
Abstract
Normal facial nerve (FN) function is very important for human being. However, if injured, FN function is difficult to restore completely. Recently, many studies reported the immune regulation function of stem cells (SCs). However, the immunomodulation function of SCs on FN injury is still unclear. Our study aims to explore the mechanism of immunomodulation effect of Sprague–Dawley rat bone marrow-derived SCs (BMSCs) on FN injury and specially focus on the regulation of Th17 and the protection effects of BMSCs on central facial motor neurons (FMNs). First, rat FNs were harvested. FN and BMSCs were cultured together or separately and levels of transforming growth factor (TGF)-β1, interleukin (IL)-6, hepatocyte growth factor (HGF), inducible nitric oxide synthase (iNOS), and prostaglandin E2 (PGE2) in supernatant were detected by enzyme-linked immunosorbent assay (ELISA). Then, after treating with or without local BMSCs injection, the proportion of Th17 in neck lymph nodes (LNs) was investigated in rat FN injury models. Furthermore, the apoptotic index of FMNs was studied in rat FN injury models that were treated with or without BMSCs. We found that BMSCs could secrete high levels of IL-6, HGF, PGE2, iNOS, and TGF-β1 in culture. The percentage of Th17 of neck LNs in BMSCs-treated group was significantly lower than that in the control group. The apoptotic index of FMNs in BMSCs-treated group was significantly lower than that in the control group. In conclusion, our research indicates BMSCs could independently secrete cytokines IL-6, HGF, PGE2, iNOS, and TGF-β1, and these cytokines could regulate the balance among subsets of CD4+ T cells and could protect FMNs by inhibiting neuron apoptosis.
Introduction
The facial nerve (FN) is a compound nerve containing special visceral motor, parasympathetic, special visceral sensory, and general somatosensory fibers and mainly plays roles in motor functions [1]. The continuity of the FN is typically restored using microsurgical techniques in the clinic; however, the recovery of neurological function remains unsatisfactory [2]. Many studies indicate that the CD4+ T cell-mediated adaptive immune response plays an important role in the maintenance of the activity of facial motor neurons (FMNs) after FN injury [3]. Jones et al. [4] and Sanders and Jones [5] proposed the adaptive immune response model based on their studies. Unknown specific antigens are released after FN injury. Peripheral antigen-presenting cells present them to CD4+ T cells in peripheral draining lymph nodes (LNs) to cause activation of CD4+ T cells. In addition, microglia in the central nervous system (CNS) release chemokines to attract peripheral activated CD4+ T cells to penetrate the blood–brain barrier. CD4+ T cells are reactivated to provide nutritional contact with microglia and other cells to maintain the survival of FMNs and/or release corresponding soluble mediating signals. Various cytokines and chemokines, such as interleukin (IL)-10 [6,7], IL-2 [8], CCL11 [9,10], and IL-15 [11], can affect microglial function and peripheral T cell recruitment [12].
Stem cells (SCs) are a group of cells with high proliferation capacity and self-renewal capacity with pluripotency. Bone marrow-derived SCs (BMSCs) can promote nerve fiber regeneration and angiogenesis, reduce neuronal apoptosis [13], and promote targeted homing to the injury site [14] and migration and differentiation of endogenous SCs [15]. Numerous studies have demonstrated that fetal bone marrow-derived mesenchymal SCs can promote Th17 cell proliferation through IL-6 and IL-1 in vitro [16]. Other studies [17,18] have shown that BMSCs can inhibit the differentiation of CD4+ naive T cells into Th17 cells in vitro, although BMSCs secrete high levels of cytokines required for Th17 differentiation, such as transforming growth factor (TGF)-β and IL-6. It might be associated with IL-10 and prostaglandin E2 (PGE2). Therefore, the effect of BMSCs on CD4+ T cell function may be associated with the release of various soluble cytokines [19].
We performed in vivo and in vitro experiments to observe the levels of inflammation-related cytokines secreted by Sprague–Dawley (SD) rat bone marrow-derived SCs (SDBMSCs) in different culture conditions, compared the Th17 cell percentage in cervical draining LNs among different experimental groups, and collected FN nuclei to compare the apoptosis of FMNs. According to the data, we analyzed the effect of SDBMSCs on the Th17 subset percentage, the underlying mechanisms, and the protective function on FMNs.
