
Abstract
Cellular therapies are emerging as a novel treatment strategy for diabetes. Thus, the induction of endogenous islet regeneration in situ represents a feasible goal for diabetes therapy. Umbilical cord blood-derived hematopoietic progenitor cells (HPCs), isolated by high aldehyde dehydrogenase activity (ALDHhi), have previously been shown to reduce hyperglycemia after intrapancreatic (iPan) transplantation into streptozotocin (STZ)-treated nonobese diabetic (NOD)/severe combined immunodeficiency (SCID) mice. However, these cells are rare and require ex vivo expansion to reach clinically applicable numbers for human therapy. Therefore, we investigated whether BMS 493, an inverse retinoic acid receptor agonist, could prevent retinoic acid-induced differentiation and preserve islet regenerative functions during expansion. After 6-day expansion, BMS 493-treated cells showed a twofold increase in the number of ALDHhi cells available for transplantation compared with untreated controls. Newly expanded ALDHhi cells showed increased numbers of CD34 and CD133-positive cells, as well as a reduction in CD38 expression, a marker of hematopoietic cell differentiation. BMS 493-treated cells showed similar hematopoietic colony-forming capacity compared with untreated cells, with ALDHhi subpopulations producing more colonies than low aldehyde dehydrogenase activity subpopulations for expanded cells. To determine if the secreted proteins of these cells could augment the survival and/or proliferation of β-cells in vitro, conditioned media (CM) from cells expanded with or without BMS 493 was added to human islet cultures. The total number of proliferating β-cells was increased after 3- or 7-day culture with CM generated from BMS 493-treated cells. In contrast to freshly isolated ALDHhi cells, 6-day expansion with or without BMS 493 generated progeny that were unable to reduce hyperglycemia after iPan transplantation into STZ-treated NOD/SCID mice. Further strategies to reduce retinoic acid differentiation during HPC expansion is required to expand ALDHhi cells without the loss of islet regenerative functions.
Introduction
D
Bone marrow-derived cells have been shown to induce endogenous islet regeneration in several preclinical models [19 –22]. Cellular transplantation from the bone marrow can contain several different progenitor cell types, including multipotent stromal cells (MSCs), hematopoietic stem/progenitor cells, and endothelial progenitor cells, all which may contribute to islet regeneration. Our first publication on islet regeneration showed that transplantation of hematopoietic stem/progenitor cells from mouse bone marrow, isolated based on c-kit expression, reduced streptozotocin (STZ)-induced hyperglycemia in nonobese diabetic (NOD)/severe combined immunodeficiency (SCID) mice [11]. Upon postmortem analyses, donor cells did not coexpress insulin and suggested that glucose reduction occurred via endogenous pancreas regeneration in response to unknown secreted stimuli from donor cells.
In addition, it has previously been shown that human umbilical cord blood (UCB)-derived hematopoietic progenitor cells (HPCs), isolated based on high aldehyde dehydrogenase activity (ALDHhi), could engraft and repopulate irradiated NOD/SCID mice [23], as well as reduce hyperglycemia after intrapancreatic (iPan) transplantation into STZ-treated NOD/SCID mice [24]. Aldehyde dehydrogenase (ALDH) is a highly conserved marker of progenitor cells in multiple lineages and has many cytoprotective roles, including the detoxification of reactive oxygen species from long-lived progenitor cells [25 –27]. Both islet size and number were increased in mice transplanted with ALDHhi cells, and these mice demonstrated improved responses to intraperitoneal glucose tolerance tests compared with mice injected with low aldehyde dehydrogenase activity (ALDHlo) cells or phosphate-buffered saline (PBS) control. Collectively, these findings suggest that ALDH-expressing HPC demonstrate islet regenerative paracrine function after β-cell ablation with STZ.
UCB-derived HPCs isolated based on ALDHhi are extremely rare, comprising <0.5% of all UCB mononuclear cells (MNCs) [28]. Therefore, ex vivo expansion without the loss of islet regenerative capacity will be necessary to accrue applicable cell numbers for transplantation in humans. As ALDH is the rate-limiting enzyme in the production of intracellular retinoic acid (RA) and RA is known to induce differentiation in hematopoietic cells, we hypothesized that the small molecule BMS 493, an inverse agonist of the retinoic acid receptor (RAR)/retinoid X receptor (RXR) heterodimer, could be used during ex vivo expansion to prevent RA-induced differentiation [29]. The aims of this study were to characterize the phenotype and functional capacity of UCB-derived ALDHhi HPC after our 6-day expansion protocol with BMS 493 and to subsequently test expanded cell populations for islet regenerative properties using cultured human islets and after transplantation into STZ-treated NOD/SCID mice.
Materials and Methods
Human UCB collection and processing
Human UCB was collected with informed consent from the birthing center at the London Health Sciences Centre, Victoria Hospital in London, Ontario (REB No. 12934). Lineage depleted MNCs were stained with Aldefluor™ reagent (StemCell Technologies, Vancouver, BC), and ALDHhi cells were isolated using fluorescent activated cell sorting (FACS) as previously described [24,30,31].
HPC expansion ex vivo
ALDHhi UCB cells were cultured in X-VIVO™ 15 media supplemented with 10 ng/mL of thrombopoietin (TPO), FMS-related tyrosine kinase ligand (FLT-3L), and stem cell factor (SCF). After 3 days, media was changed and cells were left untreated or treated with 100 nM of BMS 493 (Sigma-Aldrich, St. Louis, MO).
Cell surface marker analyses
Multiparametric flow cytometry was used to assess ALDHhi frequency and cell surface marker coexpression after 6-day expansion and with or without 100 nM BMS 493. Cells were harvested and stained with Aldefluor, CD34-Brilliant Violet 421, CD38-phycoerythrobilin, CD117-PE-CF594, CD133-allophycocyanin, and 7-AAD (all antibodies BD PharMingen, San Diego, CA).
Methylcellulose assay
Methylcellulose was used to assess the hematopoietic colony-forming capacity (CFC) of expanded cells. After 6 days, cells were either left unsorted or resorted based on ALDH activity using the Aldefluor system by FACS. Cells were resuspended in Iscove's Modified Dulbecco's Medium (Gibco, Carlsbad, CA) and were plated at 500 cells per mL MethoCult™ H4434 (StemCell Technologies) in duplicate. After 14 days, hematopoietic colonies were enumerated under light microscopy based on morphology.
