
Abstract
The functional disorders caused by central nervous system (CNS) diseases, such as ischemic stroke, are clinically incurable and current treatments have limited effects. Previous studies suggested that cell-based therapy using mesenchymal stem cells (MSCs) exerts therapeutic effects for ischemic stroke. In addition, the characteristics of MSCs may depend on their sources. Among the derived tissues of MSCs, we have focused on cranial bones originating from the neural crest. We previously demonstrated that the neurogenic potential of human cranial bone-derived MSCs (cMSCs) was higher than that of human iliac bone-derived MSCs. Therefore, we presumed that cMSCs have a higher therapeutic potential for CNS diseases. However, the therapeutic effects of cMSCs have not yet been elucidated in detail. In the present study, we aimed to demonstrate the therapeutic effects of transplantation with rat cranial bone-derived MSCs (rcMSCs) in ischemic stroke model rats. The mRNA expression of brain-derived neurotrophic factor and nerve growth factor was significantly stronger in rcMSCs than in rat bone marrow-derived MSCs (rbMSCs). Ischemic stroke model rats in the rcMSC transplantation group showed better functional recovery than those in the no transplantation and rbMSC transplantation groups. Furthermore, in the in vitro study, the conditioned medium of rcMSCs significantly suppressed the death of neuroblastoma × glioma hybrid cells (NG108-15) exposed to oxidative and inflammatory stresses. These results suggest that cMSCs have potential as a candidate cell-based therapy for CNS diseases.
Introduction
C
Previous studies reported that transplantation with MSCs may result in the repair of damaged brain tissue and functional recovery in stroke models [1 –12]. Moreover, MSCs secrete neurotrophic factors, exhibit migration, and promote endogenous neurogenesis in stroke models [13 –16]. MSCs have been isolated from various tissues, including adipose tissue and bone marrow. Some studies revealed that the characteristics of MSCs from different sources may vary [17 –20]. Therefore, the effectiveness of the transplantation of MSCs may depend on the source of MSCs, and source selection is important for cell-based therapy using MSCs.
Among sources of MSCs, we focused on cranial bones, which originate from the neural crest. Some of the bones in the cranial vault and face are derived from the cranial neural crest. On the contrary, the bones of the limbs, iliac bone, and vertebrae are derived from the mesodermal germ layer [21]. MSCs are normally isolated from the long bones of rodents and the iliac bone of humans [22 –25]. Sakai et al. [26] revealed that dental pulp-derived stem cells are derived from cranial neural crest cells and exhibit prominent neurogenic activity, which has not been observed for other adult somatic stem cells. We previously succeeded in establishing MSCs from human cranial bones and demonstrated that they have more cranial neural crest markers and show a tendency to differentiate into neuron-like cells [27]. Based on these findings, we presumed that cranial bone-derived MSCs (cMSCs) have a high therapeutic potential for CNS diseases, including ischemic stroke. However, the therapeutic effects of cMSCs have not yet been elucidated in detail. In the present study, we aimed to demonstrate the characteristics of rat cranial bone-derived MSCs (rcMSCs) and their effectiveness over rat bone marrow-derived MSCs (rbMSCs) for function recovery after transplantation into ischemic stroke model rats.
Materials and Methods
Ethics statement
All experiments involving the use of animals were in compliance with the National Institutes of Health Guidelines. All study protocols were approved by the Animal Testing Committee Guidelines at Hiroshima University.
Isolation and culture of rbMSCs and rcMSCs
We isolated rbMSCs from the femurs and tibias of six male Sprague–Dawley (SD) rats (4–5 weeks old). Bone marrow was seeded on culture dishes (Sumitomo Bakelite Co., Tokyo, Japan) using growth medium: Dulbecco's modified Eagle medium low-glucose (DMEM-L; Sigma-Aldrich Co., St. Louis, MO) with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific HyClone, South Logan, UT), penicillin (100 U/mL), and streptomycin (100 μg/mL; both from Sigma-Aldrich). Dishes were incubated at 37°C with 5% CO2. Culture media were changed every 3 days to eliminate nonadherent cells. Cells that adhered to the bottom of the culture dish were used as rbMSCs and were passaged several times.
We isolated rcMSCs from the rat cranial bone of six male SD rats (4–5 weeks old). Periosteum, muscle, dura mater, and olfactory nerves were completely removed from the cranial bones. The cranial bone samples were seeded on culture dishes containing growth medium. Samples were maintained at 37°C with 5% CO2. We detected adherent cells 7–14 days after seeding of cranial bone samples. Therefore, the culture media were first changed 14 days after seeding to eliminate floating debris and bone particles. Adherent cells were used as rcMSCs and were passaged several times.
