
Abstract
Human embryonic stem cell (hESC) derived 3D human lung organoids (HLOs) provide a promising model to study human lung development and disease. HLOs containing proximal or/and immature distal airway epithelial cells have been successfully generated in vitro, such as early staged alveolar type 2 (AT2) cells (SPC+/SOX9+) and immature alveolar type 1 (AT1) cells (HOPX+/SOX9+). When HLOs were transplanted into immunocompromised mice for further differentiation in vivo, only few distal epithelial cells could be observed. In this study, we transplanted different stages of HLOs into immunocompromised mice to assess whether HLOs could expand and mature in vivo. We found that short-term transplanted HLOs contained lung progenitor cells (NKX2.1+, SOX9+, and P63+), but not SPC+ AT2 cells or AQP5+ AT1 cells. Meanwhile, long-term engrafted HLOs could differentiate into lung distal bipotent progenitor cells (PDPN+/SPC+/SOX9+), AT2 cells (SPC+, SPB+), and immature AT1 cells (PDPN+, AQP5−). However, HLOs at late in vitro stage turned into mature AT1-like cells (AQP5+/SPB−/SOX9−) in vivo. Immunofluorescence staining and transmission electron microscopy (TEM) results revealed that transplanted HLOs contained mesenchymal cells (collagen I+), vasculature (ACTA2+), neuroendocrine-like cells (PGP9.5+), and nerve fiber structures (myelin sheath structure). Together, these data reveal that hESC-derived HLOs would be useful for human lung development modeling, and transplanted HLOs could mimic lung organ-like structures in vivo by possessing vascular network and neuronal network.
Introduction
T
During lung development in mouse, AT2 and AT1 cells are derived from a bipotent progenitor (BP) population at saccular stage and then expanded and matured at alveolar stage [1,3,10]. The BP cells co-express AT2 and AT1 cell markers (SPC+/PDPN+) [1,10] and distal lung progenitor marker SOX9 [11]. Based on single-cell RNA-seq results, BP cells (SPC+/PDPN+) can express more SPC, SPB, and downregulate early AT1 marker PDPN to generate mature AT2 cells (SPC++/SPB+/PDPN−) or express more mature AT1 markers and downregulate AT2 markers to generate mature AT1 (PDPN++/AQP5+/SOX9−) [10]. Studies on adult human/mouse lung confirm that mature AT1 cells do not express distal progenitor marker SOX9 [5,12]. It has been reported that hESC-derived HLOs in vitro contain SPC+/SOX9+ and HOPX+/SOX9+ progenitor-like cells, which resemble early staged AT2 cells and immature AT1 cells, respectively [5].
With the support of a bioartificial microporous poly (lactide-co-glycolide) (PLG) scaffold, transplanted HLOs grown in vivo possess proximal airway-like epithelium with basal, ciliated, goblet, and club cells, while the distal airway AT1 and AT2 cells are missing [13]. Similarly, orthotopic lung transplantation of dissociated bud tip organoid cells could not differentiate into distal cell types, but the proximal ciliated, goblet, and club cells [9]. Finally, although transplanted hESC-derived HLOs could give rise to AT1 and AT2 cells under the kidney capsule, the AT1 cells were immature since the mature marker AQP5 was not detected [7].
In this study, we efficiently generated AT1 and AT2 cells in long-term transplanted hESC-derived HLOs. Compared with previous differentiation methods, our strategy generates lung distal BP cells (PDPN+/SPC+/SOX9+) and mature AT1-like (AQP5+/SPB−/SOX9−) cells for the first time. These findings indicate that hESC-derived HLOs provide a promising model to study human lung development and regenerative medicine.
Materials and Methods
Maintenance of hESCs
H9 hESCs were cultured on Matrigel (BD Biosciences; 354277) coated plates in mTeSR1 medium (StemCell Technologies; 05850) at 37°C with 5% CO2. Cells were manually passaged at 1:4 to 1:6 split ratios every 4–5 days. All hESC work in this study has been approved by the Institutional Embryonic Stem Cell Research Oversight (ESCRO) Committee of Southern Medical University.
