
Abstract
Human embryonic stem cells (hESCs) have the potential to differentiate along the retinal lineage. We have efficiently differentiated human pluripotent stem cells into optic cup-like structures by using a novel retinal differentiation medium (RDM). The purpose of this study was to determine whether the retinal progenitor cells (RPCs) derived from hESCs can integrate into the host retina and differentiate into retinal ganglion cells (RGCs) in vivo. In this study, hESCs (H9-GFP) were induced to differentiate into optic cup-like structures by using our novel differentiation system. The RPCs extracted from the optic cup-like structures were transplanted into the vitreous cavity of N-methyl-
Introduction
Age-related macular degeneration (AMD), glaucoma, diabetic retinopathy, and retinitis pigmentosa (RP) are the main optic nerve and retinal diseases. These diseases are significant public health concerns, as they are becoming increasingly common in the elderly [1 –3]. The pathological basis of these irreversible retinal diseases is the degeneration or damage of some specific retinal neurons [3,4]. For example, AMD is associated with damage to photoreceptors and retinal pigment epithelial cells, and RP is a group of inherited disorders associated with progressive photoreceptor degeneration and blindness [5].
In addition, the progressive apoptosis of retinal ganglion cells (RGCs) characterizes the pathological basis for glaucoma. Unfortunately, as part of the postmitotic central nervous system, retinal neurons are typically unable to regenerate themselves. At present, the ideal therapy method for these degenerative retinal diseases such as glaucoma is still unknown. Human embryonic stem cells (hESCs) have the potential to differentiate along the retinal lineage in vivo and in vitro [6,7]. Therefore, previous studies have shown that different types of retinal cells could be differentiated from hESCs and human-induced pluripotent stem cells (hiPSCs), including retinal pigment epithelium (RPE), photoreceptors, and RGCs. Furthermore, the complicated optic-cup organ was shown to self-organize within a three-dimensional (3D) culture system [8 –12]. In particular, stem cell-derived RPE and photoreceptors have restored vision in preclinical models of human retinal degenerative disease [5,13 –15], which suggests that embryonic stem cell (ESC)-based therapy is a promising treatment option to restore vision in patients with degenerative retinal diseases.
Recently, a significant breakthrough in the use of stem cells in regenerative medicine was made by the Lamba group. Their study indicated that transplantation of hESC-derived photoreceptors can restore some visual function in Crx-deficient mice [16]. Furthermore, RPE cells derived from hESCs and hiPSCs have been used in clinical patients, and great progress has been made [15,17 –20]. A recent study reported the successful use of induced pluripotent stem cells (iPSCs) to form 3D retinal cups containing all the major retinal cell types [21], opening the path to possible new approaches.
Our group has efficiently differentiated the hESC lines (H1, H9) and the human iPSC line (IMR90-4) into optic cup-like structures using a retinal differentiation medium (RDM) without small molecular compounds [22]. A number of retinal progenitor markers were detected in these optic cup-like structures, including Chx10, Mash1, Otx2, Sox2, NeuroD, and Pax6, and the expression profiles of these transcription factors mirrored their counterparts during human embryogenesis [23,24]. Whether retinal progenitor cells (RPCs) can survive and differentiate into specific retinal neurons when transplanted into experimental eyes with glaucoma is still unknown.
To determine whether RPCs from the hESC-derived optic cup-like structures could integrate into the host retina and differentiate into RGCs when transplanted into the vitreous cavity of N-methyl-
Materials and Methods
Animals
Male C57BL/6J mice (8–10 weeks old) were obtained from Shanghai SLAC Laboratory Animal Co., Ltd. The animals were maintained in standard animal rooms with food and water ad libitum. All animal experiments were conducted in accordance with the Association for Research in Vision and Ophthalmology statement for use of animals in ophthalmic and vision research and were approved by the animal research committee at the Shanghai Medical College of Fudan University.
Maintenance and neural differentiation of hESCs
An enhanced green fluorescent protein-expressing human ES (H9 inGFPhES) cell line was gifted from the laboratory of Su-Chun Zhang (Waisman Center and Wicell Research Institute). Cells were maintained in ESC medium (see Supplementary Table S1 for media) for several passages for acclimation to culture conditions, and then were differentiated according to previously established methods (Supplementary Fig. S1). Partially differentiated colonies were manually removed before differentiation analysis. Undifferentiated hESC colonies were separated from the feeder cells and cultured in ESC medium for 4 days. On day 4, differentiation was initiated using neural differentiation medium (Supplementary Table S1). By day 7, aggregates of hESCs had differentiated into primitive neural epithelium and were transferred to adherent culture.
Differentiation toward a retinal fate was accomplished using our previously published protocols [22]. In brief, on day 14, hESC-derived neural tube-like rosettes were cultured in RDM (Supplementary Table S1). Two days later, the rosettes were mechanically detached from the dish and were cultured in suspension as cell spheres in the same medium. On days 18–22, most of the cell spheres adopted a vesicular appearance. To differentiate the cells into neural retina, bFGF (20 ng/mL; R&D Systems, Inc., Minneapolis, MN) was added to the culture medium. And the medium was changed every 3 days. On days 42–56, the optic cup-like structures were collected for further analysis. In addition, round cover slips were placed in six well plates where the transplanting cells (∼4 × 105 cells/well) were cultured, and the cell climbing pieces were collected on the third day.
