
Abstract
Several studies show that mesenchymal stem cells (MSCs) homing to tumors not only provide the microenvironment for tumor cells but also promote tumor growth and metastasis. However, the exact mechanism remains unclear. Our study aims to investigate the role of gastric cancer MSCs (GCMSCs)-derived IL15 during GC progression. The effects of IL15 secreted by GCMSCs on GC development were evaluated by detecting the stemness, epithelial–mesenchymal transition (EMT), and migration abilities of GC cell lines. The expression of IL15 in serum and tissues of GC patients was also assessed. We found that IL15 derived from GCMSCs enhanced stemness, induced EMT and promoted migration of GC cell lines. The level of IL15 was higher in GC patients both in serum and tissues compared with that in healthy donors, which was associated with lymph node metastasis. In addition, the results have shown that IL15 in GC microenvironment was mainly produced by GCMSCs. Moreover, IL15 upregulated Tregs ratio through activation of STAT5 in CD4+T cells was accompanied by elevated expression of programmed cell death protein-1 (PD-1). Our data proved that the high concentration of IL15 in tumor microenvironment, which was mainly secreted by GCMSCs, may contribute to tumor cell metastasis and offer a new opportunity to develop effective therapeutics for intercepting tumor progression.
Introduction
G
The cause and progress of GC involves complicated steps and various factors, in which tumor microenvironment (TME) has a vital role [3]. Besides tumor cells, TME stromal cells includes fibroblasts, endothelial cells, mesenchymal stem cells (MSCs) and immune cells. Among them, tumor-associated MSCs have caused more attention among these stromal cells [4]. Inflammatory TME recruit MSCs from the circulation and adjacent tissues to tumor tissues, educating these MSCs to stimulate angiogenesis, promote epithelial–mesenchymal transition (EMT), tumor growth, and metastasis. Tumor-associated MSCs can also recruit immunosuppressive cells and finally result in the formation of an immunosuppressive microenvironment. It has been shown that paracrine factors secreted by MSCs were involved in the formation of this vicious cycle [5].
IL15 is a pleiotropic cytokine of the four α helix bundle family. Upon binding to a heterotrimeric receptor complex, which consists of IL15Rα (CD215), IL2Rβ (CD122), and the common γ (CD132) chains, IL15 activates downstream signaling pathways and then leads to proliferation of T, B, and natural killer (NK) cells. As is reported, IL15 enhances antitumor effects. However, as a growth factor for immune cells and also for tumor cells, IL15 functions as an important pathogenic factor of hematological malignancies [6 –8]. It has been rare to report that IL15 has a tumor-promoting effect in solid tumors. IL15 mainly functions in a cell contact-dependent manner called trans-presentation, in which IL15Rα, expressed at the surface of IL15 producer cells, presents IL15 in trans to responder cells bearing the IL15Rβ/γ. However, signaling by cis-presentation also contributes to IL15-induced responses, in which IL15 is presented by IL15Rα to IL15Rβ/γ expressing on the same cell [9 –12]. Once capturing IL15, IL15Rβ/γ stimulates intracellular signaling activation through JAK/STAT pathway [13]. Tosiek et al. found that IL15 upregulated Foxp3 by enhanced p-STAT5 signaling and supported Tregs stability, while negatively regulated expression of RORγt and Runx1 in Th17 cells [14]. Wang et al. proved that IL15 derived from small-cell lung cancer tumor cells could induce Tregs production, and the significantly worse survival rate was related with higher ratio of Foxp3+ cells infiltration in tumor [15].
In our previous study, we found that MSCs secreted soluble molecules or exosomes promoted angiogenesis and favored tumor growth and metastasis [16 –18]. However, the exact mechanism by which MSCs promote tumor remains unknown. In this study, we aim to investigate the role of GCMSCs-derived IL15 in GC progression. The results revealed that IL15, as a pleiotropic cytokine, which was mainly secreted by GCMSCs in GC microenvironment, upregulated Tregs ratio and enhanced programmed cell death protein-1 (PD-1) expression in CD4+T cells to contribute to tumor metastasis.
Materials and Methods
MSCs, cell lines, and cell culture
GC tissues were obtained from GC patients of the Affiliated People's Hospital of Jiangsu University (Zhenjiang, China). The procedure was approved by the Ethics Committee of Jiangsu University. The informed consents were obtained from all subjects. GCMSCs were isolated from human GC tissues as previously described [19]. In brief, fresh tissues were cut into about 1 mm3-sized pieces and adhered to 35 mm cell culture dishes (Corning) and were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) with 10% fetal bovine serum (FBS; Gibco) at 37°C in humid air with 5% CO2. The culture medium was refreshed every 3 days. When the confluence of fibroblast-like cells reached about 70%, the cells were further trypsinized and passaged into a cell culture flask (Corning) for further expansion up to five passages and used for the subsequent experiments.
Bone marrow (BM) MSCs were obtained from healthy donors and this procedure was approved by the Ethics Committee of Jiangsu University. The BM cells were diluted with equal volume of phosphate-buffered saline (PBS) and separated over a gradient of 1.077 g/mL Ficoll (Dakewe), washed with PBS and cultured in DMEM with 10% FBS at 37°C in humid air with 5% CO2. After about 10 days, the adherent cells were trypsinized and passaged up to five passages for use.