Generation of hESC-derived HLOs
The hESC-derived HLOs were generated as previously described [5] with several modifications. For lung organoid induction, we first generated definitive endoderm (DE) cells from hESCs (∼90% confluence) by the combination of 1–2 μM CHIR-99021 (Tocris; 4423-10 MG) [14] and 100 ng/mL Activin A (R&D Systems; 338-AC-050) for 3 days in RPMI1640 medium and then 200 ng/mL Noggin (R&D Systems; 6057-NG-100), 500 ng/mL FGF4 (Peprotech; 100-31-1 MG), 10 μM SB431542 (Tocris; 1614-10 MG), and 2 μM CHIR99021 (Tocris; 4423-10 MG) in Advanced DMEM/F12 (Life Technologies; 12634010) for next 4 days [5]. After 7 days treatment with factors, cells were embedded in a droplet of Matrigel (BD Biosciences; 356237) and fed with Advanced DMEM/F12 containing 1% fetal bovine serum. Organoids were transferred into new Matrigel droplets every 5–8 days.
Quantitative RT-PCR
Total RNA was extracted using the TRIzol reagent (MRC, TR1187), and 1 μg of total RNA was reverse transcribed using the ReverTra Ace Kit (TOYOBO; 34520B1). cDNA was properly diluted and used as PCR template. PCRs were performed with the SYBR®Premix Ex Taq™ Kit (TAKARA, RR420A). GAPDH was used as the internal control. Three to four biological replicates were performed for each assay, and data represent mean ± SD. Primers used are listed in Supplementary Table S1 (Supplementary Data are available online at
Flow cytometry
The cells (D3, DE cells)/organoids (D21, lung lineage organoids) were incubated in Accutase for 3/20 min at 37°C and then collected gently. For FOXA2, NKX2.1, and SOX9 analysis, cells were fixed by 2% paraformaldehyde (PFA) for 20 min at room temperature (RT), followed by permeabilizing with 0.2% Triton X-100 (Sigma; T9284)/PBS for 30 min at RT and blocking with 10% fetal bovine serum (FBS) solution at RT for 1 h. Primary antibodies were added at appropriate dilutions for 30 min at RT and if necessary, incubated in the secondary antibodies for 20 min at RT. After rinsing with 2% FBS/PBS, the cells were analyzed using a BD LSRFortessa flow cytometer (BD Biosciences).
Kidney capsule transplantation
Immune deficient B-NSG (NOD-Prkdcscid IL2rgtm1 /Bcgen) mice were purchased from Beijing Biocytogen Co., Ltd. (Beijing, China). HLOs were placed under the renal capsule of anesthetized NSG mice. Briefly, the left flank was sterilized using Chlorhexidine and isopropyl alcohol. Host kidney was exposed through a left lateral incision. D21HLOs, D31HLOs, and D41HLOs (10–20 organoids (1–2 mm3) per mice) were manually placed in a subcapsular pocket of the kidney of female 6–9-week-old NSG mice using forceps. Xenografts were harvested at 14 days or 100–120 days after transplantation. All mouse work was approved by the Institutional Animal Care and Use Committee of Southern Medical University.
Immunofluorescence staining
Samples were fixed in 4% PFA overnight at 4°C and then rinsed with PBS and incubated overnight again at 4°C in 30% sucrose solution. The samples were next overlaid with OCT compound and frozen using dry ice and stored at −80°C. Ten micrometer sections were permeabilized with 0.2% Triton X-100 (Sigma; T9284)/PBS for 45 min at RT and then blocked with blocking solution (10% FBS) at RT for 1 h or overnight at 4°C. Primary antibodies were added at appropriate dilutions overnight at 4°C and secondary antibodies at RT for 1 h. Finally, samples were counterstained with 4′, 6-diamidino-2-phenylindole (DAPI) (Sigma; D9542) for 5 min and then imaged with the Zeiss LSM 880 confocal microscope (Carl Zeiss). Antibodies used in this study are listed in Supplementary Table S2. Imaris software was used to render Z-stack three-dimensional images.
