
Abstract
Glial cell line-derived neurotrophic factor (GDNF) exhibits potent neuroprotective properties in preclinical models of Parkinson's disease (PD), but challenges in GDNF delivery have been reported from clinical trials. To address this barrier, we developed a hematopoietic stem cell transplantation-based macrophage-mediated GDNF therapy platform. Here, we introduced a regulatable lentiviral vector (LV-MSP-Tet-Off-hGDNF) to allow the expression of human GDNF (hGDNF) to be adjusted or stopped by oral administration of doxycycline (Dox). C57BL/6J mice were lethally irradiated with head protection and then transplanted with syngeneic bone marrow cells transduced with either the hGDNF-expressing vector or a corresponding GFP-expressing vector, LV-MSP-Tet-Off-GFP. Suppression of vector gene expression was achieved through administration of Dox in drinking water. To create a toxin-induced Parkinsonian model, mice were injected in two cycles with MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) to yield nigral cell/striatal dopamine loss and behavioral deficits. During the presence of Dox in the drinking water, plasma GDNF was at a basal level, whereas during the absence of Dox, plasma GDNF was significantly elevated, indicating reliable regulation of therapeutic gene expression. Midbrain GDNF levels were altered in parallel, although these did not return completely to basal levels during the periods of Dox withdrawal. Motor activities of the MPTP-Tet-off-hGDNF group were comparable to those of the Tet-off-GFP (subject to no MPTP treatment) group, but substantially better than those of the MPTP-Tet-off-GFP group. Interestingly, the improvement in motor activities was sustained during the Dox-withdrawn periods in MPTP-Tet-off-hGDNF animals. Neuroprotection by therapeutic GDNF expression was further evidenced by significant amelioration of nigral tyrosine hydroxylase loss after both the first and second MPTP treatment cycles. These data suggest that neurotrophic factor expression can be upregulated to achieve efficacy or downregulated in case of off-target effects or adverse events, a feature that may eventually increase the acceptance of this potentially neuroprotective/disease-modifying PD therapy.
Introduction
P
Unfortunately, GDNF crosses the blood-brain barrier (BBB) poorly (<2%). Clinical trials delivering GDNF directly into the brain of PD patients via intraputamenal infusion have yielded mixed results, two open-label trials showing efficacy [9,10], while one placebo-controlled trial concluded a lack of benefit [11]. Clinical trials of gene therapy using NRTN (neurturin, AAV-NRTN), a close GDNF analog that shares similar biological functions, have also failed to achieve their primary endpoints in PD patients [12]. Taken together, focal administration of GDNF has been unsuccessful, as this method of delivery may cover <10% (GDNF protein) or <19% (gene transfer) of the target neurons [2,13,14]. Although infusion of GDNF using multiple catheters has been effective in monkeys having a brain one 15th the size of the human brain, the human putamen may be too large for effective delivery. Moreover, extremely low rate of molecular diffusion through brain tissue limits convection-enhanced delivery of GDNF [15 –18]. The degeneration of axonal fibers may nullify retrograde transport of GDNF to the substantia nigra (SN), further limiting this approach [12]. Consequently, a sufficient population of the degenerating neurons may not be accessible for GDNF molecules to exhibit a therapeutic effect from direct injection strategies. The concern that nonresponsiveness of PD patients to GDNF treatment was due to the absence or low levels of canonical GDNF receptor Ret in the degenerating dopamine (DA) neurons was not supported by recent findings suggesting that the GDNF signaling machinery was normal in patients with sporadic PD–that is, there was no significant decrease in expression levels for Ret, Nurr1, and other associated genes [19].
In contrast, experiments using bone marrow transplantation have shown that circulating monocytes/macrophages are preferentially recruited through the BBB to sites of neuronal degeneration where they may become microglial cells [20,21]. These results have been brought into question by findings demonstrating that microglia colonize developmentally during embryogenesis and are not replaced by bone marrow cells throughout the natural lifetime in mice, unless the brain is subjected to defined pathologic conditions such as irradiation [22 –25]. However, recent findings demonstrate that such conditioning is not essential to facilitate extravasation of circulating monocytes into the central nervous system (CNS). Both intravenously injected and endogenous monocytes/macrophages infiltrate the CNS without conditioning in mouse models of neurodegenerative diseases [26 –29] and psychological stress [30]. We and others [31,32] have shown that monocyte-derived macrophages can infiltrate the brain parenchyma of Parkinsonian mice (genetic [Mitopark] or toxin [MPTP] models) after hematopoietic stem cell (HSC) transplantation (HSCT) following head-protected irradiation conditioning. Furthermore, we observed brain infiltration of macrophages in PD mouse models without the use of irradiation or conditioning (data not yet published). Our studies have reproducibly shown that HSCT-based macrophage-mediated gene therapy effectively delivers GDNF to degenerating dopaminergic neurons, reducing DA neuron loss and ameliorating motor as well as nonmotor Parkinsonian symptoms.
HSC gene therapy is achieved through transplantation of the cells transduced with retroviral or lentiviral vectors carrying a therapeutic gene. Both retroviral and lentiviral vectors can integrate genetic materials into the host cell genome permanently, leading to stable, life-long expression of a therapeutic gene. Although this is the preferred outcome, the currently irreversible nature of HSC gene therapy may be accompanied by off-target effects and potential adverse events. Therefore, a controllable system that enables regulation of GDNF expression would be desirable. Importantly, regulation of the expression of therapeutic genes can further tailor therapies to meet the needs of PD patients. Regulatable adeno-associated virus (AAV) vectors have been tested in rodents for delivering the GDNF gene to neurons [33 –35]. Furthermore, control of gene expression in the hematopoietic system was studied using an inducible lentiviral vector [36]. Here, we report a doxycycline-regulatable lentiviral vector transduced into HSCs pretransplantation for macrophage-mediated delivery of GDNF to dopaminergic neurons in the MPTP mouse model of PD.
Materials and Methods
Animals
Eight-week-old female C57BL/6J mice were purchased from Jackson Laboratory (Bar Harbor, Maine) and used for Dox-regulated human GDNF (hGDNF) expression studies. Mice were group-housed in standard shoebox cages with ad libitum access to food and water. The room was maintained at 26°C on a 12 h light/12 h dark cycle. All animal care and use was in accordance with the Guide for the Care and Use of Laboratory Animals, 8th edition (National Research Council). The procedures for all animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Texas Health Science Center at San Antonio.