Experimental Materials and Methods
Major instruments, reagents, and supplies
Accuri C6 flow cytometer (BD Biosciences), 10% chloral hydrate (Peking Union Medical Hospital),fetal bovine serum (FBS), penicillin/streptomycin solution (Sigma), Phosphate-buffered saline (PBS), DMEM/F12 culture medium (HyClone), TrypLE Express (1 × ) (Gibco), the 0.4% Trypan blue solution, erythrocyte lysis buffer (Solarbio, China), the PMA/ionomycin mixture (250 × ), FIX&PERM kit, BFA/monensin mixture (250 × ), flow cytometry staining buffer (Lianke, China), the FITC mouse anti-rat CD4 antibody (BD Biosciences), the PE-cyanine7 anti-rat IL-17A antibody and the isotype control (eBioscience), P2 SDBMSCs (CCTCC Wuhan Cell Bank, China), the multifunctional microplate reader (Bio Tek) and the rat IL-6/hepatocyte growth factor (HGF)/PGE2/inducible nitric oxide synthase (iNOS)/TGF-β1 ELISA 48T Reagent Kit (Reanta, China).
Our study has obtained the approval of Ethics Committee of Peking Union Medical College Hospital and Ethics Committee of the Basic Medical Sciences, Chinese Academy of Medical Sciences (no. 003-2014). SD rats were purchased from the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences, and used according to the Regulations of Animal Management of the Chinese Academy of Medical Sciences (housed in sterile conditions; body weight 280–320 g). Data were analyzed using SPSS24.0.
Experimental methods
Culture of SDBMSCs in vitro: The culture medium was DMEM/F12 (containing 5% heat-inactivated FBS, 100 U penicillin/streptomycin). P4 was the last generation, and the culture was terminated thereafter. To minimize bias in experimental results, local injection into SD rats in subsequent experiments was performed using P4 cells on day 3 of passage.
Detection of cytokine expression in SDBMSCs using ELISA: The FN trunk was removed and placed in 4°C PBS for future use after surrounding connective tissues were removed. SDBMSCs were cultured in different groups as shown in Table 1, and two plates were used. Cells were cultured in a 37°C and 5% CO2 thermostat incubator for 72 h. Cytokines, including TGF-β1, IL-6, HGF, iNOS, and PGE2, in the supernatant were detected using ELISA.
Culture of Sprague–Dawley Rat Bone Marrow-Derived Stem Cells in Different Groups (12-Well Plates)
FN: FN including PBS for immersion of FN during collection and preservation of FN.
FN, facial nerve; PBS, phosphate-buffered saline; SDBMSC, Sprague–Dawley rat bone marrow-derived stem cell.
Detection of CD4+ T cells and the Th17 subset percentage in lymphocytes in cervical LNs in SD rats using flow cytometry
SD rats were grouped according to the following conditions: (A) no surgical operation; (B) bilateral dissection without transection of the FN trunk, injection of PBS at the left and SDBMSCs at the right side; (C) bilateral transection of the FN trunk, injection of PBS at the left and SDBMSCs at the right side. Each group contained six SD rats. The cervical LNs were collected 9 days after surgery, and CD4+ T cells and the Th17 subset were detected using flow cytometry.
Dissection and surgical operation of rats in groups B and C: According to the grouping, the FN trunk at the opening of the stylomastoid foramen was cut or preserved. During the transection, 3 mm was removed to prevent regeneration. The right side was injected with 1 × 106 SDBMSCs, and the left side was injected with an equal volume of PBS. When rats were awake, the SD rats in group C lost whisker movement ability. The blink reflex was slow, and the eye closure range became weaker or could not be induced. Animals were continuously housed for 9 days according to their grouping, and facial paralysis was observed daily.
Collection of LNs from rats in groups A, B, and C and preparation of the lymphocyte suspension: The left and right bilateral neck LNs were collected. LNs were ground and the cell suspension was collected.19 Erythrocyte lysis buffer was used to remove the red cells. Cells were resuspended and the cell concentration was adjusted to 2 × 106 cells/mL.
Staining of lymphocytes from rats in groups A, B, and C for flow cytometry: 1 mL of a 2 × 106 cells/mL cell suspension from each sample was collected, mixed with PMA/ionomycin and BFA/monensin, and cultured for 4 h. Sample detection tubes were set up as follows: tube 1, CD4-FITC/Rat IgG2a-pecy7; tube 2, CD4-FITC/IL-17A-pecy7.