Cell transplantation
At 6–8 weeks, hESC-derived optic cup-like structures were dissociated into single cell dilutions with 0.25% trypsin LE and dispase (Invitrogen). The isolated cells were washed three times with RDM and centrifuged to a concentration of 6.7 × 103 cells/μL in RDM. Approximately 2 × 104 isolated cells were injected into the vitreous cavity of mice that had been injected with NMDA 21 days before. The same volume of sterile RDM was injected into the contralateral eye for use as a control.
Immunofluorescence
The optic cup-like structures and fetal eye tissues (collected from human and mouse fetal tissues) were fixed in 4% paraformaldehyde (PFA; Sigma, St Louis, MO) in 0.1 M phosphate buffer, cryoprotected by immersion in 30% sucrose, embedded in OCT (Tissue-Tek, Sakura Finetak USA, Inc, CA), and frozen at −80°C until they were sectioned on a cryostat into 10 μm-thick slices and mounted onto slides. Sections were washed with 0.01 M phosphate-buffered saline (PBS) for 15 min at room temperature and incubated in blocking solution (10% donkey serum and 0.3% Triton X-100 in 0.01 M PBS) for 1 h at room temperature. Primary antibodies were incubated overnight at 4°C. The following antibodies were used: Math5 (1:2,000, ab78046; Abcam), Brn3b (1:1,000, sc31987; Santa Cruz), Brn3 (1:200, sc6026; Santa Cruz), Pax6 (1:1,000; DSHB), Nanog (1:1,000, ab21624), Oct 3/4 (1:1,000, ABP52004; Abbkine), and Tuj1 (1:1,000, Ab18207; Abcam). The corresponding secondary antibodies (Alexa Fluor 488, A11055; Alexa Fluor 488, A21202; Alexa Fluor 488, A21206; Alexa Fluor 594, A11058; Alexa Fluor 594, A21203; Alexa Fluor Cy5, A21447) were incubated for 1 h at room temperature. The nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI; Invitrogen). The staining specificity for each antigen was tested by omitting the primary antibody. Image analysis and photographic documentation were accomplished with a fluorescence microscope and a confocal laser-scanning microscope.
For whole mount retina, 4 and 5 weeks after cell transplantation, the eyes were enucleated and immersion fixed in 4% PFA for 10 min. The retinas were isolated and four radial cuts were done. Subsequently, the retinas were pasted on the nitrocellulose membrane with the GCL upturned, washed with 0.1 M PBS for 30 min to eliminate the PFA, and incubated in blocking solution (5% donkey normal serum, 1% Triton X-100 in 0.1 M PBS) for 1 h at room temperature. Primary antibodies were incubated for 3 days at 4°C. The following antibodies were used: Brn3a (1:200, sc-31984; Santa Cruz), hNu (1:1,000, MAB1281; Millipore), Tuj1 (1:1,000, Ab18207; Abcam), and GFP (1:1,000, ab6556; Abcam). The corresponding secondary antibodies (Alexa Fluor 488, A21202; Alexa Fluor 488, A21206; Alexa Fluor 594, A11058; Alexa Fluor 594, A21203; Alexa Fluor 594, A21207; Alexa Fluor Cy5, A21447; Invitrogen) were incubated for 2 h at room temperature, and the nuclei were counterstained with DAPI. Finally, the retinas were thoroughly washed with 0.1 M PBS, mounted vitreal side up on slides and covered with antifading solution (Southern Biotech). Photographic documentations were accomplished with a confocal laser-scanning microscope (TCS SP8; Leica Microsystems, Wetzlar, Germany).
Quantification
Image processing was performed with Adobe Photoshop CS4 software (Adobe Systems, San Jose, CA) and was limited to brightness/contrast adjustments. The figures were constructed with GraphPad Prism 5.0 (Adobe Systems, Inc.).
Results
The optic cup-like structures were self-organized in vitro by using our novel differentiation system
The hESCs were maintained in the undifferentiated state using mouse fibroblasts, and the morphology of stem cell clones were well defined (Fig. 1A, B). In particular, differentiated colonies were manually removed before subculture and differentiation analysis. The hESCs were identified by expression of the stem cell markers Nanog and Oct 3/4 (Supplementary Fig. S2). Following our published efficient differentiation protocol for producing optic cup-like structures from hESCs, the embryoid bodies were formed on days 3–4 (Fig. 1C, D) after detaching the hESC clones from the feeder. On days 14–16, hESC-derived neural tube-like rosettes were observed (Fig. 1E, F). On days 28–35, the optic cup-like structures were formed spontaneously and self-organized (Fig. 1G, H), and the transparent neuroepithelia structures were clearly observed within them.