Vector construction and virus preparation
A lentiviral vector containing a macrophage-specific promoter (MSP) was designed as described previously [37]. The Lenti-MSP-GDNF vector was constructed by replacing the reporter gene (GFP) in the original design with the hGDNF gene (gene bank No. NM000514, STS 201-836) using standard molecular cloning procedures. A similar lentiviral vector carrying the gene that encodes GFP driven by the MSP was also generated and used as a control. The rtTR-KRAB-2SM2 fragment derived from pLVT-hGDNF-rtTRKRAB2SM2 plasmid (Addgene, Cambridge, MA) was amplified by PCR using a set of primers for the GDNF vector (forward primer: 3′-CTCTAGAACTAGTGGATCCCC-5′, reverse primer: 3′-CGATTTAGGTGACACTATAGAATAC-5′) and second set of primers for the GFP vector (forward primer: 3′-GTACAAGTAACTCTAGAACTAGTGGATCCCC-5′, reverse primer: 3′-CGATTTAGGTGACACTATAGAATAC-5′). The Dox-regulated (tet-off) lentiviral GDNF or GFP vectors were constructed by inserting the amplified rtTR-KRAB-2SM2 fragments into the 7.8 kb BsrGI-PmeI fragments of the Lenti-MSP-GDNF or Lenti-MSP-GFP backbone (Fig. 1). The resulting construct was sequenced to verify the site of insertion.
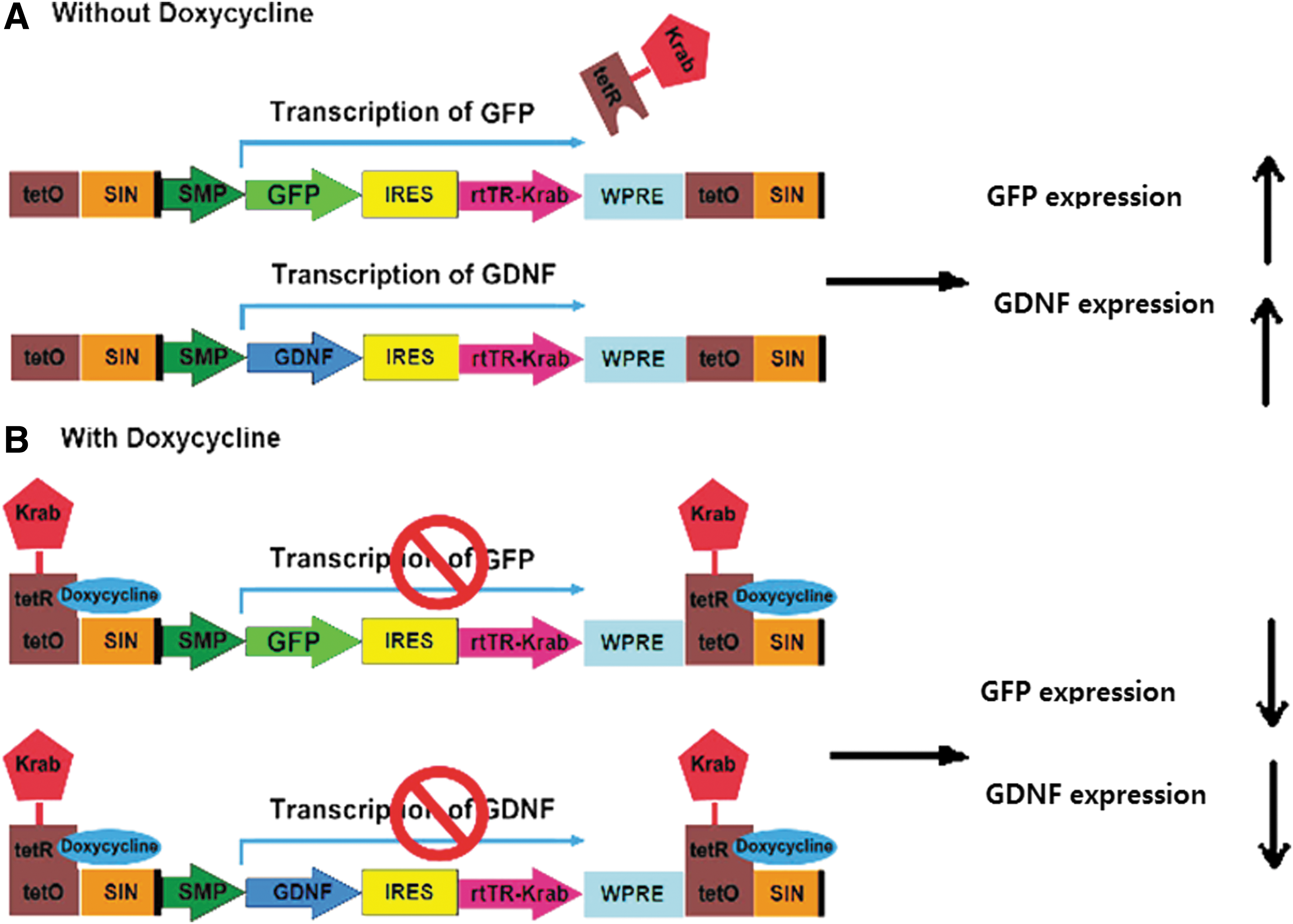
Schematic representation of the doxycycline-controllable lentiviral vector (LV-MSP-Tet-Off-hGDNF and LV-MSP-Tet-Off-GFP) design. GFP and hGDNF expression was driven by a Dox-regulated MSP.
Viruses were prepared as described previously [25,37]. Briefly, the LV-MSP-Tet-off-hGDNF or LV-MSP-Tet-off-GFP plasmid was co-transfected with packaging plasmids pMDLg/pRRE, pRSV/Rev, and pMD.G2 into 293T cells. The culture media containing lentiviral particles were collected 24 and 48 h post-transfection, filtered through 0.45-μm pore size filters, and then concentrated 1,000-fold by two rounds of ultracentrifugation at 56,000 g for 2 h and then 72,000 g for 1.5 h at 4°C. The viral pellets were resuspended in StemPro-34 SFM medium (Invitrogen, Carlsbad, CA) and stored at −80°C up to use.