Flow cytometry analysis for MSC-specific markers
rbMSCs and rcMSCs at passage 3 were collected by TrypLE™ Select (Thermo Fisher Scientific), centrifuged at 1,500 rpm for 5 min, and resuspended in phosphate-buffered saline (PBS). Aliquots containing 1 × 105 cells were incubated with phycoerythrin (PE) or fluorescein isothiocyanate (FITC)—conjugated antibodies against rat CD45, CD90, CD29 [28] (BD Biosciences, San Jose, CA), CD44 (Biolegend Co., San Diego, CA), and CD34 (Santa Cruz Biotechnology, Dallas, TX). PE-conjugated antibodies against CD29, CD90, and FITC-conjugated antibodies against CD44 were used as MSC markers, and PE-conjugated antibodies against CD34 and FITC-conjugated antibodies against CD45 were used as hematopoietic markers. PE mouse IgG1 and FITC mouse IgG1 (Biolegend) were used as isotype controls. Data acquisition and analyses were performed using FACSVerse (BD Biosciences).
Mesodermal and ectodermal lineage cell differentiation
Regarding osteogenic differentiation, cells were seeded on a six-well plate (Sumitomo Bakelite Co.) and maintained in growth medium until confluence. rbMSCs and rcMSCs were cultured in MSCs Osteogenic Differentiation Medium (PromoCell, Heidelberg, Germany) for 14 days. Medium was changed every 3 days. Cells were finally stained with Alizarin red S solution (Sigma-Aldrich) for 30 min to confirm calcium deposition. The images of stained cells were captured using a phase-contrast microscope (ECLIPSE TE300; Nikon Co., Tokyo, Japan).
Regarding chondrogenic differentiation, cells were pelleted at 1,500 rpm for 5 min. Pellets were cultured in chondrogenic differentiation medium; α-minimum essential medium eagle, alpha modification (Sigma-Aldrich) with 0.1% penicillin streptomycin (Thermo Fisher Scientific), 1% GlutaMAX (Thermo Fisher Scientific), 10−7M dexamethasone (Sigma-Aldrich), 50 μg/mL ascorbate 2-phosphate (Sigma-Aldrich), 3.5 g/L D-(+)-glucose (Sigma-Aldrich), 100 μg/mL pyruvate (Sigma-Aldrich), and 1% ITS-plus (BD Biosciences) for 21 days. The medium was changed every 3 days. Paraffin sections of pellets were stained with alcian blue solution. The images of stained cells were captured using a multifunctional microscope (BZ-9000; KEYENCE Co., Osaka, Japan).
Regarding neural differentiation, we used modified neural differentiation conditions, including neural conditioning medium and neural differentiation medium, according to previously reported methods [27]. Specifically, MSCs were seeded on 6-cm dishes (Corning Co., New York, NY) and maintained in growth medium until 80% confluent. Medium was changed to neural conditioning medium containing Dulbecco's modified Eagle's medium: Nutrient mixture F-12 (Invitrogen Co., Carlsbad, CA) with 1% FBS (Thermo Fisher Scientific HyClone), basic fibroblast growth factor (100 ng/mL; PeproTech, Rocky Hill, NJ), penicillin (100 U/mL), and streptomycin (100 μg/mL; both from Sigma-Aldrich). After being incubated in neural conditioning medium for 3 days, cells were cultured in neural differentiation medium composed of neural conditioning medium with forskolin (10 μM; Sigma-Aldrich) added for 7 days. The expressions of NF-M and βIII-tubulin were then investigated with immunostaining. The images of stained cells were captured using a multifunctional microscope (BZ-9000; KEYENCE Co.).
Reverse transcription and real-time polymerase chain reaction
When cultured cells reached confluency on dishes using DMEM-L growth medium, they were collected in PBS. Total RNA was extracted with NucleoSpin® RNA (MACHEREY-NAGEL GmbH & Co. KG, Düren, Germany). Reverse transcription was performed with ReverTra Ace-α (Toyobo Co., Ltd., Osaka, Japan). Using cDNA as the template, real-time polymerase chain reaction (PCR) was performed with the 7,500 Real-time PCR system (Applied Biosystems, Carlsbad, CA). Snail, P75, and Slug were used as neural crest-associated markers. Brain-derived neurotrophic factor (Bdnf), glial cell line-derived neurotrophic factor (Gdnf), and nerve growth factor (Ngf) were used as neurotrophic factors. B-actin (Actb) was used as an internal endogenous control.
