
Abstract
In mice, trophoblast stem (TS) cells are derived from the polar trophectoderm of blastocysts. TS cells cultured in the presence of fibroblast growth factor 4 (Fgf4) are in an undifferentiated state and express undifferentiated marker genes such as Cdx2. After removing Fgf4 from the culture medium, TS cells drastically reduce the expression of undifferentiated marker genes, stop cell proliferation, and differentiate into all trophoblast cell subtypes. To clarify the roles of the parental genomes in placentation, we previously established TS cells from androgenetic embryos (AGTS cells). AGTS cells are in the undifferentiated state when cultured with Fgf4 and express undifferentiated marker genes. After removing Fgf4, AGTS cells differentiate into trophoblast giant cells (TGCs), but not into spongiotrophoblast cells, and some of the AGTS cells continue to proliferate. In this study, we investigated the differentiation potency of AGTS cells by analyzing the expression of undifferentiated marker genes and all trophoblast cell subtype-specific genes. After removing Fgf4, some undifferentiated marker genes (Cdx2, Eomes and Elf5) continued to be expressed. Interestingly, TGCs differentiated from AGTS cells also expressed Cdx2, but not Prl3d1. Moreover, the expression of Gcm1 and Synb was induced after the differentiation, indicating that AGTS cells preferentially differentiated into labyrinth progenitor cells. Cdx2 knockdown resulted in increased Prl3d1 expression, suggesting that Fgf4-independent Cdx2 expression inhibited the functional TGCs. Moreover, Fgf4-independent Cdx2 expression was activated by Gab1, one of the paternally expressed imprinted genes via the mitogen-activated protein kinase kinase (MEK)-extracellular signal regulated protein kinase (ERK) pathway. These results suggested that the paternal genome activates the MEK-ERK pathway without the Fgf4 signal, accelerates the differentiation into labyrinth progenitor cells and controls the function of TGCs.
Introduction
In mammals, the blastocyst stage is the first step in the generation of differentiated cells. Blastocysts have two types of differentiated cells, namely, inner cell mass (ICM) and trophectoderm (TE) cells. There are two types of TE cells, mural and polar TE cells. After implantation, the mural TE cells, which are distant from the ICM, stop the proliferation and differentiate into the primary trophoblast giant cells (TGCs). In contrast, the cells of the polar TE, which is attached to the ICM, continue to proliferate and differentiate into placental tissues, including the labyrinth, spongiotrophoblast, and TGCs [1].
Trophoblast stem (TS) cells were established from the polar TE of E3.5 blastocysts in mice. TS cells continue to proliferate and remain in an undifferentiated state in the presence of fibroblast growth factor 4 (Fgf4) and primary mouse embryonic fibroblasts (MEFs) or MEF-conditioned medium. TS cells express undifferentiated marker genes such as Cdx2, Errb, and Eomes. After removing Fgf4 and MEFs or MEF-conditioned medium to induce the differentiation, TS cells stop the proliferation, repress the expression of undifferentiated marker genes and differentiate into labyrinth, spongiotrophoblast, and TGCs, and terminally into TGCs [2]. Therefore, TS cells are suitable models for analyzing the mechanism of placentation.
Mouse androgenetic (AG) embryos, which have two paternal genomes, are lethal until E9.5. Some AG embryos form a placental-like structure, but the AG placenta in vivo at E9.5 does not exhibit a functional structure due to the numerous TGCs and lack of spongiotrophoblast cells [3]. Therefore, both the parental genomes might be necessary for placentation.
To understand the roles of paternal and maternal genomes in the placentation, we established TS cells from androgenetic embryos (AGTS cells). These AGTS cells continue to proliferate and express undifferentiated TS marker genes (Cdx2 and Errb) in a manner similar to TS cells. After the differentiation, AGTS cells differentiate into TGCs [3]. Recently, we reported that some AGTS cells shift mitotic cell division into endoreduplication for TGCs formation, while others continue to proliferate after the differentiation [4]. These results suggest that AGTS cells have the ability to proliferate in an Fgf4-independent manner.
Fgf4, MEF, and MEF-conditioned medium are essential for the self-renewal of TS cells in vitro. For the self-renewal maintenance, TS cells express the transcription factor genes Eomes [5], Errb [6], Cdx2 [7], Ets2 [8], and Elf5 [9]. TS cells express these transcription factor genes in the presence of Fgf4 via the mitogen-activated protein kinase kinase (MEK)-extracellular signal regulated protein kinase (ERK) pathway [10]. After the differentiation, these transcription factor genes are repressed and the expression of placental-specific genes increases. On the contrary, the expression of these transcription factor genes and placenta-specific genes in AGTS cells have not been examined in detail. The aim of this study was to identify the expression patterns of these genes after the differentiation of AGTS cells to clarify the differentiation potency of AGTS cells and to calculate the roles of the paternal genome in placentation.
Materials and Methods
Cell culture
All mice were maintained and used in accordance with the Guidelines for the Care and Use of Laboratory Animals, as specified by the Japanese Association for Laboratory Animal Science and by the Tokyo University of Agriculture. TS cells and AGTS cells were cultured as previously described [3,4]. TS and AGTS cells were cultured in TS medium containing 25 ng/mL Fgf4 (PeproTech EC, London, United Kingdom) and 1 mg/mL heparin (Sigma-Aldrich, St. Louis, MO) with mitomycin-treated MEFs or with 70% MEF-conditioned medium (70condF4H). To induce the differentiation, TS and AGTS cells were cultured in TS medium without Fgf4, heparin, and MEF.
Immunostaining
Immunostaining of CDX2
TS and AGTS cells were fixed with 4% paraformaldehyde for 20 min at room temperature (RT) and then permeabilized in 0.25% Triton X-100 for 10 min at RT. Samples were blocked in phosphate-buffered saline (PBS) containing 1% bovine serum albumin (BSA; Sigma-Aldrich) for 1 h at RT and incubated with the antibody to anti-CDX2 (1:300; BioGenex, Fremont, CA) overnight at 4°C. After washing three times, the samples were incubated with Alexa 594 goat anti-mouse antibody (1:300; Invitrogen, Carlsbad, CA) for 1 h at RT. DNA staining and mounting on slides were conducted using VECTASHIELD WITH DAPI (Vector Laboratories, Burlingame, CA). Image acquisition was performed using confocal laser scanning microscopy (LSM710/780 ZEN; Carl Zeiss, Oberkochen, Germany).