The human GC cell lines (MGC-803 and SGC-7901), nonsmall-cell lung cancer cells (A549), and breast cancer cells (MDA-MB-231) were obtained from the Chinese Academy of Sciences Type Culture Collection Committee Cell Bank (Shanghai, China). MGC-803, A549, and MDA-MB-231 were cultured in DMEM with 10% FBS and SGC-7901 was cultured in RPMI 1640 (Gibco) with 10% FBS at 37°C in humid air with 5% CO2.
Peripheral blood mononuclear cells (PBMCs) from healthy donors were isolated by Ficoll gradient centrifugation. The subpopulation of CD4+T cells and CD8+T cells were further purified by human CD4+T Cell Isolation Kit (Miltenyi Biotec) and CD8+T Cell Isolation Kit (Miltenyi Biotec), respectively. The purity of T cells determined by flow cytometry was more than 95%. PBMCs, CD4+T cells, and CD8+T cells were cultured in RPMI 1640 with 10% FBS and preactivated in wells coated with CD3 and CD28 functional antibodies before use.
Condition medium of GCMSCs preparation
GCMSCs were cultured in cell culture flasks and the culture medium was refreshed when the confluence reached about 70%. After 48 h, cell culture medium was harvested, then centrifuged at 1,000g for 5 min to remove cells and filtered through a 0.22 μm membrane (Millipore) and stored at −80°C until use.
PBMCs treated with condition medium of GCMSCs
RPMI 1640 with 10% FBS was premixed with condition medium of GCMSCs (GCMSC-CM) at volume ratio of 1:1. 1 × 106 of PBMCs were preactivated in wells coated with CD3 and CD28 functional antibodies for 48 h and then treated with the premixed GCMSC-CM for 3 days. After treatment, the supernatant of PBMCs were harvested, prepared, and stored in −80°C until use.
Adipogenic and osteogenic differentiation
GCMSCs were seeded in 35 mm plates in adipogenic differentiation medium or osteogenic differentiation medium (Biological Industries). At the end of induction, corresponding adipo-staining kit or osteo-staining kit (Biological Industries) was used to detect the adipogenic and osteogenic potential of GCMSCs, respectively.
Enzyme-linked immunosorbent assay
The concentration of IL15 in serum and culture supernatants was measured by enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems) according to the manufacturer's instructions. The concentration of IL15 was calculated based on standard curve provided with the kit.
Immunohistochemistry
Tissues were formalin-fixed and paraffin-embedded, then dewaxed in xylene, rehydrated with gradient ethanol, and treated in citrate buffer for antigen retrieval. Then samples were stained with rabbit anti-human IL15 antibody (1:200; Peprotech) or rabbit anti-human IL15Rα antibody (1:50; Proteintech) at 4°C overnight, followed by incubation in secondary biotinylated anti-rabbit antibody for 30 min at 37°C, and finally visualized with DAB solution and counterstained with hematoxylin. Each stained sample was evaluated by three senior pathologists, and five sites were selected typically. The scores of positive cell percentage were classified by four grades: 0 (<10%), 1 (10%–30%), 2 (30%–50%), and 3 (>50%). The scores of staining intensity were also classified by four grades: 0 (none), 1 (weak), 2 (moderate), and 3 (strong). Total score = positive cell percentage score + staining intensity score. Total scores 0, 1–2, 3–4, and 5–6 were defined as negative (−), weak positive (+), moderate positive (++), and strong positive (+++), respectively.
Immunofluorescence
Tissue sections were incubated overnight at 4°C with the following primary antibody: IL15 (1:1,000; Abcam), CD105 (1:1,000; Abcam), and Foxp3 (1:1,000; Abcam). Secondary antibodies (1:500; CST) were incubated for 1 h at room temperature. Sections were mounted with DAPI Fluoromount-G and imaged using a Nikon ECLIPSE Ti-S microscope.
Western blot
Cells were lysed in RIPA buffer containing protease inhibitor cocktail (Vazyme). The protein concentration of cell lysates was quantified by a BCA Protein Assay kit (Cwbio). The primary antibodies were anti-E-cadherin (1:1,000; CST), anti-N-cadherin (1:1,000; CST), anti-Oct4 (1:300; SAB), anti-Sox2 (1:400; Millipore), anti-IL15Rα (1:500; Santa Cruz), anti-IL15Rβ (1:500; Santa Cruz), anti-p-STAT3 (1:2,000; CST), anti-STAT3 (1:1,000; CST), anti-p-STAT5 (1:1,000; CST), anti-STAT5 (1:1,000; CST), and anti-GAPDH (1:2,000; Cwbio). HRP-conjugated secondary antibody (1:2,000; CST) and ECL reagent (Millipore) were used for detection of immunoreactive proteins captured by ImageQuant LAS 4000 detection system.