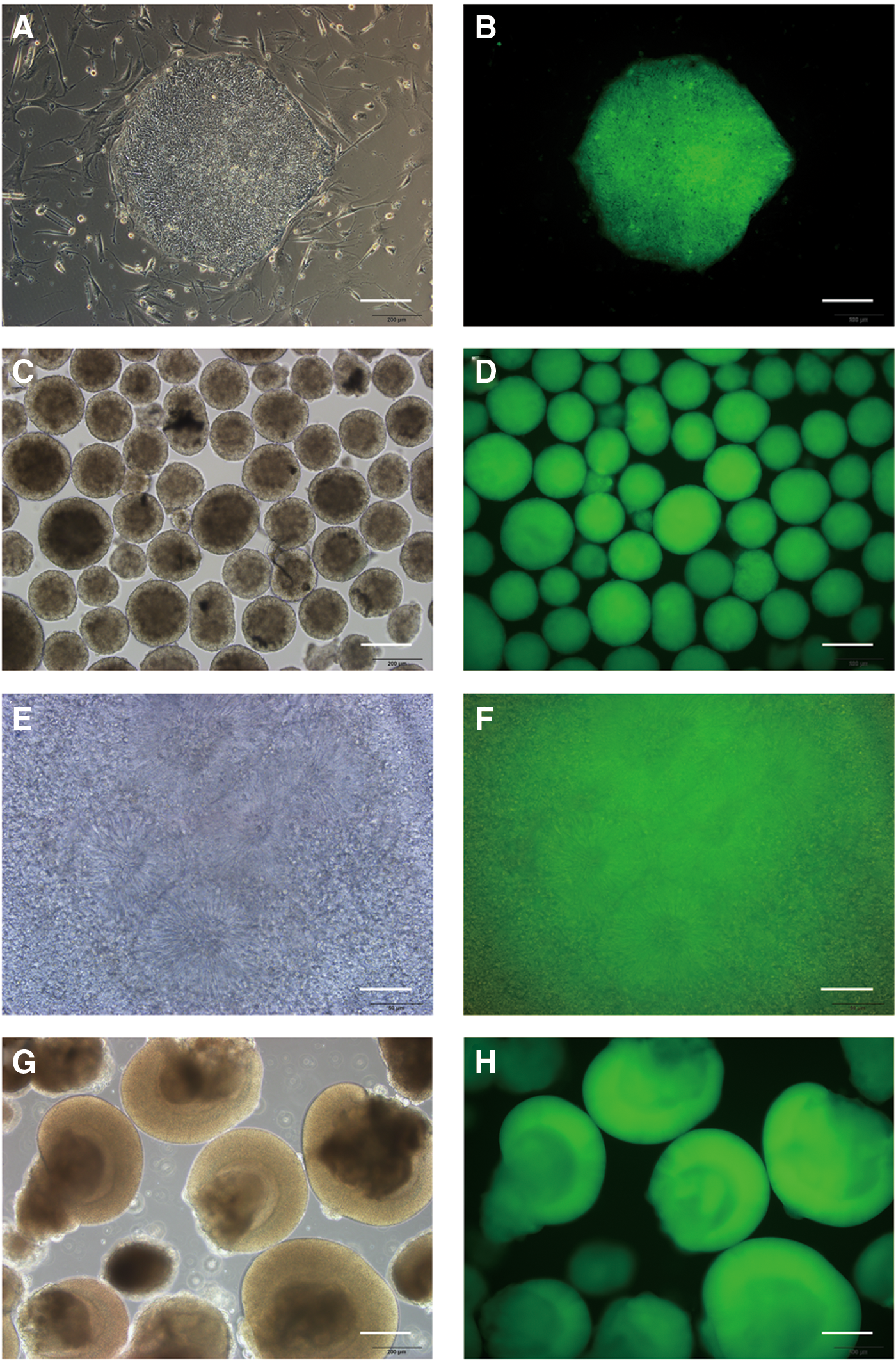
Optic cup-like structures were differentiated from hESCs in vitro. The brightfield
To test the characteristics of the cells in the optic cup-like structures, the optic cup-like structures were collected and investigated by using immunofluorescence. As expected, the eye field-specific transcription factors Math5 and Brn3b were expressed in the hESC-derived optic cup-like structures (Fig. 2A3). Previous studies have shown that Math5 (Atoh7) encodes a basic helix-loop-helix transcription factor that is specifically expressed by RPCs. This is required not only for cell cycle progression but also for early multiple retinal neuronal differentiation, including RGCs, photoreceptor, horizontal, and amacrine cells [25 –27]. The vast majority of cells in optic cup-like structures were Math5 positive (Fig. 2A1), suggesting that these cells were RPCs.