Double-immunostaining of CDX2 and PRL3D1
After fixing with 4% paraformaldehyde for 20 min at RT, blocking solution (3% donkey serum and 1% BSA in PBS-T) was added to the samples and incubated for 1 h at RT without permeabilizing. After washing three times, the primary antibodies, goat anti-PRL3D1 (PL-1, 1:100; Santa Cruz, Dallas, TX), diluted with blocking solution were added and the samples were incubated overnight at 4°C. After washing three times, the secondary antibody Alexa Fluor 488 donkey anti-goat (1:1,000; Life Technologies, Carlsbad, CA) diluted with blocking solution was added and kept standing with protection from light for 1 h at RT. For CDX2 staining, the samples were permeabilized with 0.25% Triton X-100 for 10 min at RT. After washing three times, the primary antibody, mouse anti-CDX2 (1:300; BioGenex), diluted with blocking solution was added, and the samples were incubated overnight at 4°C. After washing three times, the secondary antibody, Alexa Fluor 594 goat anti-mouse (1:1,000; Life Technologies) diluted with blocking solution, was added and incubated for 1 h at RT. All steps of CDX2 staining were performed with protection from light. Each sample was then washed a final time with PBS(-), mounted and subjected to nuclear staining by adding a drop of VECTASHIELD WITH DAPI (Vector Laboratories). Image acquisition was performed using confocal laser scanning microscopy. The size of nuclear was measured using ImageJ software (WS Rasband, National Institute of Health, Bethesda, MD;
Western blotting
TS and AGTS cells were washed once with PBS and lysed on ice in 10 mL of buffer containing 62.5 mM Tris-HCl (pH 6.8), 2% SDS, 10% glycerol, 2.5% β-mercaptoethanol, and 1.25% bromophenol blue. The cells were then boiled at 95°C for 5 min. Electrophoresis was carried out on a 10% polyacrylamide gel, and separated proteins were transferred to a nitrocellulose membrane (Hybond ECL; GE Healthcare, Piscataway, NJ). The membrane was blocked in 5% skim milk in washing buffer (25 mM Tris-HCl, 150 mM NaCl and 0.05% Tween-20) for 1 h, and incubated with primary antibodies [anti-CDX2, 1:500 (BioGenex) and anti-human G3PDH (Trevigen, Gaithersburg, MD)] overnight at 4°C, followed by anti-rabbit or anti-mouse horseradish peroxidase-conjugated secondary antibodies (1:10,000; GE Healthcare). Signals were detected with an ECL Prime Western Blotting Detection Reagent (GE Healthcare) and visualized by a luminescent image analyzer (LAS-3000; Fujifilm, Tokyo, Japan). Samples consisting of 1 × 104 or 5 × 104 cells were used for the detection of proteins.
Gene expression analysis
Total RNAs were extracted by using ISOGEN (Nippon Gene) and were treated with DNase (Promega) to eliminate genomic DNA. DNase-treated total RNA was reverse-transcribed to cDNA by using Super Script III Reverse Transcriptase (Invitrogen). The quantitative gene expression analysis was performed by real-time polymerase chain reaction (PCR) using a QuantStudio 3 system (Applied Biosystems, Carlsbad, CA). PCR products were detected with SYBR Green using Power SYBR Green Master Mix (Applied Biosystems). The primers are listed in Table 1. The relative expression of target mRNAs was calculated from target CT values and Tbp CT values using the standard curve method. The expression levels of target mRNAs were normalized relative to the amount of Tbp.
Primer Sequences Used for Quantitative Gene Expression Analysis in This Study
siRNA transfection
siRNA was transfected with Lipofectamine 3000 Reagent (Thermo Fisher Science, Waltham, MA) according to the manufacturer's protocol. The siRNA sequences are listed in Table 2. siRNA-transfected AGTS cells were cultured for 48 h in TS medium without Fgf4. siRNAs were purchased from Funakoshi (Tokyo, Japan).
Short Hairpin Sequences Against Cdx2 and Gab1 Rnas Used In This Study
U0126 treatment
TS cells were cultured in 70condF4H containing 20 μM U0126 (a selective inhibitor of the MAP kinase kinases, MEK1 and MEK2; Merck Millipore, Billerica, MA). AGTS cells were cultured in TS medium containing 20 μM U0126 for 4 days.
Statistical analysis
Results are presented as the mean ± SE of three independent experiments unless otherwise stated. Statistical analysis was performed by Student's t-test. Values of P < 0.05 were considered to indicate statistical significance.
Results
Morphology of AGTS cells
TS and AGTS cell colonies retained a tight epithelial morphology cultured in TS medium containing Fgf4 and heparin with mitomycin-treated MEFs. After the differentiation, TS and AGTS cells formed TGCs [3]. At day 6 after the differentiation, almost TS cells differentiated into TGCs, however, some AGTS cells remained the epithelial morphology (Fig. 1).
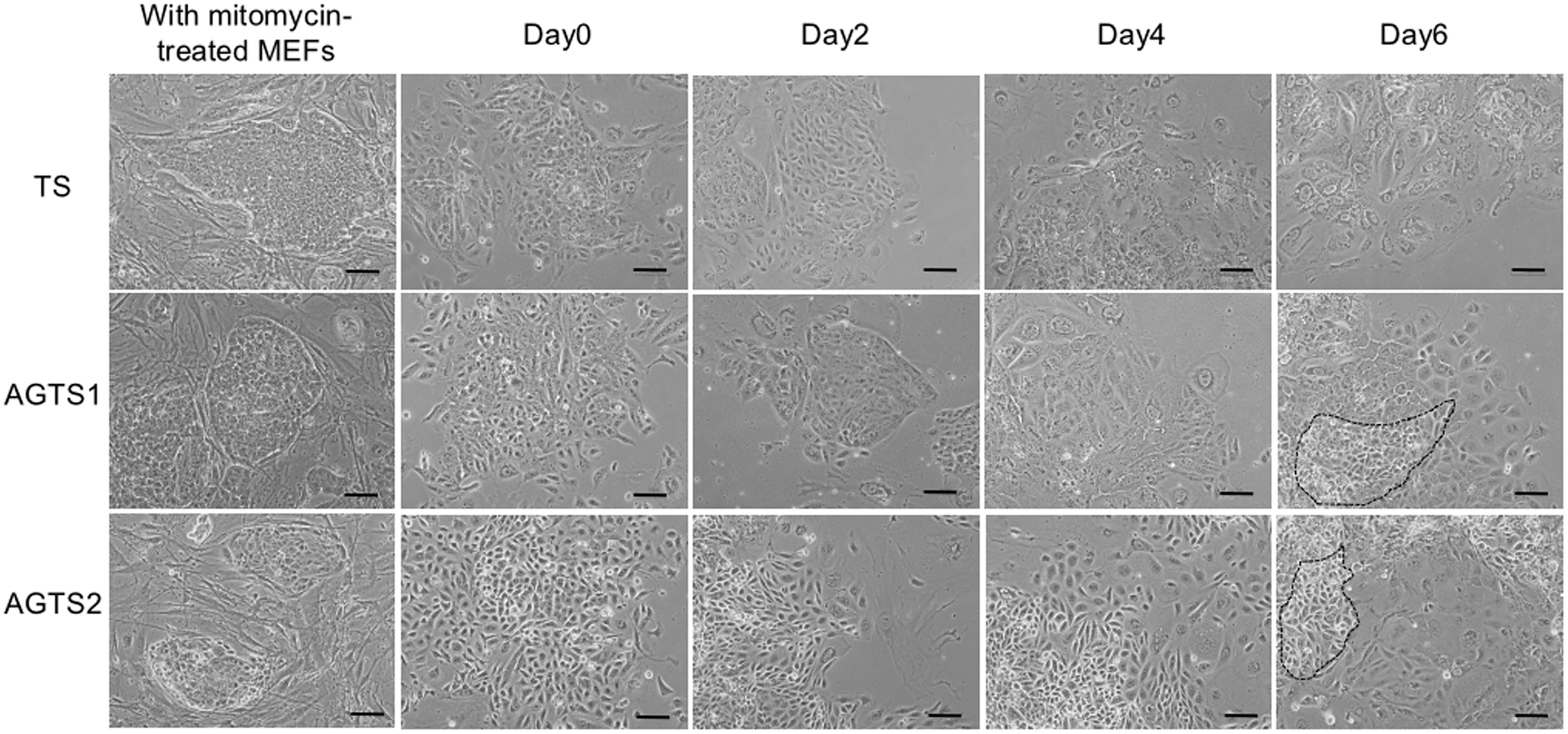
Photomicrographs of TS and AGTS cells cultured on mitomycin-treated MEFs and at days 0, 2, 4 and 6 after the differentiation. At day 6 after the differentiation, almost TS cells differentiated into TGCs, however, some AGTS cells remained the epithelial morphology (outlined by the black dotted line). Scale bars are 100 μm. AGTS, TS cells from androgenetic embryos; MEF, mouse embryonic fibroblast; TGC, trophoblast giant cell; TS, trophoblast stem.