Hematoxylin and eosin staining
D21, D31, and D41 tHLOs were overlaid with OCT compound and frozen using dry ice and stored at −80°C. Ten micrometer sections were used to perform Hematoxylin and Eosin (HE) staining following the instruction and then imaged with Nikon microscope.
Transmission electron microscopy
D41 HLOs and tHLOs were cut into small pieces and incubated in fixative containing 2.5% glutaraldehyde and 0.8% PFA solution and were postfixed with 1% osmium tetroxide, dehydrated in a series of alcohol, and embedded. Ultrathin sections were stained with uranyl acetate and lead citrate. The specimens were imaged using a Hitachi H-7500 electron microscope.
Experimental replicates and statistics
All experiments were done on at least three (n = 3) independent biological samples. Statistical analysis was done using Prism software. If only two groups were being compared, a two-sided Student's t-test was used. If more than two groups were being compared with in a single experiment, an unpaired one-way ANOVA was carried out and followed by Tukey's multiple comparison test to compare the mean of each group with the mean of every other group within the study. The differences between the groups were considered statistically significant for P ≤ 0.05.
Results
Generation of HLOs from hESCs
Several methods for the generation of hESC-derived HLOs have been previously developed [5,7,9,15 –17]. We constructed an alternative to generate HLOs from hESCs (Fig. 1A, B). In our protocol, the first step is to induce DE from hESCs with Activin A [18] and CHIR-99021 (GSK3β inhibitor) that has been proved effective in efficient induction of DE cells from hESCs [6,14]. In step 2, DE cells were treated with Noggin, SB431542, CHIR-99021, and FGF4 for 4 days [5] and changed from 2D culture to 3D culture by Matrigel embedding with medium containing 1% FBS for 34 days.

Generation of human lung organoids from hESCs.
Using quantitative real-time PCR, we checked the expression of various lineage markers at day (D) 0, 3, 21, 31, and 41 along with differentiation (Fig. 1C). The pluripotency marker POU5F1 peaked at D0, the ESC stage, and decreased when differentiation started. SOX2 is a pluripotency marker and a lung proximal progenitor marker and is highly expressed at D0, D21, and D31. SOX17 and FOXA2 are DE markers, and they were highly expressed at D3. NKX2.1, SOX9, and P63 are lung progenitor and stem cell markers and are highly expressed at D21 and D31. The markers for more differentiated cell types appeared late at D41, including SCGB1A1 (club cells), MUC5AC (goblet cells), SPC and SPB (AT2 cells), and HOPX (AT1 cells).
Next, we checked the protein expression levels of lineage markers. At D3, the DE stage, SOX17 expression was evidenced by immunostaining assay, and about 90% of cells expressed FOXA2 in a FACS assay (Supplementary Fig. S1A, B). At D21, the lung endoderm stage, about 57% of cells expressed lung lineage marker NKX2.1, and about 50% of cells expressed distal lung progenitor marker SOX9 (Supplementary Fig. S1C, D). At D41, the lung organoid stage, cells expressing NKX2.1 (lung lineage), SOX9 (distal progenitor cells), P63 (basal cells), MUC5AC (goblet cells), and SPC (AT2 cells) could be detected by immunostaining assay (Fig. 1D). Meanwhile, transmission electron microscopy (TEM) showed lamellar bodies, which are specific secretory organelles for surfactant protein trafficking and secretion in AT2 cells [1,19,20] (Fig. 1E). TEM also revealed multiple cilia with a “9 + 2” structure consisting of nine outer microtubule doublets and a central pair of microtubule singlets (Supplementary Fig. S1E), the specific features of motile cilia [21], indicating that ciliated cells were successfully differentiated in HLOs.
Finally, we investigated whether HLOs could be expanded with passaging in basal medium with 1% FBS used from D8 (Fig. 1A). qPCR data revealed that P21 (CDKN1A), G1 cell cycle inhibitor [22], was unregulated from D21 to D41 (Supplementary Fig. S1F, *P ≤ 0.05, n = 3). Meanwhile, proliferation and cell cycle gene, including PCNA, CDK4, and Cyclin E1, were downregulated (Supplementary Fig. S1F, *P ≤ 0.05, n = 3). The expression level of apoptosis-related gene BAD [22] was unregulated (Supplementary Fig. S1G, *P ≤ 0.05, n = 3). Collectively, these findings indicated that this medium without additional cytokines/small molecules could not in the long-term support the growth of HLOs in vitro. In fact, several key lung lineage markers, including NKX2.1, SOX9, and SPC, could not be detected at D100 (Supplementary Fig. S1H). Therefore, we transplanted HLOs into immunocompromised mice to test whether HLOs could expand and mature in vivo.
