
Abstract
Adipose-derived stem cells (ADSCs) are a type of multipotent mesenchymal stem cells with immunosuppressive capacities. However, the underlying mechanisms involved in the inhibitory effects of ADSCs on T cells are not completely elucidated. In this study, human peripheral blood mononuclear cells (PBMCs) stimulated with anti-CD3/CD28 antibody-coated beads were cultured with or without allogeneic ADSCs (ADSC-to-PBMC ratio, 1:5). Surface marker levels, violet-labeled cell proliferation, apoptosis, interferon-gamma (IFN-gamma) production, and nuclear factor-kappaB (NF-kappaB) phosphorylation of CD4+ and CD8+ T cells were detected using flow cytometry. It was observed that ADSCs significantly suppressed the proliferation and IFN-gamma production but enhanced apoptosis of both CD4+ and CD8+ T cells in T cell receptor (TCR)-stimulated PBMCs. The expressions of programmed death-ligand 1 (PD-L1) and galectin 9 (Gal-9) on ADSCs were significantly upregulated and induced during coculture with PBMCs. TCR-stimulated CD4+ and CD8+ T cells cultured with ADSCs had higher expression levels of programmed death-1 (PD-1) and T cell immunoglobulin and mucin-containing protein-3 (TIM-3) than those in cells cultured without ADSCs. Moreover, the suppressive effects of ADSCs on T cells in terms of proliferation and IFN-gamma production were significantly reversed in the presence of anti-PD-L1 and anti-Gal-9 antibodies. Importantly, the phosphorylation of NF-kappaB in CD4+ and CD8+ T cells cocultured with ADSCs was significantly inhibited, and this inhibition was significantly attenuated via the PD-L1 and Gal-9 blockades. In conclusion, human ADSCs perform immunoregulatory functions partially through the inhibition of NF-kappaB activation in T cells via the PD-L1/PD-1 and Gal-9/TIM-3 pathways, which provide new insights into the mechanism of human ADSC-mediated immunomodulation.
Introduction
A
However, the mechanisms underlying these activities remain unclear. The number of reports on the inhibition of proliferation of total T cells and peripheral blood mononuclear cells (PBMCs) by human ADSCs have been increasing [1,12,13], whereas related studies on T cell subsets are scant [14]. Furthermore, there are no reports on the role of human ADSCs in interferon-gamma (IFN-gamma) production and apoptosis of CD4+ and CD8+ T cells. Therefore, it is necessary to investigate the immunoregulatory effects of ADSCs on T cell subsets and the related underlying molecular mechanisms.
Several factors involved in the immunosuppressive mechanisms of human ADSCs have been reported, including indoleamine 2,3-dioxygenase (IDO) [2], interleukin-6 (IL-6), transforming growth factor β1 (TGF-β1) [11], and prostaglandin E2 [13], in addition to cell contact-dependent mechanisms. However, the inhibitors or neutralizing antibodies of IDO, IL-6, and TGF-β have failed to completely prevent T cell suppression by human ADSCs [15], suggesting that there might be other factors contributing to immunosuppression.
Programmed death-ligand 1 (PD-L1) and galectin 9 (Gal-9), which are two important inhibitory molecules, suppress the T cell immune response by binding programmed death-1 (PD-1) and T cell immunoglobulin and mucin-containing protein-3 (TIM-3), respectively [16,17]. Studies on human placenta MSCs (PMSCs), umbilical cord blood MSCs (UCB-MSCs), and BM-MSCs have found that the upregulation of constitutively expressed PD-L1 mediates a suppressive effect on T cells [18 –21]. Gal-9 is also involved in immunosuppressive function of human BM-MSCs [22,23]. However, little is known about the expression and roles of PD-L1 and Gal-9 in the immunosuppressive properties of human ADSCs [24].
Nuclear factor-kappaB (NF-kappaB) is a major transcription factor. On activation, it enters the nucleus to control differentiation, proliferation, cytokine production, and T cell apoptosis [25,26]. The ligation of PD-1 with PD-L1 and TIM-3 with Gal-9 transmits negative signals and plays an inhibitory role in T cells [16,17,27]. Therefore, we hypothesized that human ADSCs exert an immunosuppressive effect on T lymphocytes through the inhibition of NF-kappaB activation mediated by PD-L1/PD-1 and/or Gal-9/TIM-3 pathways.
In this study, we investigated the immunosuppressive effects of human ADSCs on human CD4+ and CD8+ T cells in T cell receptor (TCR)-stimulated PBMCs. We further investigated the expression and role of PD-L1 and Gal-9 on ADSCs and the mechanism underlying ADSC-mediated immunomodulation. The results in this study may have clinical implications for the development of rational strategies to delicately manipulate the immunomodulatory properties of ADSCs on T cells.
Materials and Methods
Generation of human ADSCs
Subcutaneous adipose tissues of 10 healthy female donors aged 37.07 ± 10.5 (mean ± SEM) years were obtained by surgical liposuction and processed as described in the following brief procedures [11,28]. The fat block was cut into 1–2-mm3 pieces and digested with an equal volume of 0.2% collagenase type I (Sigma) at 37°C for 45 min. After digestion, the cell suspension was centrifuged, and the pellets were cultured in basal medium [low-glucose Dulbecco's modified Eagle's medium (Hyclone), 20% fetal bovine serum (FBS; Biological Industries, Israel), 20 mM L-glutamine, and 100 U/mL penicillin/streptomycin (Gibco)] at 37°C with 5% CO2. Nonadherent cells were removed by changing the medium after 4 days. Fresh basal medium was supplemented once every 3 days. At 80%–90% confluence, the cells were digested with 0.25% trypsin and passaged at a ratio of 1:3. The initial passage was defined as passage 0. The cells of passages 3–5 were used for all the experiments in this study. The Ethical Review Board of the First Hospital of China Medical University approved the informed consent forms and study protocols. Informed consent was obtained from all donors.
Phenotyping of ADSCs
To characterize the phenotypes of ADSCs [29,30], the cells (passage 3) were stained with APC-Cy7-conjugated anti-CD10, PE-conjugated anti-CD13, PE-Cy7-conjugated anti-CD49d, and FITC-conjugated anti-lineage 1, including CD3, CD14, CD16, CD19, CD20, and CD56 (BD Biosciences), APC-conjugated anti-CD29, FITC-conjugated anti-CD90, PE-conjugated anti-CD44, PE-conjugated anti-CD34, APC-conjugated anti-CD31, PerCP-conjugated anti-CD45, PE-conjugated anti-HLA-DR (BioLegend), and their appropriate isotype controls. For the detection of PD-L1 and Gal-9 expression on ADSCs, APC-conjugated anti-PD-L1 and PerCP-conjugated anti-Gal-9 (BioLegend) were used. The stained cells were analyzed using flow cytometry (LSR-II; BD Biosciences) and FlowJo software (TreeStar, San Carlos, CA).
Differentiation potential assay
The differentiation potentials of cells (passage 3) were investigated using MSCgo™ adipogenic, osteogenic, and chondrogenic medium (Biological Industries) according to the manufacturer's instructions [31]. Briefly, the cells were seeded onto 48-well plates at 3,000 cells/cm2 in basal medium. When the cells reached 80% confluence, the medium was replaced with complete adipogenic and osteogenic medium for the induction of adipogenesis and osteogenesis, respectively, and it was changed every 2–3 days for 7–14 days. For the induction of chondrogenesis, the medium was replaced with complete chondrogenic medium when the cells reached 100% confluence and was changed every 3–4 days for 14–21 days. The cells were stained using oil red O (Sigma) for the detection of adipocytes, using NBT/BCIP alkaline phosphatase (Sangon Biotech, Shanghai) and alizarin red S (Santa Cruz Biotechnology) for the detection of osteoblasts, and using toluidine blue O (Santa Cruz Biotechnology) for the detection of chondrocytes.