Middle cerebral artery occlusion surgical procedure, neurological function test, and cell transplantation
A total of 36 adult male SD rats weighing 250–300 g were subjected to middle cerebral artery occlusion (MCAO). The right common carotid artery, external carotid artery, internal carotid artery, and pterygopalatine artery were exposed under isoflurane anesthesia using a microscope. A 4-0 nylon monofilament suture coated with silicone (403745PK10; Doccol Corporation) was inserted through the external carotid artery into the internal carotid artery and up to the origin of the middle cerebral artery. Two hours after MCAO, reperfusion was performed by withdrawing the suture. The modified neurological severity score (mNSS) was used to evaluate motor and sensory functions. In mNSS, the level of functional recovery was graded on a scale of 0–18 based on motor, sensory, balance, and reflex tests, with higher scores reflecting more severe injury [8]. Rats with mNSS of 9–12 points on day 1 were used in the present study.
Before the transplantation analysis, we assessed brain infarction with 2,3,5-triphenyl tetrazolium chloride staining 24 h after MCAO (Fig. 1A) and hematoxylin and eosin staining 28 days after MCAO (Fig. 1B) in nine MCAO rats. We confirmed brain infarction in all rats with mNSS of 9–12 points on day 1.
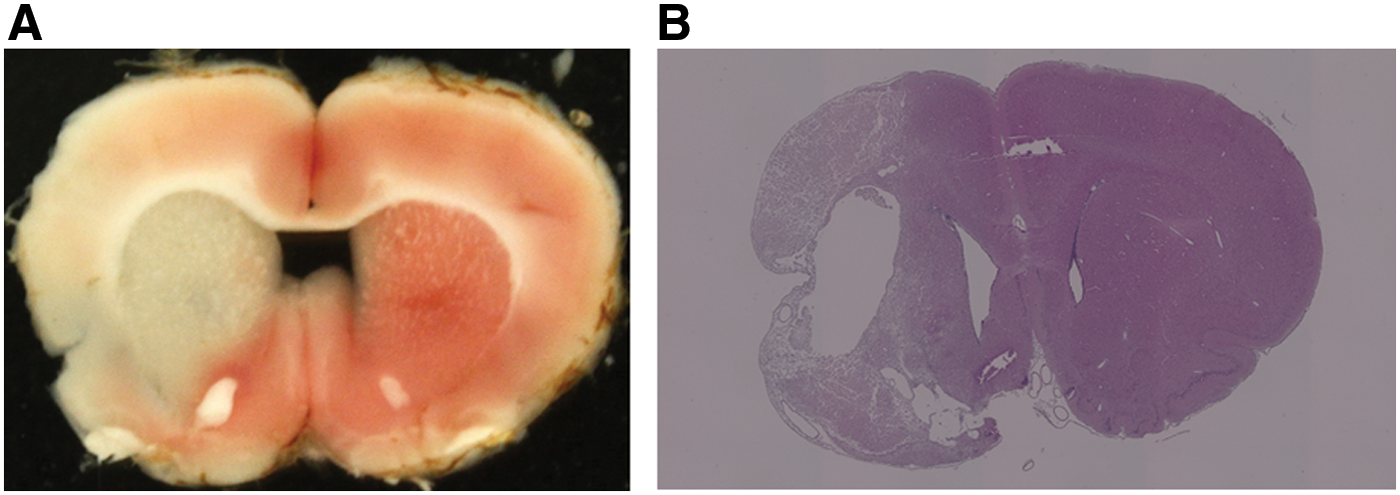
2,3,5-triphenyl tetrazolium chloride staining 24 h after MCAO
Rats were divided into the following three groups according to the treatment received: the administration of PBS only (Ctrl group, n = 8), the transplantation of rbMSCs (rbMSC group, n = 8), and the transplantation of rcMSCs (rcMSC group, n = 8). Rats in the rbMSC and rcMSC groups were injected with MSCs (1.0 × 106 cells/300 μL PBS) via the tail vein 24 h after MCAO. In all rats, mNSS was evaluated before and 1, 2, 4, 7, 14, 21, and 28 days after MCAO.
Brain sampling and analysis of the brain lesion site
To identify the transplanted cells in the recipient rats, rbMSCs and rcMSCs were prelabeled with PKH26 [29] (Sigma-Aldrich) and injected into MCAO model rats as a separate study. PKH26-labeled rbMSCs or rcMSCs transplantation was performed as described above. The rats were anesthetized 24 h after transplantation, and brains were removed and perfused with 4% paraformaldehyde in PBS. To examine the distribution of the fluorescently labeled cells, 14-μm-thick cryostat coronal brain sections were analyzed under a multifunctional microscope (BZ-9000; KEYENCE Co.). 4,6-Diamidino-2-phenylindole dihydrochloride (1:1,000; Kirkegaard and Perry Laboratories, Gaithersburg, MD) was used to stain nuclei. The brain of a rat injected with PBS via the tail vein 24 h after MCAO was used as a control (Ctrl).
