
Abstract
Key transcription factors, which activate or repress master gene regulators and signaling pathways, tightly regulate self-renewal and cell lineage differentiation of bone marrow-derived stromal cells (BMSC). Among these factors is the basic helix-loop-helix transcription factor Twist-related protein 1 (TWIST-1), which is important in BMSC self-renewal, life span, and differentiation. Another layer of gene regulation comes from microRNAs (miRNAs). miRNAs are short noncoding RNAs that interfere with translation of specific target mRNAs and thereby regulate diverse biological processes, including BMSC lineage commitment. However, little is known of how TWIST-1-regulated miRNAs control osteogenic commitment, and influence the fate of bone precursor cells. In this study, we have discovered a novel TWIST-1-regulated miRNA, miR-376c-3p. Reduced miR-376c-3p expression by a miR-376c-3p inhibitor or due to TWIST-1 haploinsufficiency promotes alkaline phosphatase (ALP) activity, mineral deposition, and expression of osteoblast-associated genes in BMSC and calvarial cells. Conversely, overexpression of miR-376c-3p using a miR-376c-3p mimic inhibited BMSC proliferation and the osteogenic potential of BMSC and TWIST-1 haploinsufficient calvarial cells. This was demonstrated by a decrease in insulin growth factor 1 receptor (IGF1R) levels, Akt signaling, ALP activity, mineral deposition, and expression of osteoblast-associated genes. Thus, miR-376c-3p reduces IGF1R/Akt signaling in BMSC and is one mechanism by which osteogenesis may be inhibited. Overall, we have identified miR-376c-3p as a TWIST-1-regulated miRNA, which plays an important role in the osteogenesis of bone precursor cells and can mediate TWIST-1 inhibition of osteogenesis. Furthermore, overexpression of miRNA-376c-3p in TWIST-1 haploinsufficient calvarial cells can decrease the aberrant osteogenesis of these cells, which contributes to increased calvarial bone volume and premature fusion of the coronal sutures.
Introduction
T
TWIST-1 belongs to the TWIST family of genes comprising of basic helix-loop-helix (bHLH)-containing transcription factors [11,12]. TWIST-1 and the other members of its family are mediators of MSC self-renewal and lineage commitment in postnatal skeletal tissues and play an important role in mediating skeletal and mesodermal tissue development. TWIST-1 acts by homo- or heterodimerizing with other bHLH molecules, thus allowing it to bind to a conserved motif known as the E box (CANNTG) present on gene promoters, leading to the transcriptional activation or repression of genes [13 –15]. In vivo, Twist-1 expression is greatest at sites of immature bone cells and decreases during endochondral and intramembranous bone development [16] and in bone cell populations during calvarial and suture development [17].
During development, Twist-1 null mice die at embryonic day 10.5 due to failure of neural tube closure and defects in the head mesenchyme, somites, and limb buds [18]. Heterozygote animals display abnormal craniofacial structures, poor suture growth, and predominantly premature fusion of the coronal suture [4,18 –20]. They also display an asymmetrical facial skeleton and polydactylia of the hind limbs. The cause of the premature bony bridging and fusion between apposed skull plates results from the loss of TWIST-1 (TWIST-1 haploinsufficiency), which results in alterations in suture mesenchyme cellular gene expression patterns and increased osteogenesis of osteoprogenitor calvarial cells, and accelerated bone growth [21 –23].
Another intrinsic property of BMSC that has been attributed to regulating cell lineage differentiation is its microRNA (miRNA) expression profile [24]. miRNAs negatively regulate target protein expression through binding to specific complementary sequences in the 3-untranslated region (3-UTR) of target mRNAs and have been reported to be involved in the regulation of a variety of biological processes, as new modulators of gene regulation [25]. Thus, a number of miRNA expression profiling studies of human BMSC have been performed during osteogenic differentiation. These have revealed novel miRNAs, which regulate key players of lineage commitment and influence the fate of bone precursors [24,26 –29]. TWIST-1 positively regulates the miR-199a/214 cluster during development of neural cell populations [30]. Postnatally, miR-199a inhibits osteoblast proliferation by regulating the WNT signaling pathway [31]. Furthermore, miR-214 has been demonstrated to inhibit osteoblast differentiation of BMSC by targeting a number of signaling pathways [32 –34]. However, the relationship between TWIST-1 and miRNAs, and the function of TWIST-1-regulated mRNAs in biopotential osteogenic progenitor cells, such as BMSC, and in more committed calvarial preosteoblasts during osteogenesis remains largely undefined.
The present study sought to identify novel TWIST-1-inducible miRNA targets involved in BMSC osteogenic differentiation. Using miRNA microarray analysis of TWIST-1 overexpressing human BMSC lines, we identified miR-376c-3p as a novel miRNA that is upregulated by TWIST-1 during growth and osteogenic differentiation. We examined the role of miR-376c-3p in BMSC cell proliferation and osteogenic differentiation. By modulating miR-376c-3p activity, we show that inhibition of miR-376c-3p increased osteogenic differentiation of BMSC, whereas overexpression of miRNA-376c-3p decreased cell proliferation and osteogenic differentiation of BMSC in vitro. In addition, we identified insulin growth factor 1 receptor (IGF1R)/Akt signaling to be involved in miRNA-376c-3p-mediated regulation of BMSC osteogenic differentiation as miR-376c-3p inhibits IGF1R transcript and protein levels and inhibits IGF1R/Akt signaling, thereby reducing the osteogenic potential of BMSC. Furthermore, reduced levels of miR-376c-3p are suggested to promote the osteogenic differentiation of TWIST-1 haploinsufficient BMSC and calvarial cells. miR-376c-3p inhibited the osteogenic potential of calvarial cells and was able to rescue dysregulated osteogenesis due to TWIST-1 haploinsufficiency.