Islet culture with HPC-conditioned media
Cells expanded with or without BMS 493 were harvested on day 6 and purified based on viability. Live cells were replated at 2 × 105 cells per mL in RPMI 1640 media (Gibco) supplemented with 10 ng/mL SCF, TPO, and FLT-3L. After 48 h, conditioned media (CM) was collected and concentrated using 3 kDa Amicon Ultra-15 Centrifugal Filter Units (Millipore, Billerica, MA). Protein concentration was quantified using a NanoDrop™ spectrophotometer (A280). Next, human islets were obtained through the Integrated Islet Distribution Program (IIDP, Duarte, CA) and 200 IEQ were plated in 3 mL RPMI 1640 supplemented with 10 ng/mL SCF, TPO, and FLT-3 L or concentrated CM. EdU (500 nM) was added to cultures 24 h before cell harvesting. On day 3 or day 7, islets were dissociated into a single-cell suspension for assessment of survival and proliferation using multiparametric flow cytometry. Total cell number was enumerated manually by hemocytometer. ß-cell frequency was assessed using FluoZin-3 AM for live cells and insulin for permeabilized cells. Live cells were stained with Annexin V, and 7-AAD to assess β-cell survival, while fixed and permeabilized cells were stained with CD45-phycoerythrobilin and EdU-ClickIT Alexa Fluor 488 to assess proliferation.
Transplantation into STZ-treated NOD/SCID mice
Seven to 8-week-old NOD/SCID mice (Jackson Labs, Bar Harbor, ME) received intraperitoneal injections of 35 mg/(kg·d) of STZ (Sigma-Aldrich) for five consecutive days as previously described [11,24,30,31]. All animal procedures were approved by the Animal Care Committee at Western University Canada (AUP 2015-033). On day 10, hyperglycemic mice (>15 mmol/L) were sublethally irradiated (300 cGy) and transplanted by direct iPan injection [24]. For cellular transplantation, mice received 2 × 105 bulk, ALDHhi, or ALDHlo cells that were untreated or treated with BMS 493 as described above or PBS in 20 μL volume. For CM injections, mice received 20 μL of CM generated from cells expanded with or without BMS 493 as described above or RPMI 1640 supplemented with 10 ng/mL SCF, TPO, and FLT-3L as a control. Nonfasted blood glucose concentration was measured on days 0, 7, 10, 14, 17, 21, 27, 35, and 42. Mice were intraperitoneally injected with 100 μL of 2.5 mg/mL EdU 24 h before euthanasia, and mouse pancreata were harvested and frozen in OCT for immunohistochemistry.
Immunohistochemistry
Pancreata were cryosectioned into 10 μm sections separated by no <100 μm between sections, and three sections were placed on the same slide. Sections were stained for insulin, human leukocyte antigen (HLA), and CD31 as previously described [30]. For quantification of immunohistochemistry, 4 images were taken per section (12 images per slide) at 10 × magnification for insulin staining or 20 × magnification for HLA and CD31 staining.
Statistics
Data are presented as mean ± standard error of the mean. ALDH activity and primitive cell surface marker retention with or without BMS 493 treatment was analyzed by two-tailed Student's t-tests (*P < 0.05). Titration of BMS 493, area under the curve (AUC) for in vivo data, and all histological data were analyzed using a one-way analysis of variance (ANOVA). Human islet culture data were analyzed using a two-way ANOVA with Dunnett's multiple comparisons test. Forty-two-day blood glucose curves were analyzed using a two-way ANOVA with Tukey's multiple comparisons test.
Results
Treatment with BMS 493 increased the frequency of ALDHhi cells after expansion
ALDHhi cells were purified from UCB using FACS (Fig. 1A–C). Various concentrations and timing of BMS 493-treatment were initially tested to optimize the retention of ALDHhi cells after expansion (Supplementary Fig. S1; Supplementary Data are available online at

Treatment with BMS 493 increased ALDHhi cell frequency and total ALDHhi cell number after 6-day expansion.
BMS 493-treated cells retained primitive cell surface marker expression
Multiparametric flow cytometry was used to assess the expression of primitive (CD34 and CD133) and mature (CD38) hematopoietic cell surface markers previously characterized on UCB-derived hematopoietic cells [32]. Representative flow cytometry dot plots are shown for untreated (Fig. 2A–C) and BMS 493-treated cells after 6-day expansion (Fig. 2D–F). Cells treated with BMS 493 showed no difference in the frequency of CD34 and CD133 expression compared with untreated controls (Fig. 2G, H). In contrast, treatment of cells with BMS 493 resulted in a significant decrease in the frequency of ALDHhi cells that acquired CD38 coexpression (P < 0.01, Fig. 2I). Due to the significant increase in total ALDHhi cell number after BMS 493 treatment compared with untreated conditions at day 6, BMS 493 treatment increased the total number of ALDHhi cells coexpressing CD34 and CD133, 1.89- and 1.86-fold, respectively (P < 0.01, Fig. 2J, K). Similarly, BMS 493 treatment resulted in a 48.5-fold decrease in the total number of ALDHhi CD38+ cells compared with untreated controls (P < 0.05, Fig. 2L). Since bone marrow cells isolated from mice based on the expression of c-kit (CD117) have been previously shown to play a role in pancreatic regeneration [11], BMS 493-treated cells were also assessed for c-kit expression after 6-day expansion. There was no significant difference in the frequency or total number of cells with high ALDH activity and c-kit expression compared with untreated conditions (Supplementary Fig. S2). Collectively, expanded BMS 493-treated cells enhanced retention of ALDHhi cells with primitive cell surface marker (CD34 and CD133) coexpression and reduced the expression of the mature hematopoietic cell surface marker CD38 compared with untreated cells.