Preparation of MSC conditioned medium and NG108–15 cell cultures
rbMSCs and rcMSCs were seeded on 6-cm culture dishes (Corning Co.) and maintained in growth medium. After cells had reached 80% confluency, medium was changed to fresh growth medium. The culture medium was collected 24 h later as conditioned medium (CM). After 0.2-μm filtration, rbMSC-CM and rcMSC-CM were stored at −80°C.
Neuroblastoma × glioma hybrid cells (NG108-15; ECACC, Porton Down, United Kingdom) were cultured in Dulbecco's modified Eagle's medium with high glucose (DMEM-H) (Sigma-Aldrich Co.) supplemented with 10% FBS (Thermo Fisher Scientific), penicillin (100 units/mL), streptomycin (100 μg/mL: both from Sigma-Aldrich), and HAT supplement (Thermo Fisher Scientific). Cells were maintained at 37°C in 5% CO2.
NG108-15 cell death assay with exposure to oxidative and inflammatory stresses
NG108-15 cells were exposed to oxidative and inflammatory stresses to evaluate the neuroprotective effects of rbMSCs and rcMSCs. H2O2 (Santoku Chemical Industries, Tokyo, Japan) was used to mimic an oxidative stimulus to cells, while lipopolysaccharide (LPS) (Wako Pure Chemical Industries, Osaka, Japan) was used to mimic an inflammatory stimulus to cells [30 –33]. NG108-15 cells were seeded on culture dishes (Sumitomo Bakelite Co.) and maintained in growth medium. Medium was changed to rbMSC-CM and rcMSC-CM (with 500 μM H2O2 or 200 ng/mL LPS) 48 h after seeding. Cells were collected 24 h after exposure to stress, centrifuged, suspended in PBS, and the cell survival rate was then assessed with a counting chamber (Sunlead Glass Corp., Saitama, Japan) using trypan blue staining. The remaining cells were collected for the mRNA expression analysis.
mRNA expression analysis of stress-exposed NG108-15 cells
Total RNA was extracted from NG108-15 cell samples and reverse transcription was performed as described above. Real-time PCR was performed using oligonucleotide primer sets corresponding to the cDNA sequences of rat b-cell leukemia/lymphoma 2 protein (Bcl2) and Bcl2-associated X protein (Bax).
Statistical analyses
Data were evaluated statistically using the Mann–Whitney U test for mRNA expression and survival rate analyses. Comparative analyses of the mRNA expression of p75 and Bdnf were performed using the delta Ct (ΔCt) method, in which ΔCt = Ct(target gene)–Ct(reference gene), and Fisher's exact test. Low ΔCt values indicated the strong expression of a gene, whereas high ΔCt values indicated the weak expression of a target gene relative to the expression of Actb [34]. A repeated-measures two-way analysis of variance with Bonferroni's test was used in the mNSS analysis. Statistical analyses were performed using JSTAT software (Sato, Japan). A P value <0.05 was considered to be significant.
Results
Cell surface marker expression in MSCs
MSC-specific markers in rbMSCs and rcMSCs were analyzed by flow cytometry. rbMSCs and rcMSCs had similar characteristics. Both types of isolated MSCs were strongly positive for CD29, CD90, and CD44 (cell surface markers associated with MSCs) but negative for CD34 and CD45 (cell surface markers associated with hematopoietic cells) (Supplementary Table S1; Supplementary Data are available online at
Differentiation potentials of both types of isolated MSCs
Both MSCs were morphologically spindle-shaped cells before differentiation (Fig. 2A). The differentiation potentials of the isolated rbMSCs and rcMSCs into osteogenic cells, chondrogenic cells, and neurons were estimated to identify them as MSCs.

Morphology and differentiation to osteogenic cells, chondrogenic cells, and neurons of both MSCs. Both MSCs were morphologically spindle-shaped cells before differentiation
Regarding osteogenic differentiation, both MSCs were negative for specific staining (Alizarin red S staining) before differentiation; however, positive cells were observed after differentiation. The number of Alizarin red-positive cells was higher among rbMSCs than rcMSCs (Fig. 2B).
Regarding chondrogenic differentiation, both MSCs were conglobulated in a micromass (Fig. 2C). Staining with alcian blue solution showed the tendency to differentiate into chondrogenic cells in both MSCs.
Regarding neural differentiation, both MSCs had similar differentiation potentials with NF-M (data not shown) and βIII-tubulin immunostaining (Fig. 2D).
Neural crest-associated gene expression profiles of both types of isolated MSCs
We used real-time PCR to evaluate neural crest-associated gene expression in rbMSCs and rcMSCs. The expression of Snail, p75, and Slug, which are associated with neural crest cells, was evaluated in rbMSCs and rcMSCs. The expression of Snail and p75 was significantly higher in rcMSCs than in rbMSCs (Snail: n = 6, P < 0.01; p75: n = 6, P < 0.05, Fig. 3A, B). The expression of Slug was slightly stronger in rcMSCs than in rbMSCs (n = 6, P = 0.083, Fig. 3C).