Materials and Methods
Isolation of human BMSC, retroviral transduction, and miRNA microarray analysis
Human BMSC were derived from bone marrow aspirates from the posterior iliac crest of normal adult volunteers (18–35 years of age) after obtaining informed consent in accordance with procedures approved by the Human Ethics Committee of the Royal Adelaide Hospital (protocol number 940911a) and cultured as previously described [8,35]. Full-length human sequence for TWIST-1 (Cat. No. MHS 1011-7508836 MGC IRAU) was subcloned into the pRuf-IRES-GFP vector and retroviral transduction of TWIST/pRuf-IRES-GFP or empty control pRuf-IRES-GFP into BMSC from three donors [8]. Stably transduced high-expressing GFP þMSC were selected by fluorescence-activated cell sorting and overexpression of TWIST-1 was confirmed by quantitative polymerase chain reaction (PCR) and western blot analysis [8]. For the miRNA microarray analysis, total RNA was isolated from 2 × 105 stably transfected BMSC previously cultured in noninductive growth medium or osteogenic induction medium for 5 days or 3 weeks using TRIzol (Sigma-Aldrich, Inc., Sydney, NSW, Australia) in accordance with the manufacturer's instructions. Microarray analysis was performed using Affymetrix miRNA microarray analysis (AGRF—http://agrf.org.au).
Isolation of mouse calvarial cells
Calvarial cells were obtained in accordance with the procedures approved by the Animal Ethics committee of the South Australian Health and Medical Research Institute (SAM142). Cells were isolated from 4-week-old wild-type and Twist-1del/+ heterozygous mice by first cutting out the calvaria from the rest of the skull using surgical scissors. Care was taken while extracting the calvaria to make sure that skin, soft tissues, and fibroblasts were removed. The extracted calvaria were cut into small pieces using a scalpel and incubated in 3 mg/mL Collagenase I and 50 U/μL DNAse I in phosphate-buffered saline at 37°C for 1 h Calvarial preosteoblasts were isolated by filtering out the cellular suspension from calvarial bone chips through a nylon falcon 70-μm cell strainer (Cat No. 342350; Becton Dickinson Australia, North Ryde, NSW, Australia). Bone chips were then subjected to a second round of Collagenase and DNAase treatment along with a second filtration step. Cells released from the digestions along with remaining bone chip fragments were cultured in growth media [alpha modification of Eagle's medium (αMEM) containing 20% fetal calf serum (FCS), 2 mM
Osteogenic differentiation and osteogenesis assays
Cells were cultured in osteogenic induction medium (Human BMSC: αMEM supplemented with 5% FCS, 2 mM
Transfection assays
Human BMSC and mouse calvarial cells were seeded at 2.2 × 104 cells per well (24-well plate) or 5 × 103 cells per well (96-well plate) the day before transfections were performed. For siRNA knockdown studies, human BMSC were transfected with either a validated sequence-specific siRNA against TWIST-1 (s14524) or negative siRNA#1 control (4390843) (Ambion/Life Technologies, Mulgrave, VIC, Australia,
Cell proliferation assay
Transfected cells were seeded at 2.5 × 103 (96-well plates) and assessed after 6 days using the Cell Proliferation ELISA, Bromodeoxyuridine (BrdU) Colorimetric Kit (Roche Products Pty Limited, Sydney, NSW, Australia) as per the manufacturer's instructions. Absorbance was read at 450 nM on an iMark microplate absorbance reader (Bio-Rad Laboratories, Hercules, CA).
RNA extractions and real-time polymerase chain reaction analysis
The anterior segment of the calvaria of day (P) 10 wild-type and Twist-1del/+ mice were retrieved and snap frozen in liquid nitrogen. Total RNA from cultured cells and mouse calvarial fragments was isolated using TRIzol Reagent (Cat No. 15596026; Invitrogen/Thermo Fisher Scientific) according to the manufacturer's instructions. Generation of cDNA and real-time PCR analysis was performed as previously described in triplicate [35]. Primer sets used in this study (GeneWorks Pty Ltd. Thebarton, SA, Australia) were: human β-actin (Fwd: 5′-gatcattgctcctcctgagc-3′; Rev: 5′-gtcatagtccgcctagaagcat-3′), Runt-related transcription factor 2 (RUNX2; Fwd: 5′-gtggacgaggcaagagtttca-3′; Rev: 5′-catcaagcttctgtctgtgcc-3′), osteopontin (OPN; Fwd: 5′-catccagtaccctgatgctacag-3′, Rev: 5′-gtgggtttcagcactctggt-3′), osteocalcin (OCN; Fwd: 5′-atgagagccctcacactcctcg-3′, Rev: 5′-gtcagccaactcgtcacagtcc-3′), TWIST-1 (Fwd: 5′-tcttacgaggagctgcagacgca-3′, Rev: 5′-atcttggagtccagctcgtcgct-3′), ID-1 (Fwd: 5′-cctcaacggcgagatcag-3′, Rev: 5′-cttcagcgacacaagatgc-3′), ID-2 (Fwd: 5′-attgtcagcctgcatcacc-3′, Rev: 5′-attcagaagcctgcaaggac-3′), IGF1R (Fwd: 5′-aggatattgggctttacaacctg-3′, Rev: 5′-gaggtaacagaggtcagcatttt-3′), mouse β-actin (Fwd: 5′-ttgctgacaggatgcagaag-3′, Rev: 5′-aagggtgtaaaacgcagctc-3′), mouse Twist-1 (Fwd: 5′-cagcgggtcatggctaac-3′, Rev: 