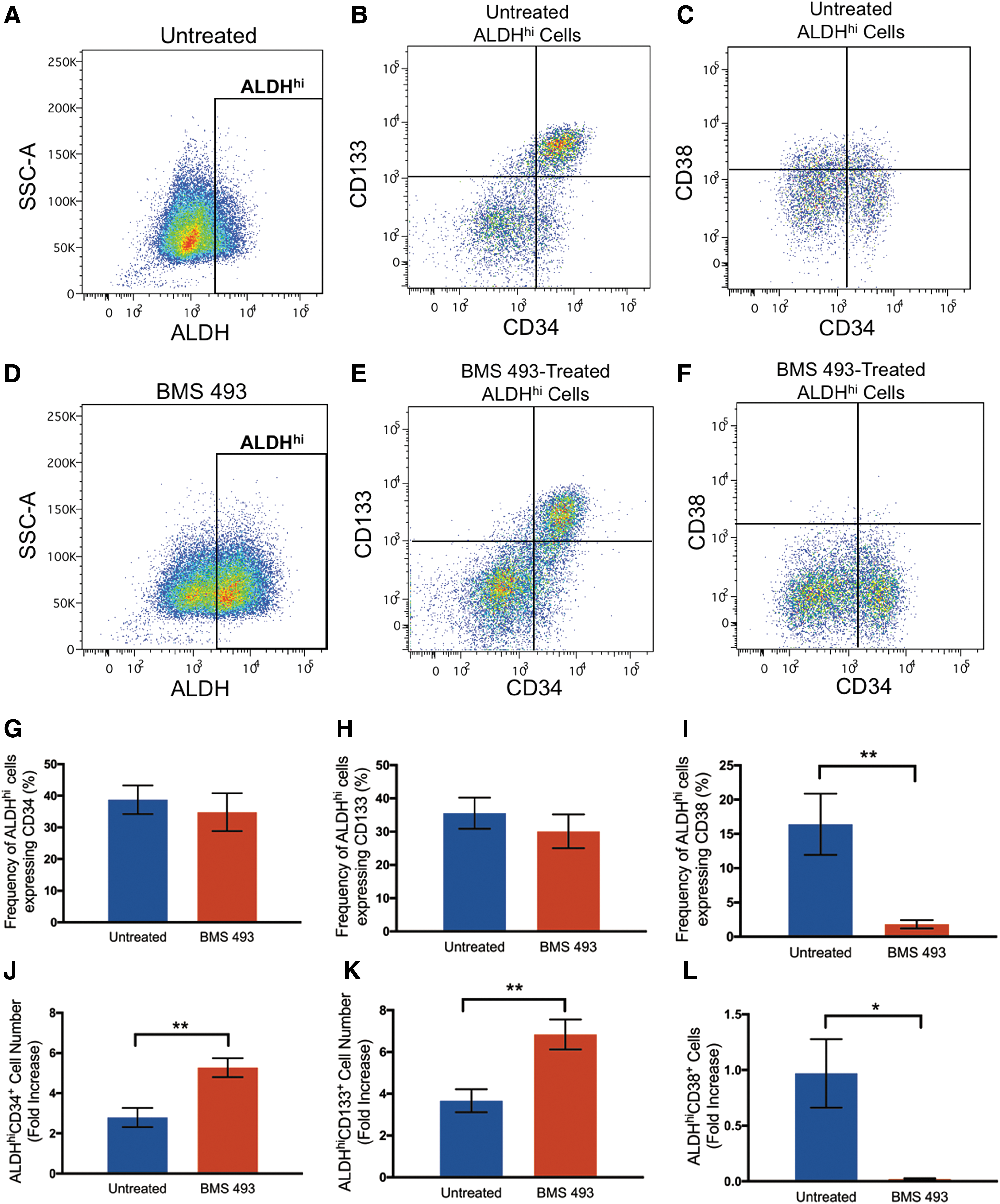
Treatment with BMS 493 increased total ALDHhi cells with primitive (CD34+ and CD133+) phenotypes and decreased total ALDHhi cells with differentiated (CD38+) phenotype. Representative flow cytometry dot plots showing ALDH activity, and CD34, CD133, or CD38, coexpression on cells expanded for 6 days under:
Hematopoietic CFC was retained after BMS 493 treatment
To test the progenitor function of cells expanded with or without the small molecule BMS 493, hematopoietic colony-forming assays were performed using semisolid methylcellulose medium. After 14 days, there was no significant difference in the total number of hematopoietic colonies formed (Fig. 3A), or in the colony subtypes formed by BMS 493-treated cells compared with untreated controls (Fig. 3B). After 6-day expansion, cells were resorted based on ALDH activity and plated in the methylcellulose medium. The ALDHhi subpopulation produced significantly more colony-forming units (CFUs) per 100 cells compared with the ALDHlo subpopulation in both BMS 493-treated (P < 0.05, 9.80% ± 1.67 and 4.55% ± 0.96) and untreated controls (P < 0.001, 12.30% ± 0.90 and 5.22% ± 0.54) (Fig. 3C). There were no differences in the frequency of colony subtypes between resorted populations of treated and untreated cells (Fig. 3D). Therefore, treatment with BMS 493 during expansion increased the total number of ALDHhi cells available for transplantation without the loss of hematopoietic colony-forming progenitor capacity.
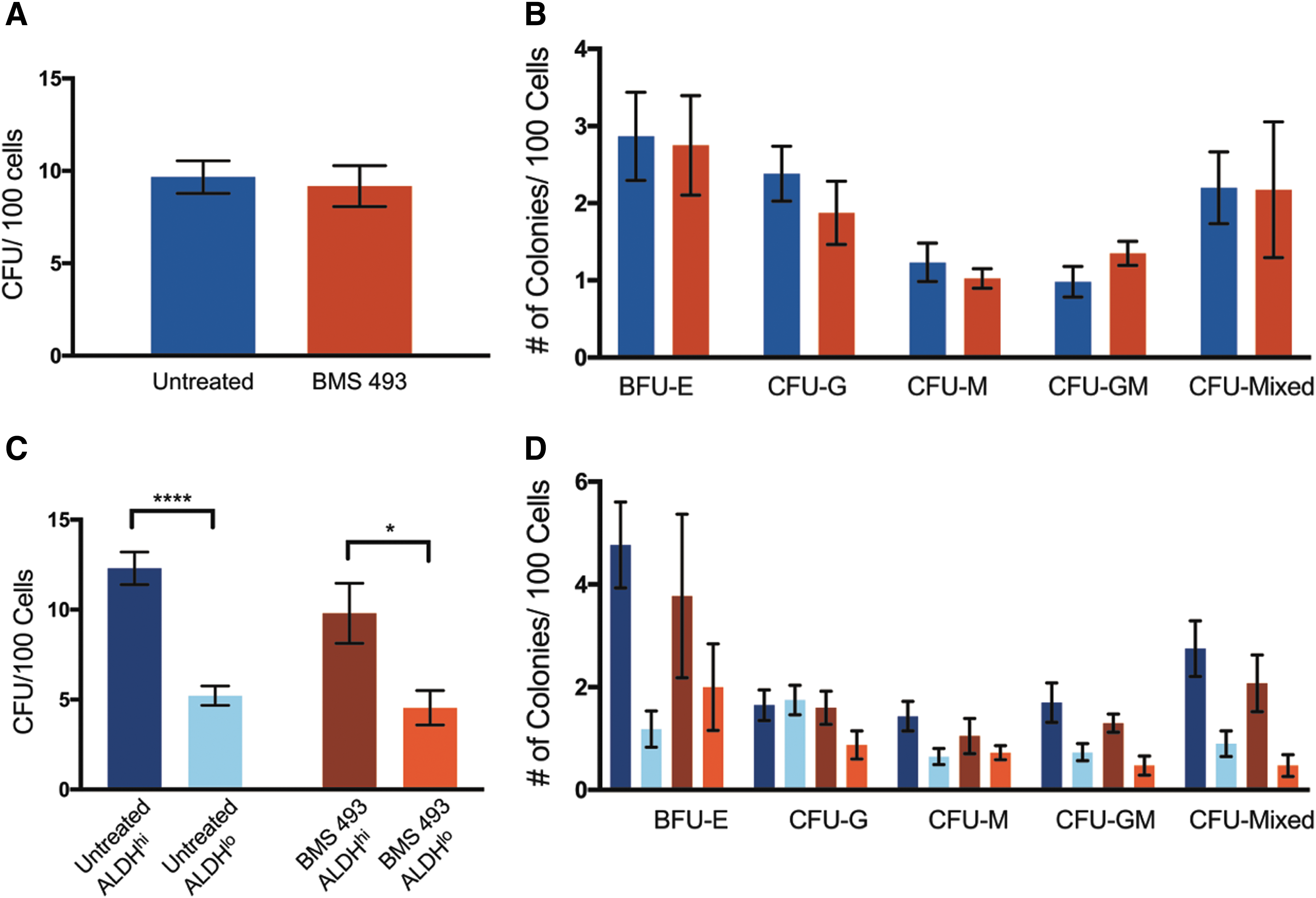
BMS 493-treated cells retained hematopoietic CFC. Compared with untreated controls (N = 4, n = 2), bulk BMS 493-treated cells (N = 6, n = 2) showed
CM from BMS 493-treated cells increased β-cell proliferation in vitro
Human pancreatic islets were cultured for 3 or 7 days in vitro with concentrated CM generated from cells expanded with or without BMS 493. There were no changes in islet morphology observed after culture with HPC CM (Supplementary Fig. S3A–C). Subsequently, islets were dissociated and assessed for β-cell survival using