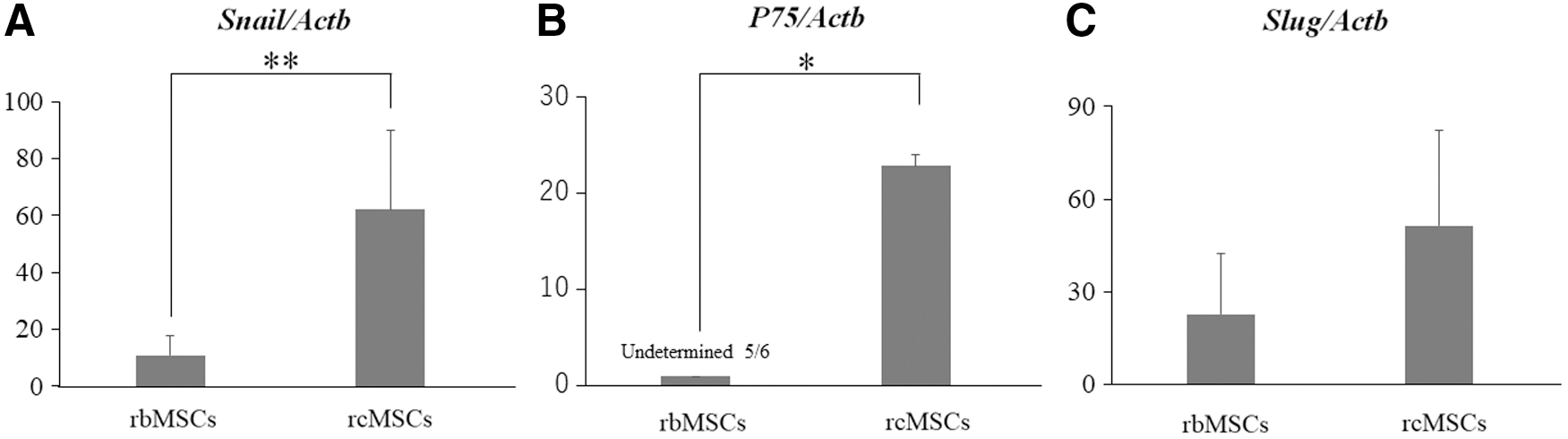
Real-time PCR analysis of neural crest-associated genes. Slug and Snail were detected in all cultured rbMSCs and rcMSCs
Neurotrophic factor gene expression profiles of both types of isolated MSCs
Neurotrophic factor gene expression in rbMSCs and rcMSCs was analyzed using real-time PCR. The expression of Bdnf, Gdnf, and Ngf, which function as neurotrophic factors, was evaluated in rbMSCs and rcMSCs. The expression of Bdnf and Ngf was significantly higher in rcMSCs (Bdnf: n = 6, P < 0.01; Ngf: n = 6, P < 0.05, Fig. 4A, C). The expression of Gdnf was slightly stronger in rcMSCs than in rbMSCs (n = 6, P = 0.057, Fig. 4B).

Real-time PCR analysis of neurotrophic factor-associated genes. The expression of Bdnf was undetermined in two out of six cultures of rbMSC
Recovery of neurological function in MCAO rats
We assessed neurological function using mNSS to compare the functional benefits of transplantation with rbMSCs and rcMSCs after MCAO. Improvements in mNSS were significantly greater in the rcMSC group than in the Ctrl and rbMSC groups 4, 7, 14, 21, and 28 days after MCAO (Fig. 5). Improvements were significantly greater in the rbMSC group than in the Ctrl group 4, 7, 21, and 28 days after MCAO.