Coculture of ADSCs and PBMCs
Human PBMCs were isolated from EDTA-coagulated blood of healthy adults using Ficoll centrifugation. PBMCs were cocultured with or without ADSCs in RPMI-1640 medium (Hycolone), containing 10% FBS and in the presence or absence of anti-CD3 and anti-CD28 antibody-coated beads (Gibco). The ratio of ADSCs to PBMCs was 1:5, and that of beads-to-cells was 1:1 as recommended by the manufacturers. The cells were cultured in 24-well plates for 4 days for T cell proliferation and apoptosis assay and for 2 days for IFN-gamma production and PD-1 and TIM-3 expression assays. For blocking experiments, anti-PD-L1 antibody (clone: 29E.2A3, 10 μg/mL; BioLegend), anti-Gal-9 antibody (clone: 9M1–3, 10 μg/mL; BioLegend), and appropriate isotype controls were added to the cultures at the beginning of the appropriate experiments. For transwell experiments, ADSCs were seeded in 6.5 mm transwells with 0.4 μm pore size (Corning). The ratio of ADSCs to PBMCs was 1:5, and that of beads-to-cells was 1:1 as recommended by the manufacturers.
T cell proliferation
The CellTrace™ Violet Cell Proliferation Kit (Invitrogen, C34557) was used according to the manufacturer's instructions. Briefly, after 4 days of coculture, violet-labeled PBMCs were stained with PE-Cy7-conjugated anti-CD3, APC-Cy7-conjugated anti-CD4, and FITC-conjugated anti-CD8 (BioLegend) antibodies. Dead cells were excluded by adding 7-aminoactinomycin D (7-AAD; BD Biosciences) before the samples were analyzed. T cell proliferation was measured using BD LSR-II flow cytometer.
T cell apoptosis
The PE-Annexin V/7-AAD staining kit (BD Bioscience, 559763) was used according to the manufacturer's instructions. Briefly, the cocultured PBMCs were collected and stained with PE-Cy7-conjugated anti-CD3, APC-Cy7-conjugated anti-CD4, APC-conjugated anti-CD8, PE-conjugated annexin V, and 7-AAD and measured using BD LSR-II flow cytometer.
IFN-gamma production by T cells
At the end of the culture, PBMCs were stained with PE-Cy7-conjugated anti-CD3, APC-Cy7-conjugated anti-CD4, and FITC-conjugated anti-CD8 antibodies, followed by intracellular staining with APC-conjugated anti-IFN-gamma (BD Biosciences) antibodies, and analyzed using LSR-II flow cytometer. For the detection of PD-1 and TIM-3 expression on T cells, PBMCs were stained with PerCP-conjugated anti-CD3, APC-Cy7-conjugated anti-CD4 (BD Biosciences), FITC-conjugated anti-CD8, PE-Cy7-conjugated anti-PD-1, and PE-conjugated anti-TIM-3 (BioLegend) antibodies.
Phosphorylation of NF-kappaB and AKT in T cells
To detect the phosphorylation of NF-kappaB (p65) and protein kinase B (AKT), cells were treated with Phosflow kits according to the manufacturer's instructions. After surface staining with PerCP-conjugated anti-CD3, APC-Cy7-conjugated anti-CD4, and FITC-conjugated anti-CD8 antibodies, the cells were fixed by adding 100 μL of fixation buffer and incubated at room temperature for 30 min. After washing twice with permeabilization buffer (eBioScience), the cells were stained with PE-conjugated antibodies of anti-Phospho-NF-kappaB p65 (Ser529; eBioscience, 12-9863-42) or anti-Phospho-AKT (Ser473; Cell Signaling Technology, 5315) for 30 min at room temperature and analyzed using flow cytometry.
Statistical analysis
SPSS 17.0 software (SPSS, Inc., Chicago, IL) and Graphpad Prism 7.0 were used to conduct statistical analyses. Comparisons of proliferation, apoptosis, and IFN-gamma production, if applicable, in the presence or absence of ADSCs and anti-PD-L1 and anti-Gal-9 blockades were performed using the paired t-test. Spearman's correlation analysis was used to analyze the correlation between the corresponding parameters, if applicable. A P value <0.05 was considered statistically significant.
Results
Characterization of ADSCs
Human ADSCs were produced by adherent culture. We then detected cell surface markers to determine the characteristics of these cultured cells. As shown in Fig. 1A, the harvested cells of passage 3 expressed high levels of CD10, CD13, CD49d, CD29, CD90, and CD44, but did not express CD34, CD31, CD45, HLA-DR, and lineage 1, including CD3, CD14, CD16, CD19, CD20, and CD56. Importantly, these cells efficiently differentiated into adipocytes, osteoblasts, and chondrocytes in the respective culture media (Fig. 1B–E), which is consistent with previous reports [11]. These observations indicated that both phenotypes and the differentiation capacities of the harvested cells adequately met the criteria for ADSCs proposed by the International Society for Cellular Therapy (ISCT) [29,30]. As shown, not all the ADSCs were CD29, CD90, or CD44 positive. The median percentage of the positive population was 82.3% for CD29 (range: 81.4%–98.1%), 84.9% for CD90 (range: 79.1%–97.6%), and 81.2% for CD44 (range: 80.2%–95.5%) (Supplementary Table S1; Supplementary Data are available online at

Characterization of ADSCs.
ADSCs suppress the proliferation of CD4+ and CD8+ T cells
We detected the proliferation of CD4+ and CD8+ T cells in TCR-stimulated PBMCs (bead-to-cell ratio, 1:1) after 4 days of coculture with ADSCs (ADSC-to-PBMC ratio, 1:5) using CellTrace violet-labeled assay and flow cytometry. The percentage of violet-positive CD4+ and CD8+ T cells in PBMCs cocultured with ADSCs significantly decreased compared with PBMCs cultured without ADSCs (both P < 0.01, Fig. 2A–D). However, no significant difference in the proliferation between CD4+ and CD8+ T cells was observed (Supplementary Fig. S1A), indicating that human ADSCs inhibited the proliferation of CD4+ and CD8+ T cells in vitro without significant difference.