Migration assay
MGC-803, SGC-7901, A549, and MDA-MB-231 with different treatments for 48 h were diluted in 200 μL of serum-free medium and planted into the upper chamber with 8 μm pores (Corning). Untreated cells were set as control groups. Medium supplemented with 10% FBS (600 μL) was added to the lower chamber. Cells were incubated at 37°C in humid air with 5% CO2 for 8–12 h. The cells remaining on the upper membrane were removed with cotton wool. The cells that had migrated through the membrane were fixed with 4% paraformaldehyde, stained with crystal violet, imaged, and counted. Three experiments were performed independently.
Flow cytometry
Monoclonal antibodies against CD105, CD90, CD29 (PE-conjugated), CD45, CD34, and CD19 (FITC-conjugated; eBioscience) were used to characterize GCMSCs according to standard protocols. The PD-1 staining used human monoclonal antibody: antigen presenting cell (APC)-anti-PD-1 (BD Biosciences). To detect Tregs, Fix/Permeabilization buffer (eBioscience) was used according to the manufacturer's instructions, and cells were stained with FITC-anti-human CD4 (eBioscience) and PE-anti-human Foxp3 (BD Biosciences). Data were acquired by a flow cytometer (FACSCalibur; BD Biosciences).
Statistical analysis
Data are shown as mean ± standard deviation. Student's t-test, one-way analysis of variance, or nonparametric testing was used to analyze the data and P < 0.05 is considered to be significant.
Results
Characterization of GCMSCs and the source of IL15 in GC microenvironment
Differentiation assay in vitro showed that GCMSCs differentiated into adipocytes and osteocytes successfully (Fig. 1A). Immunophenotype analysis displayed that GCMSCs were positive expression of CD105, CD90, and CD29; nevertheless, negative expression of CD45, CD34, and CD19 (Fig. 1B). Next, BMMSCs and GCMSCs isolated from healthy donors and GC tissues, respectively, were spindle-shaped and exhibited logarithmic growth (Fig. 1C). Besides, we compared their colony formation ability and found that GCMSCs were more capable of cloning than BMMSCs (Supplementary Fig. S1; Supplementary Data are available online at
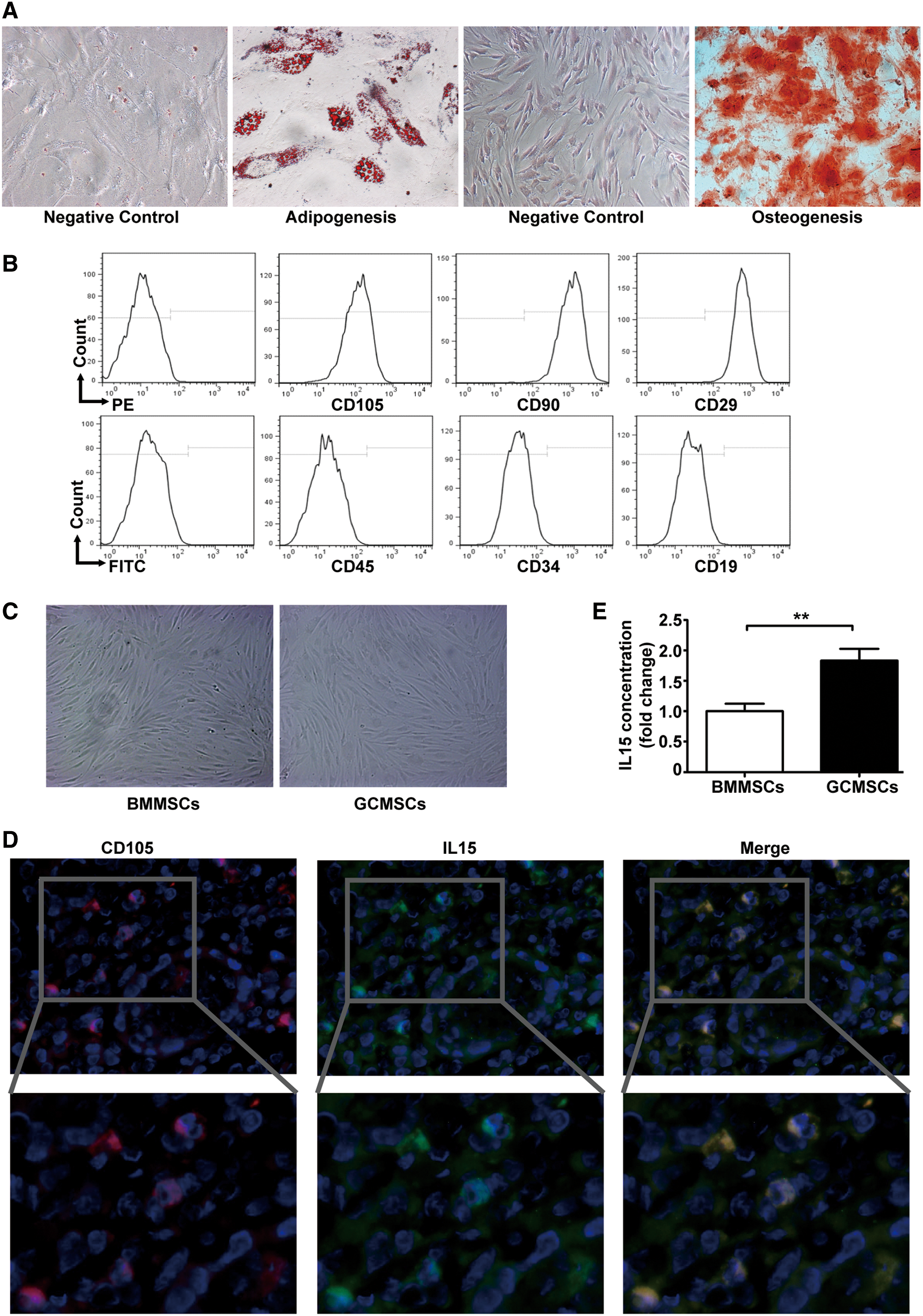
Characterization of GCMSCs and the source of IL15 in GC microenvironment.
