
Abstract
Sca-1+ progenitor cells in the adult mouse aorta are known to generate vascular smooth muscle cells (VSMCs), but their embryological origins and temporal abundance are not known. Using tamoxifen-inducible Myf5-Cre ER mice, we demonstrate that Sca-1+ adult aortic cells arise from the somitic mesoderm beginning at E8.5 and continue throughout somitogenesis. Myf5 lineage-derived Sca-1+ cells greatly expand in situ, starting at 4 weeks of age, and become a major source of aortic Sca-1+ cells by 6 weeks of age. Myf5-derived adult aortic cells are capable of forming multicellular sphere-like structures in vitro and express the pluripotency marker Sox2. Exposure to transforming growth factor-β3 induces these spheres to differentiate into calponin-expressing VSMCs. Pulse-chase experiments using tamoxifen-inducible Sox2-CreERT2 mice at 8 weeks of age demonstrate that ∼35% of all adult aortic Sca-1+ cells are derived from Sox2+ cells. The present study demonstrates that aortic Sca-1+ progenitor cells are derived from the somitic mesoderm formed at the earliest stages of somitogenesis and from Sox2-expressing progenitors in adult mice.
Introduction
V
Resident adventitial Sca-1+ (CD45−Sca-1+CD31−) progenitor cells are abundant and can differentiate into VSMCs both in vivo and in vitro [3]. They localize to the intima of atherosclerotic plaque in vein grafts [3] and to the neointima following wire-induced injury of the femoral artery [4]. Sonic hedgehog signaling has been implicated in localization and/or development of adventitial Sca-1+ progenitor cells [5]. The proliferation of Sca-1+ cells after carotid injury is dependent on c-myb, which also regulates their transforming growth factor (TGF)-β-dependent differentiation into VSMCs by activating myocardin [6]. While Sca-1+ progenitor cells serve an important physiological role, their embryological origins and temporal abundance have not been adequately detailed. Understanding the origins of VSMC progenitors may inform their role in vascular disease.
Cell types constituting the vasculature do not originate from a single embryological source, yet cells derived from distinct embryological lineages often adopt the same fate. An important example of this is the aorta, where VSMCs from distinct embryological lineages contribute to this composite organ. For example, VSMCs in the aortic root, arch, descending thoracic, and abdominal aortic segments are derived from secondary heart field, neural crest, somitic, and splanchnic mesoderm, respectively [7].
Given this heterogeneity, we postulated that like VSMCs, stem and/or progenitor cells of the aorta may also derive from distinct embryological lineages. The embryological origins of vascular progenitor populations are unknown, with Sca-1+ progenitor cells not being derived from bone marrow [3] or neural crest lineages [5]. However, given that Sonic hedgehog signaling has been implicated in homing of aortic Sca-1+ progenitor cells to the adventitia [5], and also in somitogenesis [8], we hypothesized that aortic Sca-1+ progenitor cells may originate from the somitic mesoderm.
Myf5 is a marker of the somitic mesoderm lineage. Myf5 is the earliest myogenic factor expressed in the embryo, first expressed in the dermomyotome of the early somite at E8.0 [9]. Myf5 expression is limited to the somitic mesoderm, as no detectable expression of Myf5-CreER /LacZ was previously observed outside of the somite after induction with tamoxifen at E10.5, and was only expressed in the myotome of the somite and somite-derived tissues, such as the developing musculature of the head, neck, and limbs in whole-mount E12.5 embryos [10].
In the present study, we show that the adult mouse aorta contains Sca-1+ progenitor cells of the Myf5 lineage. Sca-1+ cells are specified from the Myf5 lineage at E8.5, before Myf5 cells contribute to skeletal muscle, and continue on throughout somitogenesis. The Sca-1+ progenitor cell population expands in situ between 4 and 6 weeks of age. Myf5-derived adult aortic cells form sphere-like structures in vitro and express the pluripotency marker Sox2. We go on to show (with an inducible Sox2 lineage trap) that 35% of all adult aortic Sca-1+ cells are derived from Sox2+ progenitors. Our work sheds new light on the embryological origins and postnatal development of aortic Sca-1+ progenitor cells.
Materials and Methods
Animals
Experimental protocols were approved by and in accordance with our Animal Resource Centre at the University Health Network (AUP# 1032, 1034, and 1605). Mice used include Myf5-Cre [11] (B6;129S4-Myf5tm3(cre)Sor/J), floxed tdTomato [12] (B6;129-Gt(ROSA)26Sortm9(CAG-tdTomato)Hze /J), Sox2-CreERT2 [13] (B6; 129S-Sox2tm1(cre/ERT2)Hoch/J), Myf5-Cre ER [10] (B6;FVB-Myf5tm1(Cre/Esr1*)Trdo/J with neomycin cassette removed), and Rosa26 [14] (FVB-Gt(ROSA)26Sor/J). Mice were housed in the animal care facility and maintained on standard chow diet and water. Both males and females were used at ages specified in the text or legends.
Whole-mount β-gal activity
Organs were fixed in 0.25% glutaraldehyde for 15 min and rinsed thrice with 1× PBS. Specimens were incubated in X-gal solution [0.2% X-gal, 2 mM MgCl2, 5 mM K4Fe(CN)6, 5 mM K3Fe(CN)6] at 37°C for 1–16 h. Organs were visualized with a dissecting stereomicroscope.
