
Abstract
Primordial germ cells (PGCs), the precursors of gametes, have regulatory mechanisms involving transcription factors and epigenetic modifications. The transcription factor NANOG is a key regulator of germ cell and embryonic stem cell characteristics. However, the epigenetic regulation of NANOG with a histone deacetylase (HDAC) complex in PGCs has not been studied. In this study, we investigated the epigenetic regulation, and in particular the histone acetylation, of NANOG in chicken PGCs. Intriguingly, although NANOG was highly activated in chicken PGCs, the upstream region of its promoter was moderately suppressed by histone deacetylation. HDAC inhibition induced histone H3 lysine 9 acetylation (H3K9ac) and derepressed NANOG expression. Furthermore, knockdown studies revealed that HDAC complex members, such as RE1-silencing transcription factor (REST) and REST corepressor 3 (RCOR3), are important epigenetic modulators of NANOG expression in chicken PGCs. We demonstrate that moderate regulation of NANOG by the REST/CoREST/HDAC complex might be crucial for maintaining the integrity of PGCs.
Introduction
P
Specifically, histone H3 lysine 9 acetylation (H3K9ac) transiently and markedly increases in PGCs to establish transcriptional activation of chromatin at the point of entry into the genital ridge and decreases during colonization [3]. Also, maintenance of H3K9ac is involved in proper chromosomal segregation during the formation of gametes [4,5]. Thus, histone acetylation is a crucial epigenetic marker of germ cell specification and development.
Avian PGCs are important for biotechnology and germ cell biology due to their unique characteristics [6]. Chicken PGCs are specified by maternally inherited germ plasm, including VASA and deleted in azoospermia like (DAZL), in a predetermined manner [7,8]. In contrast with mammals, chicken PGCs migrate to the genital ridge through blood vessels after specification [9].
In terms of epigenetic regulation, the dynamics and landscape of histone modification have not been established yet in avian PGCs. In recent study, global enrichment of H3K9 trimethylation (H3K9me3) was observed in chicken PGCs [10]. Also, H3K9ac formed foci in euchromatin in chicken PGCs as the activating histone acetylation marker [10], indicating this mark could play an important role in epigenetic regulation in chicken PGCs. However, gene expression regulation by histone modification in chicken PGCs has been largely unknown.
NANOG is an important transcription factor that regulates pluripotency in early embryos, embryonic stem cells (ESCs), and induced pluripotent stem cells (iPSCs) [11]. In mammals, NANOG expression was also found from migrating germ cells to gonadal germ cells [12,13], and shown to prevent apoptosis of migrating germ cells [14]. Moreover, in later stage, NANOG is involved in differentiation and maturation of mouse PGCs in genital ridge [15 –17]. In chicken, NANOG is required for the maintenance of pluripotency in chicken ESCs [18] and was detected during early embryonic development from preovulatory oocyte to intrauterine stages [19]. Also, Jak1/Stat3 signaling supports NANOG expression and pluripotency in early chicken embryos [20].
In chicken PGCs, NANOG is expressed in the germinal crescent at Hamburger and Hamilton stage 5 (HH5) to HH8 [21]. In addition, NANOG expression was similar to the PGC-specific genes CVH and dead end homolog 1 (DND1) during PGC migration and colonization in gonads [21], indicating a role for NANOG in maintaining the self-renewal ability of chicken PGCs in various stages. The recent report showed that NANOG solely activated enhancers upstream of germ cell specifiers to generate PGC-like cells from epiblast-like cells, but not in ESCs in mouse [22], indicating that its function differs according to cell type.
The role of histone acetylation and regulation of the histone deacetylase (HDAC) complex in pluripotency and somatic cell reprogramming in ESCs and iPSCs has been reported [23 –25]. Despite the emerging role of NANOG in germ cell development, the role of histone acetylation and the HDAC complex in the regulation of NANOG expression in avian PGCs is unclear.
In this study, we demonstrated the epigenetic regulation of NANOG expression by H3K9ac and the HDAC corepressor complex in chicken PGCs. We used an HDAC inhibitor to induce aberrant histone acetylation and evaluated the expression of germ cell-specific genes, including NANOG. In addition, upstream regulators of NANOG, such as DNA-dependent factors and HDAC complexes, were examined to evaluate epigenetic regulation in chicken PGCs. This study increases our understanding of the molecular mechanisms involved in the fundamental specification of germ cells.
Materials and Methods
Experimental animals and care
The care and experimental use of White Leghorn (WL) chickens were approved by the Institute of Laboratory Animal Resources, Seoul National University (SNU-150827-1). Chickens were maintained according to a standard management program at the University Animal Farm, Seoul National University, Korea. The procedures for animal management, reproduction, and embryo manipulation adhered to the standard operating protocols of our laboratory.