IL15 derived from GCMSCs promotes GC cell lines migration in vitro
To reveal the effects of IL15 derived from GCMSCs on GC progression, GCMSC-CM was collected and used to culture GC cell lines in vitro. Untreated GC cell lines were set as control groups. We found that GCMSC-CM induced a significant upregulation of stem cell markers expression in MGC-803 and SGC-7901, such as Sox2 and Oct4. Moreover, GCMSC-CM also induced EMT in them. Nevertheless, these phenomena were reversed with IL15 neutralizing antibody added (Fig. 2A, B). Meanwhile, transwell migration analysis showed that the number of migration cells were more in rhIL15 and GCMSC-CM groups compared with control groups and the migration capability declined in groups with IL15 neutralizing antibody added compared with the corresponding GCMSC-CM group of MGC-803 (P < 0.05) (Fig. 2C, D) and SGC-7901 (P < 0.001) (Fig. 2E, F). These results suggest that IL15 derived from GCMSCs may promote GC metastasis.
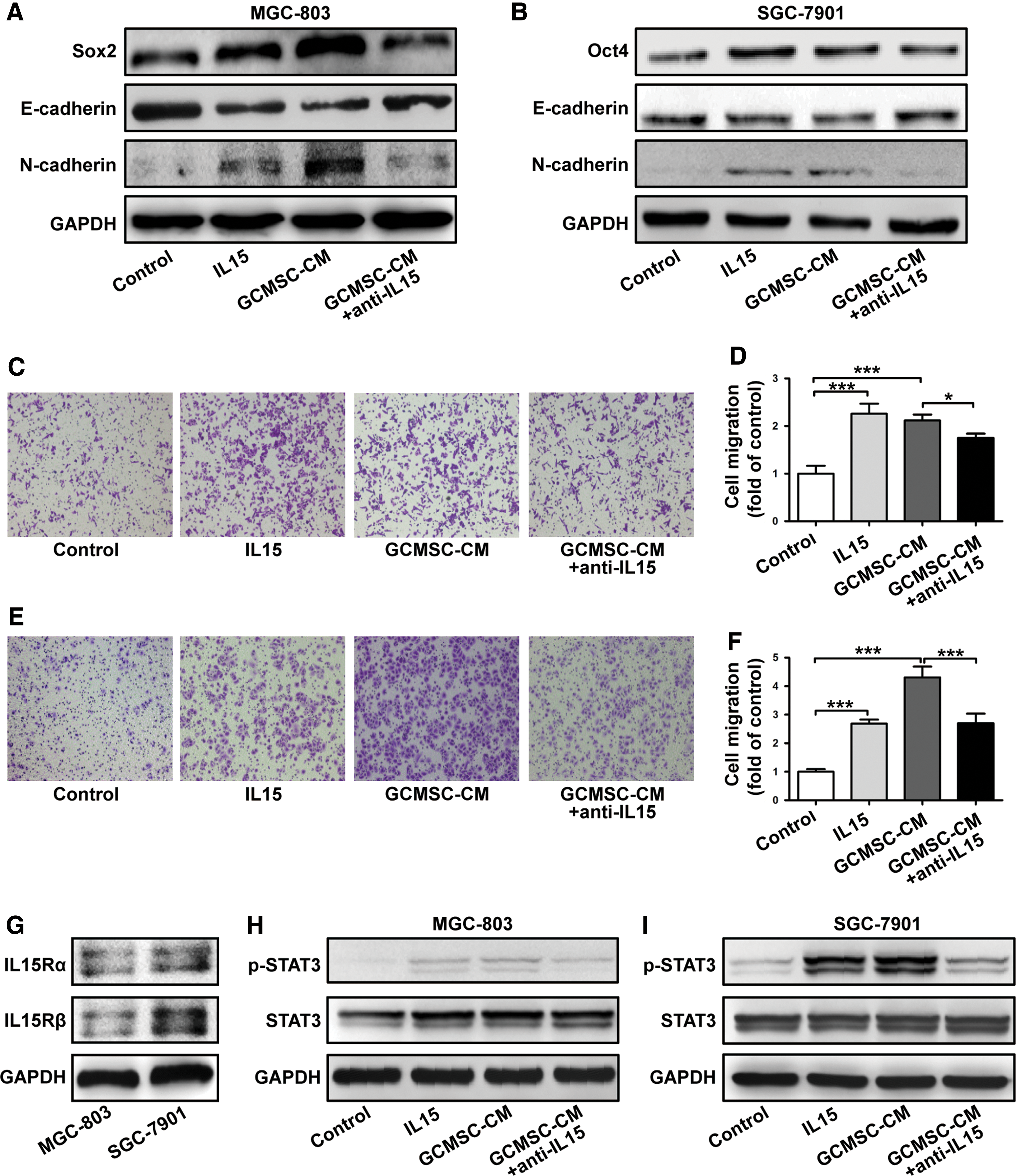
IL15 derived from GCMSCs promotes GC cell lines migration in vitro. The cells were pretreated with rhIL15 (100 ng/mL) or GCMSC-CM (with or without IL15 neutralizing antibody) for 48 h.