5′-tgaatcttgctcagcttgtcc-3′), mouse Runx2 (Fwd: 5′-cctctgacttctgcctctgg-3′, Rev: 5′-tatggagtgctgctggtctg-3′), mouse Opn (Fwd: 5′-agcaagaaactcttccaagca-3′, Rev: 5′-gattcgtcagattcatccgact-3′), and mouse Ocn (Fwd: 5′-gatgggtacctgtcccacac-3′, Rev: 5′-agcaggaggcgtgaacttag-3′). To perform real-time quantitative PCR on miRNA expression, miRNA cDNA synthesis from 10 ng of RNA was performed using 5× TaqMan primers for miR-36c-3p and U6 and the MicroRNA Reverse Transcription Kit (Cat No. 4366596; Thermo Fisher Scientific). cDNA was then amplified with specific 20× TaqMan primers for miR-376c-3p and U6 using TaqMan Universal Master Mix with UNG (Cat No. 4440038; Thermo Fisher Scientific). The PCR conditions were as follows: 82°C for 2 min, 95°C for 10 min, and 50 cycles of 95°C for 15 s, 60°C for 30 s with the elongation step of 60°C for 15 s. MiR-376c-3p expression levels were measured relative to U6 expression levels using the Pfaffl method as previously described [36] (No. 77). miRNA primers used in this study were: human TaqMan assay miRNA 376c (ID No. 002122), mouse TaqMan assay miRNA 376c (ID No. 002450), and U6 snRNA (ID No. 001973, NCBI Acc#NR_004394).
Chromatin immunoprecipitation
Chromatin was isolated from cultured human BMSC (1 × 106) and chromatin immunoprecipitation (ChIP) was performed using the Magna ChIP Kit (Millipore Corporation, Billerica, MA,
Western blot analysis
Whole cell lysates were prepared from cells cultured in 9.5 cm2 wells in triplicate in 300 μL of lysis buffer per well (50 mM Tris/HCL pH 7.4; 1 mM ethylenediaminetetraaceticacid pH 8.0; 100 mM NaCl; 1% NP-40; 10 mM NaF; 0.5 mM phenylmethylsulfonyl fluoride; 5 mM sodium vanadate; 10 mM sodium pyrophosphate; 1× complete protease inhibitor; Roche Applied Science, Roche Diagnostics GmbH, Mannheim, Germany) and western blot analysis was performed as previously described [35]. The membranes were probed overnight at 4°C with the following primary antibodies diluted in 2.5% ECL membrane blocking agent (Cat No. RPN2125; GE Healthcare Australia, Parramatta, NSW, Australia): anti-rabbit IGF-1R (Cat No. ab39675, 1:400 dilution; Abcam, Cambridge. MA), anti-rabbit RUNX2 (M-70, Cat No. SC-10758, 1:500 dilution; Santa Cruz Biotechnology, Inc., Dallas, TX), anti-rabbit HSP90α/β (H-114, Cat No. SC7947, 1:1,000 dilution; Santa Cruz Biotechnology, Inc.), anti-mouse S6 (Cat No. 2317, 54D2, 1:500 dilution; Cell Signaling Technology, Danvers, MA), anti-rabbit phosphorylated-S6 (Ser235/236, Cat No. 2211S, 1:500 dilution; Cell Signaling Technology), anti-mouse beta-actin (Cat No. 8H10D10, 1:1,000 dilution; Cell Signaling Technology), anti-rabbit Akt (Cat No. 9272S, 1/500 dilution; Cell Signaling Technology), and anti-mouse phosphorylated Akt-Ser473 (Cat No. 4051S, 1/500 dilution; Cell Signaling Technology). Blots were washed three times with Tris buffered saline/0.1% Tween 20, then incubated for 1 h at room temperature with corresponding secondary detection antibodies. Secondary detection was performed using anti-rabbit alkaline phosphatase conjugate (AP) (Cat No. ab97048, 1:2,500 dilution, RRID:AB_10680574; Abcam) and anti-mouse alkaline phosphatase conjugate antibodies (AP, Cat No. AP124A, 1:2,500 dilution, RRID:AB_92456; Merck Life Science, Bayswater, VIC, Australia). After washing off secondary antibody, the blots were developed with enhanced chemifluorescence substrate (GE Healthcare UK Ltd., Buckinghamshire, England). Quantification of signal from multiple independent blots (n = 3) was performed using Image Lab Software (Bio-Rad Laboratories Pty., Ltd, Gladesville, NSW, Australia,
Statistics
Experiments were performed in triplicate. Data analysis was carried out using Microsoft GraphPad Prism 5 (GraphPad Software,
Results
miR-376c-3p expression decreases in BMSC during osteogenic differentiation and is upregulated by TWIST-1
Vector control and TWIST-1-overexpressing BMSC were cultured for 5 days or 3 weeks under normal growth culture conditions or under osteogenic induction (OI) conditions to identify novel TWIST-1-regulated miRNAs during BMSC osteogenic commitment. Differentially expressed (DE) miRNAs were selected based on a P value <0.05 with a fold change of 1.5-fold [a log fold change (logFC) ≥|0.58| or ≥|−0.6|]. Using the selection criteria, 28 DE miRNAs were identified following OI. The miRNAs identified were ranked according to fold change (Table 1) and plotted on a volcano plot, generated by plotting statistical significance (P value) over magnitude of fold change (Fig. 1A). Of these 28 miRNAs, 16 miRNAs were downregulated in TWIST-1-overexpressing BMSC, whereas 12 miRNAs were upregulated. Four miRNAs, which were upregulated in TWIST-1-overexpressing BMSC, were miR-199b, miR-424, miR-148a, and miR-195. These miRNAs have previously been shown to be expressed in BMSC and inhibit osteogenesis [38 –41].