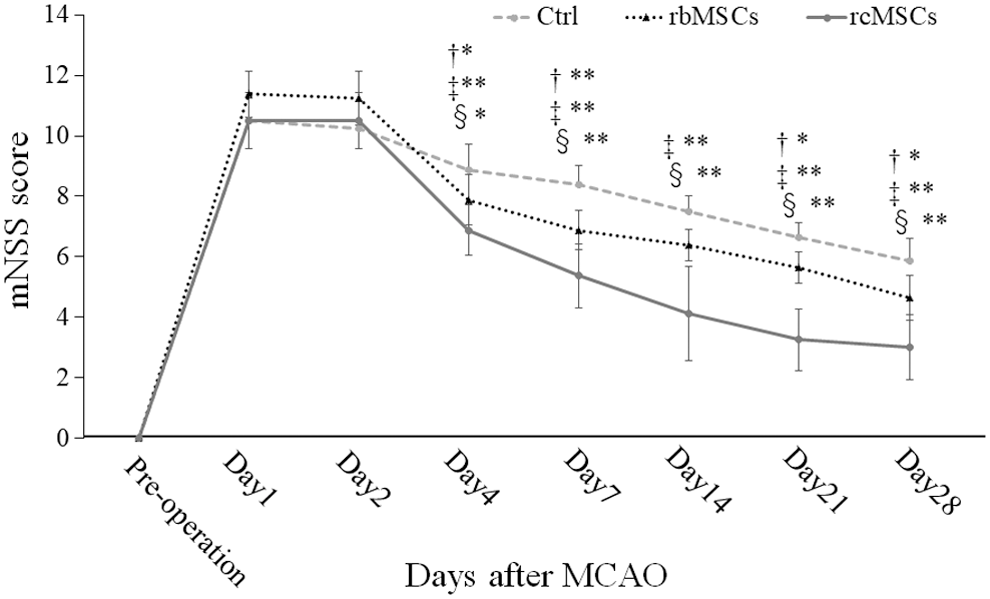
Results of the modified neurological severity score (mNSS). †Ctrl group versus rbMSC group; ‡Ctrl group versus rcMSC group; and §rbMSC group versus rcMSC group. Rats in the rcMSC group showed more significant improvements in mNSS than those in the rbMSC group 4, 7, 14, 21, and 28 days after MCAO (*P < 0.05, **P < 0.01, n = 8).
Localization of PKH-labeled cells
PKH26-labeled rbMSCs or rcMSCs injected intravenously 24 h after MCAO appeared in the brain lesion site 24 h after transplantation (Supplementary Fig. S1). There were no fluorescent cells in the Ctrl.
Survival rate of and mRNA expression in stress-exposed NG108-15 cells
rbMSC-CM and rcMSC-CM were used to estimate the paracrine effects of both types of isolated MSCs. The survival rate of NG108-15 cells exposed to oxidative and inflammatory stresses was significantly higher in those cultured in rcMSC-CM than in rbMSC-CM (oxidative stress exposure: n = 6, P < 0.05; inflammatory stress exposure: n = 6, P < 0.01, Fig. 6).
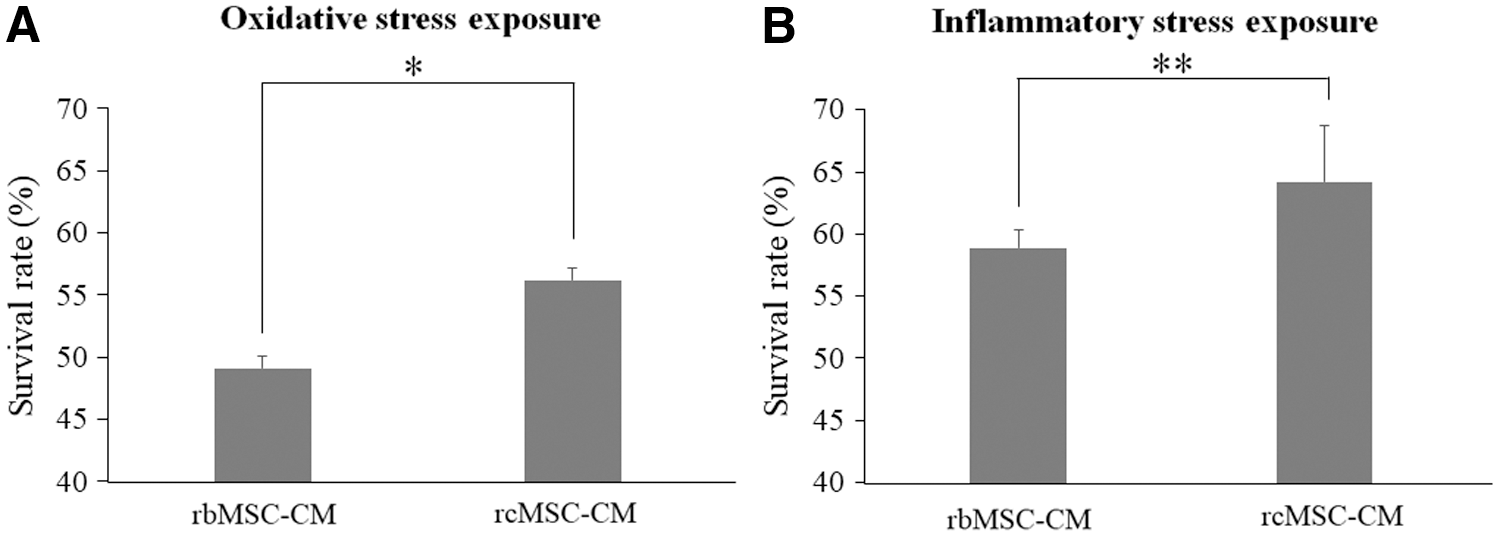
Effects of conditioned medium on the survival rate of stress-exposed NG108-15 cells. Survival rates of NG108-15 cells exposed to oxidative stress
The Bax/Bcl2 ratio in oxidative stress and inflammatory stress-exposed NG108-15 cells was significantly lower in those cultured in rcMSC-CM than in rbMSC-CM (oxidative stress exposure: n = 6, P < 0.01; inflammatory stress exposure: n = 6, P < 0.01, Fig. 7).