Lentiviral transduction and transplantation of bone marrow cells
Mice were randomly divided into three weight-matched groups with 16 animals per group. Group 1 (MPTP-Tet-off-hGDNF) was transplanted with bone marrow cells that had been transduced with the Dox-regulated lentiviral vector expressing hGDNF cDNA (Tet-off hGDNF), and subsequently treated with MPTP. Group 2 and 3 were transplanted with the bone marrow cells that had been transduced with Dox-regulated lentiviral vector expressing GFP cDNA (Tet-off GFP). Group 2 (MPTP-Tet-off-GFP) was then treated with MPTP, while group 3 (Tet-off-GFP) served as a non-MPTP control. Lentiviral transduction and bone marrow cell transplantation (BMT) were performed as described previously [38]. Briefly, syngeneic bone marrow cells were harvested from the tibias and femurs of 8-week-old C57BL/6J donor mice by flushing the bones with StemPro-34-SFM complete medium (GIBCO Invitrogen) supplemented with 2 mM L-glutamine, 100 IU/mL penicillin, 100 μg/mL streptomycin, and 5 U/mL heparin, and then passed through a 100 μm nylon mesh cell strainer (BD Biosciences, San Jose, CA) to make a single cell suspension. Bone marrow-derived stem cells were enriched in the lymphocyte medium using density gradient separation. After washing, enriched bone marrow-derived stem cells were prestimulated overnight in StemPro-34-SFM supplemented with 2 mM L-glutamine, 6 ng/mL of murine interleukin-3, 10 ng/mL of human interleukin-6, 10 ng/mL of murine interleukin-1, and 100 ng/mL of murine stem cell factor (PeproTech, Rocky Hill, NJ). The next day, harvested cells were pelleted and resuspended in 800 μL of concentrated viral supernatant that was supplemented with the aforementioned growth factors. Infections were performed on RetroNectin- (Takara, Otsu, Japan) coated plates for 6 h in the presence of 4 μg/mL of protamine sulfate (Sigma, St Louis, MO). Following infection, cells were harvested and 4 × 106 cells were transplanted into recipient mice previously irradiated with 950 cGy using a cesium γ-source while recipients' heads were protected with lead caps. Doxycycline was dissolved in 5% sucrose solution and supplied to all animals in drinking water (1 mg/mL) beginning on the day of transplantation and continuing (or not) as specified below. Doxycycline solution was freshly prepared every three to 4 days in a brown drinking bottle.
MPTP treatment and behavioral test
On day 3 post-transplant, MPTP-HCl (Sigma) dissolved in physiological saline was administered to animals in groups 1 and 2 (20 mg/kg) via intra-peritoneal injection, four times at 2-h intervals (designated first treatment cycle). Fifteen days after BMT and 12 days post-MPTP, the Dox was withdrawn (Dox−) from all mice for 2 months (Dox− = GDNF and GFP expression switch ON). Based on mouse behavioral measurement 4 months after MPTP, we performed a second MPTP treatment to achieve progressive motor deficits (and neuron loss). Doxycycline was given (Dox+ = GDNF and GFP expression OFF) and removed (Dox− = GDNF and GFP expression ON) at 2-month intervals for up to 8 months post-transplant (Fig. 2). Mouse behavioral testing was performed every 2 months. The mice were acclimatized to the testing room for a minimum of 1 h before testing. Horizontal and vertical locomotor activities were measured using the Photobeam Activity System (San Diego, CA) before and after BMT on each experimental group according to the manufacturer's protocol.

Overview of the experimental timeline. Mice were administered doxycycline in the drinking water for 2 weeks following BMT. Removal of doxycycline initiated expression of GDNF. Dox-regulated GDNF expression was alternated every 2 months for a total of 8 months. To induce DA neuron loss, MPTP treatment was performed twice: (1) 3 days post-BMT and (2) 4-months post-BMT. BMT, bone marrow cell transplantation.
Enzyme-linked immunosorbent assay
Plasma was prepared from peripheral blood collected in EDTA-treated tubes. A 0.5 mm thick coronal midbrain tissue slice containing SN (defined as SN for simplicity) was dissected out using a mouse brain slicer matrix (Zivic Insruments, Pittsburgh, PA) and stored at −80°C until analysis. The frozen tissue samples were homogenized in tissue lysis buffer containing 137 mM NaCl, 20 mM Tris (pH 8.0), 1% NP-40, 10% glycerol, 1 mM phenylmethylsulfonyl fluoride, 10 μg/mL aprotinin, 1 μg/mL leupeptin, and 0.5 mM sodium vanadate at a tissue concentration of 30 mg/mL and centrifuged at 20,000 g for 10 min at 4°C. The GDNF concentrations in plasma and SN from each experimental group were measured using a commercially available GDNF Emax ImmunoAsssay System (Promega, Madison, WI) according to the manufacturer's protocol. The antibody used in the kit reacts with both human and mouse GDNF.
Tissue processing and immunohistochemistry
Mice were anesthetized with ketamine hydrochloride solution (Butler Schein, Dublin, OH) and perfused transcardially with 20 mL of phosphate-buffered saline (PBS), pH 7.4 followed by an equal volume of 4% paraformaldehyde solution. Brains were dissected out and fixed in the same fixative overnight at 4°C, cryo-protected in sequential 10% (2 h), 20% (2 h), and 30% sucrose in PBS solution overnight, and then embedded in Tissue-Tek OCT compound (Sakura Finetek USA, Torrance, CA) and processed for cryosectioning at 30 μm thickness in the coronal plane. Four series of slides, each containing every fourth section were prepared for SN. Anatomical landmarks were determined according to Paxinos and Franklin [39]. The brain sections were treated with 1% bovine serum albumin in PBS containing 0.3% Triton X-100 for 30 min and then incubated with antityrosine hydroxylase (TH) antibody (Millipore, Billerica, MA) at 1:1,000 dilution overnight at 4°C. The sections were rinsed with PBS and incubated with biotinylated goat anti-rabbit secondary antibody (1:100) for 1 h, followed by avidin-biotin peroxidase complex (ABC Elite Kit, Vector Laboratories, Burlingame, CA) at room temperature for 1 h. The chromogen was either 3-amino-9-ethyl carbazole (AEC Chromogen Kit, Sigma) or 3,3′-diaminobenzidine tetrahydrochloride (Liquid DAB Substrate Kit, Invitrogen). The images were captured with a fluorescent microscope (Nikon Eclipse TE2000-U; Nikon Instruments, Melville, NY). The number of TH+ dopaminergic neurons in substantia nigra parc compacta (SNpc) were counted from digitized images of each of five sections per group using NIH ImageJ software (NIH, Bethesda, MD) by two blinded investigators following the same procedure and criteria.