multiparametric flow cytometry (Fig. 4A–C). Zinc markers such as FluoZin-3 AM have previously been used to label viable β-cells due to high levels of secretory granules containing insulin and zinc within exocytosis machinery [33]. There was no difference in the total number or frequency of FluoZin-3+ β-cells at day 3 or day 7 after islets were cultured with media control (RPMI 1640) or CM generated from untreated or BMS 493-treated cells (Supplementary Fig. S3D, E). In addition to FluoZin-3 AM, 7-AAD and Annexin V were used to assess β-cell viability and apoptosis. After treatment with CM generated from BMS 493-treated cells, there was no difference in the frequency or total number of live (FluoZin-3+7AAD−Annexin−), apoptotic (FluoZin-3+7AAD−Annexin+), dead (FluoZin-3+7AAD+Annexin+), or necrotic (FluoZin-3+7AAD+Annexin−) β-cells compared with media controls at both days 3 and 7 (Fig. 4D–G). Therefore, culture of human islets with CM generated from expanded ALDHhi cells with or without BMS 493-treatment did not impact β-cell survival in vitro.
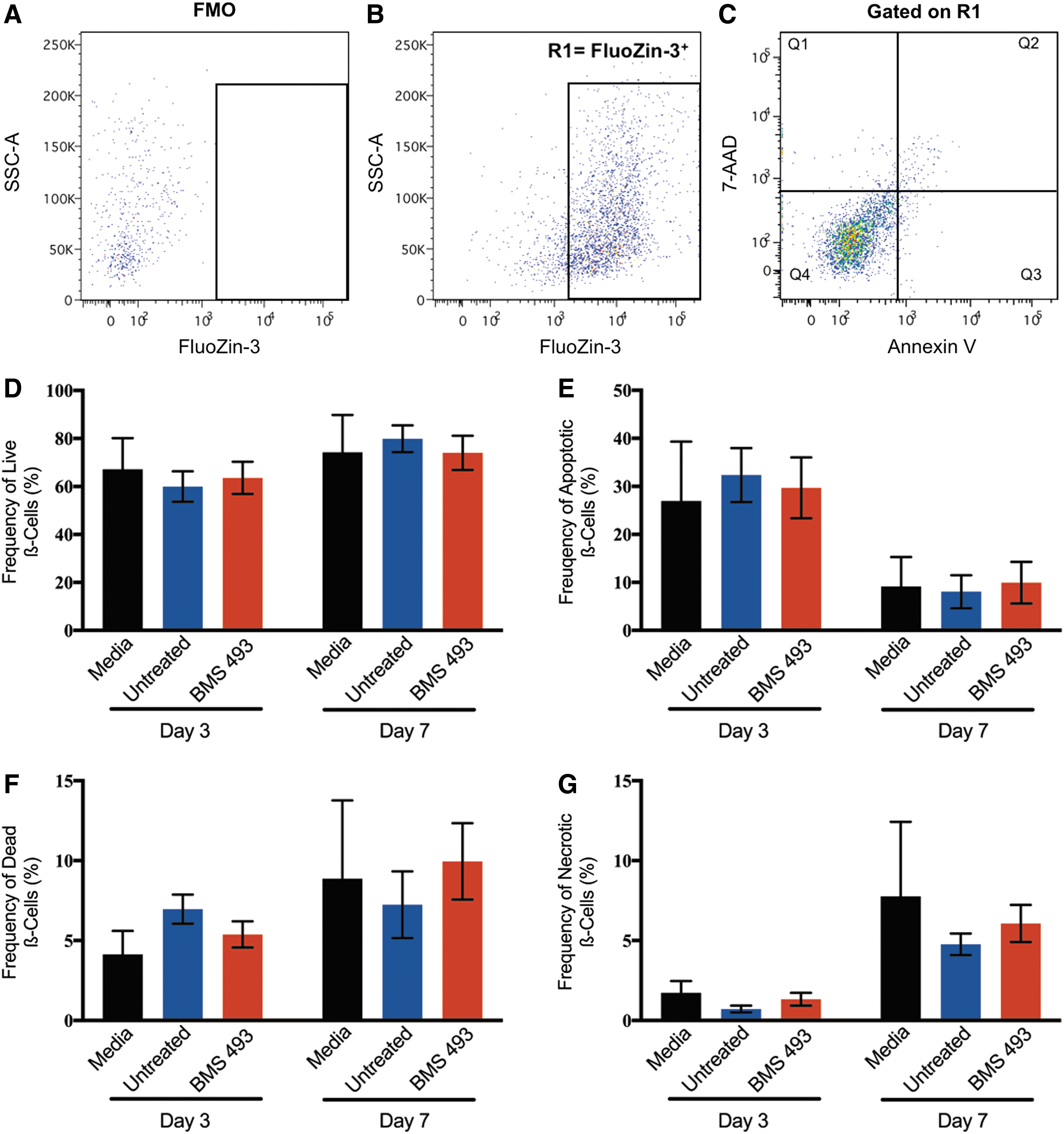
Human islets cultured with CM generated from BMS 493-treated hematopoietic cells showed equivalent ß-cell survival. Representative flow cytometry dot plots showing
To assess the potential effects of human β-cell proliferation in vitro, islets were dissociated at day 3 or day 7, fixed and permeabilized (Fig. 5A–C), and multiparametric flow cytometry was performed to measure intracellular insulin and incorporation of the thymine analog, EdU. While there were no significant differences in the overall frequency of proliferating β-cells (CD45−Insulin+EdU+) at day 3 or day 7 (Fig. 5D), the total number of proliferating β-cells was significantly increased at both day 3 (2.59-fold increase) and day 7 (2.98-fold increase) when islets were treated with CM generated from BMS 493-treated cells compared with islets treated with media alone (P < 0.05, Fig. 5E). Thus, CM generated from a 6-day expanded ALDHhi cells treated with BMS 493 significantly increased human β-cell proliferation in vitro.