GCMSCs-derived IL15 regulates STAT3 activation in GC cell lines
We observed that both IL15Rα and IL15Rβ expressed on MGC-803 and SGC-7901 (Fig. 2G). Then, we found that GCMSC-CM increases the phosphorylation level of STAT3 in MGC-803 and SGC-7901, and this effect of GCMSC-CM was partially reversed by blocking IL15 (Fig. 2H, I). We also examined that the optimal concentration of GCMSC-CM and the activation time of STAT3 in SGC-7901 were 50% and 6 h, respectively (Supplementary Fig. S2). These data indicate that IL15 in GCMSC-CM may promote GC progression through STAT3 activation in GC cells. In addition, we wanted to know whether GCMSCs had effects on non-GC cells. The results have shown that IL15 derived from GCMSCs also promoted A549 migration but had no influence on MDA-MB-231, probably because A549 expressed IL15Rα, while MDA-MB-231 do not (Supplementary Fig. S3).
IL15 derived from GCMSCs effects PBMCs to promote GC cell lines migration in vitro
To analyze the synergistic action of GCMSCs and immune cells in GC microenvironment, we collected the supernatant of PBMCs, which were treated with GCMSC-CM, and then detected MGC-803 and SGC-7901 migration abilities treated with these supernatant in vitro. The results have shown that the supernatant of PBMCs treated with rhIL15 or GCMSC-CM obviously reduced E-cadherin protein levels and increased N-cadherin protein levels in MGC-803 and SGC-7901. Nevertheless, these phenomena were reversed with IL15 neutralizing antibody added (Fig. 3A, B). Next, transwell migration analysis also showed that the number of migration cells were more in PBMCs groups treated with rhIL15 or GCMSC-CM compared with control groups and the migration capability of MGC-803 and SGC-7901 declined in PBMCs groups with IL15 neutralizing antibody added (P < 0.001) (Fig. 3C–F). These results suggest that IL15 derived from GCMSCs may effect PBMCs to promote GC metastasis.

IL15 derived from GCMSCs effects PBMCs to promote GC cell lines migration in vitro. PBMCs were preactivated in wells coated with CD3 and CD28 functional antibodies before treated with rhIL15 (100 ng/mL) or GCMSC-CM (with or without IL15 neutralizing antibody). And the supernatant was collected after 3 days.
GCMSCs-derived IL15 upregulates Tregs through activation of STAT5 in CD4+T cells
To investigate how GCMSCs influence PBMCs, we treated PBMCs with GCMSC-CM. The results have shown that the CD4+Foxp3+T cells increased with GCMSC-CM treatment and declined following IL15 neutralizing antibody added (P < 0.05) (Fig. 4A, B). Activity of Tregs is closely associated with the expression of Foxp3 transcription factor. Foxp3+Tregs have immunosuppressive properties and are associated with the presence of lymph node metastasis and tumor metastasis in many cancers. We also evaluated the expression of phosphorylated (p-) STAT5 and that is, a transcription factor related to Tregs. Transcription of Foxp3 and the resulting Tregs induction are promoted by the signaling transmitted through p-STAT5, which acts downstream of the common IL15Rβ chain. As shown in Fig. 4C, both IL15Rα and IL15Rβ expressed on CD4+T cells and GCMSC-CM increased the level of p-STAT5 in CD4+T cells, which was partially reversed by blocking IL15 (Fig. 4D). We also examined that the optimal activation time of STAT5 in CD4+T cells treated with rhIL15 and GCMSC-CM were 6 and 24 h, respectively (Supplementary Fig. S4). These results suggest that GCMSCs-derived IL15 upregulates Tregs through activation of STAT5 in CD4+T cells.