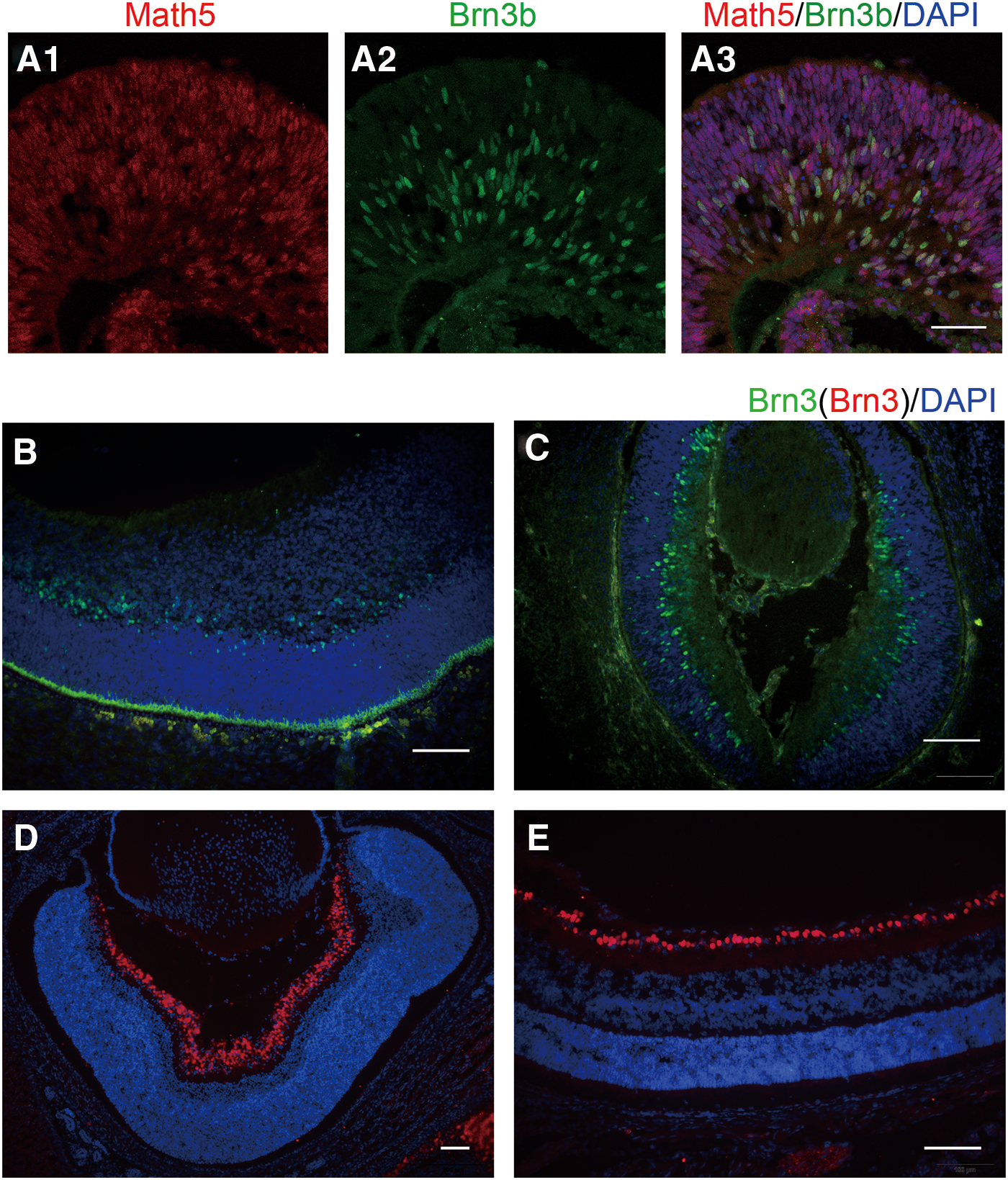
Comparison of optic cup-shaped cells from hESC-derived retinal spheres with fetal human and mouse retina tissues. Double immunofluorescence of Math5 (red), Brn3b (green), and merge (orange) in hESC-derived neural retina spheres at 6 w
Brn3b, a POU domain transcription factor, is an important regulatory gene in stem cells differentiated into RGC [28]. Brn3b specifies the RGC fate from multipotent precursors not only by promoting RGC differentiation but also by suppressing non-RGC differentiation programs as a safeguard mechanism [29]. Brn3b is the first Brn3 transcription factor to appear during RGC development. The radial expression pattern of Brn3b-positive cells (Fig. 2A2) was similar to the distribution in early fetal human retinal tissues (9 gw) and mouse retinal tissues (E12) (Fig. 2B, C). All Brn3b-positive cells were colabeled with Math5. In other words, there were many RGC progenitors in the optic cup-like structures. These data indicated that nearly all cells in the optic cup-like structures were RPCs, containing ganglion cell progenitors that could be differentiated into RGCs in the future.
The identification of RPCs
Cell transplantation might offer a convenient way to replace the lost ganglion cells in degenerative diseases such as glaucoma. The selection of an ideal cell type for this purpose needs careful consideration. To examine the aim of this study, it is, therefore, important to test the inherent properties of transplanting cells.
The climbing pieces of the cells extracted from the optic cup-like structures were collected and detected by using double immunofluorescence. As expected, we found abundant expression of a number of transcription factors, including Math5 (Fig. 3A1) and Pax6 (Fig. 3B1). Previous studies have shown that these markers were specially expressed by RPCs. Meanwhile, all nuclei of the cells derived from hESCs were efficiently labeled with human nuclei antigen (hNu) (Fig. 3A2, B2). To characterize hNu, it is more convenient to identify and quantify the transplanting cells than the GFP cells, although the expression of GFP was maintained after transplantation. In addition, <10% of the Brn3b-positive cells were observed (Supplementary Fig. S3B). These results indicated that most of the transplanting cells were in an undifferentiated stage, including ganglion cell progenitors.