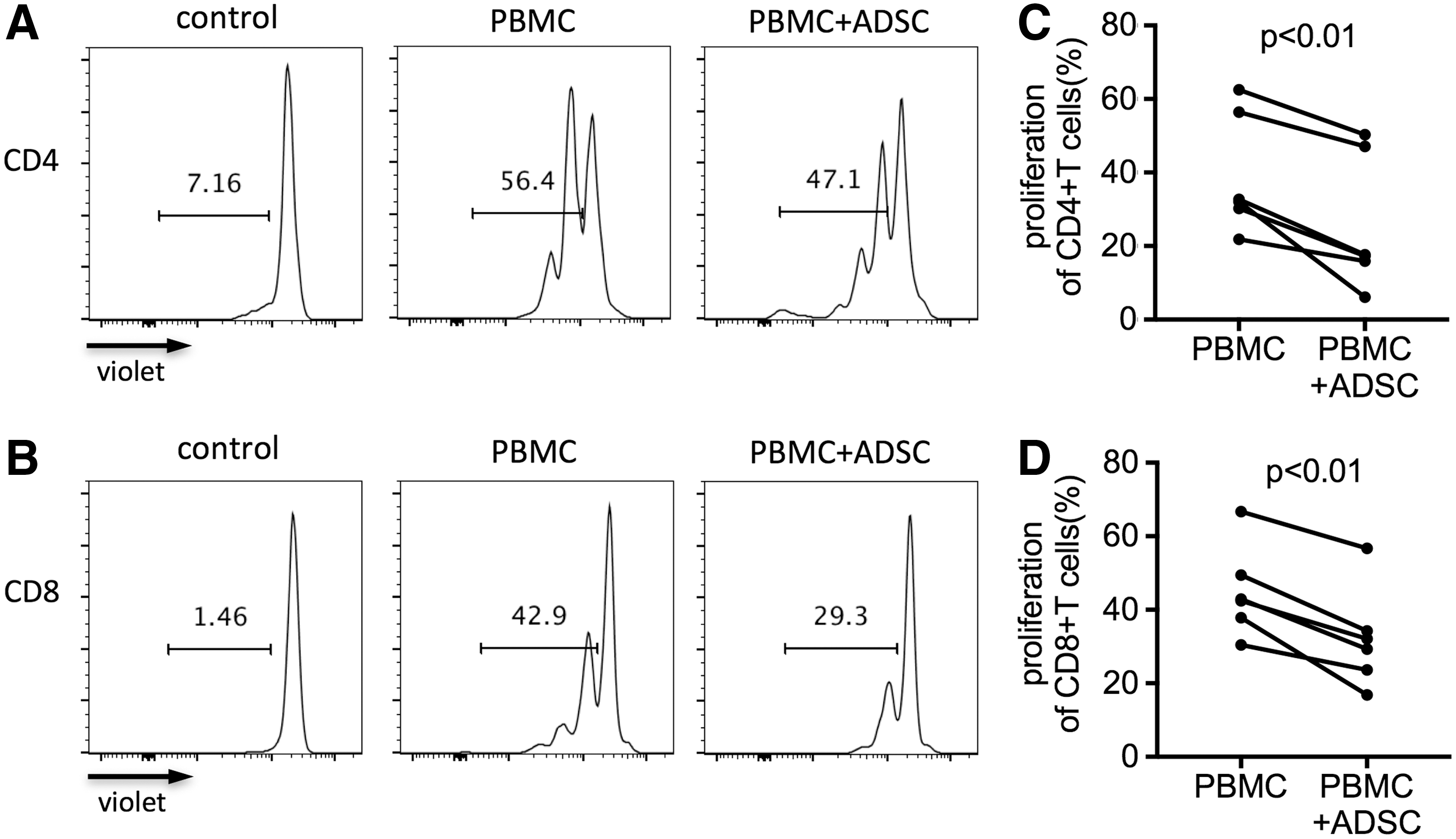
ADSC-mediated inhibition of TCR-stimulated CD4+ and CD8+ T cell proliferation. Freshly isolated PBMCs were labeled with CellTrace violet reagents and stimulated for 4 days with anti-CD3/CD28 beads in the presence or absence of allogeneic ADSCs. The beads-to-cells ratio was 1:1 and ADSC-to-PBMC ratio was 1:5. The unstimulated PBMCs were used as control.
ADSCs suppress IFN-gamma production by CD4+ and CD8+ T cells
IFN-gamma production is one of the functional characteristics of CD4+ and CD8+ T cells [32]. To determine the potential effect of human ADSCs on IFN-gamma production by T cell subsets, we used intracellular staining and flow cytometry to detect IFN-gamma production in TCR-stimulated PBMCs after coculture with ADSCs for 2 days in 24-well plates. The percentage of IFN-gamma-producing CD4+ T cells was significantly reduced in PBMCs cocultured with ADSCs (P < 0.05, Fig. 3A, C), similar to that in IFN-gamma-producing CD8+ T cells (P < 0.05, Fig. 3B, D). Moreover, the decreased IFN-gamma production in CD8+ T cells was significantly higher than that in CD4+ T cells (P < 0.05, Supplementary Fig. S1B). These results indicated that ADSCs mediated a more potent immunosuppression on CD8+ T cells than on CD4+ T cells in IFN-gamma production, suggesting that ADSCs might be more useful for the immune disorders characterized by the abnormal increase in IFN-gamma production by CD8+ T cells.
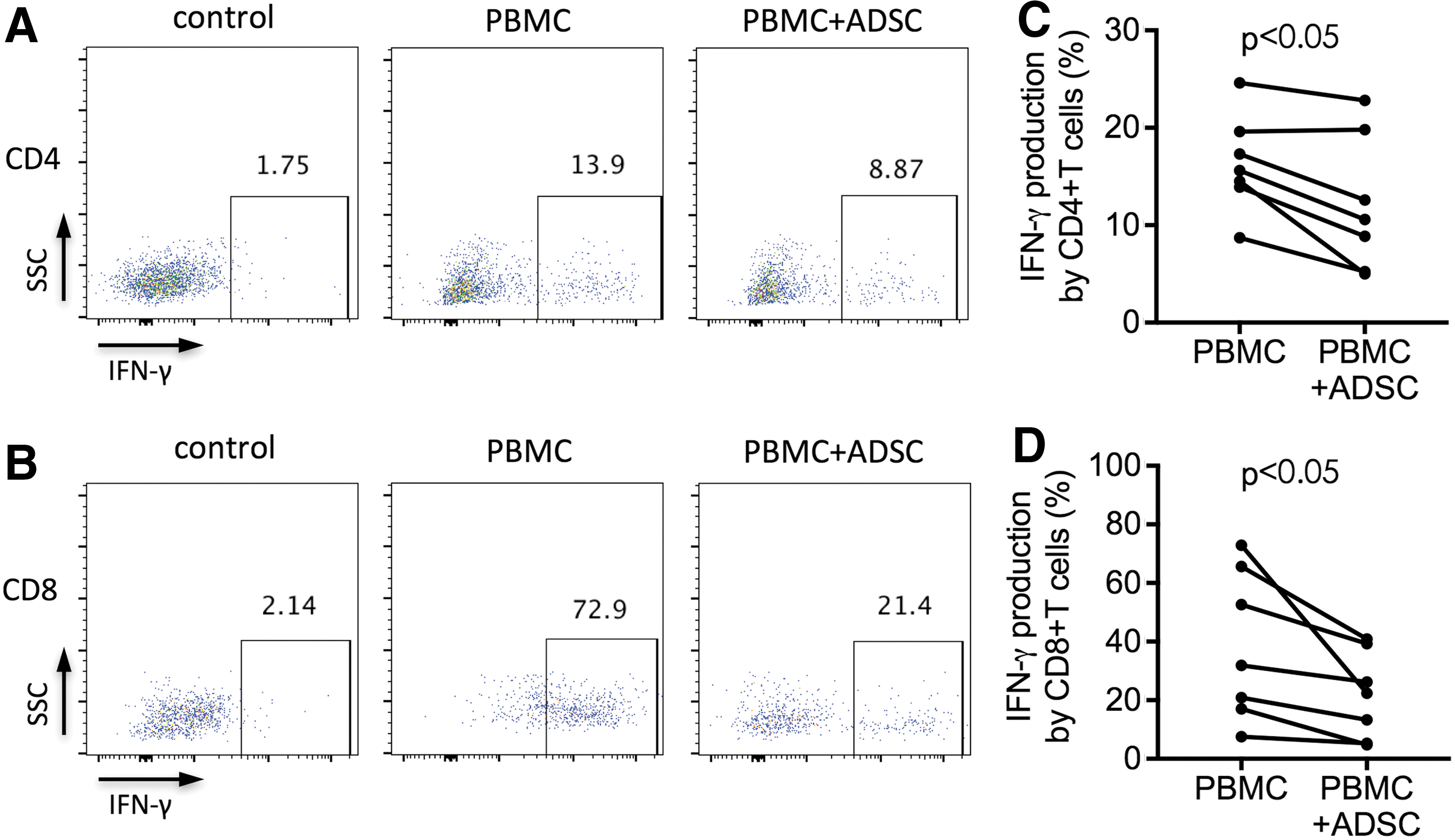
ADSC-mediated inhibition of IFN-gamma production by TCR-stimulated CD4+ T cells and CD8+ T cells. Freshly isolated PBMCs were incubated for 2 days with anti-CD3/CD28 beads at a ratio of 1:1 either alone or with allogeneic ADSCs at a ratio of 1:5 (ADSC-to-PBMC). Cells were intracellularly stained with APC-conjugated anti-IFN-gamma antibodies. The unstimulated PBMCs were used as control.