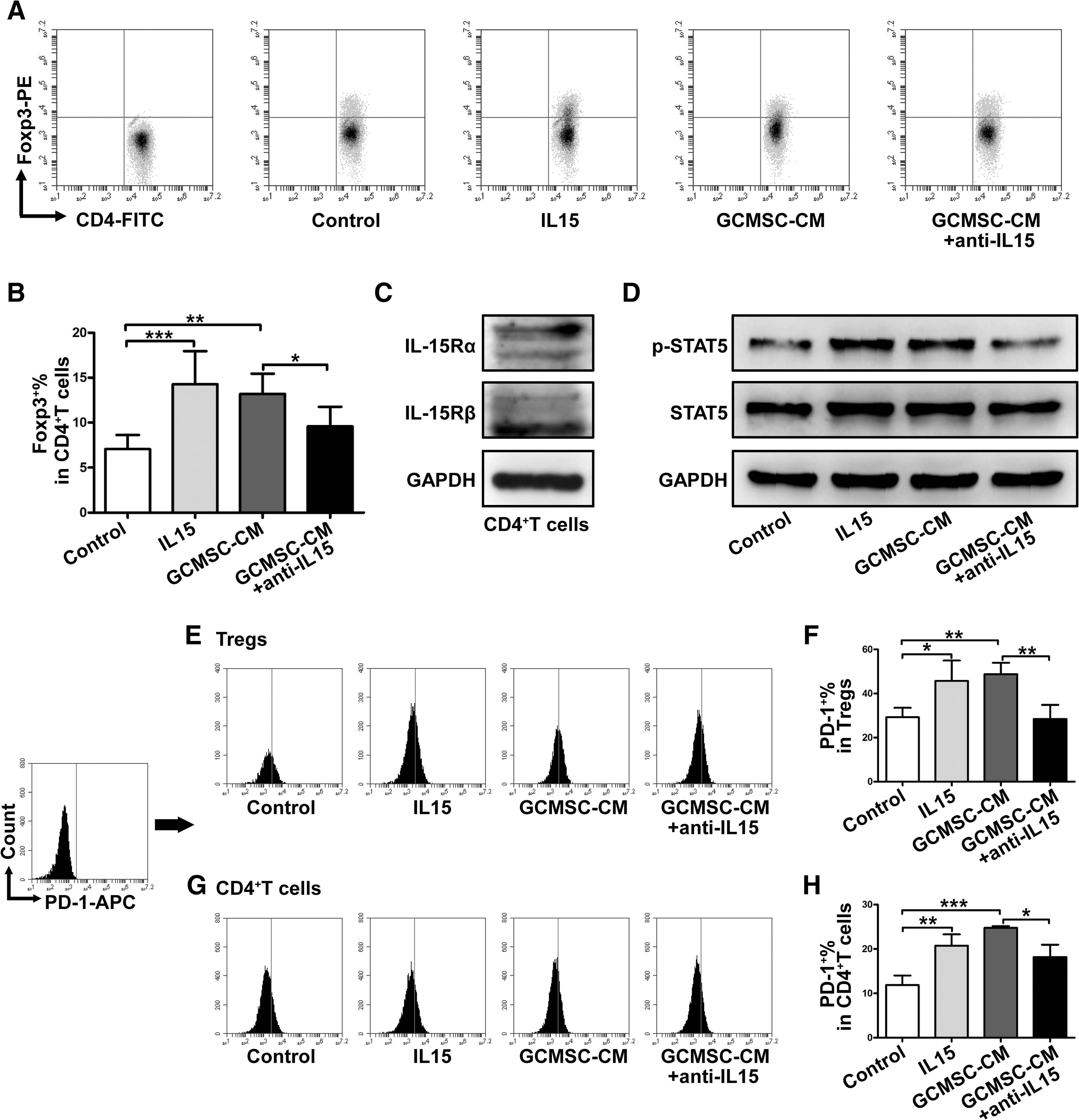
GCMSCs-derived IL15 upregulates Tregs through activation of STAT5 in CD4+T cells.
The interactions between PD-1 on T cells and PD-L1 on tumor cells or APCs mutes T cell activation and T cell-mediated tumor cell killing. Although inhibitory coreceptors are known to play important roles in the regulation of effector T cells, their functions in activated Tregs have not been well characterized. Unexpectedly, the results have shown that increased percentage of Foxp3 in CD4+T cells was accompanied by elevated expression of PD-1 in Tregs with GCMSC-CM treatment, which was downregulated after IL15 blockade (P < 0.01) (Fig. 4E, F). We also examined the PD-1 expression in total CD4+T cells, which was similar to the trend of PD-1 expression in Tregs among groups (P < 0.05) (Fig. 4G, H). However, the levels of PD-1 in total CD4+T cells were lower than that in Tregs, suggesting that PD-1 mainly expressed in the Tregs subset of CD4+T cells.
We know that CD8+T cells exhaustion is associated with high levels of expression of coinhibitory receptors, including PD-1, which is an important mechanism of tumor immune escape. Therefore, we tested the expression of PD-1 on CD8+T cells but the results have shown that there was no significant difference among each group (Supplementary Fig. S5).