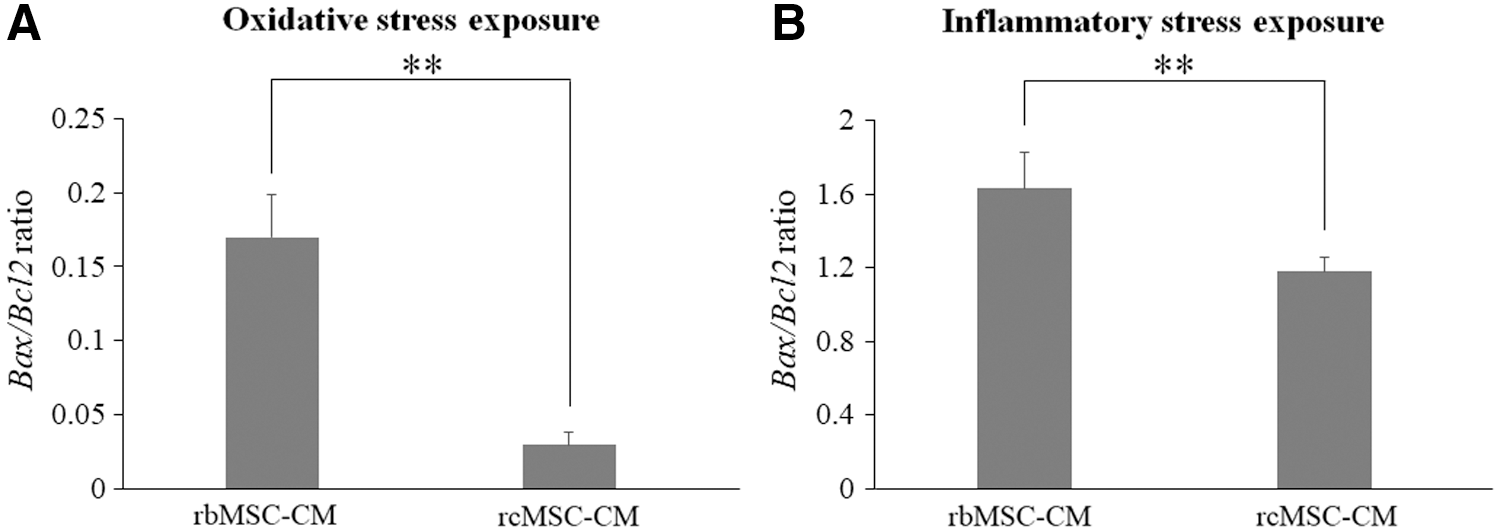
mRNA expression levels of stress-exposed NG108-15 cells. Bax/Bcl2 ratios in NG108-15 cells exposed to oxidative stress
Discussion
The present study showed that rcMSCs abundantly expressed neurotrophic factors and promoted excellent functional recovery after transplantation into ischemic stroke model rats. We successfully isolated rcMSCs and showed that these cells expressed more neural crest markers and neurotrophic factors than rbMSCs. In addition, we revealed that the transplantation of rcMSCs improved neurological functions in the rat MCAO model more than that of rbMSCs. The present results demonstrated that rcMSC-CM increased the survival rate of oxidative and inflammatory stress-exposed NG108-15 cells. The Bax/Bcl2 ratio in stress-exposed NG108-15 cells was significantly lower in those cultured in rcMSC-CM than in rbMSC-CM.
Cranial bones are flat bones that developmentally form mainly through intramembranous rather than endochondral ossification, and are mainly derived from the cranial neural crest. Bones of the limbs, iliac bone, and vertebrae are derived from the mesodermal germ layer [21]. MSCs of the long bones or iliac bone have been identified in bone marrow. Although cranial bones contain a small bone marrow space, Zhao et al. [35] showed that the sutures of cranial bones provided a niche for MSCs. In the present study, the expression of neural crest-associated genes was stronger in rcMSCs than in rbMSCs. These results suggested that rcMSCs were also derived from the neural crest and not the mesodermal germ layer. The developmental specificity of rcMSCs may lead to the abundant expression of neurotrophic factors.