Tamoxifen administration in Sox2-CreERT2 and Myf5-CreER mice
Adult Sox2-Cre ERT2 mice at 8 weeks of age received tamoxifen 3 mg/d i.p. for 5 days. Cells were traced by flow cytometry 8 weeks after the last injection. For Myf5-Cre ER , four females were caged with one male, and females were subsequently checked for plugs early the next morning. If plugged, females were removed from the harem and housed separately. At E8.5, E10.5, and E12.5, pregnant females received a single treatment with tamoxifen 75 μg/g body weight. Active recombination in these knock-in mice occurs in a small time window that does not exceed 24 h postinjection [15] and leads to irreversible expression of tdTomato in Myf5 + cells and their progeny. Pups were delivered by C-section at E19.5 and lactated with a foster mom until weaning age. Tamoxifen was prepared by dissolving 100 mg of tamoxifen in 0.5 mL of ethanol and 9.5 mL of corn oil. After resuspending, the mixture was sonicated at 37°C for 10 min. Tamoxifen was stored at −20°C and used within 4 weeks.
tdT+ aortic cell isolation and culture
Mice were perfused with HBSS through cardiac puncture. Periaortic adipose tissue was removed and aortas (all aortic layers, including adventitia) were collected. For flow cytometry analyses, aortas were digested as described [16,17] with 450 U/mL collagenase I, 125 U/mL collagenase type XI, 60 U/mL hyaluronidase type I-s, and 60 U/mL DNase 1 in HBSS containing 20 mM Hepes at 37°C for 30 min with agitation. For cell culture experiments, aortas were digested differently [18]. Briefly, aortic tissue was incubated for 30 min at 37°C in an enzymatic digestion medium consisting of low-glucose DMEM/F12 3:1 supplemented with 200 U/mL collagenase type III, 0.1 mg/mL elastase (132 U/mg), and 0.5 mg/mL soybean trypsin inhibitor. After first digestion, aortas were placed into fresh digestion medium with identical components except for collagenase 130 U/mL for an additional 45 min. After the second digestion, tissue was again placed in fresh digestion medium with collagenase 130 U/mL and incubated at 37°C in a 5% CO2 humidified incubator for an additional 1 h. tdT+ CD45
Bone marrow, blood, skeletal muscle, brain, and spleen single-cell suspensions
All single-cell suspensions acquired from tissues were from animals perfused with 20 mL of HBSS through cardiac puncture. Bone marrow preparations were obtained by crushing the femurs of mice with a mortar and pestle. Samples were filtered through a 40-μm cell strainer and subsequently washed in HBSS before use. Peripheral blood was drawn by retro-orbital sampling using heparinized capillary tubes. Skeletal muscle from the hind limb was digested using collagenase I (see digestion mixture above) for 75 min at 37°C with rigorous agitation. Splenic and brain disaggregates were prepared by applying pressure to the spleen and brain with the blunt end of a syringe through a 40-μm cell strainer and flushing with HBSS. Red blood cells (RBC) were lysed from bone marrow and spleen (and peripheral blood) preparations by washing (and incubating for 10 min) in ice-cold ammonium chloride.
Flow cytometry and cell sorting