Short-term transplanted D21 HLOs contained lung progenitor cells
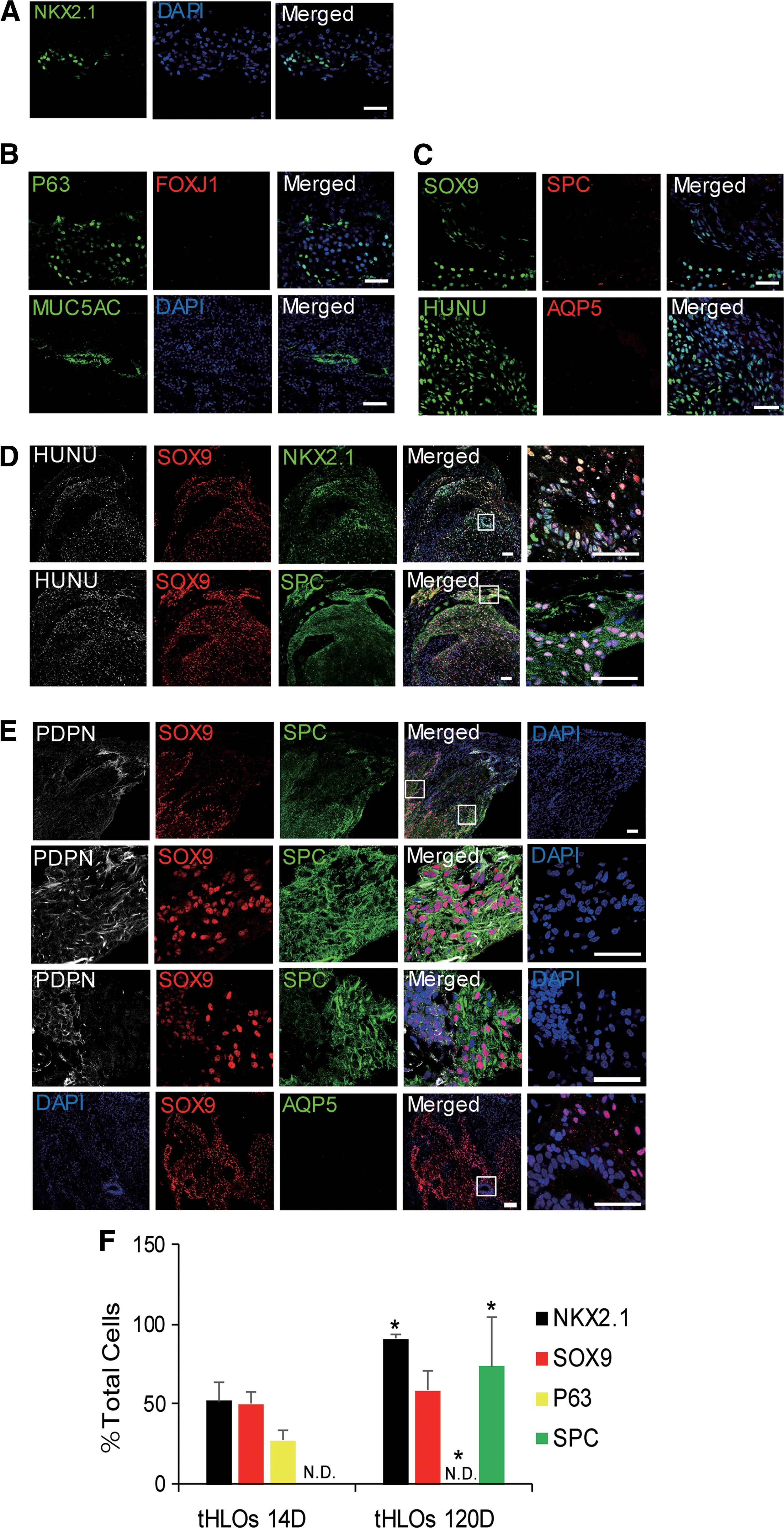
Previous studies have demonstrated that intestinal, kidney, and cerebral organoids are similar to the analogous human fetal organ [23 –25]. Organoids could mature and gain some adult structures by transplantation under the mouse kidney capsule [24,26]. Therefore, we tried to mature hESC-derived HLOs in a similar way. Since D21 is the earliest stage that lung progenitor markers (NKX2.1, SOX9, and P63) appear among all the in vitro differentiation stages checked (Fig. 1C), the stage might contain cells with high potency to differentiate into all the cell types within lung epithelium. Therefore, D21 HLOs mixed with 100% Matrigel were transplanted into three NSG mice and harvested 2 weeks later (Supplementary Fig. S2A). In contrast to the previous report, which contained no NKX2.1+ cells [13], all the three retrieved organoids in our study contained NKX2.1 positive lung lineage cells (Fig. 2A). Lung proximal cell types, specifically P63+ basal cells and MUC5AC+ goblet cells, but not FOXJ1+ ciliated cells, were evidenced (Fig. 2B). And distal cell types, specifically SOX9+ distal progenitor cells, but not SPC+ AT2 cells or AQP5+ AT1 cells, were also found (Fig. 2C). Therefore, the 2-week transplanted HLOs (tHLOs) were maintained at a less differentiated stage.
Long-term transplanted D21 HLOs possess BP cells.
Long-term transplanted D21 HLOs possess BP cells
As most cell types in the 2-week transplanted HLOs are progenitor cells, extending transplantation might promote cell differentiation and maturation. Therefore, HLOs at D21 were engrafted and harvested 100–120 days later (Supplementary Table S3). The long-term transplanted HLOs grew obviously (Supplementary Fig. S2A). No tubular structure was observed (Supplementary Fig. S2B). The tHLOs contained abundant NKX2.1+ lung lineage cells and