TS marker gene expression in AGTS cells after differentiation
The expression of six TS marker genes (Cdx2, Eomes, Elf5, Sox2, Errb, and Tfap2c) in TS and AGTS cells after the differentiation is shown in Fig. 2. These six genes were expressed in undifferentiated TS cells, and all but Tfap2c were repressed after the differentiation. These genes were also expressed in undifferentiated AGTS cells. After the differentiation, Sox2 and Errb were repressed, but Cdx2, Eomes, and Elf5 mRNA were detected until day 6 of differentiation. Among these three genes, only Cdx2 was expressed stably in both two lines of AGTS cells.
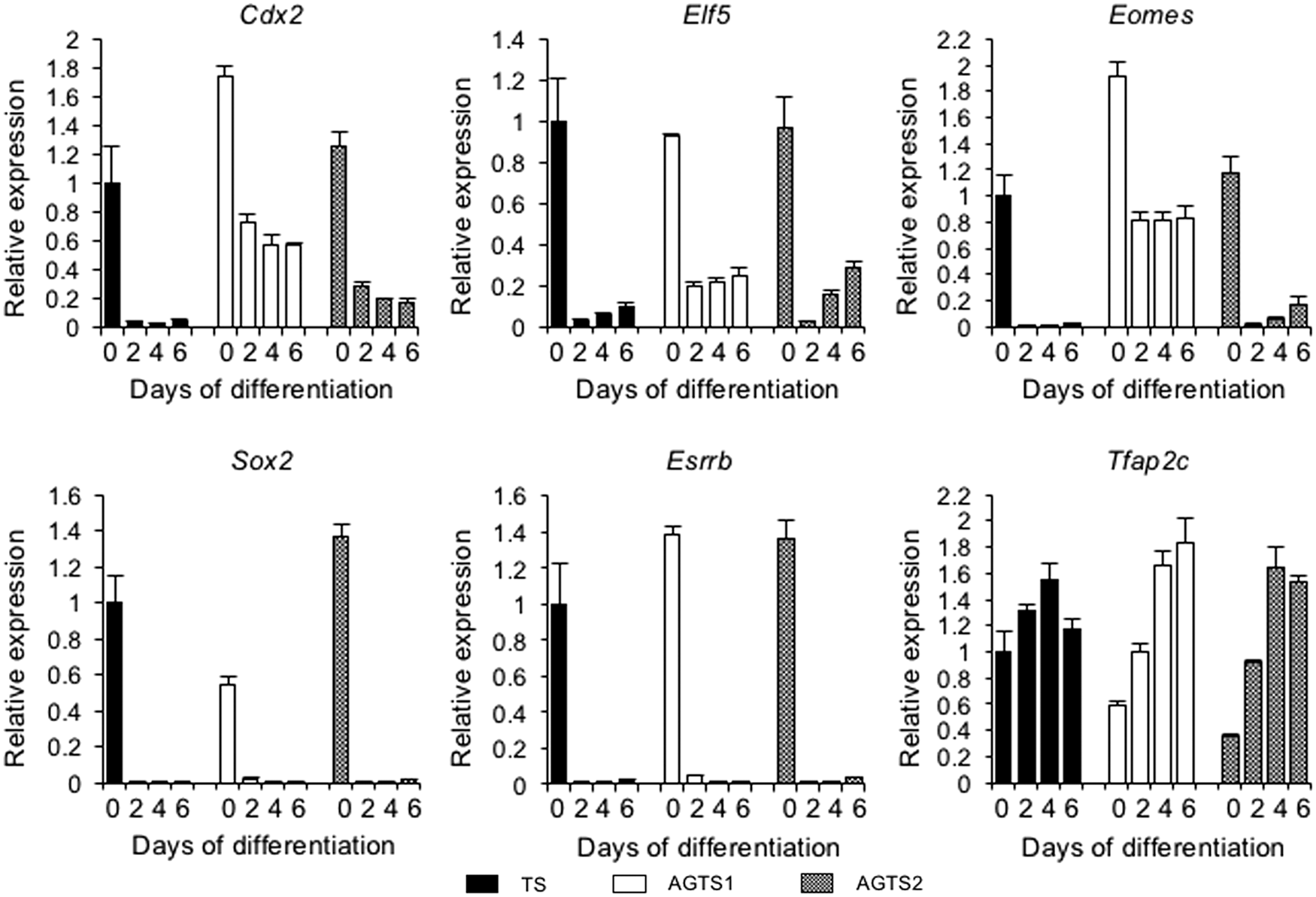
qRT-PCR analysis of the expression levels of six TS marker genes (Cdx2, Elf5, Eomes, Sox2, Esrrb, and Tfap2c) in TS and AGTS cells after the differentiation. Expression levels were all normalized with Tbp. The relative gene expression levels of each gene were calculated from TS cells at day 0 of differentiation. The data are represented as the mean ± SE. qRT-PCR, quantitative reverse transcription-polymerase chain reaction.