Pellets from cell suspensions from single aortas and peripheral blood were resuspended in 100 μL of FACS buffer [2% FBS in HBSS]. Splenic disaggregates, skeletal muscle, brain, and bone marrow preparations were resuspended in 1 mL of FACS buffer. Cells were incubated for 1 h with fluorochrome-conjugated anti-mouse antibodies for different cell surface markers listed in Supplementary Table S1 (Supplementary Data are available online at
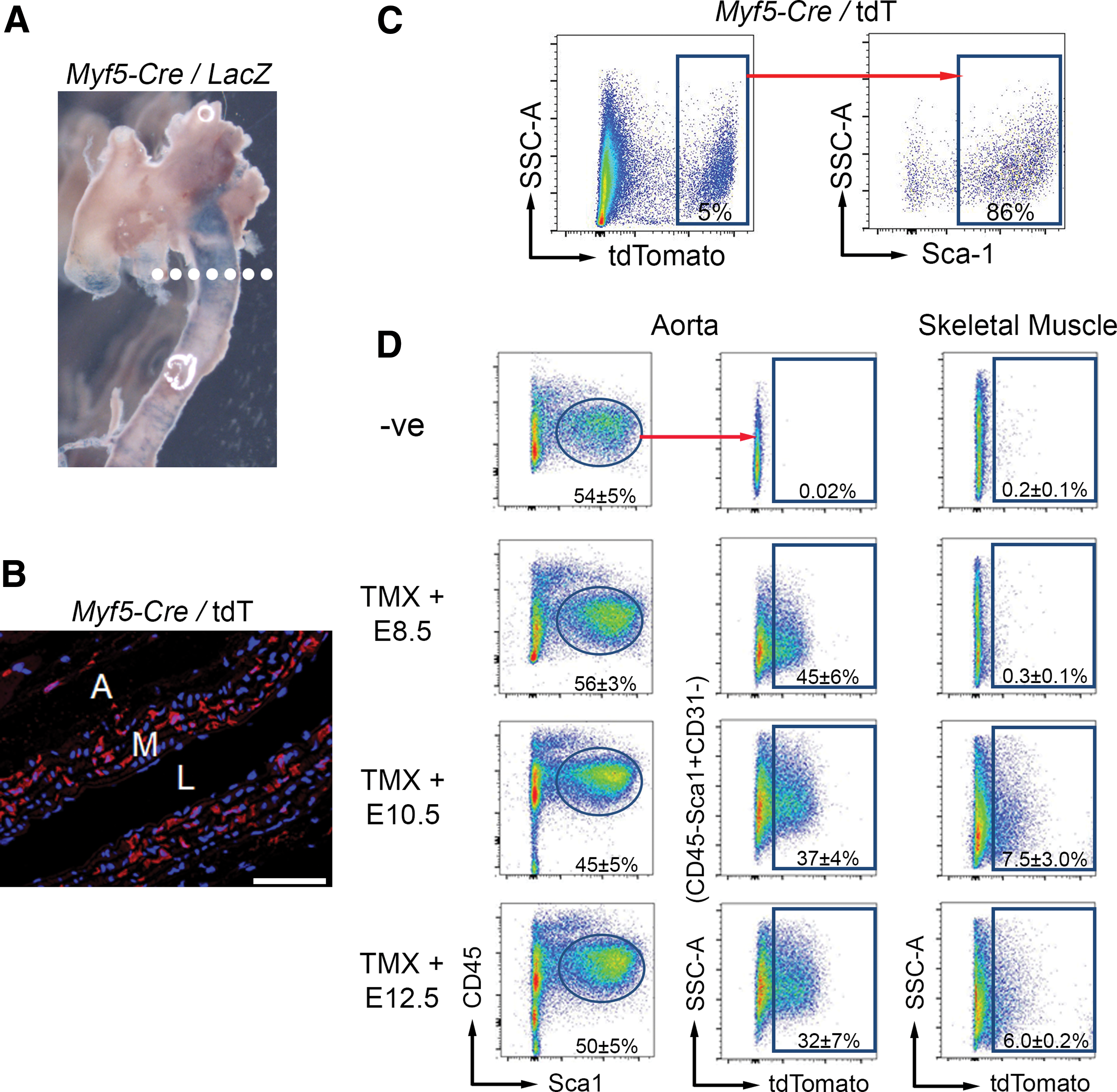
Somitic mesoderm-derived adult aortic Sca-1+ progenitor cells are specified throughout somitogenesis.
tdTomato+ Sca-1+ cell and bone marrow transplant
Aortas from 7 to 12 Myf5-Cre/tdT mice were dissected and digested in collagenase III according to the protocol above [18]. Single-cell suspensions were flow-sorted to acquire the tdT+ population of cells. The tdT+ cell populations (5–10 × 104 cells per mouse) were then injected into the tail vein of negative littermates that had been lethally irradiated with 10 Gy of radiation. Approximately 1 × 106 bone marrow cells from a negative littermate were coinjected with the flow-sorted population. Tail vein injections were performed 2–4 h postirradiation. Mice were housed for 16 weeks on normal chow diet before being sacrificed and aortas processed for flow cytometry (n = 3).
Immunofluorescence staining of cells in culture and histology
Cells were fixed by incubating in 4% PFA (Cytofix; BD Biosciences) for 15 min at room temperature and stained as described [21]. After fixation, cells were incubated in Cytowash/Perm buffer (BD Biosciences) for 30 min and blocked with normal goat serum for 1 h. Cells were incubated in primary antibody at 4°C overnight (Supplementary Table S1). After washing in 1× PBS, cells were incubated with secondary antibody for 1 h at room temperature. Nuclei were counterstained with 10 μg/mL Hoechst 33258.
Statistics
Normally distributed data were analyzed using Student's t-test. All statistical analyses were performed using GraphPad (Prism) (version 5.04). For Figure 2, 15-day-old postnatal mouse aortas were used as a control for 1-way analysis of variance (1-way ANOVA) comparisons. Statistical significance was established as a P value below 0.05 (P < 0.05). The analysis of variance was statistically significant for each analysis (****P < 0.0001).

Adult aortic Sca-1+ progenitor cells expand in situ between 4 and 6 weeks of age.
Results
Adult aortic Sca-1+ progenitor cells are specified in early somitogenesis (E8.5)
The use of an Myf5-Cre mouse crossed to a Rosa26LacZ reporter mouse confirmed somitic mesoderm involvement in aortic development. Consistent with previous reports [7], X-gal staining revealed Myf5-derived cells in the descending thoracic aorta (Fig. 1A). Using the Myf5-Cre mouse crossed to a floxed tdTomato (tdT) reporter mouse, we observed tdT+ expression in aortic sections (Fig. 1B). To assess whether Sca-1+ cells are Myf5 derived, we flow-sorted aortic cells from Myf5-Cre/tdT mice. We observed a population of tdT+ Sca-1+ cells (Fig. 1C), demonstrating their derivation from the somitic mesoderm. These data demonstrate for the first time the embryological origin of adult aortic Sca-1+ progenitor cells.