Measurement of body weight and food intake
Mice were group-housed in standard mouse cages with ad libitum access to food and water. Body weight was measured before and every 2 weeks after Dox-regulated (Tet-off) hGDNF- or GFP-expressing BMT. Food intake was determined by measuring the mass of food per cage at the same time daily for 5 consecutive days during each month. Values were normalized to the number of mice per cage.
Statistical analysis
Data are presented as mean ± the standard error of the mean (SEM). All statistical analyses of the data were performed using GraphPad Prism 6.02 (GraphPad Software, LaJolla, CA). Multiple group comparisons were analyzed by two-way ANOVA, followed by post hoc analyses using Bonferroni post-test. The nonparametric comparison between the groups was analyzed by one-way ANOVA, followed by Tukey's post-test. Differences among treatment groups were considered statistically significant at *P < 0.05, **P < 0.01, ***P < 0.001.
Results
Plasma levels of GDNF responded to doxycycline administration
Although GDNF gene therapy strategies have shown promising therapeutic effects on motor dysfunction, sustained GDNF expression may lead to unpredictable GDNF accumulation in tissues, favoring potential adverse effects. Thus, we developed a tetracycline-regulated MSP-GDNF lentiviral vector–LV-MSP-Tet-Off-hGDNF (Fig. 1)—to control GDNF expression in wild-type C57BL/6J mice. As shown in Fig. 3, blood plasma levels of GDNF in Tet-off-hGDNF-transplanted mice exhibited a sensitive response to the administration of (or lack of) doxycycline, displaying significantly higher levels during dox withdrawal periods (3.432 ± 0.323 and 3.899 ± 0.269 ng/mL during the first and second periods, respectively). During dox administration, Tet-off-hGDNF-transplanted mice expressed basal plasma levels of GDNF (0.252 ± 0.096 and 0.215 ± 0.107 ng/mL during the first and second periods, respectively), comparable to the nearly undetectable plasma levels of GDNF in Tet-off-GFP-transplanted mice, likely representing low levels of endogenous murine GDNF. These data indicate that GDNF expression can be regulated by turning the Dox switch ON or OFF.
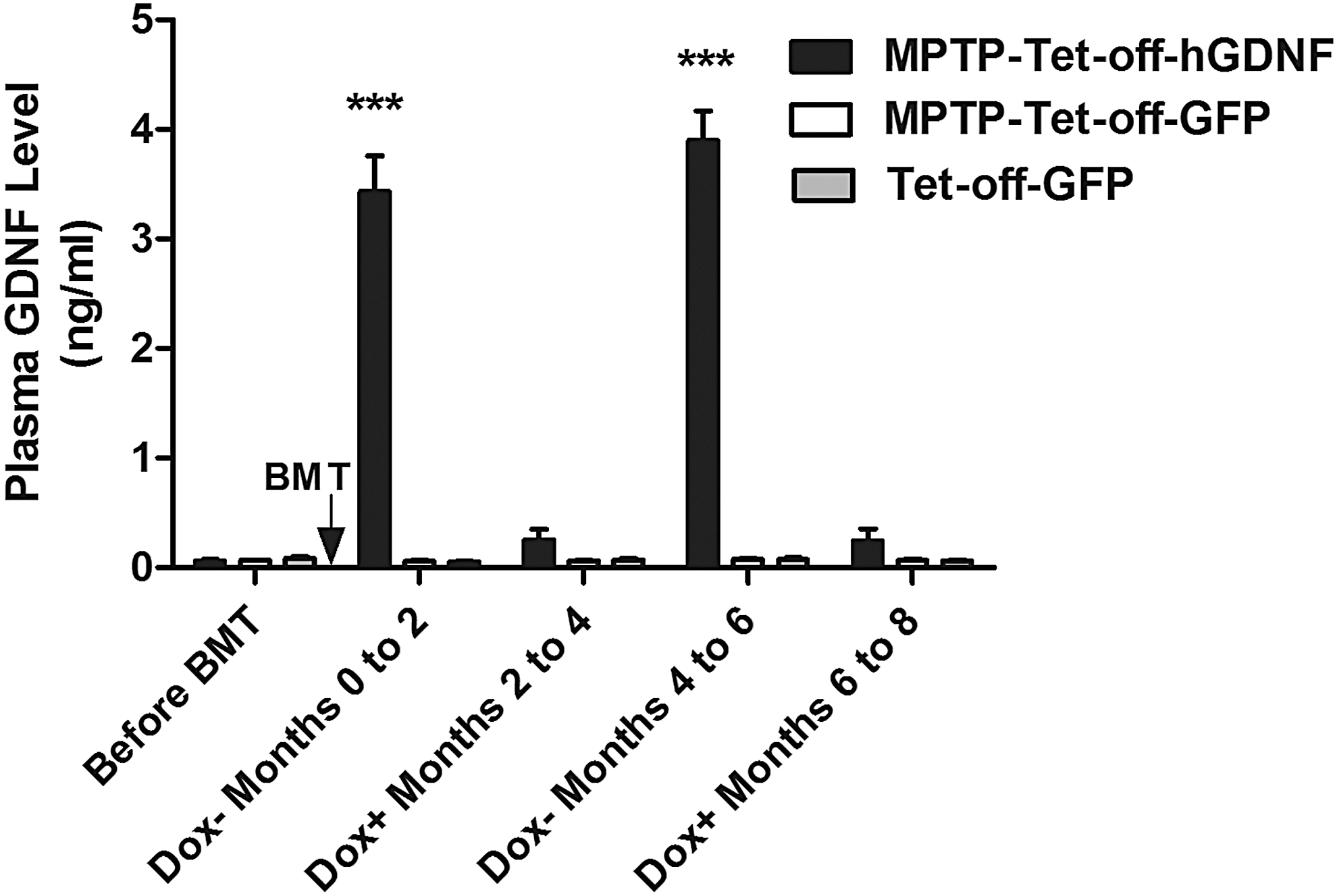
Plasma GDNF levels in Dox-regulated (Tet-off) hGDNF- and GFP-transplanted mice. Plasma samples were collected from MPTP-treated and non-MPTP-treated mice, under Dox-withdrawn and Dox–administered (1 mg/mL in drinking water) conditions, alternated every 2 months, at 2, 4, 6, and 8 months post-tet-off-hGDNF and -GFP BMT. GDNF concentrations in plasma were measured using a GDNF Emax ImmunoAsssay System (Promega, Madison, WI) according to the manufacturer's protocol. Each bar represents mean ± SEM from 16 animals per group (n = 16). ***P < 0.001 versus MPTP-Tet-off-GFP- and Tet-off-GFP-transplanted group. SEM, standard error of the mean.