ADSCs enhance the apoptosis of CD4+ and CD8+ T cells
To determine whether the induction of apoptosis contributes to the suppressive effect of ADSCs on T cell subsets, we investigated the apoptosis of TCR-stimulated T cells in PBMCs after coculturing with ADSCs for 4 days. We found that the percentages of annexin V+ and 7-AAD+ CD4+ T cells in coculture significantly increased compared with that of PBMC monoculture (both P < 0.01, Fig. 4A, C, D). We also found that the percentages of annexin V+ and 7-AAD+ CD8+ T cells in PBMCs cocultured with ADSCs significantly increased (both P < 0.01, Fig. 4B, E, F). However, no significant difference in apoptosis between CD4+ and CD8+ T cells was observed (Supplementary Fig. S1C). These findings indicated that ADSCs had the capability to equally enhance the apoptosis of TCR-activated T cells, suggesting that the modulatory effects of ADSCs on T cells may be involved in the induction of apoptosis mediated by ADSCs. To provide more information for the selection of ADSCs in the clinic, we analyzed the correlation among the levels of immunosuppression. The results showed that there was no significant correlation among the suppression of T cell proliferation, the inhibition of IFN-gamma production, and the induction of apoptosis (Supplementary Fig. S2). Nevertheless, the nonparalleled relationship among the levels of immunosuppression of ADSCs should be of concern when the ADSCs are considered to be the candidates for therapeutic use in immune disorders.
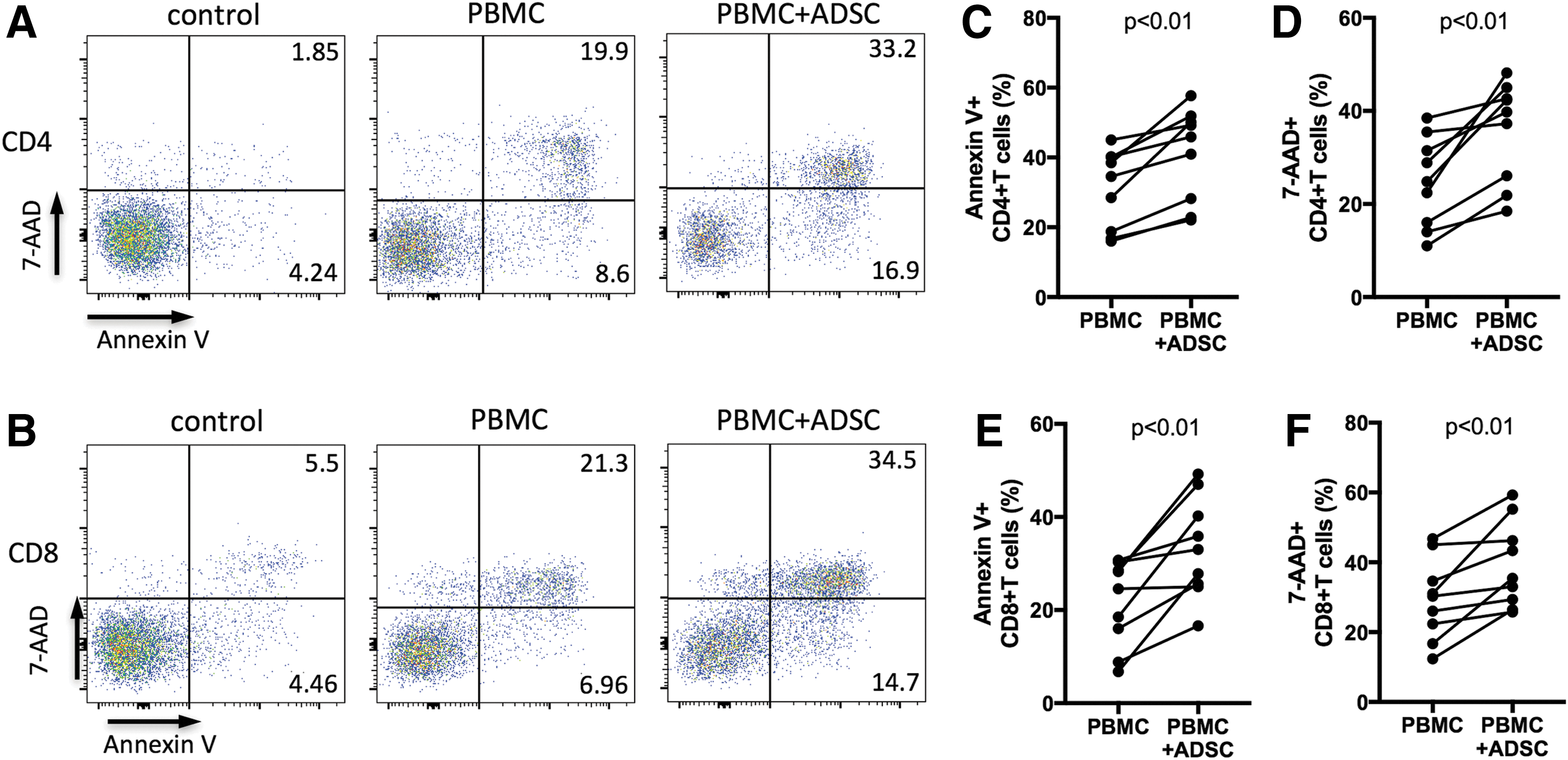
ADSC-mediated increase in apoptosis of TCR-stimulated CD4+ and CD8+ T cells. Freshly isolated PBMCs were stimulated with anti-CD3/CD28 beads at a ratio of 1:1, with or without allogeneic ADSCs at a ratio of 1:5 (ADSC-to-PBMC). After 4 days, cells were stained with PE-conjugated annexin V and 7-AAD. The unstimulated PBMCs were used as control.