Culture of chicken PGCs and chemical treatment
Chicken PGCs were cultured in accordance with our standard procedure [26,27]. Briefly, PGCs from male WL embryonic gonads at 6 days old (HH stage 28) were maintained in knockout Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY) supplemented with 20% fetal bovine serum (FBS; Hyclone, South Logan, UT), 2% chicken serum (Sigma-Aldrich, St. Louis, MO), 1× nucleosides (Millipore, Billerica, MA), 2 mM L-glutamine (Gibco), 1× nonessential amino acids (Gibco), β-mercaptoethanol (Gibco), 1 mM sodium pyruvate (Gibco), and 1× antibiotic/antimycotic (Gibco). Human basic fibroblast growth factor (Koma Biotech, Seoul, Korea) at 10 ng/mL was used for PGC self-renewal. The cultured PGCs were subcultured onto mitomycin-inactivated mouse embryonic fibroblasts at 5- to 6-day intervals by gentle pipetting without any enzyme treatment. DF-1 was cultured in DMEM (Gibco) supplemented with 10% FBS. The cultured PGCs and DF-1 were treated with HDAC inhibitors, 0.25 mM valproic acid (VPA; Sigma-Aldrich), and 50 nM romidepsin (Selleckchem, Boston, MA), for three consecutive days.
Immunostaining
VPA- and romidepsin-treated cells were harvested and washed twice in phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde for 20 min. After permeabilization with 0.1% Tween-20 and 1% Triton X-100, nonspecific binding was blocked with 1% normal goat serum. Cells were then incubated with rabbit anti-acetyl-histone H3K9 antibody (06-942; Millipore, Bedford, MA), mouse anti-acetyl-histone H3 lysine 27 (H3K27ac) antibody (GTX50903; GeneTex, CA), rabbit anti-trimethyl-histone H3K9 antibody (07-442; Millipore), and rabbit anti-trimethyl-histone H3K27 (H3K27me3) (07-449, Millipore) at 4°C overnight. Cells incubated in the absence of primary antibody were used as a negative control. After extensive washing, cells were incubated with Alexa Fluor 594-conjugated goat anti-rabbit IgG H&L (ab150080; Abcam, Cambridge, United Kingdom), Alexa Fluor 488-conjugated goat anti-rabbit IgG H&L (ab150077; Abcam), and FITC-conjugated goat anti-mouse IgG (sc-2010; Santa Cruz, CA) for 1 h. After washing, cells were mounted with 4′,6-diamidino-2-phenylindole (DAPI) and visualized using Ti-U fluorescence microscope (Nikon, Tokyo, Japan).
RNA isolation and reverse transcription-quantitative polymerase chain reaction
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. One microgram of total RNA from each sample was reverse-transcribed using the Superscript III First-strand Synthesis System (Invitrogen) according to the manufacturer's protocol. The polymerase chain reaction (PCR) mixture contained 2 μL of PCR buffer, 0.5 μL of 10 mM dNTP mixture, 10 pmole each of forward and reverse primers (Table 1), 1 μL of complementary DNA (cDNA), and 1 U of Taq DNA polymerase in a 20 μL final volume. PCR amplification involved an initial incubation at 95°C for 10 min, followed by 40 cycles at 95°C for 30 s, 60°C for 30 s, and 72°C for 30 s. PCR was terminated with a final incubation at 72°C for 5 min, and at the dissociation temperature. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was performed using a StepOnePlus real-time PCR system (Applied Biosystems, Foster City, CA) with EvaGreen (Biotium, Hayward, CA). Each test sample was assayed in triplicate. Relative quantification of target gene expression was performed using the following formula: 2−ΔΔCt, where ΔΔCt = (Ct of the target gene − (Ct of GAPDH + Ct of ACTB)/2) treatment − (Ct of the target gene − (Ct of GAPDH + Ct of ACTB)/2) control.
Histone extraction and western blot analysis
Cells were harvested and washed in ice-cold PBS for two times. Harvested cells were resuspended in Triton extraction buffer [TEB; PBS containing 0.5% Triton × 100 (v/v), 2 mM phenylmethylsulfonyl fluoride, 0.02% (w/v) NaN3]. PBS and TEB contain 5 mM sodium butyrate to preserve levels of histone acetylation. After harvest of cell pellets, the nuclear pellets were extracted with 0.2 N HCl for overnight at 4°C. Supernatants containing histone protein are neutralized by 2 M NaOH at 10% volume of 0.2 N HCl. Protein (∼10 μg) from total cell lysate was loaded in each lane for separation on 15% SDS-polyacrylamide gels. Resolved proteins were transferred onto a Hybond 0.45 PVDF membrane (GE Healthcare Life Sciences, Little Chalfont, United Kingdom) and blocked with 3% skim milk for 1 h at room temperature (Sigma-Aldrich). The blocked membrane was incubated overnight at 4°C in the presence of primary antibody (dilution: 1:3,000) and then for 1 h with horseradish peroxidase-conjugated secondary antibody (31460; Thermo Fisher Scientific, Inc., Waltham, MA) at a dilution of 1:4,000. Primary antibodies were as follows: anti-histone H3 (ab1791; Abcam) and anti-H3K9Ac antibody (06-942; Millipore). Immunoreactive proteins were visualized with the ECL western blot detection system (Thermo Fisher Scientific, Inc.). All blots were performed in triplicate, and protein levels were quantitated with ImageJ (National Institutes of Health, Bethesda, MD) [28].