Doxycycline-regulated GDNF expression protected against MPTP-induced motor deficits
The therapeutic effects of GDNF expression on motor deficits were assessed by locomotor activity test. The MPTP-Tet-off-GFP group displayed significant declines in both horizontal and vertical activities following MPTP treatment (Fig. 4A, B). Horizontal and vertical locomotor activities for these mice decreased by 32.6% and 41.4% following a single treatment with MPTP, further decreasing an additional 53.6% and 37.7% after the second treatment with MPTP, respectively (∼85% and 80% total decrease from baseline). In contrast, the MPTP-Tet-off-hGDNF group revealed significant protection from MPTP-induced motor deficits under the Dox-regulated GDNF expression, exhibiting horizontal and vertical locomotor activities similar to those of non-MPTP-treated Tet-off-GFP controls.
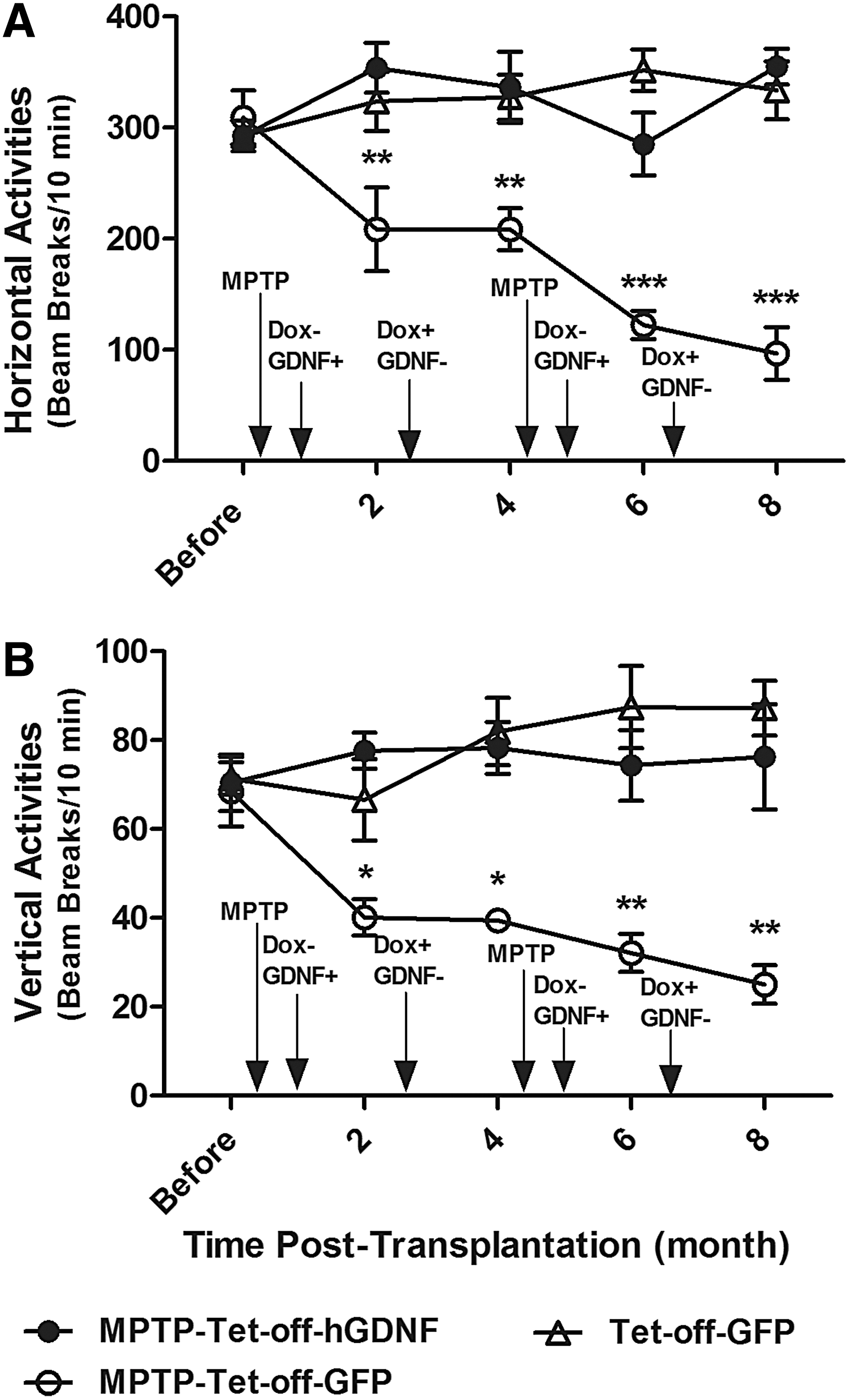
Doxycycline-regulated GDNF expression protected against MPTP-induced motor deficits. Ten-week-old female C57BL/6J mice were transplanted with bone marrow cells that had been transduced with Dox-regulated (Tet-off) lentiviral vectors containing either hGDNF or GFP cDNA (Fig. 1) and given 1 mg/mL of Dox in drinking water. Three days after transplantation, MPTP was administered to Tet-off-hGDNF (MPTP-Tet-off-hGDNF) and Tet-off-GFP (MPTP-Tet-GFP) transplanted animals. The control group (Tet-off-GFP) received donor cells containing a Tet-off-GFP vector, but were not treated with MPTP. Fifteen days after BMT and 12 days post-MPTP, Dox was withdrawn (Dox−) from drinking water for 2 months (Dox− = GDNF or GFP expression ON). Thereafter, administration of doxycycline was alternated every 2 months until 8 months post-BMT. The horizontal
Doxycycline-regulated GDNF expression ameliorated MPTP-induced dopamine neuron loss in SN
Next, we investigated whether Dox-regulated expression protected against DA neuron loss during MPTP administration. DA neuron loss was characterized in brain tissues from five randomly selected mice from each cohort (GFP, MPTP-GFP, and MPTP-hGDNF) after either one or two treatments with MPTP. A significant loss of DA neurons was observed in the SNpc of Tet-off-GFP transplanted mice after one MPTP treatment cycle, and the loss became substantially more severe after the second treatment cycle (Fig. 5). Mice expressing GDNF (Tet-off-hGDNF) revealed considerable neuroprotection against DA neuron loss, even after two MPTP treatment cycles (Fig. 5A). Approximately 2,665 ± 193 TH-stained DA neurons were observed in the SNpc isolated from non-MPTP-treated Tet-off-GFP control mice. We observed a 63% decrease in DA neurons in Tet-off-GFP MPTP mice after the first MPTP treatment cycle, plus an additional 22% decrease after the second treatment cycle (total ∼85% loss). Importantly, Dox-regulated expression of GDNF in Tet-off-hGDNF MPTP mice revealed significant DA neuroprotection, exhibiting only 30% and 12% (total 42%) decreases in DA neurons after the first and second MPTP treatment cycles, respectively (Fig. 5B).