miR-376c-3p expression is upregulated by TWIST-1.
Differentially Expressed microRNA (≥1.5-Fold) in Twist-1-Overexpressing Bone Marrow-Derived Stromal Cells Versus Empty Vector Control Bone Marrow-Derived Stromal Cells
One novel miRNA, miR-376c-3p (previously named miR-376c and miR-368), was found to be upregulated by more than two-fold during osteogenesis in TWIST-1-overexpressing BMSC compared with wild-type BMSC (Fig. 1A; Table 1). The microarray results obtained were validated using quantitative real-time PCR. Relative miR-376c-3p expression was higher in TWIST-1-overexpressing BMSC compared with empty vector control cells (Fig. 1B). During OI, miRNA-376c-3p expression decreased in both vector control and TWIST-1-overexpressing BMSC (Fig. 1B). However, the relative expression levels of miR-376c-3p during OI were higher in TWIST-1-overexpressing cells compared with vector control cells (Fig. 1B).
TWIST-1 expression was then knocked down in BMSC following transfection with a TWIST-1-specific siRNA-mediated molecule, which resulted in reduction in TWIST-1 expression, as determined by quantitative real-time PCR (Fig. 1C). BMSC in which TWIST-1 was knocked down exhibited a decrease in miR-376c-3p expression levels when cultured under normal growth culture conditions or under osteogenic induction conditions, compared with scrambled siRNA transfected control cells (Fig. 1D). These data imply that TWIST-1 regulates expression of miR-376c-3p.
To determine whether miR-376c-3p is directly regulated by TWIST-1, we used in silico analysis to determine the presence of E-box sites within the coding region of miR-367c on chromosome 14. One putative binding site, (CATATG) S1, was located at 3.2 kb upstream from the miR-376c stem loop. ChIP analysis was subsequently employed to assess genomic DNA isolated from human BMSC under normal growth culture conditions and during osteogenic induction conditions to demonstrate that TWIST-1 was recruited to the E-box binding site (Fig. 1E, F). The results indicated that TWIST-1 directly binds to the regulatory region of miR-376c during normal cell growth and osteogenic differentiation, and that TWIST-1 directly upregulates miR-376c-3p expression.
mir-376c-3p inhibits BMSC proliferation
To evaluate the effect of miR-376c-3p on BMSC proliferation, cells were transfected with either a miR-376c-3p inhibitor, a miR-376c-3p mimic, a nonspecific inhibitor control (miR-IC) or a nonspecific mimic control (miR-MC). Transfection of BMSC with miR-376c-3p inhibitor or miR-376c-3p mimic effectively decreased or increased total miR-376c-3p expression levels, respectively, compared with cells transfected with miR control molecules as demonstrated by quantitative real-time PCR (Fig. 2A, B). The relative proliferation rates of the different transfected cell lines were measured by BrdU incorporation. While reduced levels of miR-376c-3p in BMSC had no effect on cell proliferation, increased levels of miR-376c-3p reduced cellular proliferation when compared with the respective controls (Fig. 2C, D). Furthermore, real-time PCR analysis was utilized to observe any changes in the expression of two bHLH factors associated with cellular proliferation, ID-1 and ID-2 [42], previously shown to be upregulated in TWIST-1-overexpressing BMSC [8]. Decreased levels of miR-376c-3p did not alter ID-1 and ID-2 expression levels (Fig. 2E). However, an increase in miR-376c expression reduced the expression of ID-1 and ID-2 compared with control (Fig. 2F). Interestingly, miR-376c-3p overexpression led to reduced TWIST-1 transcript levels in BMSC even though there are no conserved binding sites for miR-376c-3p on the 3′UTR region of TWIST-1 (Fig. 2G). These results indicate that BMSC proliferation is inhibited by miR-376c-3p.