To determine whether somitogenesis contributes specifically to the developmental origins of Sca-1+ progenitor cells in the adult mouse aorta, we next utilized an inducible somitic mesoderm lineage-tracing approach. Tamoxifen-inducible Myf5-CreER mice were crossed to floxed tdT reporter mice, and tamoxifen was administered at E8.5. This experiment revealed unequivocal evidence of tdT+ (ie, Myf5 lineage-marked) Sca-1+ cells in the aortas of 6-week-old adult mice (Fig. 1D), demonstrating their derivation from the onset of somitogenesis. Consistent with previous reports, Myf5 + cells contribute to skeletal muscle at E10.5 [22].
To determine the significance of our finding, we determined the embryological/somitic contributions of CD45−Sca-1+CD31− progenitors in the 6-week-old adult aorta of mice induced with tamoxifen at E8.5. In this experiment, 45% ± 6% of all CD45−Sca-1+CD31− cells from the aorta were derived from the Myf5 lineage (ie, were tdT+) (Fig. 1D). By contrast, many aortic CD31+ cells were Myf5-Cre ER /tdT+, which suggests that some endothelial cells also originate from the somitic mesoderm (Fig. S1).
Sca-1+ specification continues throughout somitogenesis
To determine the kinetics of somitic contributions to the Sca-1+ progenitor cell population in the adult mouse aorta, we also induced recombination with tamoxifen in pregnant females at E10.5 and E12.5. When Myf5-CreER /tdT recombination was induced at E10.5, only 37% ± 4% of the adult aortic CD45−Sca-1+CD31− cells were tdT+ (Fig. 1D). When tamoxifen-induced recombination begins at E12.5, 32% ± 7% of the adult aortic CD45−Sca-1+CD31− cells were tdT+. These data indicate that adult aortic Sca-1+ progenitor cells are embryologically specified throughout somitogenesis, but of all adult aortic CD45−Sca-1+CD31− cells, only 8% are specified between E8.5 and E10.5, while 32% are specified at E12.5 and beyond (ie, contribution of Myf5-derived progenitors to the adult CD45−Sca-1+CD31− population is greater at late stages of somitogenesis). Focusing on this more robust stage of somitic derivation, we used tamoxifen induction at E12.5 for further evaluation.
Somite-derived Sca-1+ progenitor cells expand significantly between 4 and 6 weeks after birth
To uncover the temporal abundance of aortic Sca-1+ progenitor cells, we induced embryos with tamoxifen at E12.5 and quantified them at several postnatal time points. In the aortas of 1-day-old neonates induced with tamoxifen at E12.5, there was little tdT+ labeling (Fig. 2). Examining subsequent time points, we found that the proportion of Sca-1+ cells in the aorta expands most significantly between 4 and 6 weeks of age, which remains true even after excluding (ie, gating out) CD45+ and CD31+ cells (Fig. 2A, D). The absolute numbers of total tdT+ and CD45−Sca-1+CD31−tdT+ cells also increase after 4 weeks of age (Fig. 2C, E). Thus, we identify 4–6 weeks of age as a significant time point in the postnatal development of the adult mouse aorta.
Somite-derived adult aortic Sca-1+ progenitor cells do not home from the hematopoietic tissues
Aortic Sca-1+ cells have been shown by our group and others to be proliferative [3,4,6]. Given the postnatal expansion of Myf5-derived adult aortic Sca-1+ cells observed, we sought to determine whether these cells contribute to the aorta from other sources or expand in situ. Following tamoxifen injection at E8.5, E10.5, or E12.5, the bone marrow, spleen, and blood from tamoxifen-injected Myf5-Cre ER /tdT mice were absent of tdT+ cells (Fig. 3A). We then assessed whether circulating Myf5-derived cells home to the adult aortic Sca-1+ population. For this experiment, we flow-sorted tdT+Sca-1+ cells from aortas of Myf5-Cre/tdT mice and injected them into the tail vein of lethally irradiated mice with the bone marrow of tdT-negative littermates. Over 80% of these tdT+ aortic cells were Sca-1+ (Fig. 1C); thus, we estimate that ∼50,000–100,000 Sca-1+ cells were injected per mouse. At 16 weeks post-bone marrow transplantation (BMT), no tdT+ Sca-1+ cells could be identified in aortas of recipient mice (n = 3) (Fig. 3B), demonstrating that Myf5 lineage-derived aortic Sca-1+ cells do not home to the aorta and suggesting that Myf5-derived adult aortic Sca-1+ cell expansion occurs in situ.