Enhanced expression of PD-L1 and Gal-9 on ADSCs and PD-1 and TIM-3 on CD4+ and CD8+ T cells in ADSC and PBMC coculture
Current knowledge of the mechanisms used by ADSCs to inhibit T cells has recently expanded [33 –35]. PD-L1 and Gal-9 are coinhibitory molecules whose roles in the modulatory mechanisms of human PMSCs, BM-MSCs, and UCB-MSCs have been reported [18,20 –23,36]. Moreover, IFN-gamma pretreatment significantly induced PD-L1 expression on human ADSCs [24,37]. It is unclear whether ADSCs play immunosuppressive roles through PD-L1 and Gal-9. Thus, we measured the expression of PD-L1 and Gal-9 on ADSCs and that of their receptors PD-1 and TIM-3, respectively, on T cells in coculture of ADSCs with TCR-stimulated or nonstimulated PBMCs (ADSC-to-PBMC ratio, 1:5). We found that the percentage of ADSCs expressing PD-L1 and Gal-9 significantly increased after coculture with TCR-stimulated PBMCs for 2 days (both P < 0.01, Fig. 5A–D) compared with ADSCs cocultured with nonstimulated PBMCs. So did the mean fluorescence intensity (MFI) of PD-L1 and Gal-9 expression by ADSCs (P < 0.001 and P < 0.01, respectively; Supplementary Fig. S3A, B). The increased expression of PD-L1 and Gal-9 on ADSCs was not statistically correlated to the ADSC-mediated immunosuppression on T cells (data not shown). Furthermore, the percentage of PD-1-expressing CD4+ T cells in nonstimulated and TCR-stimulated PBMCs cocultured with ADSCs was significantly increased (both P < 0.05, Fig. 6A, B) compared with PBMCs cultured without ADSCs. Similarly, the percentage of PD-1-expressing CD8+ T cells in nonstimulated and TCR-stimulated PBMCs cocultured with ADSCs was significantly increased (both P < 0.05, Fig. 6C, D). Furthermore, the nonstimulated CD4+ and CD8+ T cells expressed very low levels of TIM-3, whereas TCR-stimulation alone could increase TIM-3 expression. This finding was similar to those in previous studies [38]. Surprisingly and interestingly, we found that coculture with ADSCs significantly increased the frequencies of TIM-3-expressing CD4+ T cells in both nonstimulated and TCR-stimulated PBMCs (both P < 0.05 and P < 0.05, respectively; Fig. 6E, F). The percentage of TIM-3-expressing CD8+ T cells also increased in these cell populations (P < 0.05 and P < 0.05, respectively; Fig. 6G, H). Meanwhile, we also found a significant increase in MFI of PD-1 and TIM-3 expressed by T cells (Supplementary Fig. S4A–H). Taken together, our findings indicated that ADSC–PBMC coculture increased the expression of PD-L1 and Gal-9 on ADSCs and that of PD-1 and TIM-3 on CD4+ and CD8+ T cells, suggesting that PD-L1/PD-1 and Gal-9/TIM-3 pathways may play a role in ADSC-induced T cell inhibition.
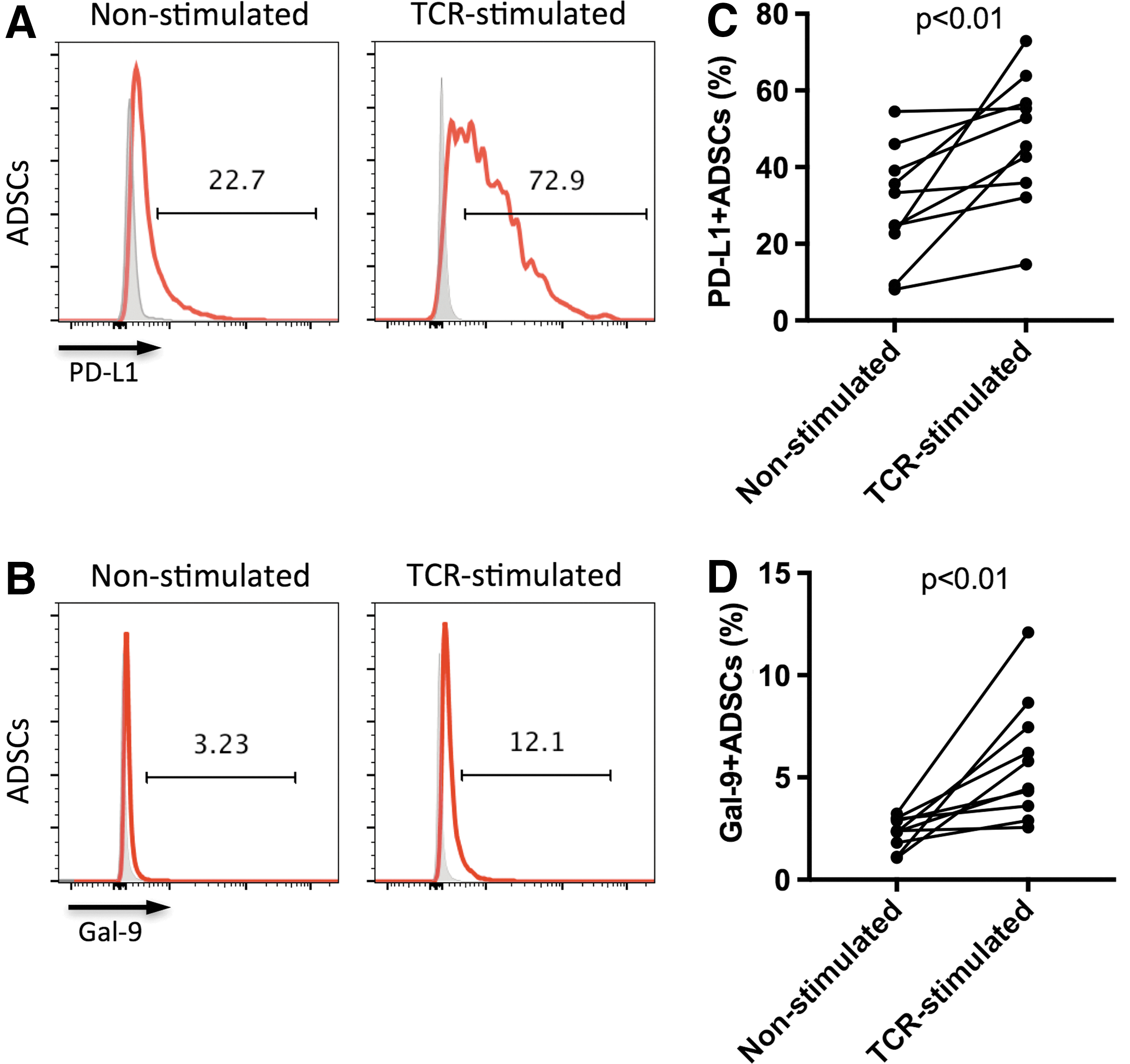
Increased expression of PD-L1 and Gal-9 on ADSCs in coculture of ADSCs and PBMCs. ADSCs were cocultured with freshly isolated PBMCs (ADSC-to-PBMC ratio 1:5) in the presence or absence of anti-CD3/CD28 beads (beads-to-cells ratio 1:1). After 2 days, PD-L1 and Gal-9 expressions by ADSCs cultured with nonstimulated PBMCs and those cultured with TCR-stimulated PBMCs were detected using flow cytometry.