CDX2 protein expression in differentiated AGTS cells
The expression of CDX2 in TS and AGTS cells at days 0, 2, 4, and 6 after the differentiation is shown in Fig. 3A and B. In TS cells, CDX2 was repressed at days 2, 4, and 6 after the differentiation. On the contrary, AGTS cells expressed CDX2 until day 6 after the differentiation. These CDX2 protein expression patterns corresponded with Cdx2 mRNA expression. CDX2-positive cells were detected by immunostaining (Fig. 3C). Under the undifferentiated conditions, more than 80% of cells were CDX2-positive both in TS and AGTS cells. In TS cells, the percentages of CDX2-positive cells were about 20% and 5% at days 2 and 4 of differentiation, respectively. In contrast, in AGTS cells, CDX2 was also detected after the differentiation, and the percentage of CDX2-positive cells was about 80% at day 6 of differentiation (Fig. 3D).
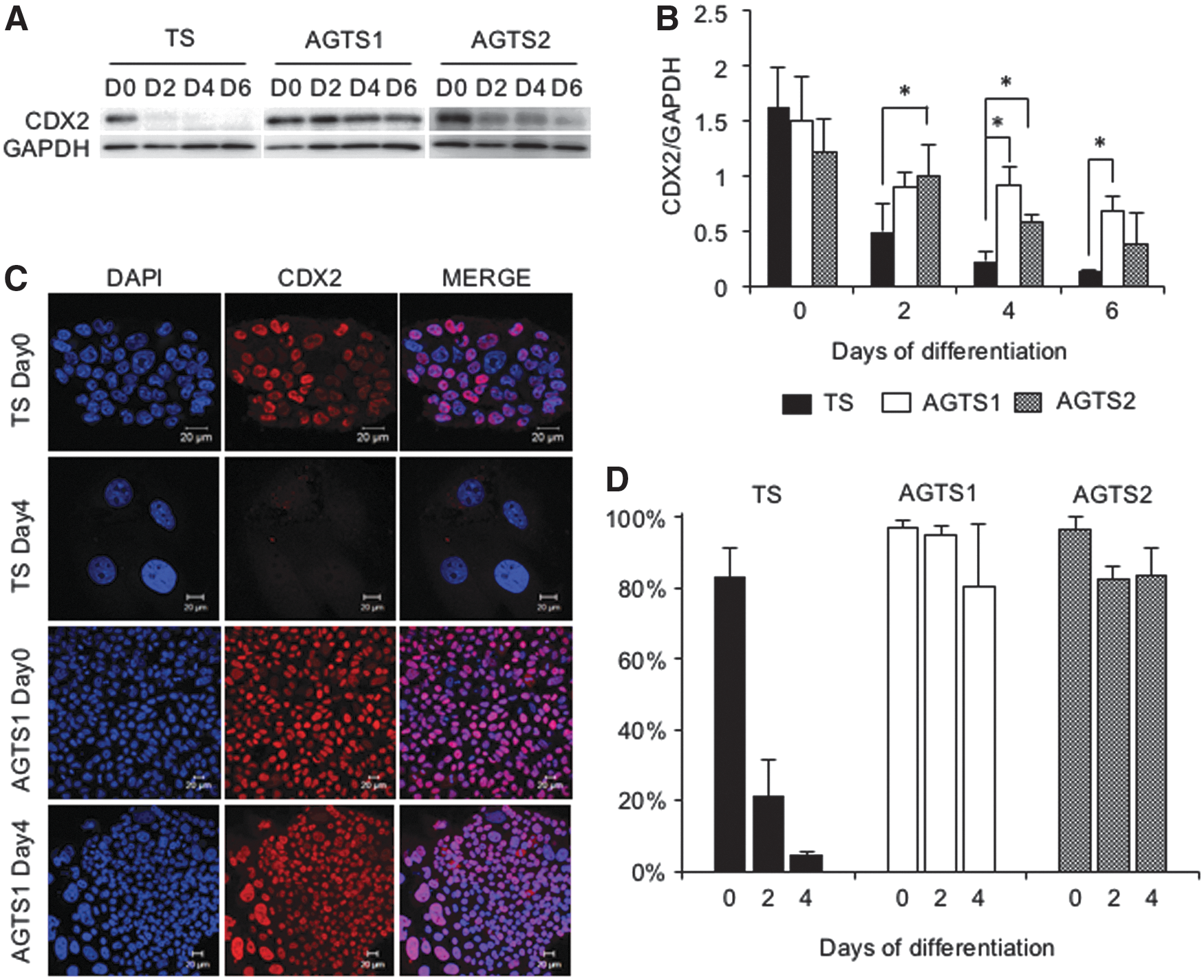
Expression of the CDX2 protein in TS and AGTS cells after the differentiation.
TGCs differentiated from AGTS cells expressed CDX2, but not PRL3D1
Both TS and AGTS cells formed TGCs after the differentiation [3]. The results of ploidy status by flow cytometric studies showed that at day 4 of differentiation, octaploid (8N) cells were appeared both in TS and AGTS cells, although the population of 8N cells was slightly lower in AGTS cells (data not shown). In TS cells, TGCs [with nuclear sizes of more than 20 μm (TS and AGTS1) and 15 μm (AGTS2) in diameter] never expressed CDX2. Although AGTS cells were also differentiated into TGCs, almost all these cells expressed CDX2 (Fig. 4A).