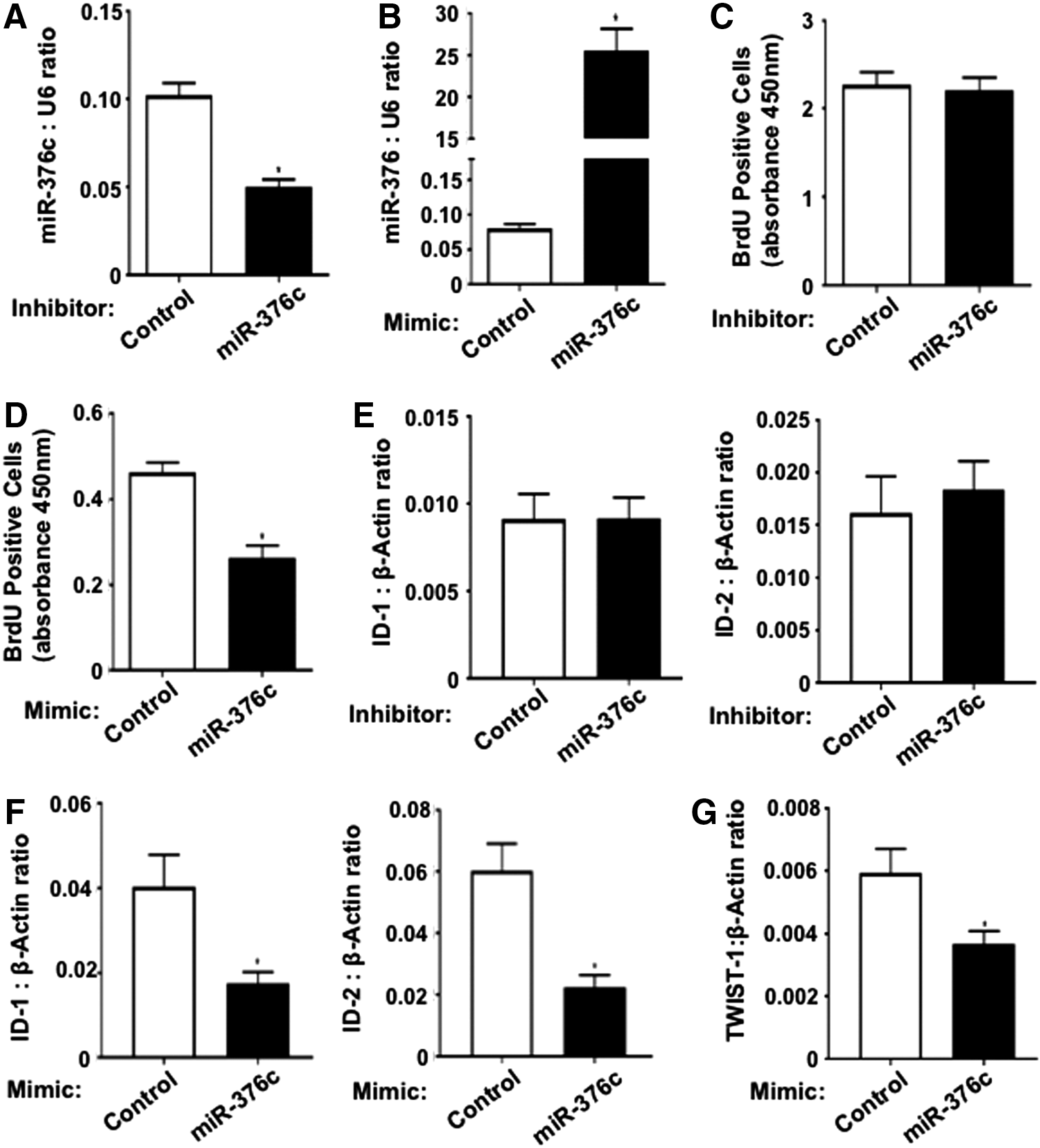
miR-376c-3p inhibits BMSC proliferation. Real-time PCR analysis of BMSC transfected with
Osteogenic differentiation and IGF1R/Akt signaling in BMSC is negatively regulated by miR-376c-3p
TWIST-1-overexpressing BMSC have reduced osteogenic potential [8] and increased miR-376c-3p expression. To evaluate the effect of miR-376c-3p on BMSC osteogenic differentiation, BMSC were transfected with either miR-376c-3p mimic or miR-MC, then cultured under osteogenic induction conditions. Transfection of BMSC with miR-376c-3p mimic effectively increased total miR-376c-3p expression levels compared with cells transfected with miR-MC as demonstrated by quantitative real-time PCR (Fig. 3A). ALP activity, an early marker of osteogenesis and osteoblast function in vitro, was measured after 1 week of osteogenic induction conditions. Staining for ALP activity showed visibly reduced levels in cells transfected with miR-376c-3p mimic compared with control, which was confirmed in quantitative studies of ALP enzymatic activity (Fig. 3B). Transfection of miR-376c-3p mimic also inhibited the expression of the osteogenic markers, Runt-related transcription factor 2 (RUNX2), Osteopontin (OPN), and Osteocalcin (OCN) compared with control when cells were cultured in osteogenic induction conditions for 2 weeks (Fig. 3C). Conversely when BMSC were transfected with miR-376c-3p inhibitor or miR-IC and cultured under osteogenic induction conditions, total miR-376c-3p expression levels were reduced (Fig. 3D) and was associated with increased levels of ALP activity (Fig. 3E), an increase in the formation of Alizarin Red-positive mineral deposits (Supplementary Fig. S1; Supplementary Data are available online at

Modulating miR-376c-3p levels can alter the osteogenic potential of BMSC.