SOX9+ distal progenitor cells (Fig. 2D). E-cadherin, an epithelial-specific marker, was co-expressed with SPC (Supplementary Fig. S2C). Most SPC+ cells co-expressed distal epithelial marker SOX9 (Fig. 2D). The SPC and SOX9 double positive cells resembled a population of cells localized to the distal epithelial tips that might finally differentiate into alveolar cells in human [9,27]. Meanwhile, further analysis revealed that most PDPN+ cells co-expressed SOX9 and SPC (Fig. 2E), resembling the BP cells (SPC+/PDPN+/SOX9+) at saccular stage in mouse lung development [10]. Interestingly, part of PDPN+ cells weakly co-expressed SOX9 and SPC (Fig. 2E). We speculated that the PDPN+/SOX9low/SPClow cells were the early AT1 progeny of BP cells which would finally mature into AT1 cells. To check whether tHLOs proceeded to a more mature AT1 development stage, AQP5 was examined by immunostaining. AQP5 (aquaporin 5) is a water channel protein, which plays an important role in transporting water across the air space-capillary barrier in the distal lung [28], and it is a functional marker of mature AT1 cells [1,10,27,29] However, we failed to detect any AQP5+ cells (Fig. 2E).
The proximal cell types in retrieved tHLOs were also examined. Club cells expressing CC10 (encoded by the SCGB1A1 gene) were observed (Supplementary Fig. S2D). Very few cells expressed the goblet cell marker MUC5AC, and no cells expressed the basal cell marker P63 (Supplementary Fig. S2D), while rare multiciliated cells could be identified by TEM (Supplementary Fig. S2E). Finally, we quantified and compared the NKX2.1+, SOX9+, P63+, and SPC+ cells between 14D- and 120D-retrieved tHLOs and found that the latter contained more SPC+ alveolar cells and less P63+ proximal cells (Fig. 2F, *P ≤ 0.05, n = 3). In summary, the present results indicated that long-term transplanted D21 HLOs promote BP (SPC+/PDPN+/SOX9+) cell differentiation.