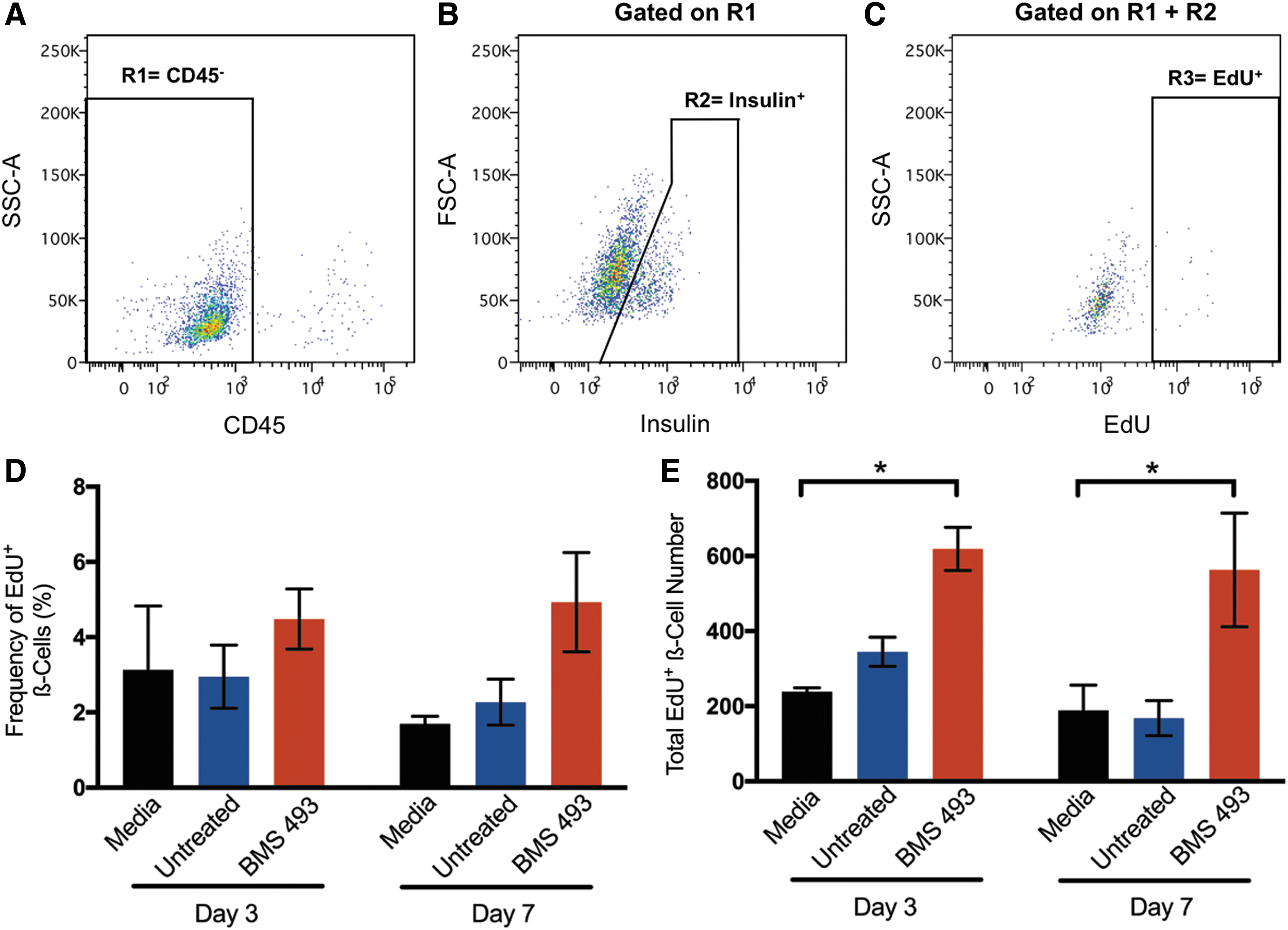
Human islets cultured with CM generated from BMS 493-treated hematopoietic cells contained an increased number of proliferating ß-cells. Representative flow cytometry plots showing
CM from expanded cells did not reduce hyperglycemia in STZ-treated NOD/SCID mice
Following the augmented human β-cell proliferation in vitro after treatment with CM generated from cells expanded with BMS 493, we next tested whether direct iPan injection of concentrated CM into STZ-treated NOD/SCID mice (Fig. 6A) could increase murine β-cell mass, leading to a reduction in hyperglycemia in vivo. Although CM generated from BMS 493-treated and untreated cells were matched for total protein concentration, injection of CM from expanded cells did not reduce blood glucose or AUC compared with mice injected with media controls (Fig. 6B, C). Postmortem, there was no significant difference in islet size, islet number, or total β-cell mass when mice injected with CM were compared with media-injected control mice (Supplementary Fig. S4, Fig. 6D–F). Collectively, these data indicate that direct iPan injection of concentrated CM from expanded ALDHhi cells was unable to reduce hyperglycemia in vivo and showed no effect on the recovery β-cell mass postinjection.