Studies of miR-376c have previously demonstrated its binding to the 3′UTR of IGF1R in melanoma cell lines [43]. IGF1R is a receptor tyrosine kinase and stable overexpression of miR-376c in melanoma cell lines resulted in decreased IGF1R mRNA and protein [43]. The effects of increased levels of miR-376c-3p in BMSC during osteogenic differentiation were investigated further by analyzing the levels of IGF1R by quantitative real-time PCR and western blot analysis. BMSC transfected with miR-376c-3p mimic expressed reduced levels of IGF1R transcript, IGF1R protein, phosphorylated Akt (Ser473), and the downstream target phosphorylated ribosomal S6 kinase (Ser235/236) protein during osteogenic differentiation compared with control with no changes in total Akt or total ribosomal S6 kinase protein levels (Fig. 4A–C). In addition, RUNX2 protein levels were reduced, confirming the observed decrease in RUNX2 transcript expression levels (Figs. 3C and 4B, C). Moreover, increased levels of miR-376c-3p in BMSC resulted in significant reduction in the formation of Alizarin Red-positive mineral deposits, corresponding to a decrease in levels of extracellular calcium compared with control (Fig. 4D). These results indicate that miR-376c-3p inhibits the osteogenic potential of BMSC by reducing IGF1R levels and IGF1R/Akt signaling. Supportive studies found that IGF1R expression was decreased in TWIST-1-overexpressing cells compared with vector control cells, under osteogenic induction conditions (Supplementary Fig. S2A). Therefore, TWIST-1 appears to regulate IGF1R levels during osteogenic differentiation and suggests that miRNA-376c-3p may mediate TWIST-1 inhibition of BMSC osteogenesis in vivo.
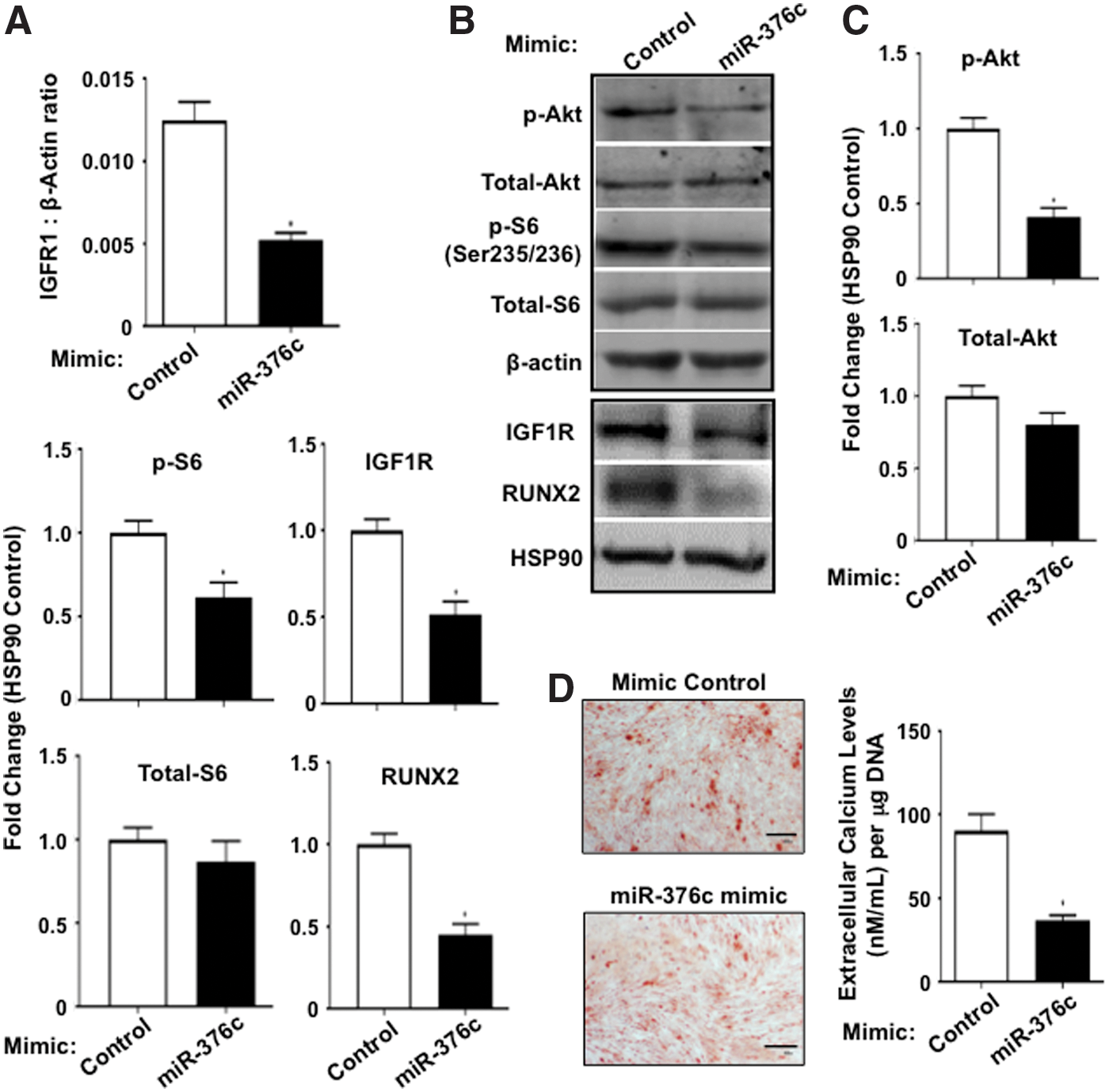
Overexpression of miR-376c-3p reduces IGF1R/Akt signaling and the osteogenic potential of BMSC.