Long-term transplanted D41 HLOs possess mature AT1-like cells
In principle, different seed cells might give rise to different cells in vivo. Along in vitro differentiation, the expression of lung progenitor and stem cell genes NKX2.1, SOX9, SOX2, and P63 reached peaks at D21 or D31 and downregulated at D41 (Fig. 1C). On the contrary, genes of distal alveolar epithelial cells (SPC, SPB, and HOPX) were upregulated at D41 HLOs (Fig. 1C). Therefore, we hypothesized that D41 and/or D31 HLOs might differentiate into more mature AT2 and/or AT1 cells than D21 HLOs after long-term transplantation. To this end, HLOs at D31 and D41 were engrafted and harvested 100–120 days later (Supplementary Table S1). Similar to D21 tHLOs, D31 and D41 tHLOs grew obviously (Supplementary Fig. S3A, B) and no tubular structure appeared (Supplementary Fig. S3C). The epithelial-specific marker EpCam was expressed in tHLOs, indicating successful differentiation of epithelial cells (Supplementary Fig. S3D). Distal alveolar cell lineages were also more efficiently differentiated than proximal cell lineages in D31 and D41 tHLOs (Fig. 3A–C and Supplementary S3E, F). However, different from D21 and D31 tHLOs which contained no AQP5+ cells, D41 tHLOs possessed abundant HUNU+/AQP5+/SOX9− mature AT1-like cells (Figs. 2D and 3B, C). Since SPB is an important functional marker of mature AT2 cells [1,10,27,29], we co-stained SPB, AQP5, and SOX9 in D41 HLOs and identified two cell populations, AQP5+/SPB−/SOX9− cells and SPB+/SOX9+/AQP5− cells (Fig. 3C). Interestingly, these two populations resembled mouse mature alveolar AT1 and AT2 cells, respectively [1,10]. Furthermore, mature AT2 cells were also evidenced with visualization of lamellar body organelles by TEM (Fig. 3D). Finally, we quantified and compared the NKX2.1+, SOX9+, SPC+, and AQP5+ cells between D21, D31, and D41 tHLOs and found that D31 and D41 tHLOs contained less NKX2.1+ and SOX9+ progenitor cells and that D41 tHLOs were the only ones containing AQP5+ cells (Fig. 3E, *P ≤ 0.05, # P ≤ 0.05, n = 3). All the above results suggested that D41 tHLOs could differentiate into mature AT1-like (PDPN+, AQP5+/SPB−/SOX9−) cells and AT2 (SPC+, SPB+/SOX9+/AQP5−) cells and that D21 tHLOs could differentiate into BP cells (SPC+/PDPN+/SOX9+) after long-term engraftment.

Long-term transplanted D41 HLOs possess mature AT1-like cells.
Transplanted HLOs possess vascular network and neuronal network
Native airways are surrounded by mesenchymal cells, which express high level of collagen I [30,31]. The tHLOs showed that collagen I+ cells were close to SOX9+ distal progenitor cells (Fig. 4A). It has been reported that transplanted HLOs with the support of biomaterial PLG scaffold were associated with abundant vasculature [13]. Consistently, we observed vasculature using ACTA2 staining (Fig. 4B, C), a vascular smooth muscle cell marker [32 –34]. The vasculature was also evidenced by 3D image reconstitution (Supplementary Video S1 and Fig. 4C). Furthermore, several clusters of neuroendocrine-like cells were observed by neuroendocrine marker PGP9.5 [7,35] staining (Fig. 4D and Supplementary Video S2), which resembled pulmonary neuroendocrine bodies, unique structures in lung to function as airway sensors [36,37]. During the early development of the lungs, a complex neuronal network is laid out. Previous study has demonstrated that elimination of neural cells abolishes lung branching and diminishes endothelial and epithelial cell proliferation [38]. Other studies have revealed that nerves are a critical component of the progenitor cell niche during organ development [39]. In the present study, blood vessel structure and myelin sheath structure of nerve fiber in tHLOs were also evidenced by TEM (Fig. 4E). Collectively, our findings suggested that tHLOs possessed vascular network and neuronal network resembling human lung.