In the present study, we used ischemic stroke model rats. Many events are involved in neural cell death in the ischemic stroke brain. Apoptosis is initially triggered by calcium influx, impaired mitochondria, and energy depletion, and this is followed by glutamate excitotoxicity as a result of the depletion of oxygen and glucose [36]. The release of nitric oxide, oxygen free radicals, and other reactive oxygen species (ROS) then induces further damage to neurons. In addition, the destruction of the blood brain barrier by the release of matrix metalloproteinases and other proteases from endothelial cells leads to the infiltration of immune cells [37]. Cytokines released by immune cells also promote inflammatory reactions and increase brain injury. Thus, the control of apoptosis or inflammation may be an obvious therapeutic target for the treatment of the ischemic stroke brain, particularly in the acute phase.
Previous studies showed that MSCs exhibited the potential for neural differentiation in vitro [38 –42]; however, their differentiation potential in vivo remains unclear. Cell replacement may not be the main therapeutic effect of the transplantation of MSCs on ischemic brain injury in vivo, and it has been proposed and commonly accepted that the functional benefits of the transplantation of MSCs are due to paracrine effects via a series of secreted trophic factors, which promote ischemic brain tissue repair [43 –47]. We also speculate that rcMSCs secrete abundant neurotrophic factors that induce functional recovery of the MCAO rats transplanted rcMSCs in our in vivo study. Neurotrophic factors exert a number of neuroprotective effects that include the suppression of local immune reactions and inflammation, the inhibition of fibrosis (scar formation) and apoptosis, enhancements in angiogenesis, and the stimulation of tissue-intrinsic stem cell proliferation and differentiation [48]. In the present study, the expression of Bdnf and Ngf was significantly stronger in rcMSCs than in rbMSCs. Brain-derived neurotrophic factor (BDNF), in particular, is an important neurotrophic factor that promotes neurogenesis and angiogenesis, offers neuroprotection, modulates inflammation and apoptosis, and may improve synaptic plasticity after ischemic brain injury [49 –51]. Matsuda et al. [52] reported that BDNF prevented the endothelial barrier dysfunction induced by interleukin-1β and tumor necrosis factor-α (TNF-α) and regulated inflammation. Takeda et al. [53] found that BDNF protected human endothelial cells from TNF-α-induced apoptosis by regulating the phosphatase and tensin homolog/AKT pathway, while Liu et al. [54] showed that BDNF reduced neuronal apoptosis by increasing the expression of the Bcl-2 antiapoptotic protein and inhibiting intracellular calcium overload. Nerve growth factor (NGF) is also a neurotrophic factor and its neuroprotective effects have been demonstrated in vitro [55 –57]. In the ischemic brain, the ROS-induced activation of apoptosis signal-regulating kinase 1 (ASK1) plays crucial roles in oxidative stress-mediated cell death [58]. Kirkland et al. [59] reported that the rapid activation of antioxidant defenses by NGF suppressed ROS. Thus, NGF also exerts suppressive effects on apoptosis associated with oxidative stress. In the present study, rcMSCs abundantly expressed neurotrophic factors, including BDNF and NGF. Therefore, the transplantation of rcMSCs may induce excellent functional recovery in MCAO rats.
The results of our in vitro study using stress-exposed NG108-15 cells appear to support those of the in vivo study. The survival rate of NG108-15 cells was significantly higher in the presence of rcMSC-CM than in rbMSC-CM. Regarding mRNA expression in NG108-15 cells, the Bax/Bcl2 ratio was significantly lower in rcMSC-CM when these cells were exposed to oxidative and inflammatory stresses. These results demonstrate that rcMSC-CM suppressed apoptosis in NG108-15 cells following the exposure to oxidative and inflammation stresses. We assumed that rcMSC-CM included abundant amounts of BDNF and NGF secreted from cultured rcMSCs, which suppressed apoptosis in response to oxidative and inflammatory stresses and inhibited NG108-15 cell death.
We assumed that rcMSCs secreted abundant neurotrophic factors, and their transplantation induced excellent functional recovery in ischemic stroke model rats. Therefore, cranial bones may be an ideal source of MSCs for the treatment of CNS diseases. The present study establishes the foundation for further studies to assess the efficacy of using cranial bones in cell-based therapy. Further studies on transplantation using MSCs derived from human cranial bones are needed before the initiation of clinical trials.
Conclusion
We herein provide novel evidence to show that cell-based therapy using rcMSCs resulted in excellent functional recovery in ischemic stroke model rats, at least through the secretion of abundant neurotrophic factors. Although further studies using MSCs derived from human cranial bones are needed, our results suggest that cell-based therapy using MSCs derived from cranial bones is a useful therapeutic strategy for ischemic stroke.
Footnotes
Acknowledgments
This work was partially supported by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (JSPS KAKENHI grant no. 16K10724). A part of this work was carried out at the Analysis Center of Life Science, Natural Science Center for Basic Research and Development, Hiroshima University.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.