miR-376c-3p alleviates aberrant osteogenic differentiation of TWIST-1 haploinsufficient calvarial cells
TWIST-1 haploinsufficiency results in aberrant osteogenesis, accelerated bone growth, and premature fusion of the coronal sutures of the cranium [21 –23]. The coronal sutures of Twist-1del/+ heterozygous mice fuse between postnatal (P) day 9 (P9) and day 15 (P15) [19,20]. By age P10, there is not only evidence of coronal suture fusion but also an increase in bone volume in Twist-1del/+ heterozygous mice crania compared with wild-type littermates [44]. Analysis of Twist-1 and miR-376c-3p expression levels in samples isolated from the coronal sutures of P10 animals revealed a significant decrease in both Twist-1 and miR-376c-3p transcript levels in Twist-1del/+ heterozygous mice, compared with wild-type controls (Fig. 5A). Furthermore, calvarial cells from Twist-1del/+ heterozygous mice exhibited increased ALP activity (Fig. 5B) and formation of Alizarin Red-positive mineral deposits, corresponding to increased levels of extracellular calcium (Fig. 5C), when cultured under osteogenic induction conditions, compared with calvarial cells from wild-type animals. These data suggest that reduced levels of miR-376c-3p in calvaria of Twist-1del/+ heterozygous mice may play a role in promoting the osteogenic differentiation of these cells and facilitating coronal suture fusion and increased bone volume in vivo.

Overexpression of miR-376c-3p can reduce the osteogenic potential of Twist-1 haploinsufficient calvarial cells.
To determine the effects of increasing miR-376c-3p levels in TWIST-1 haploinsufficient calvarial cells during osteogenesis, cells were transfected with miR-376c-3p mimic or miR-MC and cultured under osteogenic induction conditions. Cell cultures transfected with miR-376c-3p mimic exhibited a significant reduction in ALP activity (Fig. 5D), and decreased Alizarin Red-positive mineral deposits and levels of extracellular calcium (Fig. 5E), when compared with miR-MC controls. In agreement with these findings, TWIST-1 haploinsufficient calvarial cells transfected with miR-376c-376c mimic resulted in a significant reduction in the expression of the osteogenic markers, Runx2, Opn, and Ocn, when compared with miR-MC controls (Fig. 5F). Transfection of miR-376c-3p mimic was able to rescue the aberrant osteogenesis of calvarial cells from Twist-1del/+ mice, thus the increased osteogenic potential of TWIST-1 haploinsufficient calvarial cells is inhibited by miR-376c-3p.
Discussion
In an attempt to better understand the relationship between TWIST-1 and miRNAs in the regulation of human BMSC cell fate determination, we performed a miRNA microarray analysis to identify novel TWIST-1-regulated miRNAs during osteogenic differentiation. This array identified a number of miRNAs, which were upregulated or suppressed during osteogenic differentiation. TWIST-1 expression inhibits the osteogenic potential of BMSC and due to the suppressive action of miRNAs on downstream targets; we decided to focus on TWIST-1-regulated miRNAs that were upregulated by the expression of TWIST-1. One miRNA identified as a novel putative TWIST-1 target normally suppressed in BMSC during osteogenic differentiation was miR-376c-3p. In human BMSC, miR-376c-3p expression was greater in TWIST-1-overexpressing BMSC during growth conditions and osteogenic differentiation. ChIP analysis confirmed that TWIST-1 directly binds to the regulatory region of miR-376c during growth and osteogenic induction conditions and positively regulates its expression.
Previous studies have demonstrated that miR-376c inhibits cell proliferation and invasion in a number of cancer cell types, including in osteosarcoma [45], oral squamous cancer cells [46], and cervical cancer cells [47]. In BMSC, miR-376c-3p inhibited cellular proliferation and the expression of the bHLH factors, ID-1 and ID-2. ID proteins are generally considered as positive regulators of cell cycling [42] and facilitate the S phase of the cell cycle in embryonic-derived fibroblasts [48]. TWIST-1 upregulates miR-376c-3p expression in BMSC. However, TWIST-1 also promotes cell growth and the expression of ID-1 and ID-2 [8]. Conversely, when TWIST-1 is knocked down, expression of ID-1 and ID-2 are decreased [8]. In the present study, gene expression analysis found that TWIST-1 expression was decreased in miR-376c-3p-overexpressing BMSC, indicating that miR-376c-3p can modulate TWIST-1 expression indirectly during cell growth. Decreasing the levels of TWIST-1 could be one mechanism by which miR-376c-3p inhibits proliferation of BMSC and the decrease in ID-1 and ID-2 expression. This also alludes to the existence of a negative feedback mechanism between TWIST-1 and miR-376c-3p.
Our study demonstrates for the first time that during osteogenic differentiation TWIST-1 positively regulates miR-376c-3p. TWIST-1-overexpressing BMSC were shown to have reduced osteogenic potential and an increase in miRNA-376c-3p expression. To determine whether miR-376c-3p directly regulates BMSC osteogenic differentiation we employed the use of a miRNA-376c-3p mimic or inhibitor to overexpress or inhibit levels of miRNA-376c-3p, respectively. miR-376c-3p overexpression inhibited the osteogenic differentiation of human BMSC as assessed by a decrease in ALP activity, mineral nodule formation, extracellular calcium production, and the reduced expression of the early preosteoblast marker, RUNX2, and the osteoblast-associated markers, OPN and OCN. In contrast, inhibition of miR-376c-3p expression increased the osteogenic differentiation of human BMSC. The inhibitory effect of miR-376c3p on human BMSC osteogenic differentiation is consistent with a recent study demonstrating that miR-376c can inhibit osteogenic differentiation of cultured murine calvarial cells [49]. The reduced osteogenic potential observed in both human BMSC and mouse calvarial cells due to an increase in miR-376c-3p expression demonstrates for the first time the functional role of miR-376c-3p in the osteogenic differentiation in both biopotential osteogenic progenitor cells and in more committed osteoblasts.