Prediction of the RE1-silencing transcription factor-binding site in the proximate promoter of NANOG, POUV, and VASA
The region upstream of NANOG, POUV, and VASA transcriptional start site (TSS) was predicted by MatInspector, a Genomatix program (
Chromatin immunoprecipitation
Treated cells were fixed in 1% methanol-free formaldehyde (Thermo Fisher Scientific, Inc.) in DMEM for 20 min at room temperature, followed by 10-min blocking in 125 mM glycine. Cells were rinsed two times in ice-cold PBS and harvested. Suspension cultured cells were fixed in the plate and harvested by pipetting after washing with PBS. Fixed cell pellets were dissociated in radioimmunoprecipitation (RIPA) lysis buffer (10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 0.5 M EGTA, 1% Triton X-100, 0.1% SDS, 0.1% Na-deoxycholate, and 140 mM NaCl; Thermo Fisher Scientific, Inc.) with protease and phosphatase inhibitors (Sigma-Aldrich). Chromatin was sheared to an average length of 500–700 base pairs (bp) using a sonicator (Q500; Qsonica, Newtown, CT). DNA concentration and the efficiency of shearing were determined by agarose gel electrophoresis. An anti-H3K9Ac antibody (06-942; Millipore) was used for immunoprecipitation. Ten micrograms of sonicated chromatin samples was used. Chromatin immunoprecipitation (ChIP) was performed using a One-Step ChIP Kit (ab117138; Abcam) according to the manufacturer's instructions (ab117138; Abcam) and nonimmune IgG was used as a negative control. The bound chromatin/antibody complex was incubated in reverse crosslinking buffer for 2 h at 65°C, followed by treatment with proteinase K.
Chromatin immunoprecipitation-polymerase chain reaction and chromatin immunoprecipitation-quantitative polymerase chain reaction
For total chromatin control samples (inputs) purified before immunoprecipitation, add 10 μL of each chromatin sample to 88 μL of buffer and 2.5 μL of proteinase K. For chromatin immunoprecipitation-polymerase chain reaction (ChIP-PCR), the reaction mixture comprised 2 μL of PCR buffer, 0.5 μL of 10 mM dNTP mixture, 10 pmol of each of the forward and reverse primers (Table 2), 1 μL of ChIP DNA, and 1 U of Taq DNA polymerase in a 20 μL final volume. ChIP-PCR amplification involved an initial incubation at 95°C for 10 min, followed by 30 cycles at 95°C for 30 s, 60°C for 30 s, and 72°C for 30 s. The reaction was terminated by a final incubation at 72°C for 5 min. For chromatin immunoprecipitation-quantitative polymerase chain reaction (ChIP-qPCR), the reaction mixture, thermal conditions, and all other procedures were as stated above. ChIP-qPCR was performed using a StepOnePlus real-time PCR system (Applied Biosystems) with EvaGreen (Biotium). Each test sample was assayed in triplicate. Relative quantification of gene expression was performed using the following formula: ((2−ΔCt) × 100%), where ΔCt = (Ct [IP] − Ct [Input × DF]). The DF (default input fraction) was 1%, which is a dilution factor of 6.644 cycles (ie, log2 of 100).
Small interfering RNA transfection in chicken PGCs
Chicken PGCs were seeded at 2.0 × 105 in 1 mL of medium per well of a 12-well plate. Cells were transfected with 100 pmol of each HDAC small interfering RNA (siRNA) (Table 3) using RNAiMAX (Invitrogen). Negative control siRNA with no complementary sequence in the chicken genome was used as a control. After transfection for 48 h, total RNA was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. The knockdown efficiency of HDAC complex siRNAs and their effects on the expression of pluripotency-, germ cell-, and three germ layer-specific marker genes were assayed by RT-qPCR.
Nonspecific siRNA has no complementary sequence in the chicken genome and was used as a control for gene silencing.
siRNA, small interfering RNA.
Statistical analysis
Data are expressed as mean ± standard error from three independent biological experiments. Student's t-test was performed to evaluate differences between experimental groups using GraphPad Prism v.5 (GraphPad Software, La Jolla, CA). A value of P < 0.05 was considered indicative of statistical significance (***P < 0.001, **P < 0.01, and *P < 0.05).