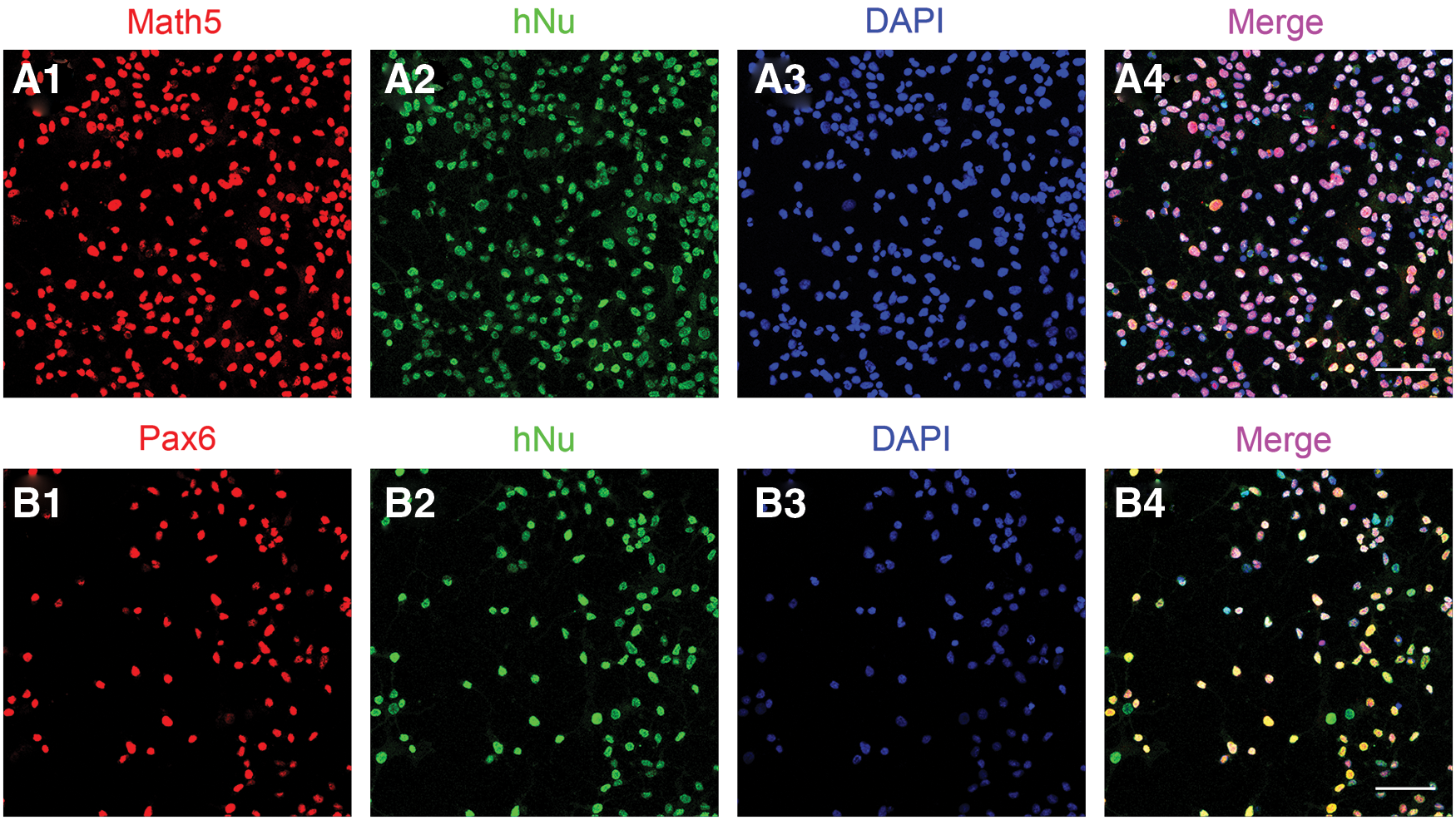
Cells of optic cup-like structures express retinal progenitor markers. Most of the transplanted cells from optic cup-like structures derived from hESCs expressed the retinal progenitors specific markers Pax6 and Math5
Surprisingly, with the extended incubation time, the cells extended numerous long processes that could be viewed under a bright light microscope. We found that the majority of cells were immunopositive for Map2. Map2 is a neuron-specific marker. These data demonstrated that the RPCs will differentiate into neurons.
The migration and integration of the transplanting cells
Retinal cells are generated by common multipotent RPCs in an evolutionarily conserved temporal sequence [30]. Recently, it was found that photoreceptor-like progenitors, derived from hESCs and iPSCs, could differentiate into photoreceptors and form a cell layer connected to host retinal neurons after transplantation into mice with end-stage retinal degeneration. Furthermore, visual function was partially restored in these animals [10]. These data validated that transplanting RPCs might be a better choice for retinal degenerative diseases.
To determine whether the RPCs could migrate and integrate into the host retina, the RPCs were extracted from the optic cup-like structures derived from hESCs and injected into the vitreous cavity of NMDA-treated mice. The host retinas were analyzed by immunohistochemistry on the fourth and fifth weeks after transplantation. Our previous work demonstrated that excitotoxicity induced by NMDA could cause acute and severe damage to conventional RGCs [31]. Therefore, on the fourth week after transplantation, a significant reduction in Brn3a-positive cells (red) were observed in the NMDA-treated retina (Fig. 4A1, B1) compared with the control group. This demonstrated that the experimental glaucoma models were made successfully. Meanwhile, many hNu-positive cells were also clearly observed in the host retina (Fig. 4A2, B2). These data showed that the transplanting cells migrated from the vitreous cavity to the retina and survived. Surprisingly, it seemed like these cells had a complementary distribution (Fig. 4A3, B3) with the host residual conventional RGCs. To determine whether they were localized in the same layer of the retina, a 3D image was restructured using confocal software. The complementary distribution of transplanting cells and the host surviving ganglion cells were clearly observed (Fig. 4C), which colocalized in the GCL of the host retina. The same results were further certified from the level of retinal sections (Supplementary Fig. S4). These data definitely indicated that the RPCs could migrate and integrate into the GCL of the host retina.