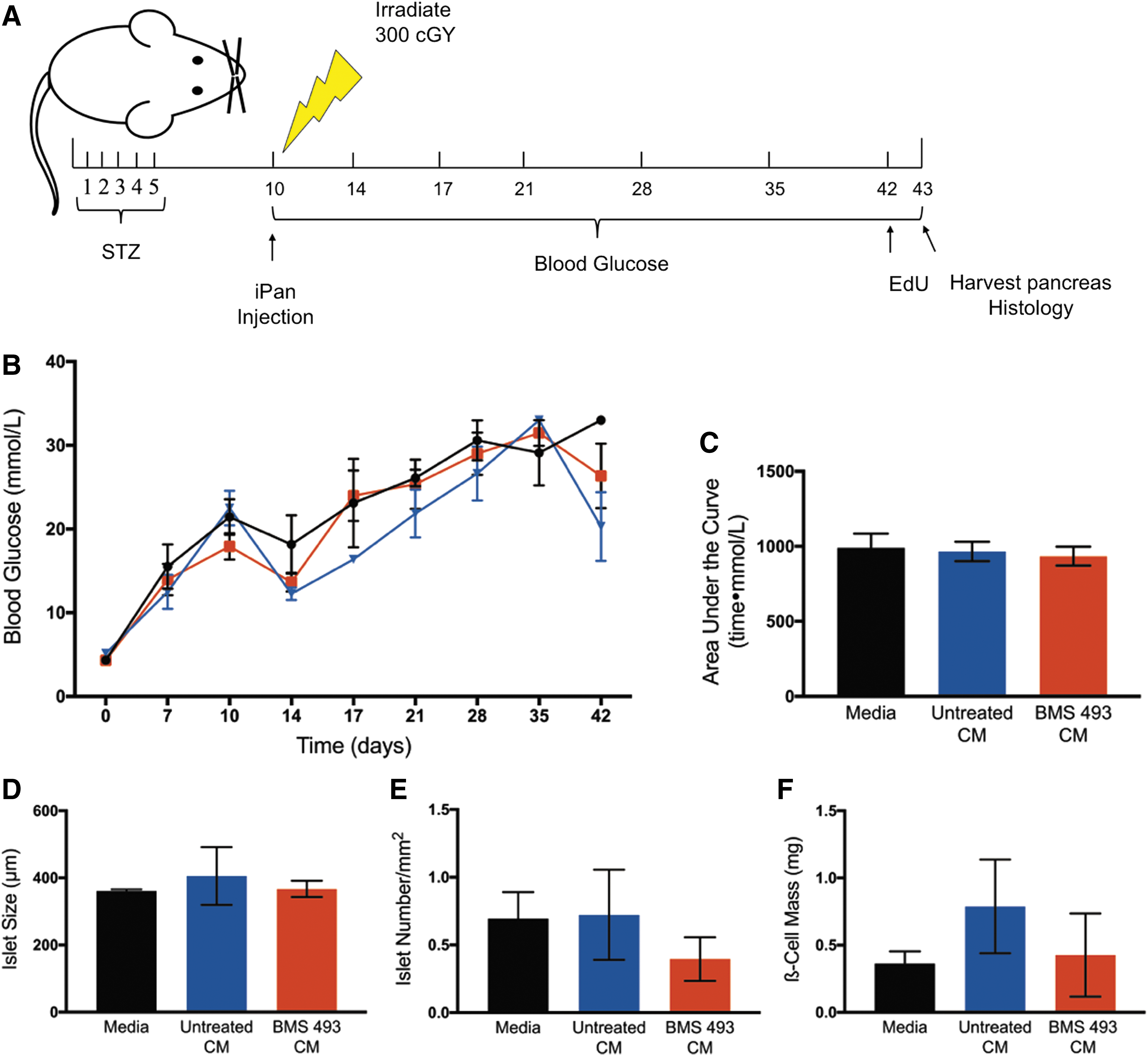
CM from BMS 493-treated cells did not reduce hyperglycemia after iPan injection into STZ-treated NOD/SCID mice.
Transplantation of expanded cells did not reduce hyperglycemia in STZ-treated NOD/SCID mice
Previously, we have shown that fresh UCB-derived cells purified for high ALDH activity have the capacity to reduce STZ-induced hyperglycemia in NOD/SCID mice [24]. Therefore, we tested the ability of 6-day expanded cells to reduce hyperglycemia after iPan transplantation. After 6-day expansion, iPan transplantation of 2 × 105 untreated and BMS 493-treated cells did not reduce hyperglycemia compared with PBS-injected controls (Fig. 7A). AUC over 42 days similarly showed no significant difference between either BMS 493-treated or untreated expanded cells compared with PBS controls (Fig. 7B). Fresh UCB ALDHhi cells were concurrently transplanted as a positive control. As previously published, there was a significant reduction in AUC after transplantation of fresh ALDHhi cells compared with PBS controls (P < 0.01, Fig. 7B). Taken together, these data indicated that despite BMS 493 treatment, expansion of ALDHhi cells for 6 days ex vivo significantly reduced islet regenerative capacity after iPan-injection.
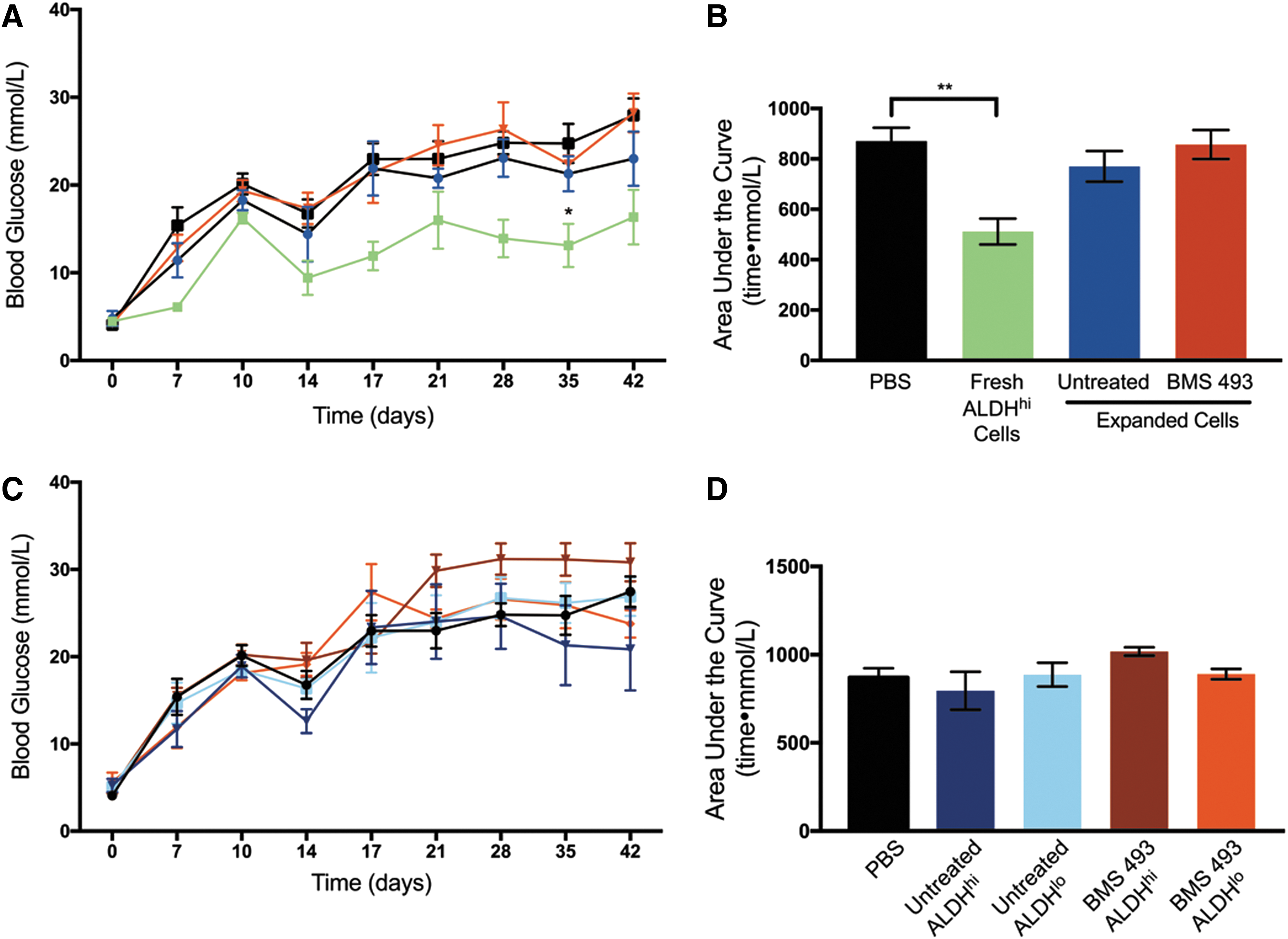
Expanded cell transplantation did not reduce hyperglycemia after iPan injection into STZ-treated NOD/SCID mice.