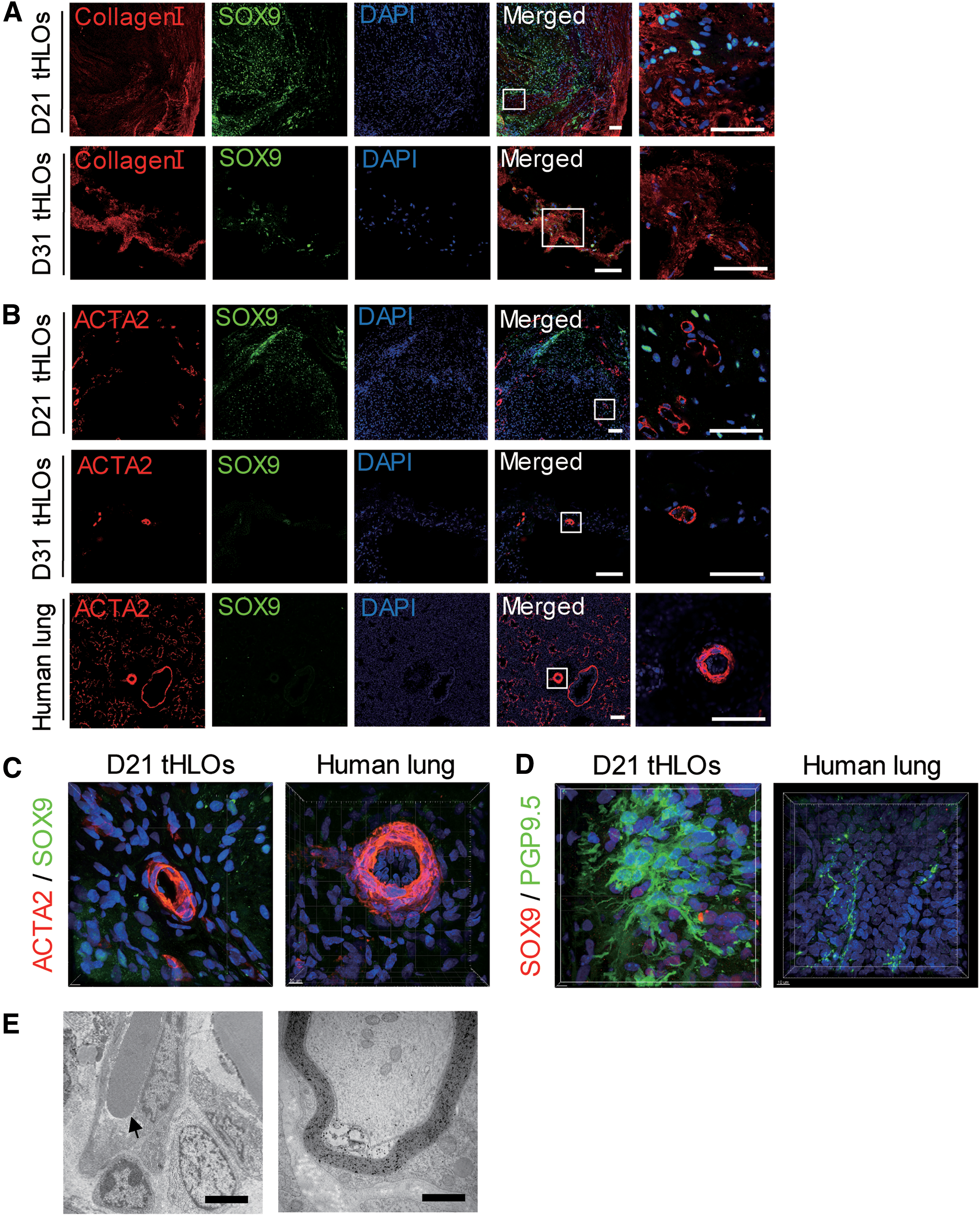
Transplanted HLOs possess vasculature and neuroendocrine-like cells.