The flow cytometry staining buffer was added to resuspend cells. The suspension was filtered and loaded onto the machine for analyses.
Analysis of the effect of SDBMSCs on the survival of FMNs using the TUNEL staining technique
SD rats were grouped based on the following conditions: (D) without any operation; (E) bilateral transection of the FN trunk, injection of PBS at the left and SDBMSCs at the right side. Each group contained six SD rats. On day 21 after surgery, groups D and E received cardiac perfusion for fixation and brain collection. Coronal brain slices containing FN nuclei were sectioned for TUNEL staining to analyze the survival of FMNs.
Dissection and surgical operation of rats in the D and E groups
The FN trunk at the opening of the stylomastoid foramen was cut. The right side was injected with 1 × 106 of SDBMSCs, and the left side was injected with an equal volume of PBS. When rats were awake, they were continuously housed for 21 days.
Cardiac perfusion for fixation and brain collection in SD rats: Exposed the heart and the surrounding large blood vessels. Rapidly perfuse 150–200 mL 4°C PBS until the four limbs of rats became white and the liquid flowing from the right auricula was colorless and transparent. The perfusion was continued using 150–200 mL of 4% paraformaldehyde at 4°C. If rats exhibited a struggling reaction and the four limbs and tail became stiff, the perfusion and fixation were successful. The perfusion was continued for an additional 1–2 min. The brain tissues were completely removed and placed in 4% paraformaldehyde at 10 times volume and stored at 4°C for future section staining and analysis.
TUNEL staining of coronal brain sections of SD rats: Brain stem tissue blocks containing FN nuclei were resected according to The Rat Brain in Stereotaxic Coordinates [20] and prepared into 4-μm sections. Five fields of each section were randomly selected under a high-power field. The total number of apoptotic motor neurons and FMNs were counted, and the mean value was calculated. The apoptotic index was calculated based on the following formula: apoptotic index = total number of apoptotic FMNs/the total number of motor neurons × 100%. The apoptosis indexes between the right and the left sides were compared. Statistical analysis was performed using SPSS24.0. An independent samples t-test was performed with significance set at P < 0.05 (Table 2).
Comparison of the Apoptosis Index of Facial Motor Neurons Between the Left and the Right Sides (
Compared with left side, P < 0.05.
Results
Detection of cytokine expression in SDBMSCs using ELISA
The levels of TGF-β1, IL-6, HGF, iNOS, and PGE2 in the SDBMSCs group (331.12 ± 17.15 pg/mL, 13.73 ± 0.72 pg/mL, 5.59 ± 0.58 ng/mL, 43.79 ± 5.80 IU/mL, and 72.30 ± 8.97 pg/mL) were significantly increased compared with those in the Blank group (189.52 ± 11.33 pg/mL, 10.79 ± 0.61 pg/mL, 3.90 ± 0.78 ng/mL, 28.55 ± 4.63 IU/mL, and 53.34 ± 7.62 pg/mL; P < 0.05). The cytokine levels in the FN+SDBMSCs group (362.53 ± 20.08 pg/mL, 16.82 ± 1.42 pg/mL, 8.03 ± 1.04 ng/mL, 45.62 ± 5.89 IU/mL, and 86.45 ± 10.06 pg/mL) were significantly increased compared with those in the FN group (199.12 ± 16.26 pg/mL, 11.41 ± 1.16 pg/mL, 4.29 ± 0.79 ng/mL, 25.18 ± 4.32 IU/mL, and 55.55 ± 6.82 pg/mL; P < 0.05) (Fig. 1). Besides, the cytokine levels in the FN + SDBMSCs group were higher than those in the SDBMSCs group.