We have also shown that expanded cells reselected for high ALDH activity could reduce hyperglycemia in STZ-treated NOD/SCID mice after intravenous (IV) transplantation [30]. To assess whether the ALDHhi subpopulation of cells expanded with BMS 493 could impact hyperglycemia after iPan transplantation, 6-day expanded cell populations were repurified based on ALDH activity and iPan injected into STZ-treated NOD/SCID mice. Unfortunately, there was no significant change in hyperglycemia over 42 days after transplantation of BMS 493-treated ALDHhi, BMS 493-treated ALDHlo, untreated ALDHhi, or untreated ALDHlo cells compared with PBS controls (Fig. 7C, D). Collectively, these data suggested that 6-day culture ex vivo leads to the loss of islet regenerative capacity, with or without BMS-493 treatment.
Immunohistochemistry was performed to assess islet size, number, and total β-cell mass after 42 days. Mice transplanted with BMS 493-treated cells showed no difference in islet size, number, or total β-cell mass compared with untreated cells and PBS controls (Fig. 8G–L). Pancreas sections were also stained with HLA (A, B, C) and platelet endothelial cell adhesion molecule (PECAM, CD31) to assess human cell engraftment and islet vascularity, respectively. After 42 days, no human cells were detected in the pancreas of mice transplanted with BMS 493-treated or untreated cells (Supplementary Fig. S5). Finally, there were no differences in the total number of CD31+ cells per islet area in the pancreas of mice transplanted with BMS 493-treated cells and PBS-injected controls (Supplementary Fig. S6). Similar to their respective unsorted populations, mice transplanted with BMS 493-treated or untreated cells reselected based on ALDH activity showed no differences in islet size, islet number, total β-cell mass, or islet vascularity as measured by CD31+ cells per islet area compared with PBS-injected control mice.
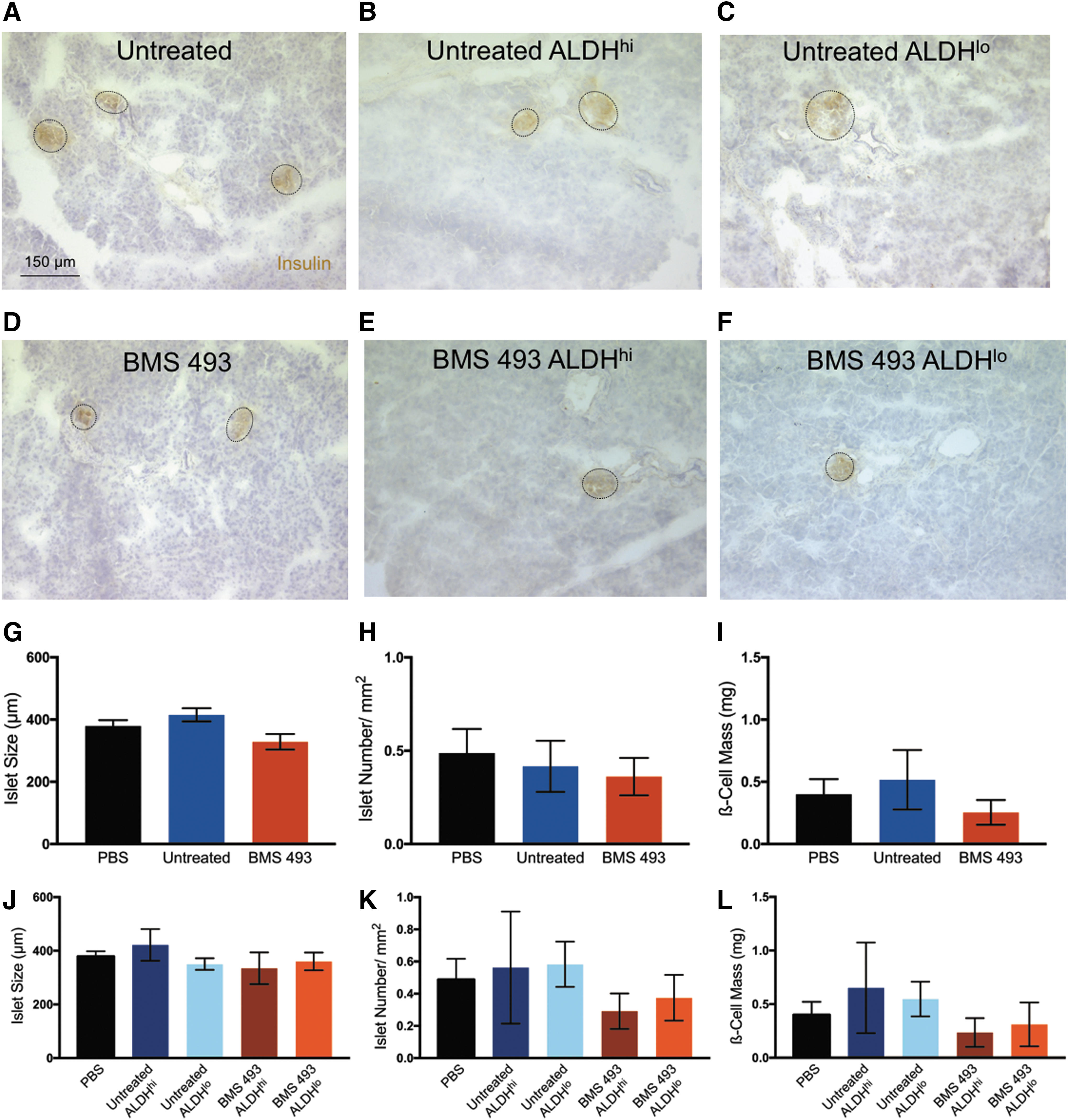
Expanded cell transplantation did not alter islet size, islet number, or ß-cell mass after iPan transplantation into STZ-treated NOD/SCID mice.
Discussion
Previously, we identified a subset of UCB-derived cells with high ALDH activity that stimulate endogenous islet repair after iPan transplantation into NOD/SCID mice with STZ-induced hyperglycemia [24]. Unfortunately, due to the efficient production of RA, ALDHhi cells differentiate quickly when cultured, rapidly losing primitive progenitor cell phenotype after 6-day expansion [30]. Chute et al. [34,35] first suggested that inhibition of retinoid signaling during HPC expansion may prevent RA-induced differentiation down the hematopoietic hierarchy. By using N,N-diethylaminobenzaldehyde (DEAB), a direct-inhibitor of ALDH1-activity during short-term culture, Chute et al. [34] showed increased numbers of CD34+CD38−Lin− cells after 7-day expansion ex vivo. Expanded cell progeny had increased number of SCID repopulating cells after IV transplantation into irradiated NOD/SCID mice, and decreased expression of cEBPɛ, a downstream target of RAR-signaling. It has also recently been shown that expansion of HPC ex vivo with DEAB in culture can increase the number of ALDHhi cells after 9 days that retain their regenerative properties after transplantation into a murine model of femoral artery ligation [36]. Alternate molecules such as BMS 493 block RA-induced differentiation further downstream in the canonical RA pathway are needed to develop cell therapy approaches to treat nonhematopoietic disorders such as diabetes and its cardiovascular comorbidities.