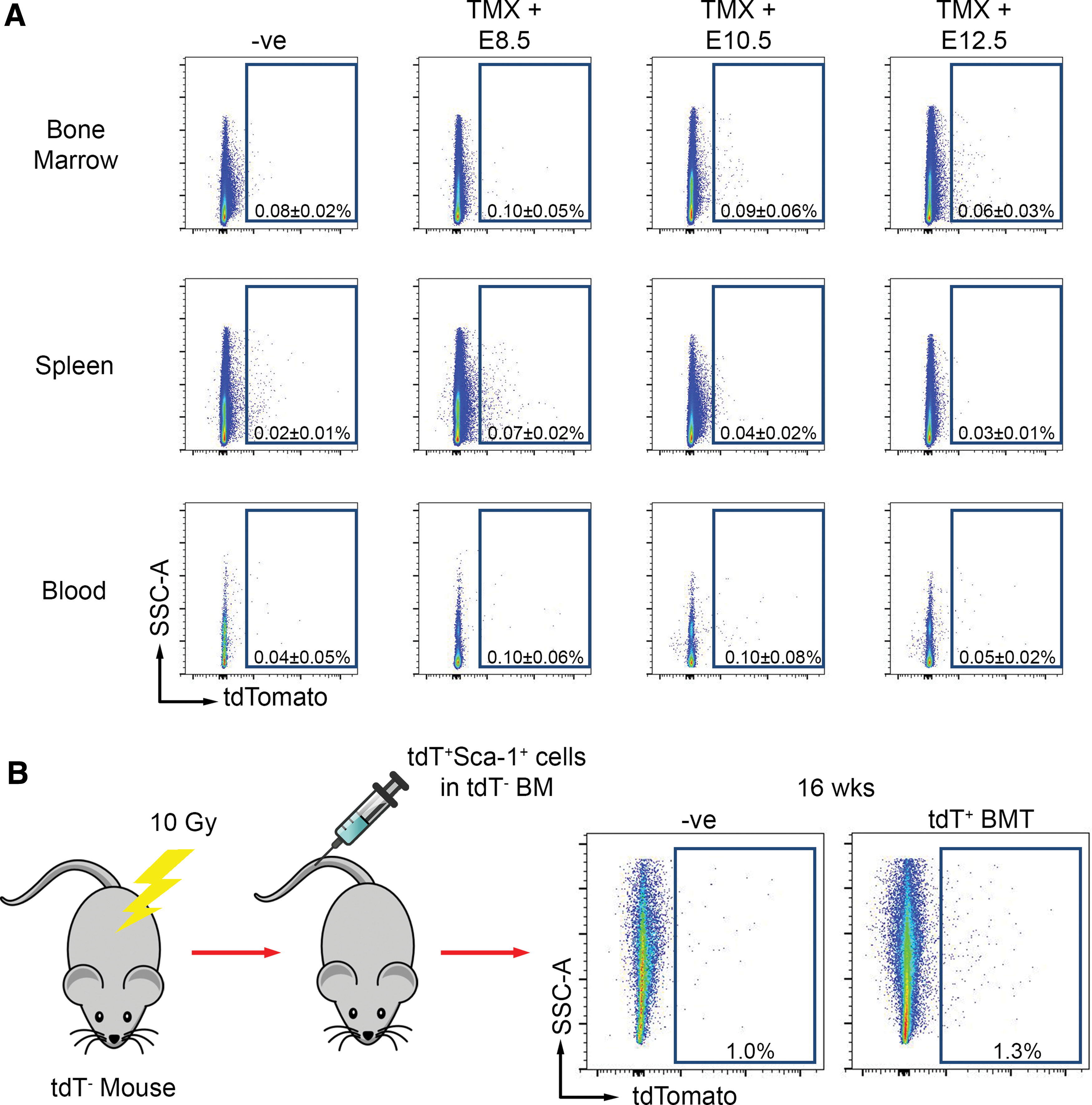
Somite-derived adult aortic Sca-1+ progenitor cells are not found in hematopoietic organs and do not home following bone marrow transplantation.
Somite-derived adult aortic cells form sphere-like structures in culture, express the pluripotency marker Sox2, and can differentiate into VSMCs
Next, we sought to demonstrate the stem/progenitor-like potential of Myf5-derived adult aortic cells. Having previously established a sphere culturing protocol [21], we next determined the sphere-forming capacity of Myf5-derived aortic cells. We flow-sorted 1–4 × 104 tdT+ cells, then using serum-free medium containing EGF, FGF, and B27, we plated cells at low density (10 cells/μL) in 1.5% methylcellulose to promote clonal expansion of single cells. From all Myf5-derived cells, we successfully generated spheres using 0.7% ± 0.5% of tdT+ cells from the proximal aorta and 0.5% ± 0.3% of tdT+ cells from the distal aorta (Fig. 4A). Importantly, Myf5-derived adult aortic cells were found to express the pluripotency marker Sox2 [21,23], as determined by immunofluorescence of tdT+ spheres (Fig. 4B).