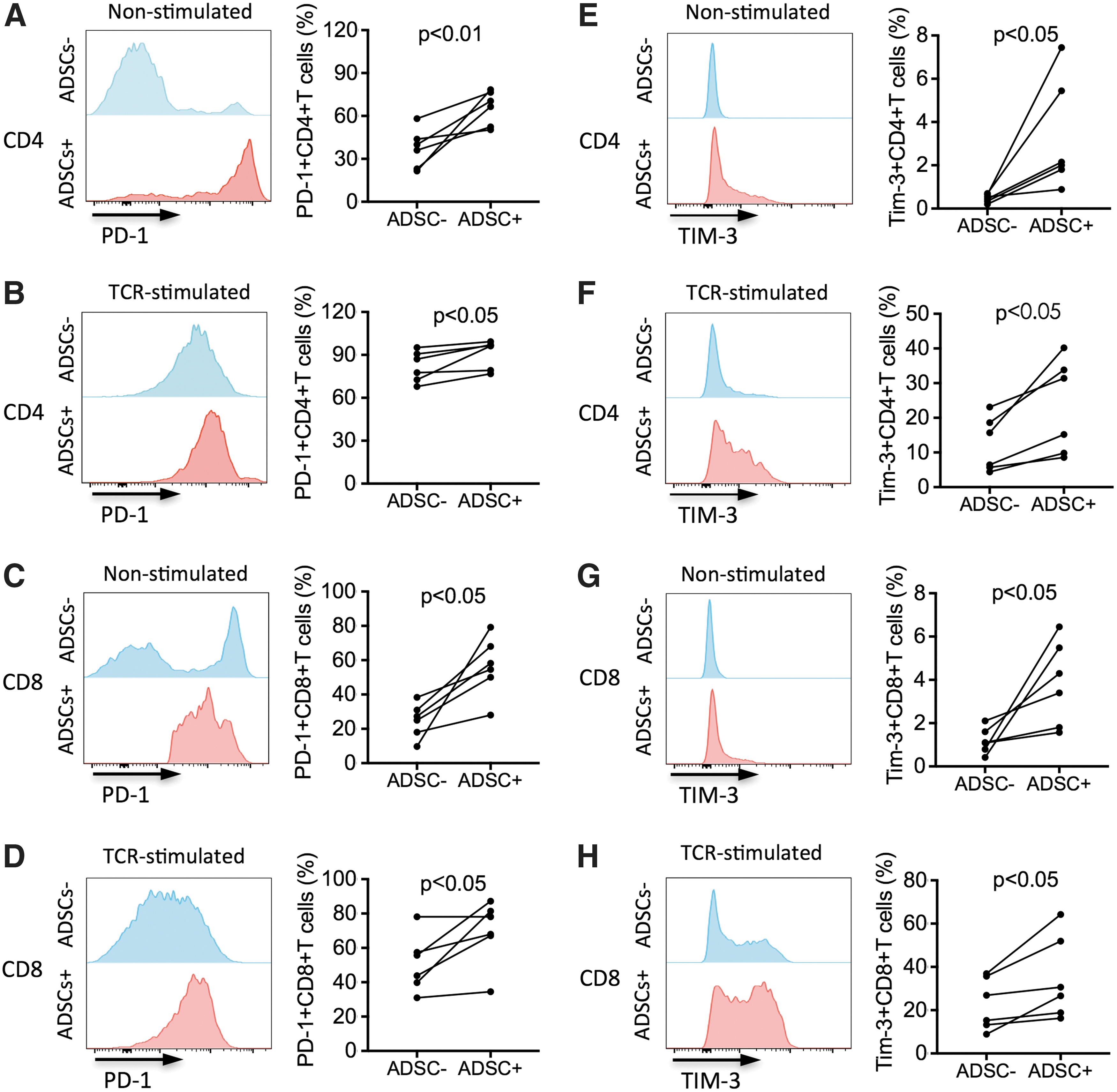
Increased expression of PD-1 and TIM-3 on T cells in coculture of ADSCs and PBMCs. ADSCs were cocultured with freshly isolated PBMCs (ADSC-to-PBMC ratio 1:5) in the presence or absence of anti-CD3/CD28 beads (beads-to-cells ratio 1:1).
Blocking PD-L1/PD-1 and Gal-9/TIM-3 pathways decreases the suppressive activity of ADSCs
To determine whether the upregulated expression of PD-1 and TIM-3 on T cells via interaction with ADSCs was involved in ADSC-mediated immunosuppresion, we further investigated the impact of PD-L1 and Gal-9 blockades on the inhibitory effect of ADSCs. Blocking antibodies for PD-L1 and Gal-9 and control antibodies were added to the cell culture. We found that compared with isotype controls, either PD-L1 or Gal-9 blocking antibodies significantly reversed the inhibitory effects of ADSCs on the proliferation (P < 0.01 and P < 0.05, respectively; Fig. 7A, B) and IFN-gamma secretion (both P < 0.05, Fig. 7E, F) by CD4+ T cells cocultured with ADSCs. Correspondingly, PD-L1 and Gal-9 blockades significantly increased the proliferation (both P < 0.05, Fig. 7C, D) and IFN-gamma secretion (P < 0.01 and P < 0.05, respectively; Fig. 7G, H) of ADSC-inhibited CD8+ T cells. These results indicated the critical role of the PD-L1/PD-1 and Gal-9/TIM-3 pathways in the immunomodulatory functions of ADSCs. Since we demonstrated that both PD-L1/PD-1 and Gal-9/TIM-3 pathways appeared to be involved in the immunosuppression of T cells, we compared the two pathways to determine which pathway was more predominant in this process. Our analysis revealed that there was no significant difference in the T cell proliferation (or IFN-gamma production) between PD-L1 and Gal-9 blockade (Supplementary Fig. S5). These results indicated that both PD-L1/PD-1 and Gal-9/TIM-3 pathways contributed to the ADSC-mediated inhibition of T cells without detectable difference.
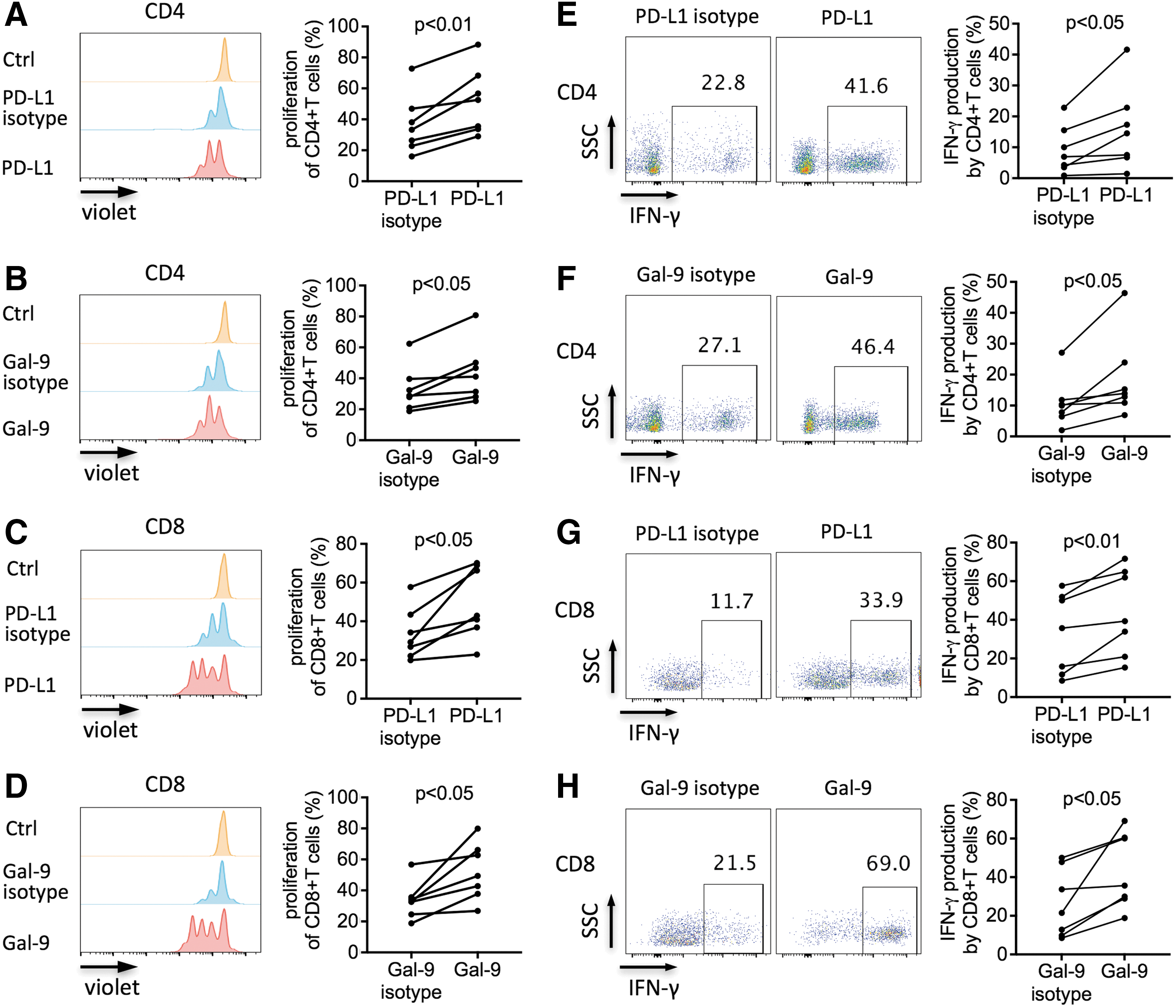
Increased proliferation and IFN-gamma production by ADSC-inhibited CD4+ and CD8+ T cells via the PD-L1 and Gal-9 blockades.