Results
Upregulation of H3K9ac and gene expression in PGCs by HDAC inhibitors
To assess the influence of HDAC on epigenetic regulation and gene expression in chicken PGCs, we treated PGCs with 0.25 mM VPA, an inhibitor of class I HDACs, closely associated with transcriptional activity [29]. Treatment of PGCs with VPA in vitro resulted in a global increase in H3K9ac compared with untreated control PGCs (Fig. 1A). Next, PGCs were treated with 50 nM Romidepsin in vitro, a specific inhibitor of HDAC1 and 2 [30], which also resulted in a global increase in H3K9ac (Fig. 1A). Quantification of H3K9ac after chemical treatment confirmed the significant elevation of H3K9ac in VPA- and romidepsin-treated PGCs compared with control PGCs (Fig. 1B, C). However, romidepsin is a more effective HDAC inhibitor in PGCs compared with VPA (Fig. 1C). Therefore, only romidepsin was used for subsequent treatments. In addition to H3K9ac, we examined H3K27ac, H3K9me3, and H3K27me3 in romidepsin-treated PGCs (Supplementary Fig. S1; Supplementary Data are available online at
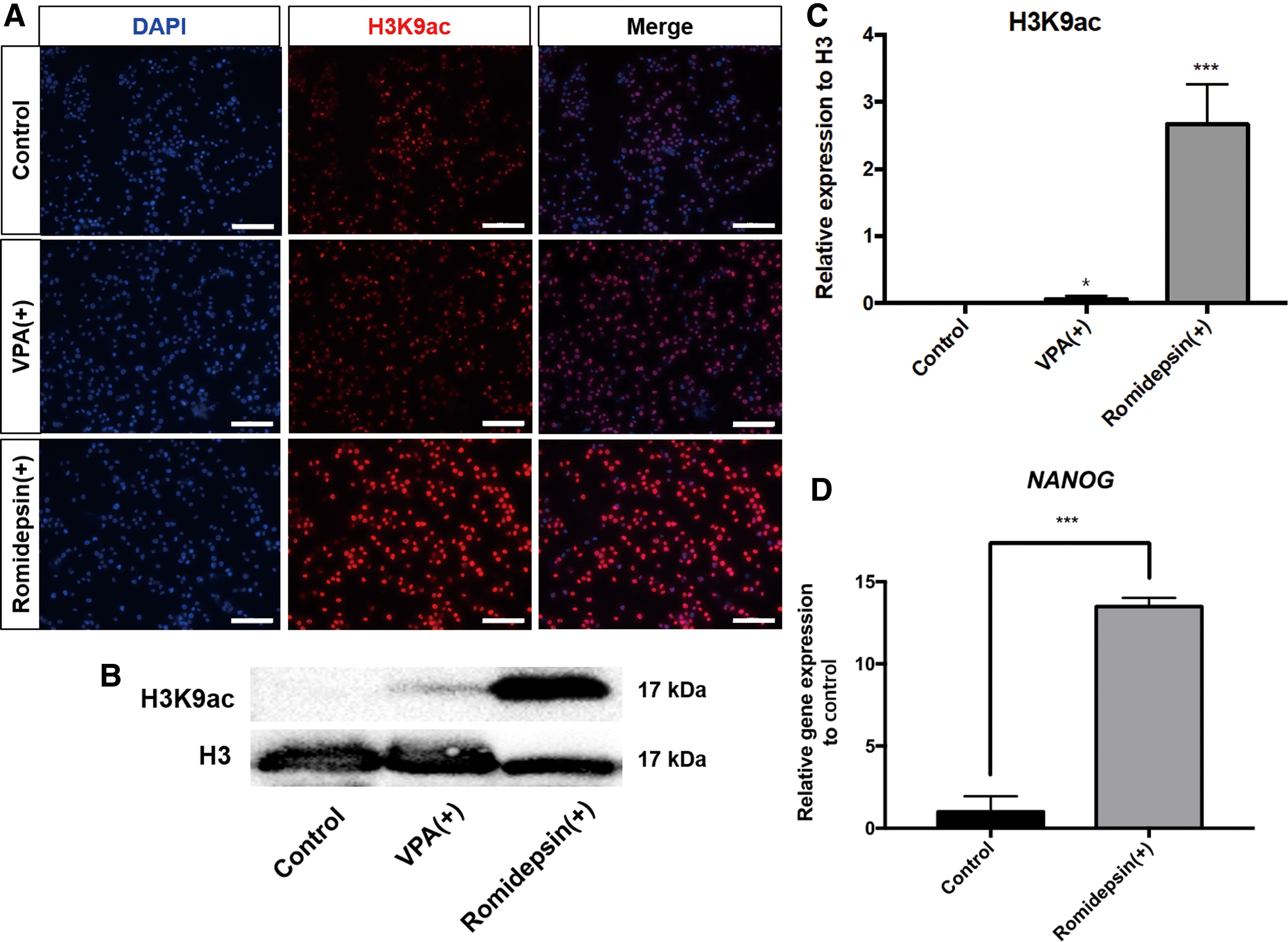
Effect of HDAC inhibitors on H3K9ac and expression of NANOG in chicken PGCs.