The transplanted cells migrated and integrated into the host retina. On the fourth week after transplantation, the surviving transplanted cells (hNu+) (green)
The generation of the RGCs in vivo
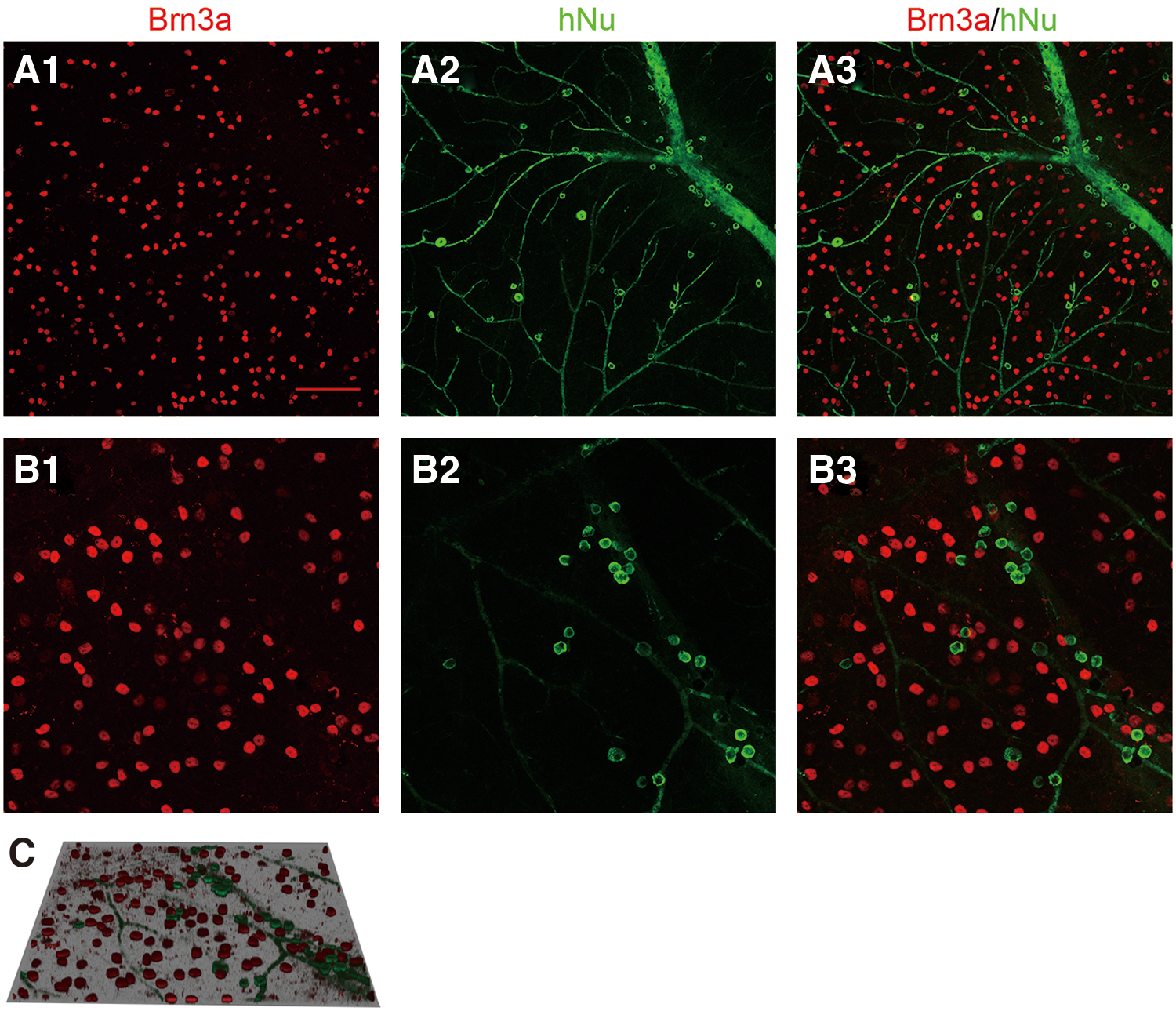
RPCs derived from hESCs could provide a limitless source of new cells for cell replacement strategies to substitute lost cells, and our aforementioned data have shown that the transplanting RPCs could migrate and integrate into the GCL of the host retina. However, it is unknown whether these cells could differentiate into ganglion cells and restore the visual function of the host. To determine whether the RGCs could be generated by the transplanting RPCs derived from hESCs, the host retinas were analyzed by using triple immunofluorescence staining on the fifth week after transplantation (Fig. 5). Brn3a, a mature ganglion cell marker, was used to label the host residual RGCs in the mouse retina (Fig. 5A1, B1). As expected, the complementary distribution of hNu-positive cells and Brn3a-positive cells was also observed in the GCL of mouse retinas (Fig. 5A4). Surprisingly, a small number of hNu-positive cells were colabeled with Brn3a (Fig. 6B1, C1, D1, E1), which indicated that the mature RGCs have been generated even although the expression level of Brn3a was lower than in the host conventional RGCs. The aforementioned data showed that the transplanting RPCs contained the ganglion cell progenitors, but the percentage is lower; some new RGCs may be differentiated from RPCs under the influence of the host retina microenvironment.