ADSCs modulate NF-kappaB activity in human T cells via PD-L1/PD-1 and Gal-9/TIM-3 pathways
Because the inhibition of NF-kappaB activation induces T cell inhibition [39], we speculated that modulation of NF-kappaB activity might be the mechanism by which ADSCs exerted suppressive effects via PD-L1/PD-1 and Gal-9/TIM-3 interactions. To verify this hypothesis, we used the Phosflow assay and flow cytometry to detect the phosphorylation of NF-kappaB (p65) in CD4+ and CD8+ T cells. We demonstrated that p65 phosphorylation in these T cell subsets in TCR-stimulated PBMCs cocultured with ADSCs was significantly decreased compared with that in PBMCs cultured alone (both P < 0.05, Fig. 8A–D), indicating that ADSCs regulated NF-kappaB activation of T cell subsets in TCR-stimulated PBMCs. To determine whether PD-L1 and Gal-9 were involved in the ADSC-induced suppression of NF-kappaB activity in T cells, we cultured TCR-stimulated T cells with ADSCs in the presence or absence of anti-PD-L1 (10 μg/mL) or anti-Gal-9 (10 μg/mL) antibodies. After 3 h of coculture, we detected NF-kappaB (p65)-positive T cells. We found that the proportion of NF-kappaB (p65)-positive CD4+ and CD8+ T cells was significantly increased in the presence of anti-PD-L1 (both P < 0.05, Fig. 8E, F) and anti-Gal-9 antibodies (both P < 0.01, Fig. 8G, H). These results suggested that both PD-L1/PD-1 and Gal-9/TIM-3 pathways were involved in ADSC-induced suppression of NF-kappaB activation in TCR-stimulated CD4+ and CD8+ T cells.
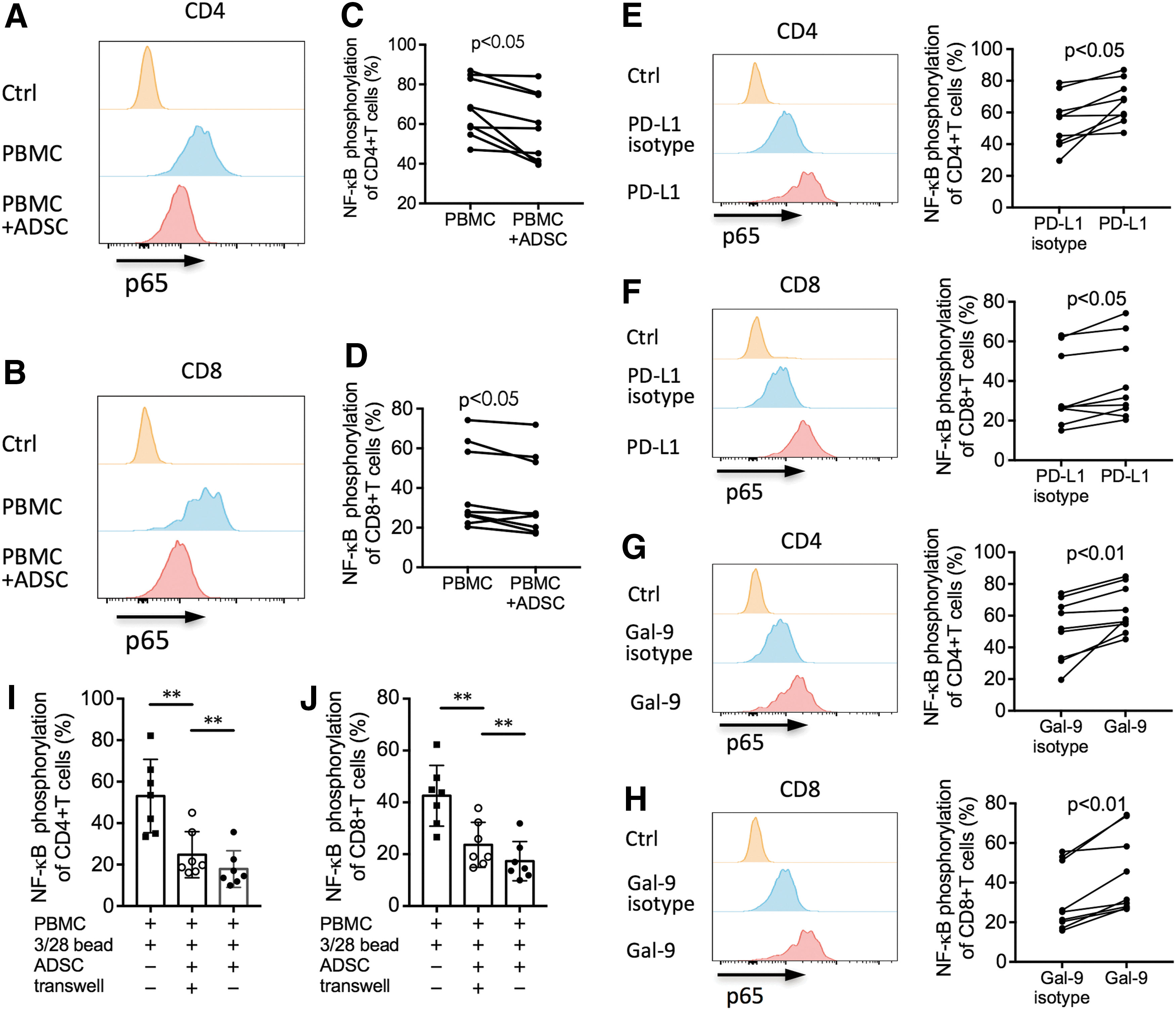
Reversion of NF-kappaB phosphorylation in ADSC-inhibited CD4+ and CD8+ T cells via the PD-L1 and Gal-9 blockades. Freshly isolated PBMCs were stimulated with anti-CD3/CD28 beads at a ratio of 1:1, with or without allogeneic ADSCs at a ratio of 1:5 (ADSC-to-PBMC). For each blocking experiment, anti-PD-L1 antibody, anti-Gal-9 antibody, or their appropriate control antibodies were added to the culture at the beginning of the experiment. After 30 min, cells were intracellularly stained with PE-conjugated anti-p65 antibodies.
Since the two pathways, PD-L1/PD-1 and Gal-9/TIM-3, appeared to be involved in the immunosuppressive activities of ADSCs through the regulation of the NF-kappaB signaling pathway, it should be worthy to address the difference in the level of p65 phosphorylation between these two inhibitory pathways. We found that the expression of phosphorylated p65 by CD4+ and CD8+ T cells, in terms of both the positive percentage and MFI, was significantly higher in the presence of anti-PD-L1 antibody compared with anti-Gal-9 antibody (all P < 0.01, Supplementary Fig. S6A–D). When both antibodies were added, the level of p65 phosphorylation in CD4+ and CD8+ T cells was similar to that in the presence of anti-PD-L1 antibody alone, but significantly higher than that in the presence of anti-Gal-9 antibody alone (all P < 0.01, Supplementary Fig. S6A–D). These findings indicated that the PD-L1/PD-1 pathway had more potent inhibitory capability than Gal-9/TIM-3 pathway did. There was no additive effect when both blocking antibodies were used together.