HDAC inhibitor increases acetylated H3K9 levels at RE1-silencing transcription factor-binding sites in the NANOG promoter
To identify the transcriptional repressor involved in modulation of NANOG expression by romidepsin, we predicted the regions affected by HDAC inhibition in the putative promoter of NANOG. Upstream of NANOG, we found a binding site of the transcriptional repressor, RE1-silencing transcription factor (REST), 188 bp from the TSS using MatInspector (Fig. 2A). Next, to examine the effect of the HDAC inhibitor on the REST-binding site, we performed ChIP analysis on this site of the NANOG promoter (188 bp from TSS). ChIP-PCR and ChIP-qPCR analyses showed that romidepsin treatment resulted in a significant increase of H3K9ac in the region nearby the REST-binding site in the NANOG promoter (Fig. 2B, C). Therefore, H3K9ac level and the expression of NANOG could be regulated by an HDAC complex possibly via the REST-binding site in chicken PGCs.

Effect of romidepsin on acetylated H3K9 levels at the REST-binding site in the NANOG promoter.
REST represses NANOG expression via H3K9ac in chicken PGCs
We next performed an expression and knockdown analysis of REST in chicken PGCs. RT-qPCR showed that REST expression was significantly higher in PGCs than DF-1 cells (Fig. 3A). Subsequently, we conducted a loss-of-function study of REST on gene expression in PGCs. REST expression was efficiently knocked down using REST-targeting siRNA. Following successful knockdown of REST (Fig. 3B), we quantified the expression of pluripotency- and germness-related genes by RT-qPCR. NANOG and VASA expression was significantly upregulated; the increase in NANOG expression was considerable. In contrast, POUV and DAZL expression was unchanged (Fig. 3C). REST-binding sites were also found in the proximal POUV and VASA promoters (Supplementary Fig. S4). Otherwise, REST knockdown increased VASA expression but not POUV expression (Fig. 3C), indicating REST to be involved in transcriptional regulation of VASA but not POUV. ChIP-PCR and ChIP-qPCR showed that suppression of REST increased H3K9ac at the REST-binding site in the proximate NANOG promoter (Fig. 3D, E). These results indicate that the REST repressor regulates the level of H3K9ac to control NANOG transcription in chicken PGCs.