IL15 may play an important role in GC metastasis in vivo
To verify the expression and the role of IL15 in vivo, the expression of IL15 was assessed in 40 GC tissues using immunohistochemistry. The intensities of IL15 immunostaining were diverse in different GC tissues (Fig. 5A) and showed positive correlation with lymph node metastasis (P < 0.01), but not associated with other clinicopathological parameters, including sex, age, tumor differentiation, TNM stage, and so on. (Table 1). The serum level of IL15 was also detected in 37 healthy controls (HC) and 73 GC patients. The results have shown that the level of serum IL15 in GC patients was significantly higher than that in HC (P < 0.001) (Fig. 5B). We generated receiver operating characteristic curve to evaluate the performance of IL15 as serum biomarker for the detection of GC. An area under the curve value for serum IL15 in discriminating GC patients from HC was 0.915 (Fig. 5C). Statistical analysis showed that elevated serum IL15 level in GC patients was significantly associated with lymph node metastasis (P < 0.05), but not with other clinicopathological parameters, including sex, age, tumor differentiation, tumor size, and so on. (Table 2). Moreover, we detected the expression of IL15Rα in GC tissues and adjacent normal tissues and we found that IL15Rα expression in GC tissues was stronger than that in adjacent normal tissues (Fig. 5D, E). These data verify that IL15 may play an important role in GC progression in vivo.
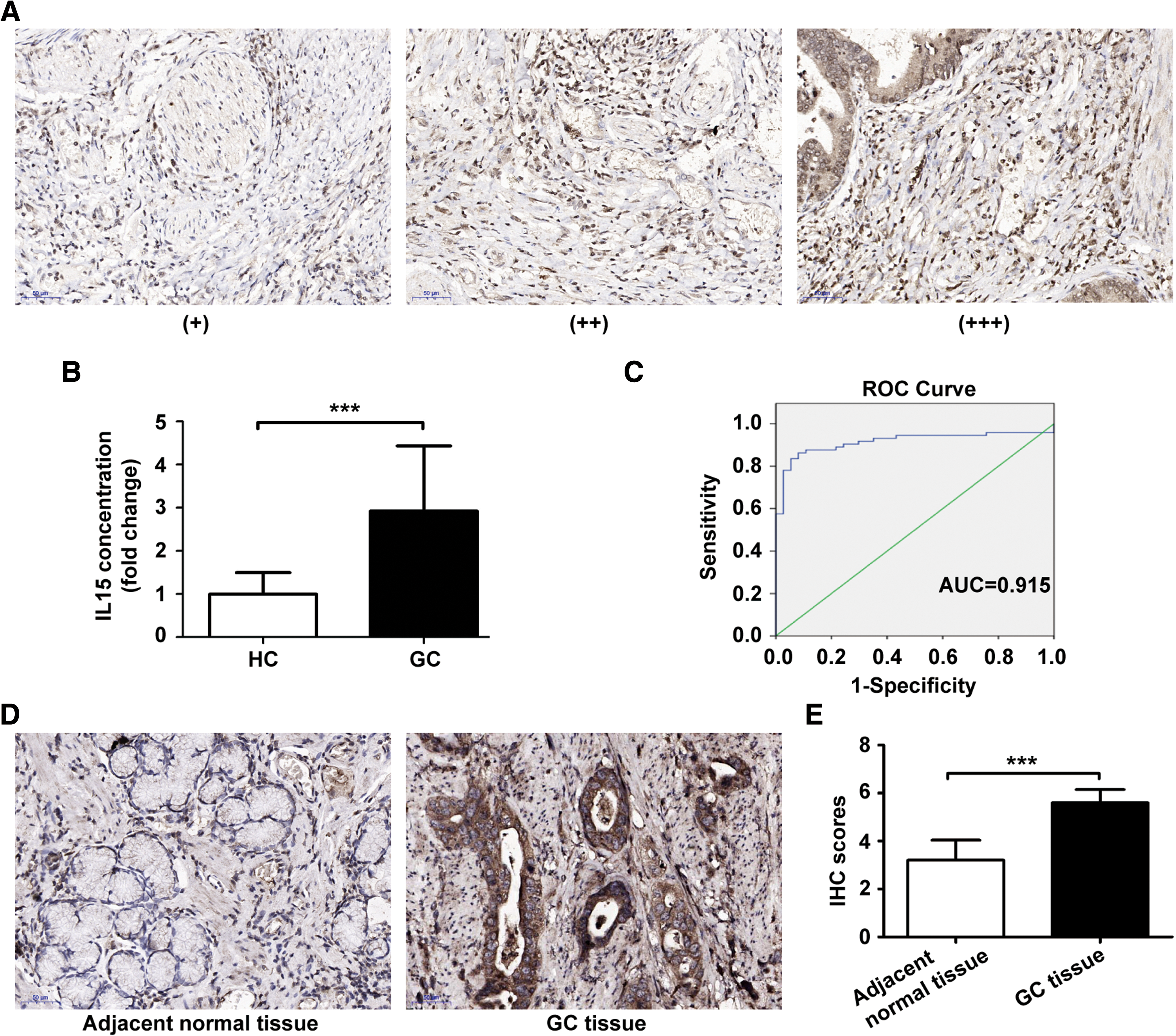
The expression of IL15 in GC patients.