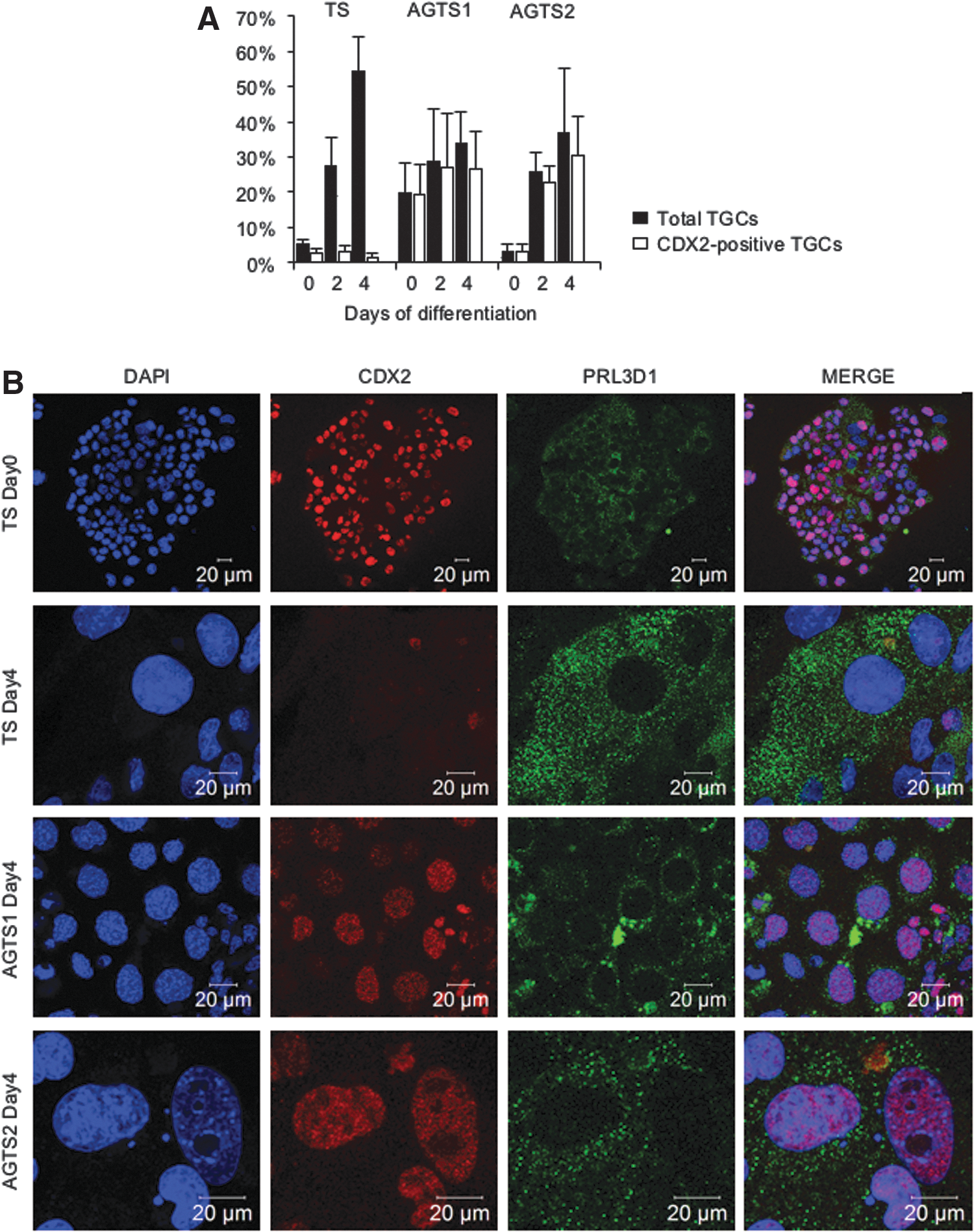
Expression of CDX2 protein in TGCs from TS and AGTS cells after the differentiation.
To clarify the features of CDX2-positive TGCs, double-immunostaining of CDX2 and PRL3D1 was carried out. TGCs from TS cells did not express CDX2 in nuclei and localized PRL3D1 in the cytoplasm. In contrast, TGCs from AGTS cells also expressed CDX2 in nuclei. Interestingly, these CDX2-positive TGCs did not express PRL3D1 in the cytoplasm (Fig. 4B).
AGTS cells preferentially differentiated into labyrinth progenitor cells
The expression of placenta-specific genes in TS and AGTS cells after the differentiation is shown in Fig. 5. The placenta-specific genes included spongiotrophoblast markers (Ascl2 and Tpbpa), labyrinth markers (Gcm1, Cebpa, Esx1, Syna, and Synb) (Fig. 5A), and TGC markers (Hand1, Prl3b1, Prl3d1, Prl2c2, and Prl4a1) (Fig. 5B). The spongiotrophoblast markers were not expressed in AGTS cells. These results supported the previous reports in vivo and in vitro [3,4]. Among labyrinth marker genes, Gcm1, Cebpa, Esx1, and Synb, but not Syna, were expressed in AGTS cells after the differentiation. The expression levels of Gcm1, Cebpa, and Synb were higher in AGTS2 than in AGTS1 cells. After the differentiation, none of the TGC markers except Hand1 were detected in AGTS cells. However, at day 6 after the differentiation, Prl3d1 and Prl4a1 were expressed in AGTS2 cells.

qRT-PCR analysis of placenta-specific genes in TS and AGTS cells after the differentiation.

Cdx2 knockdown induced Gcm1 and some of TGC markers expression in AGTS cells
To analyze the effect of Cdx2 expression after Fgf4 depletion on the differentiation potency in AGTS cells, Cdx2 was knocked down by siRNA. We used three types of siRNA (siCdx2-a, -b, and -c) and examined the expression levels of CDX2 protein by western blotting and immunostaining. The expression levels of CDX2 were markedly decreased by transfection of all three types of siRNA for Cdx2 (Fig. 6A, B). Therefore, siCdx2-a was used for gene expression analysis. Cdx2-knockdown AGTS cells showed a slight reduction of Elf5. Among labyrinth markers, Gcm1 was induced the expression, and some of TGC markers (Prl3d1 and Prl3b1) were induced the expression in Cdx2-knockdown AGTS cells (Fig. 6C). Spongiotrophoblast markers (Ascl2 and Tpbpa) were not detected in Cdx2-knockdown AGTS cells (data not shown).
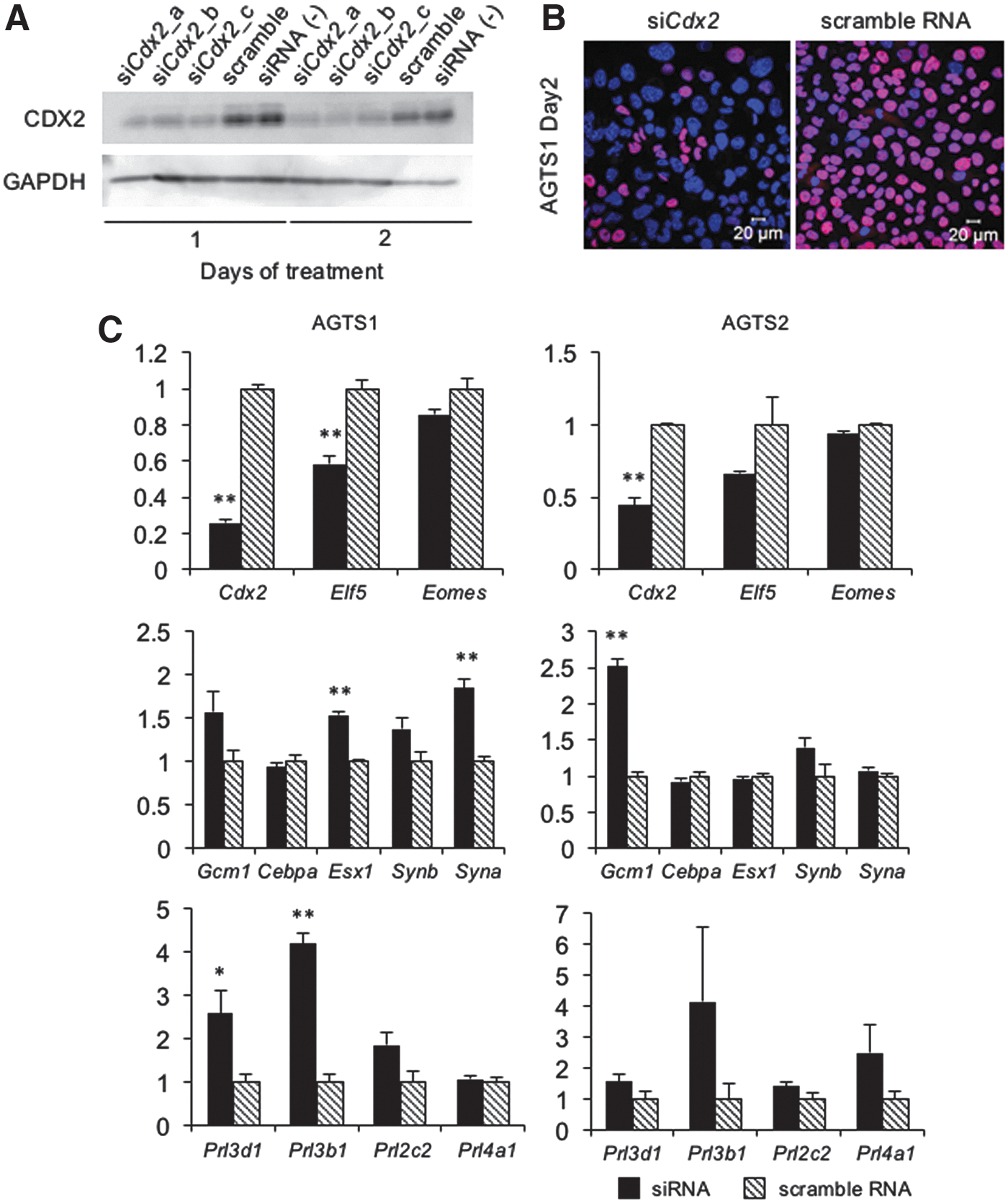
Knockdown of Cdx2 in AGTS cells by siRNA.