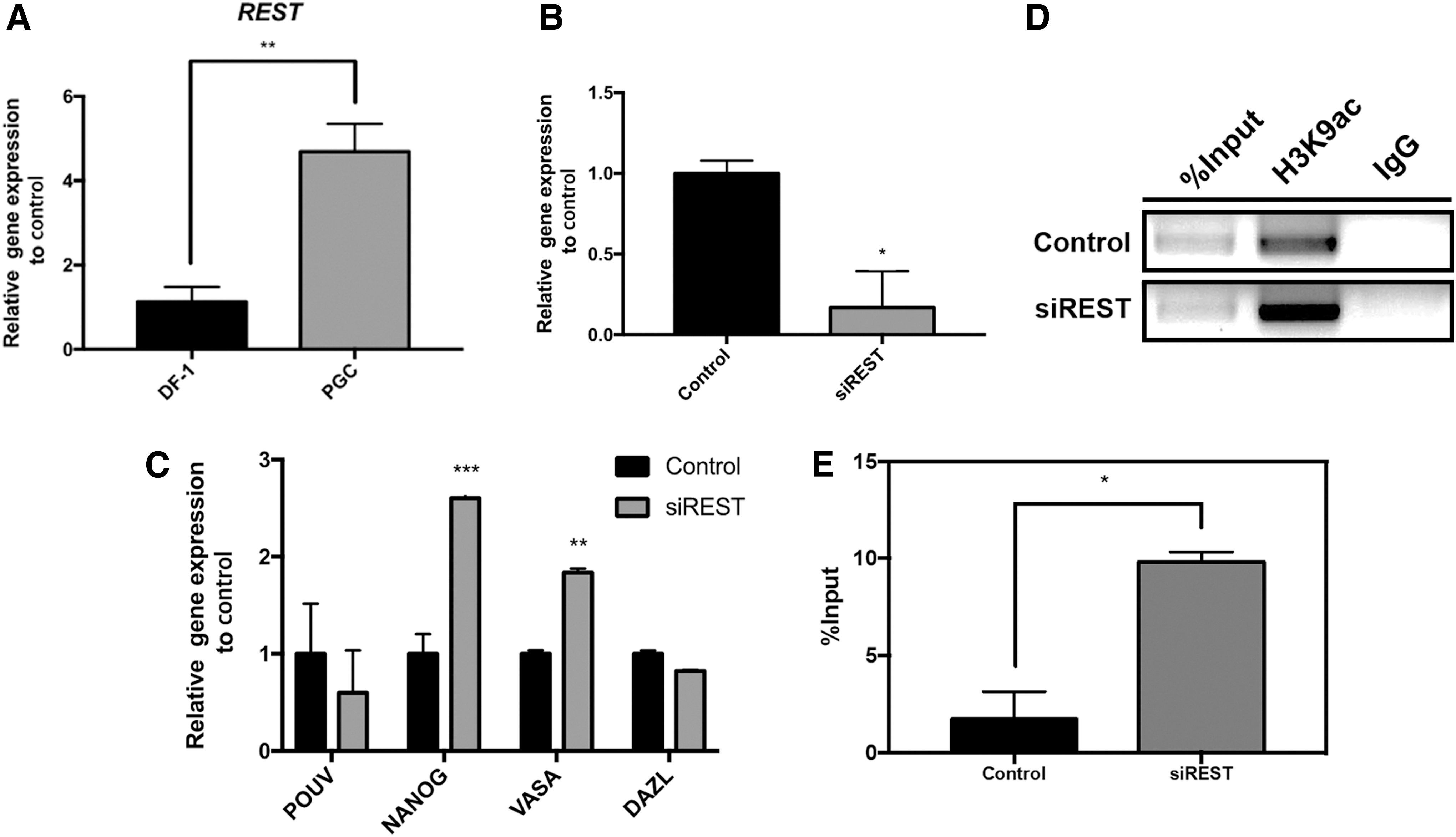
REST regulates H3K9ac to repress NANOG expression in chicken PGCs.
The CoREST complex member RCOR3 mediates epigenetic regulation of NANOG expression
Next, we elucidated the HDAC complex that mediates the change in H3K9ac in PGCs. We first determined the expression of candidate members of the HDAC complex, including metastasis-associated 1 (MTA1), MTA3, REST corepressor 1 (RCOR1), RCOR3, SIN3 transcription regulator family member A (SIN3A), and SIN3B, in chicken PGCs and DF-1 cells. Reverse transcription-polymerase chain reaction (RT-PCR) showed that all HDAC complex members, except for SIN3A, were expressed in PGCs and DF-1 cells (Fig. 4A). Second, the HDAC complex members MTA1, MTA3, RCOR1, RCOR3, and SIN3B were knocked down in PGCs using specific siRNAs. After successful knockdown of HDAC complex members in PGCs (Fig. 4B), we examined the expression of POUV, NANOG, VASA, and DAZL using RT-qPCR. Among the five members of the HDAC complex, only knockdown of RCOR3 showed significant upregulation of NANOG expression (Fig. 4C). In addition, repression of RCOR3 by siRNA enriched the acetylated H3K9 residue on the REST site in the NANOG promoter (Fig. 4D, E). These results suggest that RCOR3, together with REST, deacetylates the region upstream of the NANOG promoter, which represses NANOG expression in chicken PGCs.
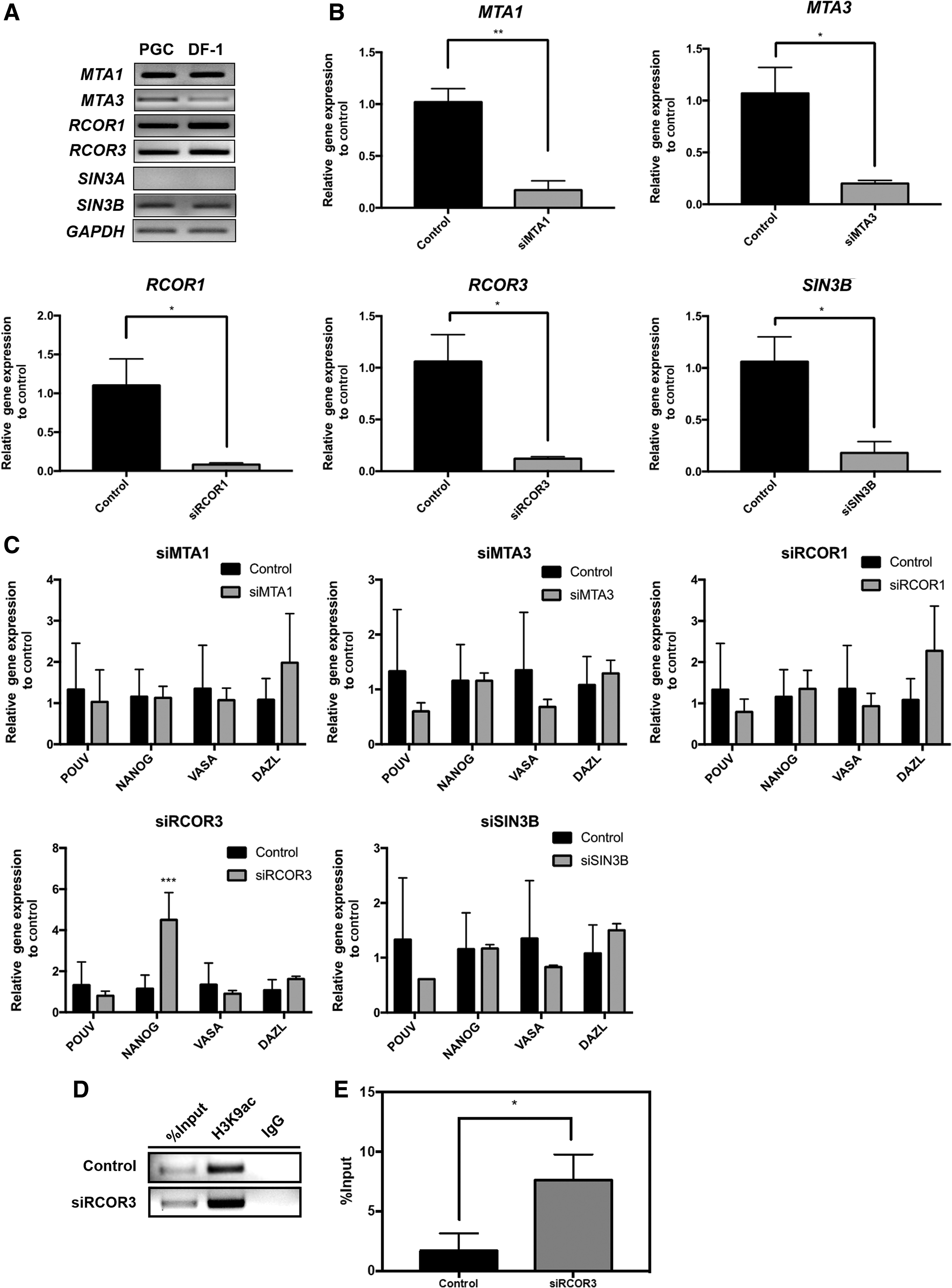
The CoREST complex member RCOR3 modulates epigenetic regulation of NANOG expression.
Discussion
NANOG is a notable homeodomain transcription factor involved in maintaining and establishing pluripotency in mammals [11,31,32]. During somatic cell reprogramming and self-renewal in ESCs, NANOG cooperates with the SIN3A/HDAC corepressor complex to activate pluripotency genes and repress reprogramming barrier genes [25]. In terms of germ cell specification from the epiblast in vitro, NANOG activates the expression of germ cell-specific transcription factors such as PRDM1 and PRDM14 as an enhancer in mouse [22], suggesting a crucial role for NANOG in germ cells. In chicken, NANOG is expressed in both ESCs and PGCs [18,21]; however, epigenetic regulation of NANOG expression by the HDAC complex in PGCs has not been investigated.