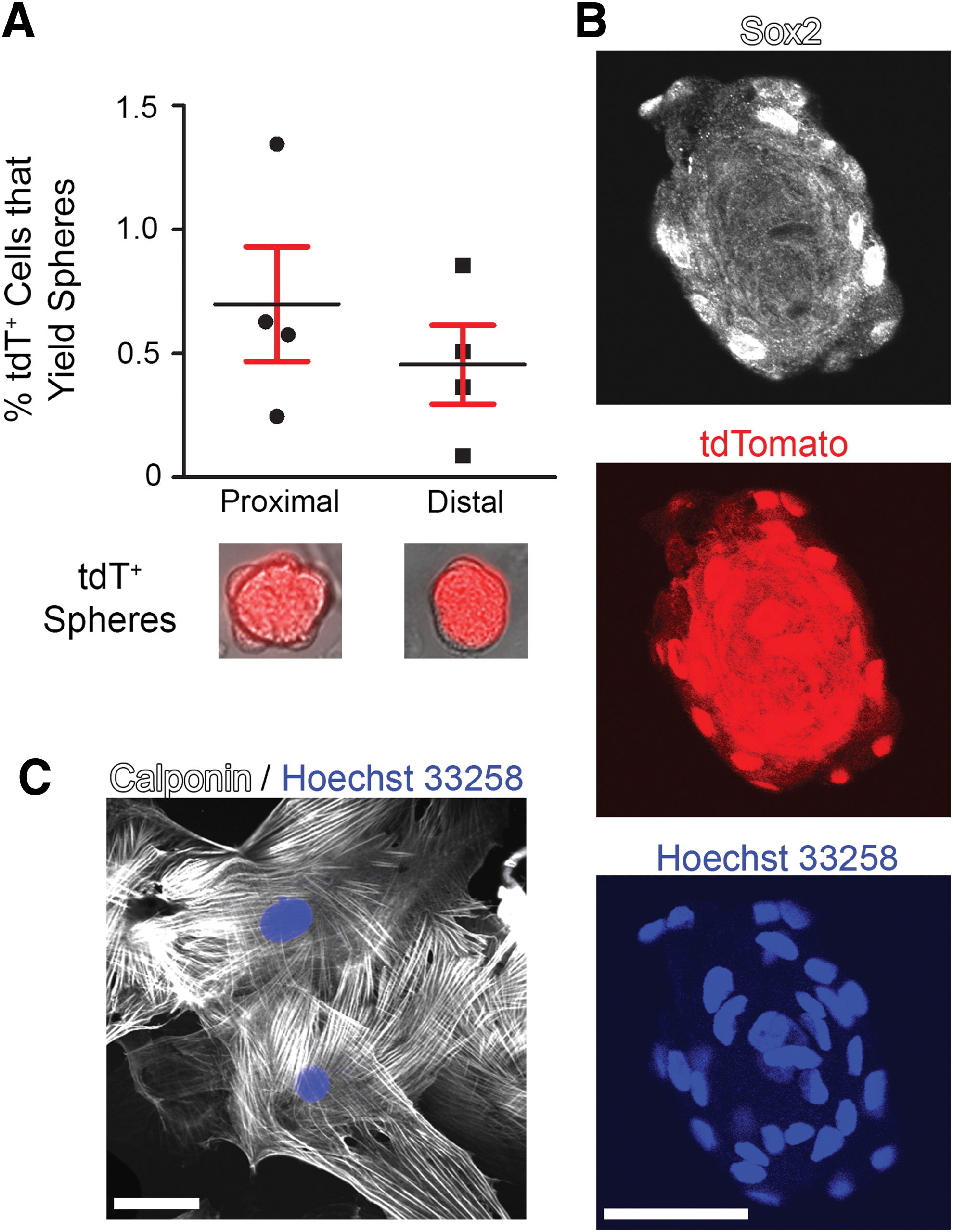
Somite-derived adult aortic cells form sphere-like structures, express the pluripotency marker Sox2, and can differentiate into VSMCs in culture.
To demonstrate that Myf5-derived adult aortic cells are capable of becoming VSMCs, we subjected these cells to an established VSMC-directed differentiation protocol. This experiment revealed that Myf5-derived tdT+ adult aortic cells could become calponin-expressing VSMCs (Fig. 4C), demonstrating their progenitor-like property.
Aortic Sca-1+ cells are also produced from Sox2-expressing progenitors in adult mice
Given that Myf5-derived progenitor cells expressed Sox2 in vitro, we hypothesized that Sox2+ cells in vivo would also give rise to Sca-1+ progenitors in the adult aorta. To this end, we sought to investigate the relationship between adult aortic Sox2+ cells and aortic Sca-1+ cells in the steady state. We acquired a tamoxifen-inducible Sox2-CreERT2 lineage-tracing mouse crossed to a floxed tdT reporter. Here, 8-week-old mice that received tamoxifen daily for five consecutive days were processed for cell sorting 8 weeks after the last injection. This pulse-chase experiment revealed that 15% ± 6% of all aortic cells were derived from Sox2-tagged cells (Fig. 5A). As a positive control, 39% ± 4% of the brain was tdT+, consistent with the known extent of Sox2+ activity in the brain (Fig. 5A). Supporting the physiological relevance in vivo of our earlier in vitro finding that Myf5-derived cells could express Sox2 and contribute to the Sca-1+ progenitor population, we found that 34% ± 5% of Sca-1+ cells in the adult aorta were Sox2 derived (Fig. 5B). Immunofluorescence-based detection of tdT and Sca-1 confirmed their expression in the aortic vessel wall (Fig. 5C). No tdT+ cells were observed in the bone marrow, blood, spleen, or skeletal muscle of these animals (Supplementary Fig. S2). Taken together, these data reveal that Sox2+ cells in the adult aorta contribute to aortic Sca-1+ progenitor cells in the steady state.