An MEK inhibitor induced Gcm1 and TGC markers expression in AGTS cells
TS cells express Cdx2 by an Fgf4-dependent MEK-ERK pathway. To clarify whether the expression of Cdx2 without Fgf4 was controlled by the MEK-ERK pathway in AGTS cells, we examined the effect of MEK-ERK inhibition by U0126 on Cdx2 expression without Fgf4. When TS cells were cultured in 70condF4H containing 20 μM U0126 for 4 days, the expression level of CDX2 decreased (Fig. 7A). This result indicates that 20 μM U0126 is sufficient to inhibit the MEK-ERK pathway to reduce Cxd2 expression in the presence of Fgf4. AGTS cells cultured with U0126 also repressed CDX2 (Fig. 7A). The expression of Cdx2 was also repressed in these cells, and Gcm1 and TGC markers (Prl3b1, Prl3d1, Prl2c2, and Prl4a1) were upregulated (Fig. 7B). Spongiotrophoblast markers (Ascl2 and Tpbpa) were not detected in U0126-treated AGTS cells (data not shown).
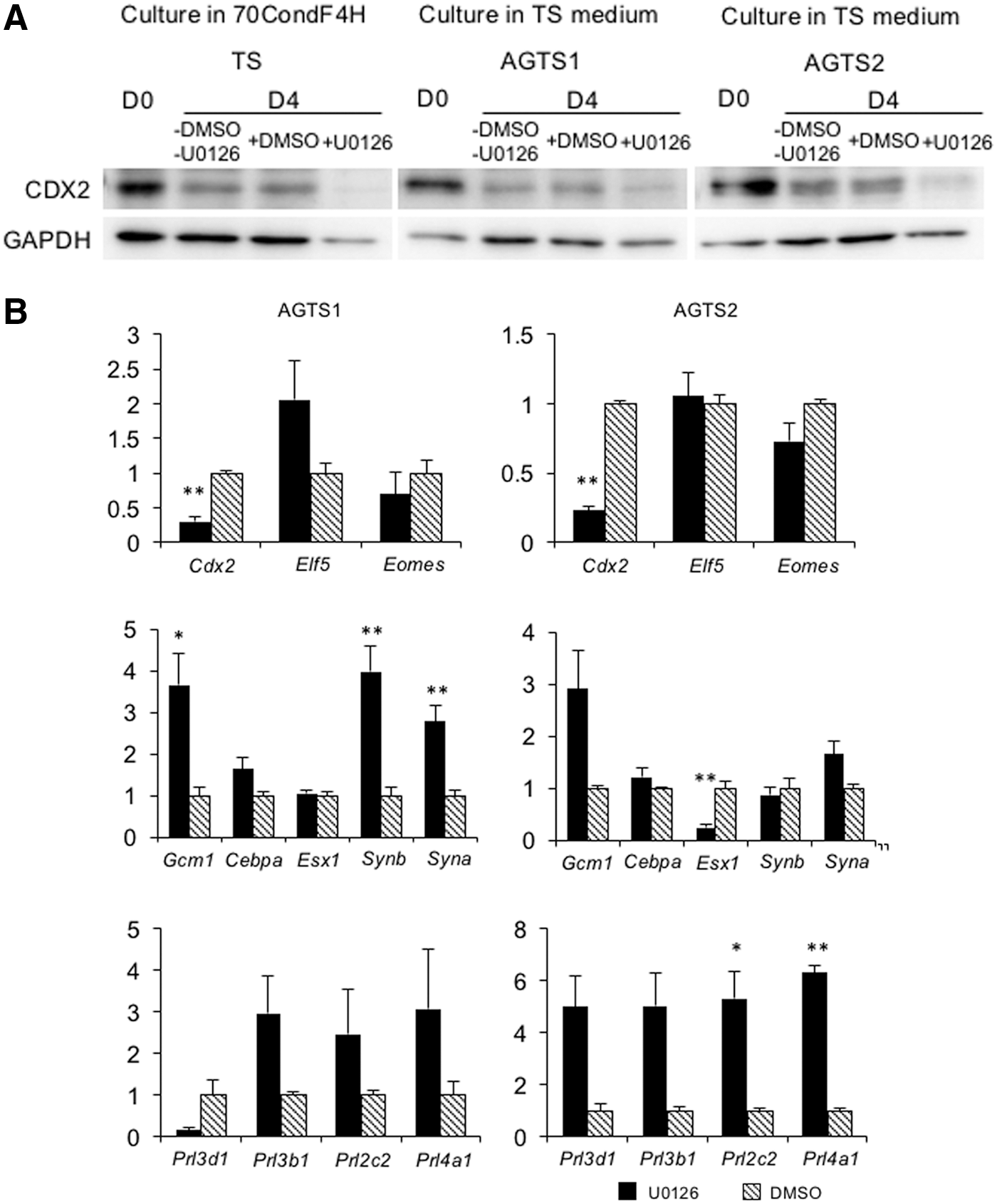
Gene expression analysis in U0126-treated AGTS cells.