Generally, acetylation of histone tails, including H3K9ac, activates gene expression. In mouse, H3K9ac was increasing sharply in PGCs at embryonic day (E) 10.5 when entering into genital ridges, but decreasing after settlement [3]. In migrating chicken PGCs, H3K9ac was restricted in foci of loose chromatin structures, which could be involved in germ cell-specific gene expression [10]. Treatment with an HDAC inhibitor resulted in global elevation of the H3K9ac level. Indeed, treatment with the HDAC1/2-specific inhibitor romidepsin resulted in a greater elevation of the H3K9ac level, implying that HDAC1 or 2 could be the active deacetylase in chicken PGCs. Surprisingly, although NANOG, as well as POUV and VASA, is expressed at high levels in chicken PGCs, the expression was increased following the increase in H3K9ac or indirect effects induced by treatment with HDAC inhibitor. NANOG expression was also increased in romidepsin-treated chicken DF-1 cells, suggesting that the transcription of NANOG is epigenetically regulated in chicken, regardless of cell type. In contrast, the expression levels of differentiation markers were unaffected. In mammalian ESCs, treatment with low concentrations of HDAC inhibitors results in enrichment of H3K9ac to promote self-renewal and pluripotency [33,34]. However, PGCs, unlike ESCs, generate sperm and oocytes and transmit genetic information to the next generation; therefore, gene expression in PGCs should be tightly regulated. In mammals, overexpression of NANOG causes germ cell tumors [35,36]; in contrast, suppression of NANOG induces apoptosis of germ cells [14]. Correspondingly, germ cell tumor markers, such as AFP, MMP7, and NR6A1 [37], could be upregulated by NANOG overexpression due to romidepsin treatment in chicken PGCs. Also, we found that the migration-related gene in chicken, CXCR4 [38], was downregulated, indicating the possibility of aberrant migration of those treated PGCs. The misallocated cells that could not undergo apoptosis in extragonadal sites could give rise to germ cell tumors [39]. Therefore, gene expression in PGCs may need to be regulated to the proper level.