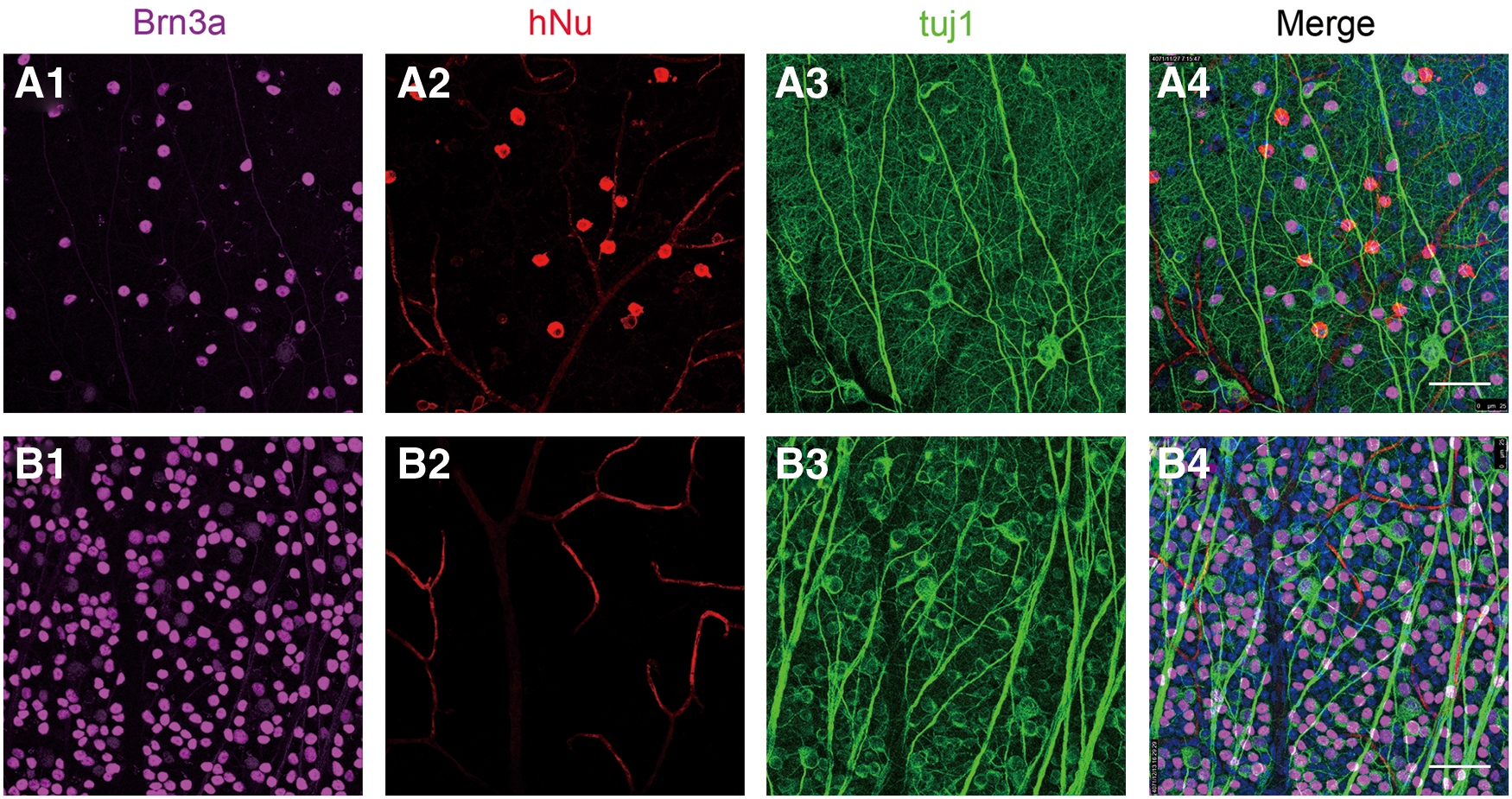
The surviving transplanted cells were well integrated in the host GCL. Compared with control group

RGC-like cells were differentiated from retinal progenitors in vivo. On the fifth week after transplantation, all of the surviving transplanted cells (hNu+) (red) were well integrated in the host GCL
Furthermore, the same results were further certified from the level of retinal sections (Supplementary Fig. S5); the surviving transplanted cells (hNu+) (red) were clearly observed in the host retina GCL (Supplementary Fig. S5A1, A2), and they also expressed Brn3a (pink) (Supplementary Fig. S5B1, B2), which is a classic marker of mature RGCs, confirming their potential to differentiate into RGCs. Meanwhile, other surviving transplanting cells localized in the GCL were Brn3a negative (Fig. 6F1). These cells may be amacrine cells or another type of ganglion cell. Either way, the complementary distribution of these cells with remaining host RGCs might be helpful for the retina. On the whole, the overall efficiency integration was poor (∼5%) on the fifth week after transplantation. Excitingly, there was no teratoma appeared in our experiment. These data represent morphological evidence that differentiated ganglion cells could be generated by the RPCs derived from hESCs in vivo.
Discussion
Glaucoma is one of the leading causes of irreversible blindness in the elderly, and degeneration of RGCs in the neural retina is a common cause of blindness. The conventional strategies for glaucoma are based on surgical or medical interventions to reduce intraocular pressure and limit visual loss. However, the number of RGCs still decreases progressively, irrespective of treatment in many patients. On account of the fact that RGCs cannot regenerate themselves, and there are no efficient treatments available to prevent the loss of RGCs, new, safe, and effective therapies should be developed for this sight-threatening disease.