Gab1 knockdown reduced Cdx2 expression in AGTS cells
Gab1, a key gene for extracellular signal regulated protein kinase (ERK)-mitogen-activated protein kinase (MAPK) signaling, is known to be important for placental development [11], and confirmed as the paternally expressed imprinted gene [12]. The expression of Gab1 in AGTS cells was significantly higher than that in TS cells under the undifferentiated state and at days 4 and 6 after the differentiation (Fig. 8A). To confirm whether the higher amount of Gab1 mRNA after the differentiation regulates the Cdx2 expression, AGTS cells were knocked down by Gab1 siRNA (3.75 and 7.5 nM) at day 2 after the differentiation for 48 h. Gab1-knockdown AGTS cells showed the reduction of Cdx2, however, the expression of TGC markers (Prl3b1 and Prl4a1) was not fluctuated (Fig. 8B)
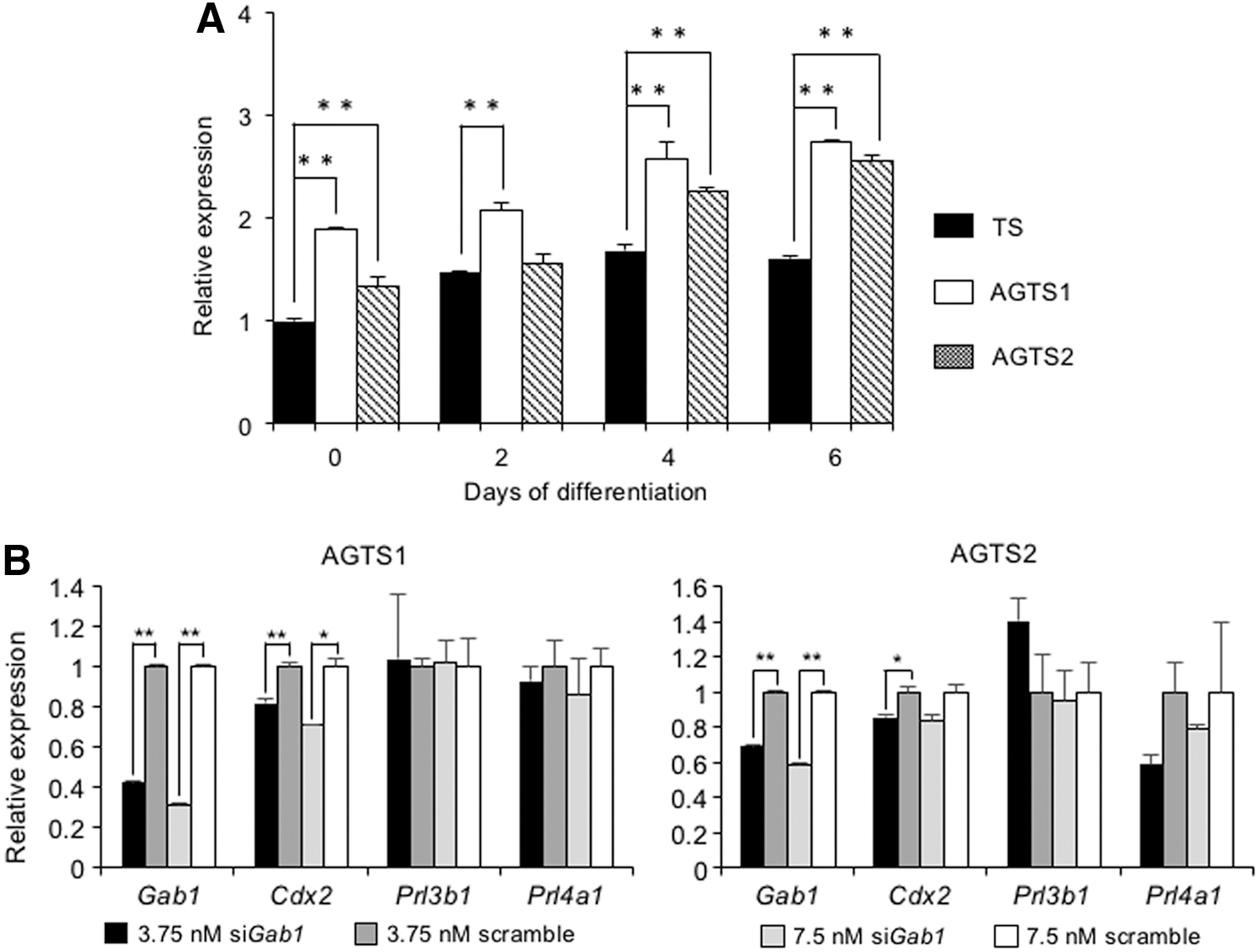
Effect of Gab1 on Cdx2 expression in AGTS cells.
Discussion
We previously established AGTS cells to elucidate the roles of the parental genomes during placentation. These AGTS cells expressed the paternally expressed imprinted genes (Peg1, Peg3, Peg10, Snrpn, U2af1-rs1, and Igf2), but did not express the maternally expressed imprinted genes (H19, Igf2, and Tssc3) [3]. Moreover, a genome-wide DNA methylation analysis in AGTS cells revealed three differentially methylated regions, which are paternally methylated regions, localized in the Gpr1-Zbdf2 region on chromosome 1, and identified a novel paternally expressed imprinted gene named Gpr1 [13]. These results suggest that AGTS cells have the paternal epigenetic modification. AGTS cells differentiated into TGCs, but not into spongiotrophoblast cells [3]. Their abilities to differentiation were thus similar to those of AG trophoblast cells in vivo [3]. Therefore, AGTS cells might be a useful tool to examine the roles of the parental genomes during placentation. The aim of this study was to obtain further insights into the differentiation potency of AGTS cells after the removal of Fgf4.
Many transcription factors, including Cdx2, Eomes, Elf5, Sox2, and Errb, are essential for TS self-renewal [14]. TS self-renewal is also dependent on Fgf4 signaling. After the removal of Fgf4, the TS cells repressed the transcription factors mentioned above, and stopped the proliferation. In contrast, AGTS cells continued to proliferate after the removal of Fgf4 [4]. Moreover, although the expression of Sox2 and Errb was completely repressed, the expression of Cdx2, Eomes, and Elf5 was sustained. These results indicate that some transcription factors (Cdx2, Elf5, and Eomes in this study) continued to be expressed without Fgf4 signaling in AGTS cells. Sox2 and Errb were the fastest and most profoundly downregulated genes [15], revealing that these two genes were primary targets and direct mediators of Fgf signaling in TS cells [16]. The combined forced expression of Sox2 and Errb supports Fgf-independent self-renewal of TS cells. Given that Sox2 and Errb can replace Fgf signaling and regulate crucial trophoblast genes, these factors would seem to be at the top of the transcription factor hierarchy that operates in TS cells [14]. However, the downregulation of Sox2 and Errb did not affect the expression of other TS transcription factors, such as Cdx2, Elf5, and Eomes, in AGTS cells. In TS cells, Eomes, Elf5, and Tfap2c are essential roles for cell fate determination. In the presence of high amounts of Eomes, the Eomes-Elf5-Tfap2c troika sustains the TS self-renewal [14]. When TS cells start to differentiate, Eomes is downregulated and the Elf5-Tfap2c complex binds to early differentiation-associated genes and accelerates their expression [14]. Therefore, the expression of Eomes, Elf5, and Tfap2c after Fgf4 removal might continue to proliferate in AGTS cells.