Discussion
In this study, we established an efficient method to induce alveolar epithelial cells from hESCs using HLOs. Several in vitro differentiation methods have been reported to derive lung lineage from hESCs (Table 1). These protocols are based on 2D monolayers [7,40,41] or 3D HLOs [5,7 –9,17,42], which use different cytokines and small molecules and consume different time. And four groups reported that the differentiated 3D HLOs could be passaged in vitro. Compared with those reported methods, our protocol is simple and uses less cytokines and small molecules and therefore costs less.
N.D. means that was not mentioned in the article. Cytokines/small molecules mean all cytokines and small molecules used for hPSC derived lung lineage induction. If the same cytokine or molecule was used in two induced stage, it was counted 2. For example, as it was mentioned in the Methods in the present study, CHIR99021 was first used for definitive endoderm (DE) induction from hESCs, then it was used again for further anterior foregut stage induction, it was counted as 2. For several reports used Definitive Endoderm Kit for DE induction, it was counted as 1 cytokine. The present study was indicated in boldface.
hPSC, human pluripotent stem cell.
To date, no study can generate alveolar structures by ectopic transplantation of human pluripotent stem cell derived HLOs [7,13] (Table 2). During lung development, the pressure of the fluid against the lung tissue may provide a mechanical stimulus that promotes lung growth [43,44]. Studies revealed that the extent of amniotic fluid and lung fluid correlated with fetal lung volume [45]. More recent study revealed mechanical forces that have an essential function in promoting alveolar progenitor cell fattening [46]. Moreover, alveologenesis arises from the fact that the process is coordinated by multiple interactions through the fibroblastic, epithelial, and microvascular lung components and with extracellular matrix (ECM) [47]. The lung's unique ECM plays critical role in lung development by providing essential biochemical and biomechanical cues directing morphogenesis [48]. Alveolar septation and maturation was accompanied by marked changes in lung ECM composition [49]. It is known that the relative lung contents of collagen and fibronectin markedly increase coincidentally with alveolar septation [47,50]. Tenascin-C, an ECM protein, is required for the process of alveolar septation [51]. Based on these reasons, generation of alveolar structures by HLO ectopic transplantation is challenging. Only one group reported that transplanted 3D HLOs could develop into proximal and distal cell lineages [7], but AT1 cells were immature (AQP5−) (Table 2). However, our study generates mature AT1-like cells (PDPN+, AQP5+/SPB−/SOX9−) (Table 2). Furthermore, we also generate hESC-derived lung BP cells (SPC+/PDPN+/SOX9+) for the first time.
N.D. means that was not mentioned in the article. The present study was indicated in boldface.
Recently, SOX9 and SPC have been reported to be expressed in human lung bud tip progenitor cells, which will give rise to alveolar cells at late development stages [9,27]. In mouse lung development, the tip progenitor cells differentiate into BP progenitors and then AT1 and AT2 cells from canalicular stage to saccular stage [1,25,52,53]. In our system, tip progenitors (SOX9+/SPC+) and BP progenitors (PDPN+/SPC+/SOX9+) could be found in D21 and D31 tHLOs, while all the four types of cells could be found in D41 tHLOs. Thus D21 and D31 tHLOs might reflect a development time point between canalicular stage and saccular stage, and D41 tHLOs might reflect the saccular stage. Therefore, hESC-derived HLOs in our system might be useful to model human lung development.
In summary, long-term engraftment could promote distal airway AT2 and mature AT1-like cell differentiation in hESC-derived HLOs. Different seed HLOs could give rise to cell types at different lung development stages. tHLOs possessed vascular network and neuronal network resembling human lung. These findings indicate that hESC-derived HLOs provide a promising model to study human lung development. Future studies might focus on optimizing the protocol to direct the cell outcome and use the differentiated cells for cell therapy.
Footnotes
Acknowledgments
The authors thank Rong and Lin lab members for helpful discussions. The research was funded by the National Natural Science Foundation of China (81670093 and 81372494), the Program of Department of Science and Technology of Guangdong Province (2014B020212018), the Natural Science Foundation of Guangdong Province (2017A030310331), the Program of Guangzhou Science Technology and Innovation Commission (201508020120), and the Thousand Young Talents Program of China.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.