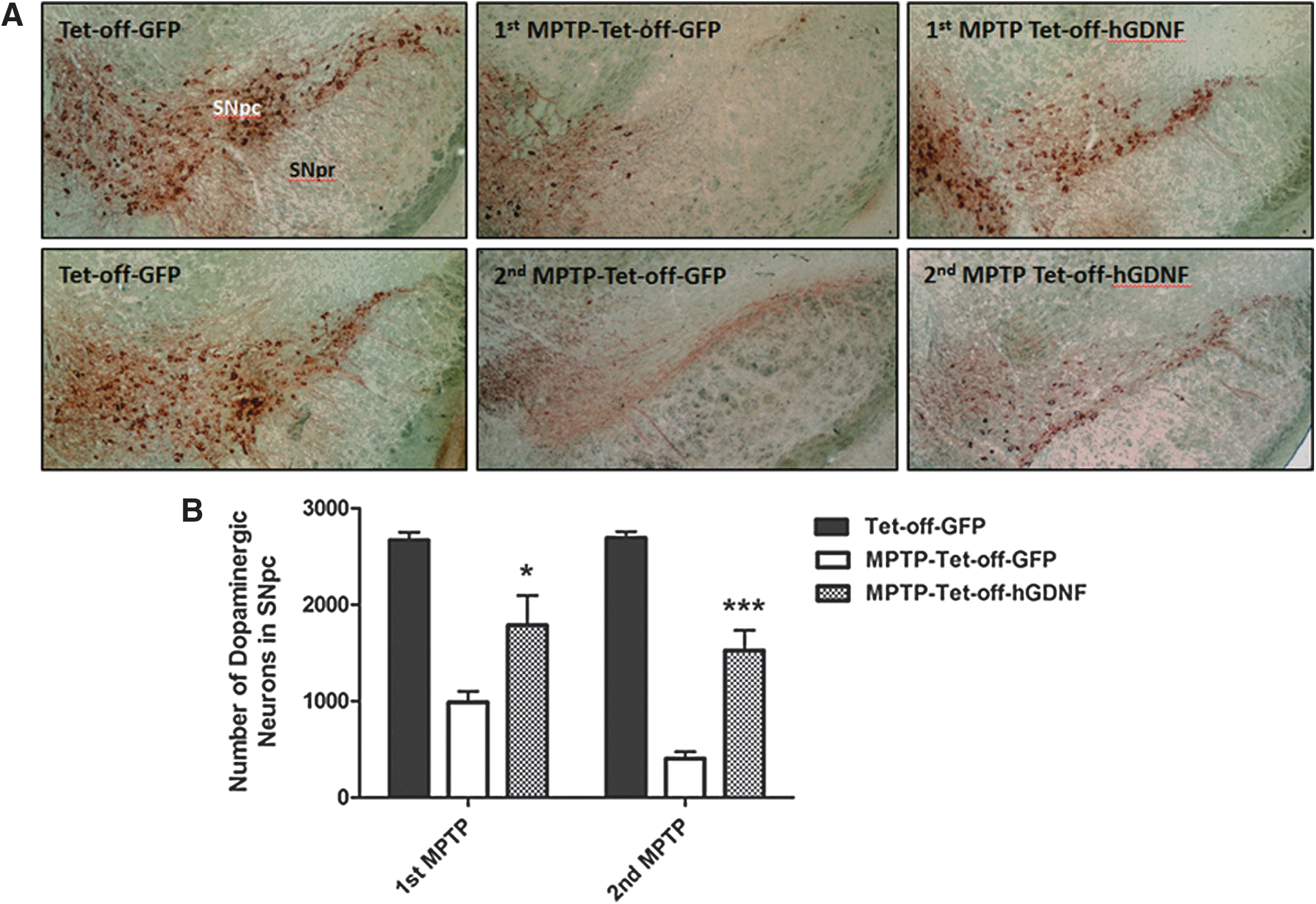
Doxycycline-regulated GDNF expression protected against MPTP-induced loss of dopaminergic neurons in mouse SN. Ten-week-old female C57BL/6J mice were transplanted with bone marrow cells (BMT) that had been transduced with Dox-regulated (Tet-off) lentiviral vectors expressing hGDNF or GFP cDNA. MPTP was administered to Tet-off-hGDNF (Tet-off-hGDNF MPTP) and Tet-off-GFP (Tet-off-GFP MPTP) -transplanted mice. First treatment at 3 days post-transplantation; second treatment 4 months post-transplantation. Control group (Tet-off-GFP) received Tet-off-GFP transplantation without MPTP treatment.