One known gene target of miR-376c in melanoma is IGF1R [43]. Insulin growth factor-1 (IGF-1) signals through IGF1R to promote the osteogenic differentiation of rat MSC [50]. Decoupling of IGF1R signaling from IGF-1 in osteoblasts derived from osteoporosis patients, results in reduced differentiation and bone loss [51]. Furthermore, disruption of IGF1R in mouse osteoblasts in vivo ablated IGF signaling and impaired the rate of bone formation and mineralization, resulting in decreased cancellous bone volume and altered trabecular structures [52]. In BMSC, we found that miR-376c-3p targets IGF1R during osteogenic differentiation. miR-376c-3p overexpression reduced the levels of IGF1R transcript, IGF1R protein, and its downstream signaling target, phosphorylated ribosomal S6 kinase (Ser235/236) protein without affecting total S6 kinase protein levels. In addition, western blot analysis demonstrated a decrease in Akt signaling demonstrated by reduced phosphorylated Akt (Ser473) protein levels without affecting total Akt protein levels. Akt signaling has been demonstrated to be a crucial regulator of osteoblasts by promoting their differentiation and survival [53,54]. Reduced Akt has been shown in MSC to reduce osteoblast differentiation [54]. Thus, one molecular mechanism by which miR-376c-3p appears to inhibit the osteogenic potential of BMSC is through the inhibition of IGF1R/IGF/Akt signaling. Overall, miR-376c-3p expression reduced IGF1R/Akt signaling, resulting in reduced ALP activity, decreased RUNX2, OPN, and OCN expression, and reduced formation of Alizarin Red-positive mineral deposits, corresponding to a decrease in levels of extracellular calcium. Freshly isolated immature human BMSC express high levels of TWIST-1, which is downregulated during maturation [5,8]. TWIST-1 overexpression resulted in increased levels of miR-376c-3p, a key regulator of BMSC osteogenesis in vitro. High levels of miR-376c-3p in TWIST-1-overexpressing cells inhibit IGF1R levels during osteogenic differentiation. This alludes to the possibility that during osteogenic differentiation, the level of IGF1R present in BMSC is a downstream effector of a TWIST-1-miR-376c-3p axis, which exists in BMSC and that miRNA-376c-3p mediates TWIST-1 inhibition of BMSC osteogenesis in vitro and in vivo.
Another target of miR-376c found in osteoblast is Wnt3 [49], a member of the Wnt family, which plays an important role in osteoblastogenesis and bone formation [55]. MiR-376c is thus able to act through various molecular mechanisms to inhibit the osteogenic potential of BMSC and osteoblasts.
Knockdown of TWIST-1 resulted in reduced miR-376c-3p expression in BMSC. Furthermore, calvarial cells from Twist-1del/+ heterozygous mice expressed reduced levels of miR-376c-3p compared with cells from wild-type littermates. In our study, we observed that inhibition of miR-376c-3p expression increased the osteogenic potential of human BMSC. In addition, we found that calvarial cells from Twist-1del/+ heterozygous mice exhibited a greater osteogenic potential than calvarial cells from wild-type littermates, when cultured under osteogenic induction conditions. This was assessed by increased ALP activity, increased number of Alizarin Red-positive mineral deposits and increased extracellular calcium levels compared with wild-type calvarial cells. Furthermore, reduced miR-376c-3p expression in mouse calvaria from P10 Twist-1 del/+ animals correlated with coronal suture fusion and increased cranial bone volume when compared with crania from wild-type littermates. This suggests that reduced levels of miR-376c-3p promote the osteogenic differentiation of TWIST-1 haploinsufficient BMSC and calvarial cells.
miR-376c inhibits osteogenic differentiation of calvarial osteoblasts [49]. To study the relationship between TWIST-1 and miR-376c-3p in more committed calvarial preosteoblasts during osteogenesis, we investigated whether overexpression of miR-376c-3p in TWIST-1 haploinsufficient calvarial cells could inhibit their osteogenic potential. Our study showed that transfection of TWIST-1 haploinsufficient calvarial cells from Twist-1 del/+ mice with mouse miR-376c-3p mimic resulted in decreased ALP activity, mineral deposition rates, and bone gene expression levels. Collectively, these findings indicate that a rescue of the aberrant miR-376c-3p levels found in TWIST-1 haploinsufficient calvarial osteogenic progenitor cells can reduce the osteogenic potential of these cells in vitro.
In conclusion, we find that miR-376c-3p is a novel target of TWIST-1, which plays a role in BMSC proliferation. In addition, miR-376c-3p mediates TWIST-1 inhibition of BMSC osteogenic potential by repressing IGF1R expression and IGF1R/Akt signaling. Furthermore, we show, for the first time that miR-376c-3p inhibited the osteogenic potential of Twist-1 mutant calvarial cells, and is a novel miRNA with the ability to rescue dysregulated osteogenesis due to TWIST-1 haploinsufficiency. It appears that miR-376c-3p is a negative regulator of osteogenesis in both BMCS, osteogenic progenitor cells, and in calvarial cells, more committed pre-osteoblasts. Thus, miR-376c-3p or its target genes may be exploited in the future as novel therapeutic tools to alleviate skeletal abnormalities due to TWIST-1 haploinsufficiency.
Footnotes
Acknowledgments
This study was supported, in part, by NHMRC project grant #APP1046053 and Channel 7 Children's Research Foundation grant #151142.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.