When treated with BMS 493, UCB-derived ALDHhi cells expanded for 6 days in culture showed a twofold increase (31.16 ± 3.09%) in the frequency of cells that retained high ALDH activity compared with untreated controls. Despite the preservation of primitive HPC phenotypes (ALDHhi/CD34+/CD133+ cells), total cell expansion over 6 days was not inhibited by BMS 493 treatment. Therefore, treatment with BMS 493 for 6 days resulted in an 8.3 ± 1.3-fold increase in the total number of ALDHhi cells available for transplantation, without the loss of primitive cell surface marker coexpression. ALDH activity represents the rate-limiting step in the conversion of vitamin A to RA, a powerful morphogen and differentiation factor [37,38]. In contrast to expansion with DEAB, BMS 493 acts further downstream on the RAR/RXR heterodimer required for RA-pathway-targeted gene transcription. Since BMS 493 does not directly affect the production of RA in the cell, levels of RA are likely unaffected by treatment. Measurement of downstream targets of RA signaling pathways may help elucidate how manipulating the actions of RA during ex vivo culture affects the differentiation and regenerative capacity of HPC.
In the context of cell therapies for diabetes, several preclinical studies have used hematopoietic MNCs or MSCs, primarily from human bone marrow (BM), to induce endogenous islet regeneration in situ. Our laboratory has previously shown that direct iPan delivery of uncultured, UCB-derived ALDHhi cells could reduce hyperglycemia and increase serum insulin levels in STZ-treated NOD/SCID mice [24,31]. Furthermore, mice showed improved functional recovery after a glucose challenge (intraperitoneal glucose tolerance test) compared with mice injected with an equivalent dose of ALDHlo cells or PBS controls. Importantly, iPan transplantation of UCB-derived ALDHhi cells expanded in culture lost the ability to reduce hyperglycemia in the same mouse model, with or without BMS 493-treatment. Although ALDHhi HPC can be expanded efficiently ex vivo, secretory and prosurvival functions may be altered and compromised by culture-induced differentiation, and transplantation at higher cell doses may be required to achieve a regenerative effect after injection.
The endocrine pancreas is an extremely well vascularized organ. Islets have their own capillary networks that are glomerular-like and can take up as much as 10% of the total islet volume [39,40]. In addition, islets are ∼10 times more vascular than surrounding exocrine tissue. Replacing lost β-cell mass in type 1 diabetes mellitus may be the primary goal of new cellular therapies; however, ensuring proper vascularization of β-cells is imperative in achieving functional recovery and glucose homeostasis. While little is known regarding the mechanisms of islet regeneration after expanded ALDHhi cell transplantation, there is emerging evidence surrounding the proangiogenic paracrine function of expanded ALDHhi HPC in models of vascular injury. It has previously been shown that UCB-derived ALDHhi cells can aid in the revascularization and perfusion of injured muscle tissue after femoral artery ligation in NOD/SCID mice, an important finding given the prevalence of peripheral artery disease as a complication of diabetes [28,41]. However, there were no differences in the frequency of CD31+ cells found in islets after iPan transplantation of HPC expanded with BMS 493, and subsequently, β-cell mass was not increased. Conversely, islet vascularization may not have been induced since there was insufficient insulin production after STZ treatment to warrant new vasculature for transport. Although we did not observe a reduction in hyperglycemia after iPan transplantation of expanded HPC alone, expansion of ALDHhi HPC may be important to accelerate islet revascularization after an alternate regenerative stimulus is delivered, particularly in islet replacement strategies that increase β-cell mass requiring efficient vascularization and anastomoses.
CM collected from BMS 493-treated HPC increased the number of proliferating β-cells in vitro. This finding supported previously published data showing a reduction in hyperglycemia after combinatorial transplantation of bone marrow-derived MSC and UCB-derived HPC into STZ-treated NOD/SCID mice [31]. This suggested that human cell-secreted effectors were highly conserved and had the capacity to impact murine islet function after transplantation. Furthermore, it was suggested that MSC played a greater role in neo-islet formation, while HPC reduced hyperglycemia by inducing β-cell proliferation. Recently published work has suggested that CM from MSC activated for the Wnt pathway augmented the survival and proliferation of human β-cells in culture [42]. This leads us to further study the effects of CM on islet regeneration after iPan transplantation of concentrated CM into STZ-treated NOD/SCID mice. Transplantation of CM was predicted to be a potent stimulus because CM can be concentrated using filtration units and transplanted directly into the mouse pancreas using low volumes [42,43]. In contrast to our in vitro human islet cultures that showed an increase in β-cell proliferation, these effects may not occur in vivo due to the inability of murine islets to receive signals from human cell secreted effectors. While it is believed that these effectors are highly conserved between species from previously published data using human cells in murine models, further in vitro experiments are necessary to determine if effectors from human HPC CM stimulate murine islet cells to proliferate comparable to effects shown with human islet cultures [31]. In addition, the lack of effect in vivo could also be attributed to extensive β-cell damage as a result of STZ-treatment and the different pancreas microenvironment.
In summary, BMS 493, an inverse RAR agonist, was used to inhibit RA-induced differentiation of UCB-derived ALDHhi cells in culture. After 6 days, BMS 493-treated ALDHhi cells showed a 2-fold increase in ALDHhi cells without the loss of primitive cell surface marker expression or CFC. Treating human islets in culture with CM generated from BMS 493-treated ALDHhi cells showed increased β-cell proliferation. iPan transplantation of cell progeny after expansion of ALDHhi cells with or without BMS 493 showed no reduction of hyperglycemia in STZ-treated NOD/SCID mice. Thus, UCB-derived ALDHhi cells effectively lost islet regenerative capacity during ex vivo expansion. Future studies are necessary to investigate the proangiogenic role of HPC expanded with small molecules and their use for other applications such as peripheral artery disease or in combination with other cell types for the development of novel cell therapies in the treatment of diabetes mellitus.
Footnotes
Acknowledgments
This work was supported by the Juvenile Diabetes Research Foundation (JDRF-2-SRA-2015-60-Q-R). We acknowledge Kristin Chadwick for cell sorting and Madeline Harvey for animal care.
Portions of the data in this article were presented at the Till and McCulloch Meetings on October 24th, 2016 and November 7th, 2017.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.