To investigate the effect of the inflammatory milieu on ADSC-suppressed NF-kappaB activation in TCR-stimulated T cells, we performed transwell experiments. TCR-stimulated PBMCs were cultured with ADSCs in the transwell plates to avoid the T cell contact with ADSCs. We found that, compared to TCR-stimulated T cells without ADSCs, a significant inhibition of p65 phosphorylation in T cells was observed when T cells were cultured with ADSCs in transwell system (both P < 0.01, Fig. 8I, J). In the nontranswell plates that allow the T cell contact with ADSCs, the inhibited p65 phosphorylation in T cells was further decreased compared with that in transwell plates (both P < 0.01, Fig. 8I, J). These results indicated that both soluble factors in the culture milieu and the cell–cell contact mechanism were involved in the suppression of NF-kappaB activation in T cells mediated by ADSCs.
To further explore how PD-L1/PD-1 and Gal-9/TIM-3 interferes with NF-kappaB pathway, we performed experiments to investigate the role of AKT, which is an upstream molecule participating in NF-kappaB activation, in the relation between PD-L1/Gal-9 and NF-kappaB [40]. We found a significant reduction of phosphorylated AKT in T cells mediated by ADSCs (both P < 0.01, Supplementary Fig. S7A, B). Blocking assays showed that the phosphorylation of the AKT kinase was recovered in the presence of blocking antibodies of either PD-L1 or Gal-9 (all P < 0.01, Supplementary Fig. S7C–F). Our finding was consistent with the report that PD-1 signaling inhibited the TCR-mediated activation of the AKT kinase in T cells [41]. These results suggested that the inhibition of AKT phosphorylation may be one of the mechanisms underlying the effects of PD-L1/PD-1 and Gal-9/TIM-3 on NF-kappaB signaling in T cells.
Discussion
While most previous studies have illustrated the suppressive modulation of ADSCs on total T cells or PBMCs [2,11,13,24,37], we aimed to confirm their inhibitory effect on the proliferation of T cell subsets, which was also observed in human BM-MSCs and Wharton's Jelly MSCs [14,42]. To further evaluate the inhibitory effect of ADSCs, we detected apoptosis of T cell subsets. Our results showed that induction of apoptosis in activated CD4+ and CD8+ T cells was the mechanism underlying the inhibitory effect of human ADSCs. Notably, conflicting results concerning the effects of human BM-MSCs on T cell apoptosis have been reported, including (1) induction of apoptosis [21,43], (2) suppression of apoptosis [44,45], and (3) no change in apoptosis [42,46,47]. Moreover, Gu and Chang reported that human PMSCs did not affect T cell apoptosis [18,48]. These conflicting results might be attributed to different tissue sources of MSCs, coculture settings, and stimulation agents, suggesting that these factors should be considered in clinical MSC-based therapy.
To further address the immunomodulatory effect of human ADSCs on T cell subsets, we added PBMCs to the preadherent ADSCs followed by stimulation with anti-CD3/CD28 beads and detected IFN-gamma production by CD4+ and CD8+ T cells. We found that the presence of human ADSCs significantly decreased IFN-gamma production in both CD4+ and CD8+ T cells, suggesting that the immunomodulation of ADSCs was not confined to a specific T cell subset, which highlights the clinical application of ADSCs for controlling unfavorable immune responses, such as graft versus host disease and autoimmune disorders.
Several studies along with our present research have reported that IFN-gamma-treated or activated T cells can increase PD-L1 expression of MSCs. However, it is unknown whether the upregulated expression of PD-L1 on ADSCs acts in turn to modulate IFN-gamma production by T cells. Interestingly, we found that PD-L1 blockade partially reversed the ADSC-inhibited IFN-gamma production by T cells in stimulated PBMCs, indicating that inhibition of IFN-gamma production was mediated by the upregulated PD-L1 expression on ADSCs. Additionally, the inhibition of T cell proliferation was also attenuated by the PD-L1 blockade. This result indicated that the effect between T cells and ADSCs was reciprocal, which may be helpful for switching from the inflammatory to the repair phase during tissue repair.
Notably, but not surprisingly, in this study, PD-L1 blockade led to partial but not total restoration of cell proliferation and IFN-gamma production in T cells, indicating that other molecules might be involved in ADSC-mediated immunosuppression. In this study, we found that the important inhibitory molecule Gal-9, which is not constitutively expressed on the surface of ADSCs, could be significantly induced when cocultured with stimulated allogeneic PBMCs. To investigate whether Gal-9 plays an important role in the immunosuppressive function of ADSCs, we performed coculture experiments similar to those of PD-L1. The Gal-9 blocking assay demonstrated that induction of Gal-9 expression on ADSCs was also responsible for the inhibition of cell proliferation and IFN-gamma secretion in activated CD4+ and CD8+ T cells. These results implied the overlapping pathways involved in ADSCs immunomodulation activities, thereby suggesting that the interplay between ADSCs and T cells merits further investigation.
It has been reported that appropriate inflammatory stimulation is requisite to trigger the immunosuppressive function of MSCs. Further investigation into the underlying signaling pathways in T cells is necessary. Therefore, we observed PD-1 and TIM-3 expression on T cells in TCR-stimulated PBMCs cocultured with ADSCs. Unexpectedly and importantly, a significant increase in PD-1 and TIM-3 expression was observed on both CD4+ and CD8+ T cells in coculture. It is of clinical significance to provide novel target molecules for delicate manipulation to ensure optimal immunomodulation in ADSC-based therapy.
In this study, we provided the first evidence that human ADSCs exerted modulatory effects on T cell subsets via the suppression of NF-kappaB activation mediated by the PD-L1/PD-1 and Gal-9/TIM-3 pathways. This finding explains why the Jagged blockade could partially but not completely reverse the Jagged-1/Notch-related inhibition of NF-kappaB signaling and T cell proliferation [49]. Our results showed that PD-L1/PD-1 and Gal-9/TIM-3 interactions were alternative pathways responsible for the immunomodulatory effect of ADSCs on T cells via NF-kappaB signaling. Therefore, it is necessary to determine the overlapping and complicated interactions among the PD-L1/PD-1, Gal-9/TIM-3, and Jagged-1/Notch pathways to finely tune immunomodulation in the future.
It should be pointed out that all ADSC donors in this study were female. The role of ADSCs from men in immunomodulation of T cell subsets should be investigated because male MSCs have a more potent osteogenic capacity compared with female [50,51]. In addition, some caution should be taken in predicting the in vivo role of ADSC-mediated immunomoduation because it could be a phenomenon restricted to in vitro conditions and not found in vivo due to the complexity of immunomodulation.
In conclusion, we found that ADSCs performed immunoregulatory functions partially through the inhibition of NF-kappaB activation in TCR-stimulated T cells via the PD-L1/PD-1 and Gal-9/TIM-3 pathways. Our data provided new insights into the mechanism of ADSC-mediated immunomodulation, which may have clinical implications for the development of novel strategies to precisely manipulate the immunomodulatory effects of ADSCs on T cells.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (no. 51272286), National Natural Science Foundation of Liaoning Province (no. 20102296), and Public Welfare Industry Research Project of the National Health and Family Planning Commission (no. 2015SQ00049).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.