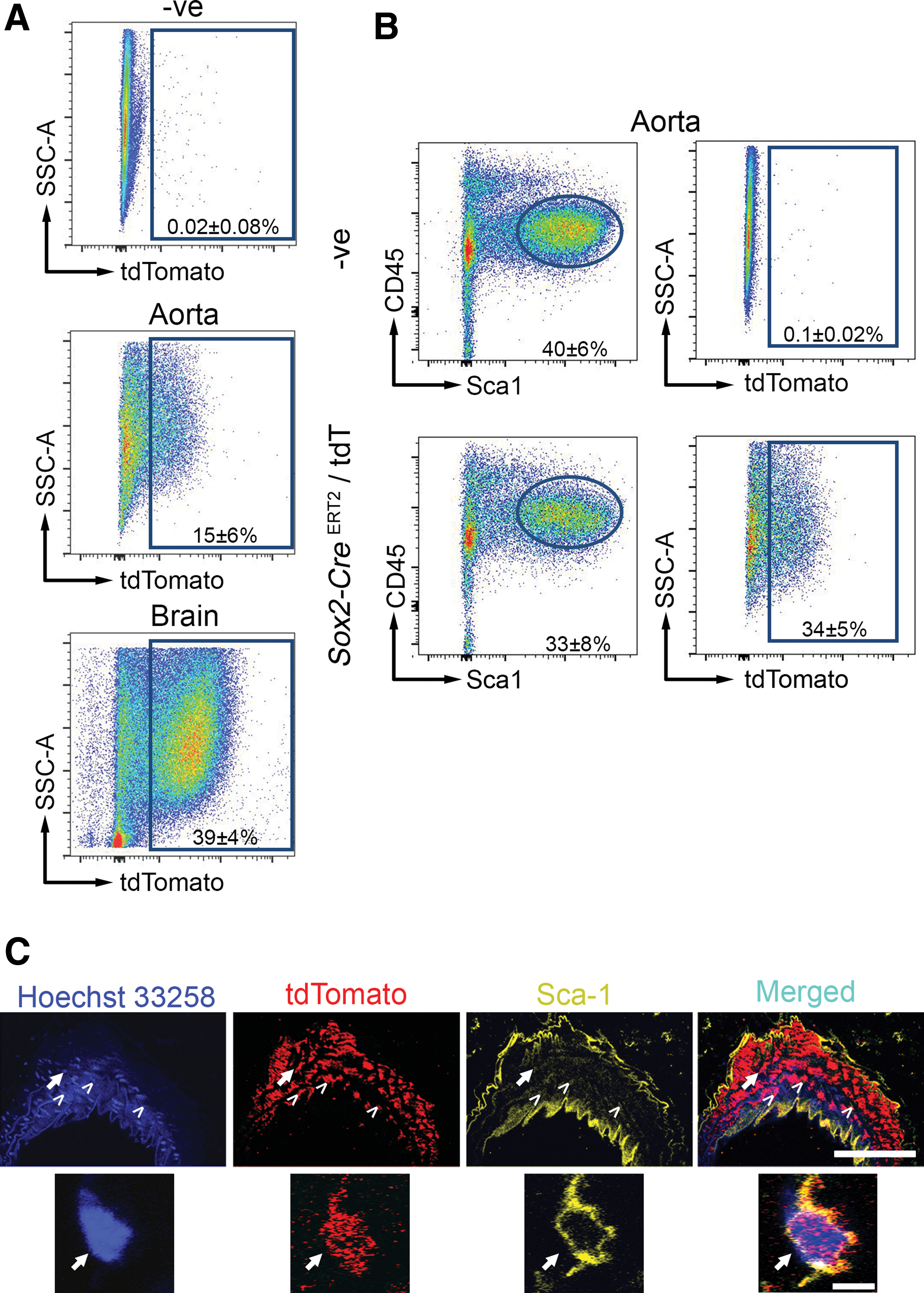
Aortic Sca-1+ cells are also produced from Sox2-expressing progenitors in adult mice. Sox2-CreERT2
/tdT mice at 8 weeks of age were given 1 injection of tamoxifen per day for five consecutive days. Mice were sacrificed 8 weeks after the last injection and their tissues were isolated for flow cytometry.
Discussion
Aortic Sca-1+ progenitors are key mediators of vascular homeostasis; however, the origin and abundance of these cells are not known. In the present study, using a tamoxifen-inducible Myf5-CreER system, we lineage traced aortic Sca-1+ progenitor cells to the somitic mesoderm. We observed lineage-marked Sca-1+ cells when Myf5-CreER /tdT embryos had been induced with tamoxifen at E8.5, E10.5, and E12.5. Thus, we were able to define that labeling of the aorta occurs at the beginning of somitogenesis (E8.5) before Myf5+ cells populate the skeletal muscle, and Myf5 + cells continue to contribute to the aorta throughout somitogenesis. In contrast, expression of alpha smooth muscle actin and SM22α in the aorta [24] and skeletal muscle specification in the limbs occur at E9.5 [25,26]. Previously, at similar time points in the Myf5-CreER /tdT mouse embryo, tamoxifen induction at E10.5 and whole-mount X-gal staining at E11.5 demonstrated that Cre activity is detected only in myotomes of somites [10]. When embryos are induced with tamoxifen at E10.5 and E11.5 and whole-mount X-gal staining is performed at E12.5, Cre activity is detected in myotomes of somites as well as the developing musculature of the head, neck, and limbs [10]. Our work corroborates studies demonstrating that VSMCs and endothelial cells of the aorta can be derived from the somitic mesoderm [27,28]. Recently, Majesky et al. used VSMC lineage tracing to show that adventitial Sca-1+ progenitor cells of several large mouse arteries can be derived from VSMCs [29]. We now have identified a novel source of Sca-1+ progenitor cells in the adult aorta.