Cdx2 is a class I homeobox transcription factor that belongs to the caudal-related homeobox gene family [17]. Cdx2 has been reported to be expressed specifically in TE at the blastocyst stage, and its expression is maintained in the proliferating ExE [18]. Cdx2 mutant blastocysts fail to maintain trophoblast differentiation and to implant and are not established TS cells [19]. Moreover, embryonic stem (ES) cells can be induced to differentiate into the TE lineage by overexpression of Cdx2 [20], indicating that Cdx2 is a key gene for redirecting the ES cell fate toward the TE lineage. Taken together, these facts suggest that Cdx2 is essential for the establishment and maintenance of TS cells and the differentiation into TE lineage. After the differentiation, Cdx2 is repressed in TS cells, but sustained in AGTS cells. These results indicate that the paternal genome might regulate Cdx2 expression after the differentiation of TS cells.
Fgf4 and conditioned medium from MEF are essential for the self-renewal of TS cells in vitro [21]. In particular, Fgf4 acts as a stimulator of the signaling pathway that includes FRS2-Shp2-Ras-Raf-MEK-ERK [22] to preserve the expression of TS marker genes, including Cdx2. Although Fgf4 was added to the medium, U0126 repressed Cdx2 expression (Fig. 7A), confirming that Fgf4 regulates Cdx2 expression via the MEK-ERK pathway in TS cells. In contrast, the Cdx2 expression was repressed by U0126 in AGTS cells without Fgf4, indicating that Cdx2 is expressed in AGTS cells in an Fgf4-independent manner due to activation of the MEK-ERK pathway. Gab1 is a key gene for this pathway and is known to be important for placental development [11] and confirmed as the paternally expressed imprinted gene [12]. In this study, as the expression level of Gab1 is higher in AGTS cells than TS cells (Fig. 8A), we examined the effect of Gab1 on the Cdx2 expression in AGTS cells by knockdown. As expected, the knockdown of Gab1 downregulated the expression of Cdx2, indicating that Gab1 is a candidate gene for Fgf4-independent Cdx2 expression via the MEK-ERK pathway in AGTS cells.
After Fgf4 depletion, TS cells differentiate into TGCs via endoreduplication. Endoreduplication occurs when a cell undergoes multiple S-phases without entering mitosis and without undergoing cytokinesis [23], and it results in large polyploidy cells to form TGCs [24]. TS cells knockouted p57/kip2, the cyclin-dependent kinase-specific inhibitor, did not undergo endoreduplication, and formed multinuclear cells, suggesting that endoreduplication is triggered by p57/kip2 [23]. Although p57/kip2 is a maternally expressed imprinted gene [25], AGTS cells expressed p57/kip2 [4]. Therefore, AG trophoblast cells were able to differentiate into TGCs and not to form multinuclear cells in vivo and in vitro because of p57/kip2 expression [4]. TGCs produce a broad range of hormones that regulate several maternal adaptations to pregnancy [26]. In particular, the 23 prolactin/placental lactogen genes are spatially and temporally expressed in TGCs [27]. After Fgf4 depletion, although AGTS cells also differentiate into TGCs [4], the expression of Prl3b1 remains negligible. Surprisingly, TGCs from AGTS cells expressed CDX2 in the nuclei, but not PRL3B1 in the cytoplasm. Moreover, the expression of Prl3b1 was increased in Cdx2-reduced AGTS cells by siRNA or U0126, suggesting that Cdx2 expression did not disturb the formation of TGCs, but suppressed their function. Indeed, Cdx2 is known to bind widely across the TS cell genome at more than 10,000 loci. These loci contain Prl family genes such as Pr3b1 [15,28], suggesting that Cdx2 binds to Prl genes to repress their expression in TS cells.
The labyrinth of the rodent placenta contains villi that are the site of nutrient exchange between mother and fetus. The labyrinth is located in the innermost compartment of the placenta, and contains both chorion (trophoblast)-derived epithelial cells and allantois (extraembryonic mesoderm)-derived vascular cells. At E8.5, the chorion shows Syna, Gcm1, Synb, and Cebpa expression. Gcm1 has been shown to regulate Synb and Cebpa, but not Syna [29]. The cells expressing these genes differentiate into the syncytiotrophoblast (STB) layer II at E11.5. The STB layer II is one of the trilaminar trophoblast layers that separates the maternal and fetal blood spaces within the labyrinth [30]. In the present study, AGTS cells expressed Gcm1, Synb, and Cebpa, but not Syna after the differentiation. These results indicated that AGTS cells preferentially differentiated into the chorion and STB layer II.
Mouse AG placenta lacks spongiotrophoblast cells [3]. This morphology is similar to the placenta from the paternal duplication of distal chromosome7 or from the deletion of Dnmt3L [31 –33]. Furthermore, Mash2 (known as Ascl2) deficient embryos suppress the spongiotrophoblast differentiation [34]. AGTS cells could not acquire the expression of spongiotrophoblast markers (Ascl2 and Tpbpa) by Cdx2 knockdown or MEK-ERK inhibition because Ascl2 is a maternally expressed imprinted gene [35]. Taken together, the maternal genome is crucial for spongiotrophoblast formation in mouse placentation.
In summary, we have compared the differentiation potency of AGTS cells with that of TS cells. Our results showed that TS and AGTS cells expressed TS marker genes for self-renewal in an undifferentiated state. After the differentiation, although those marker genes were repressed in TS cells, the expression of some genes (Cdx2, Elf5, and Eomes) was sustained in AGTS cells. We also demonstrated that differentiated AGTS cells showed the giant cell morphology, but did not express TGC-specific genes. Moreover, after the differentiation, AGTS cells increased the expression of labyrinth-specific genes, indicating that AGTS cells preferentially differentiated into labyrinth cells. An overview of the differentiation and gene expression of TS and AGTS cells is given in Fig. 9A and B, respectively.

Differentiation potency of TS and AGTS cells.
Footnotes
Acknowledgment
This work was supported by a grant from the Ministry of Education, Culture, Sports, Science and Technology, Japan (no. 25450470).
Author Disclosure Statement
No competing financial interests exist.