Recently, tremendous progress has been made in generating RGC-like cells from stem cell populations, and stem cell research has shown promising applications of stem cells in therapeutic treatment [9]. For example, ESCs can generate RGC-like cells by the addition of FGF2 and Shh to the differentiating cells. However, <2% of these cells express RGC-specific markers such as ATH5, BRN3B, RPF-1, Thy1, and ISLET1, and intravitreal transplantation of these cells into mouse retina was not promising [32]. Ganglion-like cells derived from mouse iPS cells have also been induced by the addition of Noggin, DKK1, and DAPT as well as overexpression of Math5. These cells had elongated neurites and expressed RGC-specific markers such as Brn3b, Islet-1, and Thy1.2 [33]. Unfortunately, these cells survived but failed to integrate into the host retina after transplantation. Recently, functional human RGCs with target specificity were generated from pluripotent stem cells by chemically defined recapitulation of developmental mechanisms [34]. However, it remains unclear whether these cells could be used to integrate into the host retina.
As reported, the GCL consists of approximately equal numbers of RGCs and displaced amacrine cells in murines [35 –37]. Meanwhile, previous studies have shown that displaced amacrine cells are also damaged in several models of experimental glaucoma [38,39]. Hence, the transplantation of ganglion cells only may be inefficient. In retinal development, six retinal cell types are generated from a common pool of multipotent progenitors in vivo, and each progenitor can produce clones with one to six cell types [24,40,41]. This suggests that the multipotent RPCs could be used as a rich and excellent source to produce the missing target cells. Transplantation of cells from eye-like structures derived from mouse ESCs has been performed. Retinal ganglion-like cells were differentiated, and a small proportion of the ES-derived cells extended axon-like processes toward the optic disk of the host. Surprisingly, the transplanted cells formed a new RGC layer (multi- or monolayer) attached to the host retina after transplantation [42]. However, the complementary distribution of the transplanted cells and the host residual RGCs were not observed in the same GCL. In our opinion, there was no real integration for transplanting cells. Furthermore, apparent teratoma formation induced by ES cell-derived eye-like structures has been observed 10 days after transplantation [42]. Therefore, it is necessary to evaluate the feasibility of transplanting RPCs for degenerative retinal diseases. Furthermore, it is important to note that the transplanting RPCs should not contain undifferentiated cells.
Previous studies have shown that when neural stem cells or progenitors are transplanted into the murine vitreous cavity, their differentiation into mature neurons is limited, and they rarely form retina-specific neurons [43,44]. In our experiments, ∼20,000 RPCs extracted from hESC-derived optic cup-like structures were transplanted, and ∼0.5% of these cells successfully integrated into NMDA-treated mouse retina on the fifth week after transplantation. Our data suggested that hESC-derived RPCs have the capability to integrate and differentiate into RGC-like cells after transplantation into NMDA-treated mouse retina. We present clear evidence to support the utility of hESC-derived RPCs for ganglion cell replacement therapy.
For cell replacement therapy to treat retinal degenerative diseases, the transplanted retinal neurons must survive, integrate, and function physiologically. Although functional integration was not investigated in our work, the morphological integration of differentiated retinal ganglion-like cells derived from transplanted RPCs was well observed. Considering the transplantation of cells for the treatment of RGC degenerative diseases, there are three main obstacles to overcome. First, the largest challenge associated with this method is that new RGCs could be generated from transplanting cells to replace the damaged cells in the diseased retina. The generated RGCs are required not only to integrate in the GCL layer but also to extend long axons that connect to the optic nerve and develop synaptic connections to the target brain. Therefore, cell replacement therapy for RGCs is considered the most challenging of all retinal cells. Second, it is important to note that the GCL consists of approximately equal numbers of RGCs and displaced amacrine cells, both of which are affected in many degenerative diseases such as experimental glaucoma and optic nerve transection [45]. Transplantation of just one of these two cell types may not be enough to repair the damaged retina. However, a mixture of RGCs and amacrine cells might be a better choice. Lastly, the purity of transplanting cells is an important obstacle to overcome because the remaining undifferentiated cells in the culture could lead to teratoma formation in vivo. A recent published report found that tumors were reduced by ESCs after long-term survival after engraftment into the subretinal space [46]. Future transplantation studies will seek to develop safe efficient therapies using hESCs.
Conclusion
In conclusion, the data obtained in this study demonstrate that the RPCs from hESC-derived optic cup-like structures have the potential to migrate and integrate in the GCL of the degenerated mouse retina. They also may differentiate into RGC-like cells in vivo. Although some limitations still remain, our results suggest that hESCs could be used as an ideal source for producing enough transplanted RPCs, and transplanting the RPCs derived from hESCs might be another efficient strategy to restore visual function of individuals/patients with glaucoma in the future.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (grant nos. 31571238 and 31371083) and the research foundation for doctors of Xinxiang Medical University (grant no. XYBSKYZZ2015010). We gratefully appreciate the help by Professor Su-chun Zhang (Waisman Center, University of Wisconsin) and the support project for the disciplinary group of Physiology and Neuroscience, Xinxiang Medical University.
Author Disclosure Statement
The authors declare no conflicts of interest with regard to the publication of this article.
Supplementary Material
Supplementary Figure1
Supplementary Figure2
Supplementary Figure3
Supplementary Figure4
Supplementary Figure5
Supplementary Table1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.