Changes in nigral GDNF content by doxycycline management
Next, we measured the GDNF concentration in the SN of GFP, MPTP-GFP, and MPTP-hGDNF mice during Dox-OFF or Dox-ON stages using enzyme-linked immunosorbent assays (ELISA). The SN GDNF concentrations in the MPTP-Tet-off-hGDNF-transplanted mice were 62.33 ± 8.05 and 78.96 ± 12.70 pg/mg tissue after the first and second withdrawals of doxycycline, respectively (Fig. 6). These concentrations of GDNF were 8.4-fold and 12.6-fold higher than the levels of endogenous murine GDNF detected in Tet-off-GFP-transplanted MPTP-treated and non-MPTP-treated mice. These data indicate that hGDNF-expressing cells infiltrate into MPTP-induced lesions in the SN and drive the production of GDNF in the absence of doxycycline. The concentration of GDNF in the SN of MPTP-treated Tet-off-hGDNF-transplanted mice decreased significantly during administration of doxycycline, however, the concentration of GDNF did not return to basal levels.
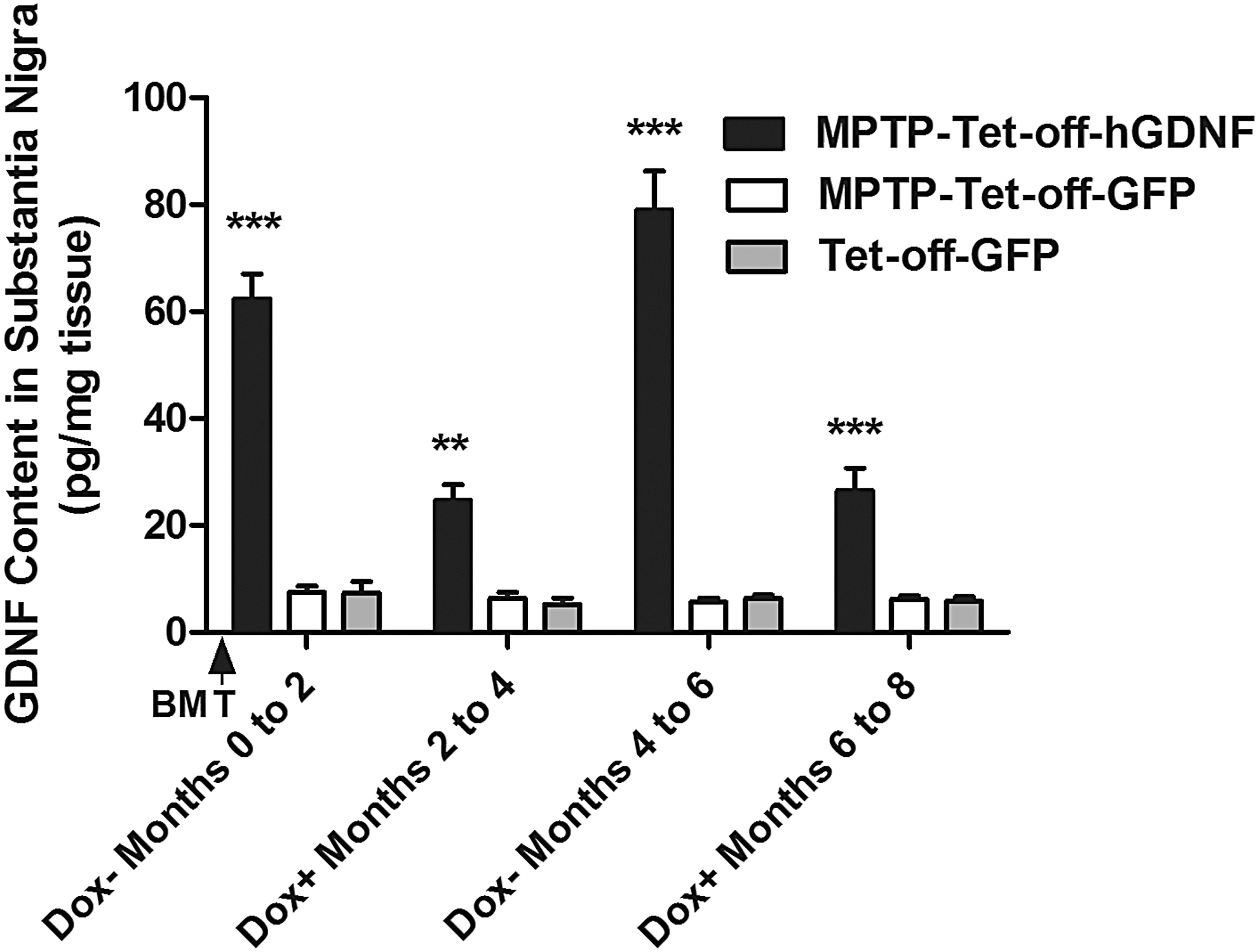
SN GDNF content responded to doxycycline following MPTP treatment. SN was dissected out using a mouse brain slicer matrix (Zivic Insruments, Pittsburgh, PA) from MPTP-treated and non-MPTP-treated mice, under Dox-withdrawn and Dox-administered (1 mg/mL in drinking water) conditions. The brain tissue samples were homogenized and GDNF concentration was measured using a GDNF Emax ImmunoAssay System (Promega, Madison, WI) according to the manufacturer's protocol. Each bar represents mean ± SEM from four brain samples per group (n = 4). **P < 0.01, ***P < 0.001 versus MPTP-Tet-off-GFP- and Tet-off-GFP- transplanted group.
Doxycycline-regulated GDNF expression did not affect body weight or food intake
We next investigated whether the body weight or food intake were affected by GDNF expression by the transplanted cells. Summarized in Fig. 7, we monitored the body weight and food intake of all mice during the Dox-regulated GDNF therapy. Overall, we observed no loss of body weight in any of the groups throughout the 8-month course of the experiments (Fig. 7A). Although food intake was variable within groups, we observed no significant differences among groups (Fig. 7B). These data indicate that body weight and food intake were not affected by either Dox-regulated GDNF expression or treatment with MPTP.
Doxycycline-regulated GDNF expression does not affect mouse body weight or food intake.