As very little tdT+ expression was observed 1 day after birth, we investigated the postnatal development of Sca-1+ cells in the aorta. By examining the kinetics of aortic Sca-1+ progenitor (CD45− CD31−) cells, we observed that the abundance of Sca-1+ cells (and tdT expression overall) accelerated between 4 and 6 weeks of age rather than during embryonic development, identifying this as a critical time point in the postnatal development of the aorta. This finding also suggests that Sca-1+ progenitor cells expand in situ rather than by colonizing the aorta from other sources. Our findings complement an earlier report that suggested Sca-1+ expression in the developing aorta begins between E15.5 and E18.5 [5]. In that study, Passman et al. performed immunofluorescence against Sca-1 in aortic root cross-sections from mouse embryos and detected Sca-1 expression only at E18.5 and onward. Sca-1 expression was also detected in the descending aorta at E18.5. Our findings support an embryological origin for these Sca-1+ cells as we find that Myf5 lineage trapping as early as E8.5, E10.5, and E12.5 ultimately yields Sca-1+ cells in the adult mouse aorta. Thus, we suggest that Sca-1+ cells are specified beginning at E8.5, and consistent with Passman et al., Sca-1+ expression does not begin until later stages. We add to this knowledge by establishing that postnatal Sca-1+ expression markedly expands in situ, starting at 4 weeks of age. Nonetheless, the determinants that trigger this postnatal expansion and its pathophysiological significance remain to be determined.
We sought to demonstrate that Myf5-derived aortic cells are indeed the source of cells with stem/progenitor-like capabilities. To do so, we demonstrated their sphere-forming capacity, Sox2+ expression, and VSMC differentiation potential. Although the spheres we cultured contained Sox2+ cells, not all of the cells within our spheres stained Sox2+. This heterogeneity may be due to several factors: (a) Sox2+ expression may be greater (and thus easier to detect) in proliferating cells [30]; (b) there may be variable penetration of the Sox2 antibody; and (c) the spheres themselves may not be clonal structures. Arguing against the latter, other studies employing the same 1.5% methylcellulose conditions we used have shown very low risk of cell aggregation (ie, nonclonality) [31], suggesting that most, if not all, of our spheres are the result of clonal cell expansion. This may suggest that paracrine factors are required to promote expansion of these cells in culture. Nonetheless, 34% ± 5% of Sca-1+ aortic cells were found to be derived from Sox2+ cells, which suggests a role for Sox2+ cells in the specification of adult aortic Sca-1+ cells.
While our study reveals that the Myf5 lineage is a major source of adult aortic Sca-1+ cells, other lineages may also contribute to this population. For example, VSMCs can arise at E7.5 from hemangioblasts in the primitive streak of the developing embryo [32], which may represent another origin of aortic Sca-1+ cells. We also demonstrate that Sca-1+ cells can be embryologically derived from Sox2 lineage-marked cells, with a portion of Sox2+ cells themselves originating from the Myf5 lineage. However, it remains to be determined whether Sox2 + cells are derived from other distinct lineages.
Importantly, differences between Sca-1+ cells of varying lineages remain to be examined. For example, it has been observed in macrophages that their developmental origin can influence the phenotype [33], raising the possibility that Sca-1+ cells from distinct embryological origins may have different functional properties or dictate the phenotype of their differentiated cell types (eg, contractile versus synthetic VSMCs [34]).
Further studies are required to determine the physiological function of Sca-1+ cells and the potential role of Myf5-derived Sca-1+ cells in cardiovascular disease. Given that conditions such as hypertension, atherosclerosis, and aortic aneurysm directly affect aortic structure and may do so, in part, by modulating repair and/or regenerative responses of the aortic Sca-1+ progenitor population, studying the pathophysiological relevance of postnatal Sca-1+ cells and their differing embryological origins may be of particular interest. For example, whether genetic or environmental disruption of the Myf5-derived aortic Sca-1+ cell expansion between 4 and 6 weeks of age would lead to vascular deficits or increased susceptibility to aortic disease is not yet known. In contrast to other Sca-1+ populations, which can be bone marrow derived and/or hematopoietic, we demonstrated that the CD45−Sca-1+CD31− cell population in question expands in situ. We noted that Myf5-derived cells populate an atherosclerosis- and aneurysm-susceptible region of the mouse aorta. In numerous studies, grafts from these atherogenic regions of the aorta to atherosclerosis-resistant areas caused them to develop atherosclerosis [7]. This was considered to be due to innate differences in vessel profiles of these respective regions [7]. Thus, our findings raise the possibility that developmental origins of various aortic regions may have a role in disease and that Myf5-derived adult aortic cells are worthy of further investigation.
Footnotes
Acknowledgments
This study was funded by operating grants to MH from the Canadian Institutes of Health Research (CIHR) (MOP136850 & MOP14648), Heart and Stroke Foundation (Ontario; T6757), and McEwen Centre for Regenerative Medicine (Acceleration Award). S.K.S. was funded, in part, by a fellowship from the Heart and Stroke Richard Lewar Centre of Excellence.
This work was presented, in part, at the 2016 Scientific Sessions of the American Heart Association.
Author Disclosure Statement
The authors have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.