REST is a zinc finger transcription factor that represses the expression of multiple genes during embryogenesis, and regulates neuronal gene expression and neural stem cell (NSC) fate in vertebrates [40 –42]. In mouse, Rest has differential regulatory roles in ESCs and NSCs [43]. Even, Rest shared common targets with pluripotency-related factors, including Nanog [43], indicating the possible interaction between REST and NANOG in other cell types. In the case of chicken PGCs, to moderately regulate gene expression, especially NANOG, repressors such as REST seem to promote histone deacetylation of gene promoters. According to our results, REST is involved in repression of NANOG expression and reduction of H3K9ac in promoter.
Furthermore, among the members of HDAC corepressor complexes, such as CoREST [23], MTA [24], and SIN3 [25], RCOR3 is a major upstream regulator of histone acetylation in NANOG promoter in chicken PGCs. Mouse ESCs showed varying numbers of recruited corepressors with Rest in target sites, indicating different interactions among their complexes in diverse regions [44]. In terms of early neuronal differentiation in mouse ESCs, the REST-CoREST complex modulates gene expression [45]. In this study, our results demonstrated that the REST/CoREST/HDAC complex modulates the H3K9ac level in upstream of NANOG to regulate its moderate expression in chicken PGCs.
In conclusion, we report that the CoREST/HDAC complex with REST, which binds to the NANOG promoter, functions as a transcriptional corepressor to regulate NANOG expression in chicken PGCs (Fig. 5). The CoREST/REST/HDAC repressor complex in PGCs is involved in maintaining moderate expression of NANOG, which is a critical transcription factor for germ cell characteristics. Therefore, regulation of NANOG expression by the REST/CoREST/HDAC complex could be involved in maintaining the PGC integrity.
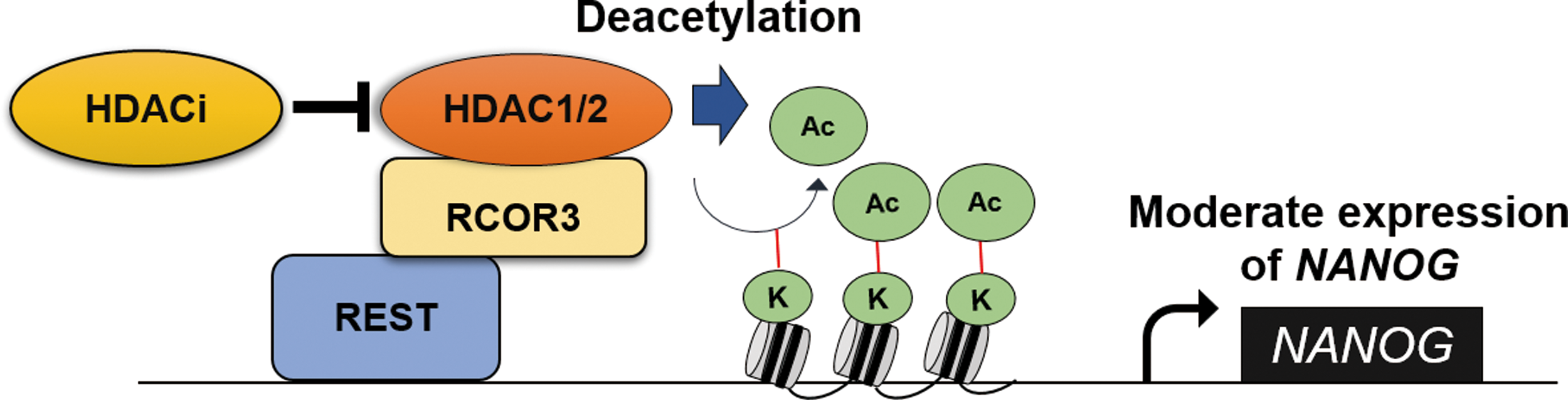
Epigenetic regulation of NANOG expression in chicken PGCs. Epigenetic regulation of HDAC complex controls NANOG expression in chicken PGCs. HDAC inhibitor represses deacetylation activity of the HDAC, resulting in upregulation of NANOG. The transcription factor REST binds to the upstream region of the NANOG promoter and represses NANOG expression with RCOR3 and HDAC. REST/CoREST/HDAC complex maintains moderate NANOG expression in chicken PGCs. HDAC, histone deacetylase; REST, RE1-silencing transcription factor; RCOR3, REST corepressor 3; PGCs, primordial germ cells. Color images available online at
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (no. NRF-2015R1A3A2033826). This research was also supported by the International Research & Development Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning of Korea (NRF-2016K1A3A1A21005676).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.