Discussion
Clinical trials of GDNF protein therapies for PD were halted in 2004 due to efficacy and safety concerns. Safety issues comprised the development of neutralizing antibodies, and an unexpected cerebellar neurodegeneration involving both Purkinjie cells and granular cell layer [40]. We previously reported that the HSCT-based macrophage-mediated GDNF delivery dramatically reduced degeneration of DA neurons in SN and their terminals in the striatum, pointing to a potentially promising therapeutic approach for PD [20,37,38]. However, this irreversible cell-based GDNF delivery raises concerns due to lack of information on the potential long-term adverse effects of sustained GDNF overexpression. Therefore, cell-based delivery of GDNF for PD patients may require regulatable vectors to enable dosage adjustment of GDNF delivery. The most regulatable system currently used in viral vectors is the tetracycline (Tet)-mediated transcriptional regulation system [39]. Several versions of the tet-regulatory system have been used to overexpress GDNF or reporter genes in vivo [41 –44]. Here, we constructed a Dox-regulated lentiviral GDNF vector, selecting a Tet-OFF system to observe the neuroprotective benefits of GDNF expression during doxycycline withdrawal. Expression of GDNF in the plasma and SN/midbrain of Tet-off-hGDNF-transplanted mice responded sensitively to both the administration (basal GDNF) and withdrawal (high GDNF) of doxycycline. Interestingly, the concentration of GDNF detected in the SN of Tet-off-hGDNF-transplanted mice was ∼10-fold higher during doxycycline withdrawal compared with basal levels of GDNF detected during doxycycline administration. However, administration of doxycycline for 2 months did not return levels of GDNF to basal levels. This sustained expression of GDNF may be due to either leaky activity of the Tet-On/Off system [45], low penetration of doxycycline through the BBB [46 –48], or their combination. The concentration of GDNF in a healthy human brain is generally low and exhibits regional differences, with undetectable concentrations in cerebrospinal fluid (<8 pg/mL), low concentrations in the cerebellum and frontal cortex (10–15 pg/mg protein), and high concentrations (∼43–70 pg/mg) in the caudate nucleus, putamen, and SN [49]. Similar levels of GDNF were detected in the SN of Tet-off-hGDNF-transplanted mice after one (62.33 ± 8.05 pg/mg) or two (78.96 ± 12.70 pg/mg) MPTP treatment cycles, suggesting that the Dox-regulated system could reversibly express GDNF at therapeutic levels required in a clinical setting.
Administration of GDNF by intraputamenal infusion in a rhesus monkey model was accompanied by acute adverse effects on treatment days, along with fluctuations in mean body weight and and intake of food and water [50]. Others have reported either a reduction in food intake or weight gain in response to GDNF in various models [51 –53]. Thus, we assessed whether alternating GDNF expression using the Dox-regulated GDNF lentiviral vector delivery was accompanied by similar adverse effects. Here, we observed a slight weight gain during the expression of GDNF in Tet-off-hGDNF mice, although this effect was not statistically significant in comparison with Tet-off-GPF mice. Although we observed fluctuations in food intake in Tet-off-hGDNF treated mice, the overall consumption of food was maintained, indicating that the influence of Dox-regulated GDNF expression or treatment with MPTP on mouse body weight and food intake is minimal.
In toxin-induced models for PD, GDNF has reproducibly displayed neuroprotective effects and improvements in motor function when delivered into dopamine-deficient striatum or SN [54 –61]. Here, we observed ∼85% DA neuron loss in MPTP-treated tet-off-GFP mice, whereas nearly half of the DA neurons were protected in mice expressing GDNF. We demonstrated that Dox-regulated GDNF expression via macrophage-mediated delivery prevented MPTP-induced motor deficits compared with non HSC-transplanted normal controls. These data agree with our previous findings wherein we showed that macrophage-mediated delivery of neurotrophic factors—either GDNF or neurturin—ameliorated MPTP-induced degeneration of DA neurons in the SN [37,38]. Moreover, others have reported that an AAV vector expressing GDNF increased locomotor performance and alleviated progressive motor dysfunction in rodent models of PD, including MitoPark mice, a genetic model [62]. However, lentiviral or AAV-mediated delivery of hGDNF failed to improve α-synuclein (α-syn) -induced motor impairment or prevent neurodegeneration in rat models [63,64]. Viral rodent models overexpressed human wild-type or mutant α-syn to levels much higher than endogenous levels (at least fourfold), resulting in blockade of the intracellular signaling response to GDNF in nigral DA neurons [19,51,52]. The blockade was caused by downregulation of transcription factor Nurr1 and downstream target GDNF receptor tyrosine kinase Ret. However, human PD data demonstrated that α-syn gene expression levels are not increased in the early-stage PD or in association with disease progression and that the majority of SN neurons contain normal levels of Nurr1 and Ret [19].
Our study showed proof of concept that donor HSCs transduced with Dox-regulated lentiviral vectors can differentiate into macrophages to express and deliver GDNF to target neurons effectively. We also demonstrated that Dox-regulated GDNF expression can be controlled via administration of doxycycline in drinking water. Dox-regulated GDNF expression was effective in protecting against MPTP-induced motor deficits and DA neuron loss in SN. These findings indicate that HSC-based macrophage-mediated Dox-regulated GDNF delivery should exert efficient long-term benefits that relieve and possibly rescue parkinsonian pathologies with minimized GDNF-based adverse effects. In these studies, transplantation of transduced HSCs was achieved via head-shielded irradiation-based conditioning to remove endogenous bone marrow cells. Head protection from irradiation would avoid inducing neuroinflammation and compromising BBB integrity, a potential confounding factor in macrophage brain infiltration-based therapies. Because the use of chemotherapeutic drugs and/or total body irradiation to sufficiently condition recipients is highly toxic, having multiple short- and long-term adverse effects [65 –70], others have designed transplantation regimens that limit toxicity by depleting endogenous cells without irradiation or chemotherapeutic drugs [71,72]. The accompanying toxicities of these new approaches are not yet fully characterized. In ongoing studies, we are developing a nontoxic mobilization-based conditioning regimen compatible with current autologous procedures (article in preparation). With multiple options to reduce conditioning becoming available in the near future, this novel approach to gene therapy may eventually increase clinical feasibility and acceptance for the treatment of PD in the future.
Footnotes
Acknowledgments
We thank Ms. Qiong Zhang for excellent animal care work and assistance with behavioral testing. This work was supported by Merit Review Award [2I01BX000737] from the United States (U.S.) Department of Veterans Affairs Biomedical Laboratory Research and Development Service and the William and Ella Owens Medical Research Foundation to SL. The study was also partially supported by Guizhou Major Basic Research Projects ([2014]6008) and Guizhou Major Science and Technology Research Projects ([2015]2003) and the Technology Department of Guizhou Province of China (SY-2014-3024, J-2015-2012).
Author Disclosure Statement
No competing financial interests exist.