Discussion
The critical role of the resident stromal cells in leading to the immunosuppressive TME was well documented [20]. Several studies have reported that MSCs can promote the growth and metastasis of solid tumors directly [18,21] or have protumorigenic effects by interfering with antitumor immune responses [22,23]. MSCs exert strong anti-inflammatory and immunosuppressive effects on immune cells through either cell–cell contact interactions [24] or soluble factors they secreted, such as IDO, PGE2, and NO [25]. Liao et al. proved that cancer-associated fibroblasts (CAFs) were related to a shift in the polarization of T cells from a T helper 2 (TH2) to a TH1 phenotype [26]. Murdoch et al. reported that CAFs reduced the recruitment of M2 macrophages, which were correlated with CD8+T cells infiltration into TME [27]. CAFs also serve as an important source of various chemokines which shape the immune landscape within the TME. CCL5 derived from CAFs has been proved to recruit Tregs compared with effector T cells preferentially and contributes to the formation of immune suppressive TME [28]. In this study, we provide evidence that GCMSCs, as important stromal cells in GC microenvironment, secreted IL15 could promote tumor cell EMT and induce Tregs ratio increase to affect GC progression. In addition, we detected whether GCMSCs had effects on non-GC cells. The results have shown that IL15 derived from GCMSCs also promoted A549 migration but had no influence on MDA-MB-231, probably because that A549 expressed IL15Rα, while MDA-MB-231 do not.
Several studies have shown that IL15 could enhance antitumor effects [29,30]. However, as a growth factor for immune cells and also for tumor cells, IL15 functions as an important pathogenic factor of hematological malignancies [6 –8]. Up to date, it has been rare to report that IL15 has a tumor promoting effect in solid tumors. Under normal physiologic conditions, IL15 is produced by cells at very low levels [31]. On the contrary, under the pathological circumstances, high-doses of IL15 have untoward side effects, for example, stimulating tumor cell growth, activating PD-1 in CD8+T cells, and functioning as an “oncogene” resulting in leukemia, which have limited its benefits in the clinic [32 –34]. In this study, we found that IL15 was abundant in tumor tissues of GC patients. Moreover, the level of serum IL15 in GC patients was significantly higher than that in healthy donors. Also, we further identified that CD105, as one of MSCs-associated markers [35], positive MSCs were the main source of IL15 in GC microenvironment.
Mishra et al. reported that prolonged in vitro exposure of wild-type large granular lymphocyte to rhIL15 (100 ng/mL) resulted leukemic transformation [7]. Our results have shown that that IL15 derived from GCMSCs promoted GC cell lines migration in vitro. In addition, serum IL15 level in GC patients and the expression of IL15 in GC tissues were significantly associated with lymph node metastasis.
IL15 mainly functions in a cell contact-dependent manner. It means that IL15 signals are delivered through trans-presentation, in which IL15 is trans-presented by its receptor, IL15Rα, expressed on the surface of producer cells, to the IL15Rβ/γ expressed on effector T, B, and NK cells [32]. This specificity is due to the capacity of the α chain to bind IL15 with a high affinity. In this study, we found that besides IL15, IL15Rα was also abundant in tumor tissues of GC patients. Once capturing IL15, IL15Rβ/γ stimulates intracellular signaling activation through JAK/STAT pathway. However, signaling by cis-presentation also contributes to IL15-induced responses, in which IL15 can be presented by IL15Rα to IL15Rβ/γ expressing on the same cell and triggers NK cell effector functions [12]. We found that SGC-7901, MGC-803, and CD4+T cells expressed both IL15Rα and IL15Rβ. Hence, we speculated that IL15 secreted by GCMSCs was captured by tumor cells or CD4+T cells and then, initiated intracellular signaling activation. Foxp3 transcription and induction are promoted by p-STAT5, which acts downstream of the common IL15Rβ. Tosiek et al. found that IL15 upregulated Foxp3 by amplifying p-STAT5 signaling and supported Tregs stability, while negatively regulated expression of RORγt and Runx1 in Th17 cells [14]. Carvalho et al. reported that abundant Foxp3+Tregs were associated with the lymph node metastasis in malignant tumors [36].
We know that the connection of CD8+T cells exhaustion and high levels of expression of coinhibitory receptors, including PD-1, is closely associated [37]. Therefore, we tested PD-1 expression on CD8+T cells, but the results have shown that there was no significant change after treated with GCMSC-CM or rhIL15. What is significant is that IL15 in GCMSC-CM-induced Tregs ratio increase was associated with the PD-1 expression on Tregs. Moreover, PD-1 expression on CD4+T cells was mainly concentrated on Tregs. Asano et al. proved that IL2 induces PD-1 expression on Tregs and the blockade of PD-1 promotes Tregs differentiation and apoptosis [38]. Therefore, it is reasonable to believe that IL15, which has been assigned to the IL2 family based on similarity in their three-dimensional structure, may affect the level of Tregs by affecting the expression of PD-1.
In summary, this study shows significant findings that IL15, as a pleiotropic cytokine, which was mainly secreted by GCMSCs in GC microenvironment, upregulated Tregs ratio and enhanced PD-1 expression in CD4+T cells to contribute to tumor cell EMT. Our findings demonstrate a new mechanism, through which GCMSCs-mediated IL15 may contribute to tumor metastasis, and offer a new opportunity to develop effective therapeutics for intercepting tumor progression.
Footnotes
Acknowledgments
This study was supported by the National Science Foundation of China (grant no. 81472334) and Jiangsu Province's Project of Key Research and Development Plan (Social Development; grant no. BE2017694).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.