharmacologic targets in controlling the K+ concentration at the suitable range and extending the time window during inner ear replacement therapy, because the K+ levels were reported to gradually recover after flushing the EL with PL or the mixture of both, which might narrow down the time window for inducing the differentiation of transplanted stem cells and integrating them into the auditory epithelium.
Fortunately, our study shows that MSCs can endure K+ concentrations as high as 130 mM, making them good candidates for cochlear transplantation therapies. Nevertheless, this optimism must be tempered, as the proliferation of MSCs is inhibited in groups with high K+ concentrations compared with the control group. Previous studies have reported that elevated K+ concentrations can inhibit the proliferation of many types of cells [53 –57] and prevent cells from entering the S phase of the cell cycle, both of which are supported by our findings [58].
For the enhancement of stem cell proliferation in vitro, various small molecules and growth factors may be needed. In 2017, McLean et al. [59] reported that during the clonal expansion of Lgr5+ cells (ie, a kind of progenitor cell in the inner ear) in vitro, their proliferation was greatly enhanced by adding glycogen synthase kinase 3β inhibitor CHIR99021 (CHIR), basic fibroblast growth factor, EGF, and transforming growth factor β receptor (Alk5) inhibitor (616452). These studies can partially explain our results. In this study, moderately high potassium concentrations might have activated the metabolism of glycogen and enhanced the survival of MSCs in vitro. However, artificial EL was lacking of some growth factors that could promote cell proliferation. In the future, proliferation of MSCs might be enhanced by some methods, such as adding a series of growth factors into the EL in vitro.
To further understand the mechanisms underlying these phenomena, we analyzed the morphological changes of MSCs under various K+ concentrations. Several apoptotic MSCs were observed in the 153.8, 30, and 5 mM K+ artificial EL groups, as expected. In contrast, there were virtually no apoptotic cells in the 130, 105, 80, and 55 mM K+ groups; instead, many glycogen particulates and membrane vacuoles manifested within the cytoplasm of the MSCs. This is quite interesting, as a few studies have been conducted to explore the relationship between high potassium levels and glycogen formation. In 2013, Xu et al. [60] reported that glycogen could provide energy for the uptake of excess extracellular K+. In addition, DiNuzzo et al. [61] found that glycogen is necessary for the clearance of neuronally released K+ from the extracellular space. Therefore, in this study, when K+ concentrations were 55–130 mM in artificial EL, the glycogen accumulated, enabling MSCs to use glycogen as a source of fuel to survive and produce energy for the uptake of excess extracellular K+.
Meanwhile, we cannot ignore other components of the EL such as sodium and calcium that might also influence MSC survival despite their low concentrations. The influence of these components on MSC survival should also be addressed in subsequent research.
In summary, we discovered that different K+ concentrations in artificial EL could significantly influence the survival of MSCs in vitro. K+ concentrations of 55–130 mM in artificial EL can enhance the survival of MSCs in vitro. While MSCs exhibited reduced proliferation in all artificial EL groups, we proposed a potential method for enhancing MSC proliferation, which will be the focus of future studies. Our results suggest that MSCs are promising candidates for cochlear transplantation therapies. Our findings provide novel insights into methods of conditioning the EL for MSC transplantation and provide new hope for cochlear stem cell therapy.
Footnotes
Acknowledgments
This project was supported by grants from the National Key R&D Program of China (grant nos. 2016YFA0101000 and 2016YFA0101002) and the National Natural Science Foundation of China (grant nos. 81470698 and 81370466).
Author Disclosure Statement
The authors declare that they have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.