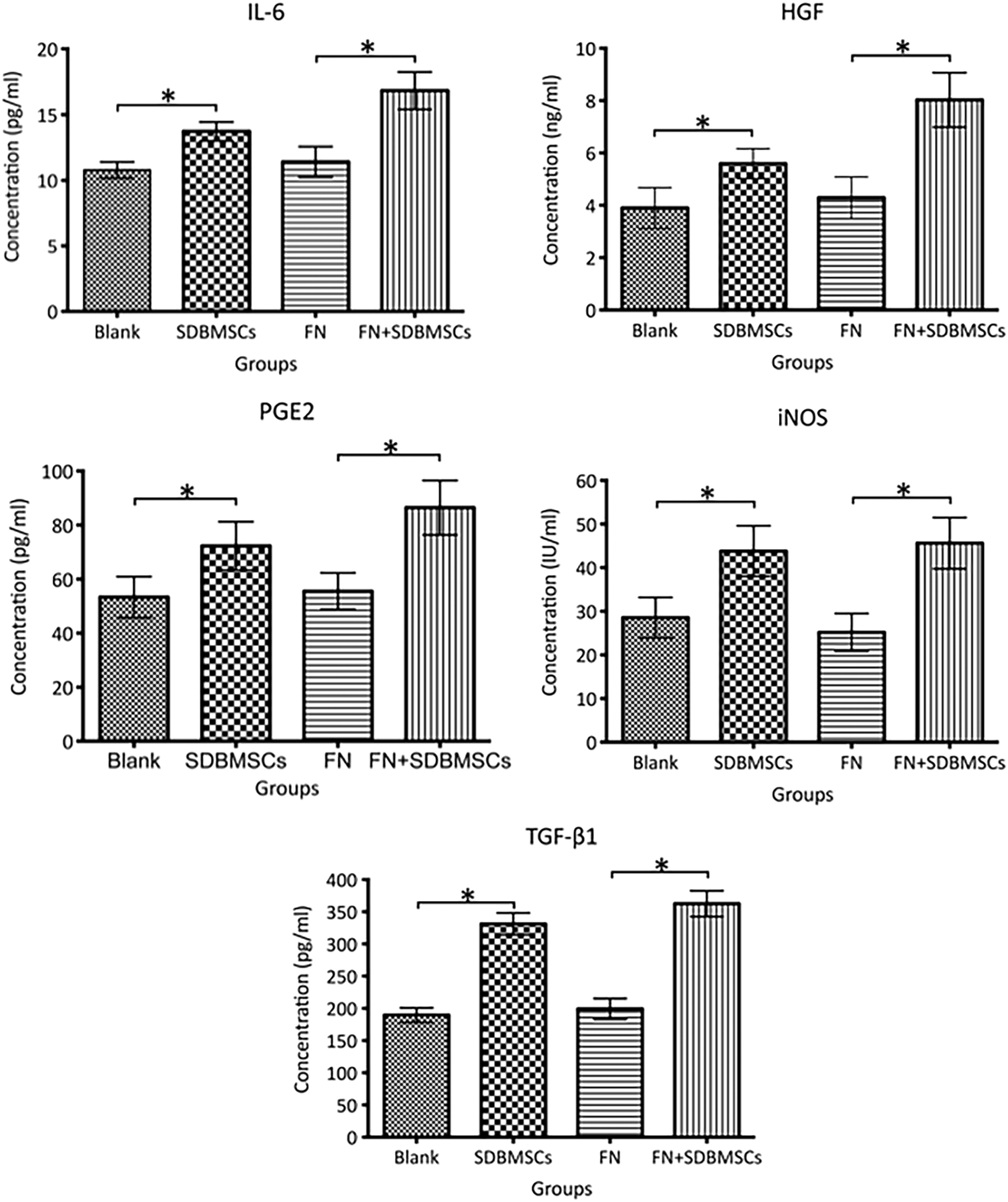
IL-6, HGF, PGE2, iNOS, and TGF-β1 levels in the culture supernatant in different experimental groups detected by ELISA. *P < 0.05. HGF, hepatocyte growth factor; IL, interleukin; iNOS, inducible nitric oxide synthase; PGE2, prostaglandin E2; TGF-β1, transforming growth factor-β1.
Detection of CD4+ T cells and Th17 subset percentages in lymphocytes in cervical draining LNs of SD rats using flow cytometry
The Th17 subset percentages in the bilateral cervical draining LNs in groups B (2.67% ± 0.18%) and C (3.08% ± 0.69%) that underwent surgery were increased compared with those in group A (2.10% ± 0.11%; P < 0.05). The Th17 subset percentage in bilateral cervical draining LNs in group C was increased compared with that in group B. The Th17 subset percentages in the right cervical draining LNs injected with SDBMSCs in groups B (2.13% ± 0.30%) and C (2.19% ± 0.13%) were both reduced compared with those in the left cervical draining LNs without injection of SDBMSCs in groups B (2.67% ± 0.18%) and C (3.08% ± 0.69%; P < 0.05) (Fig. 2).
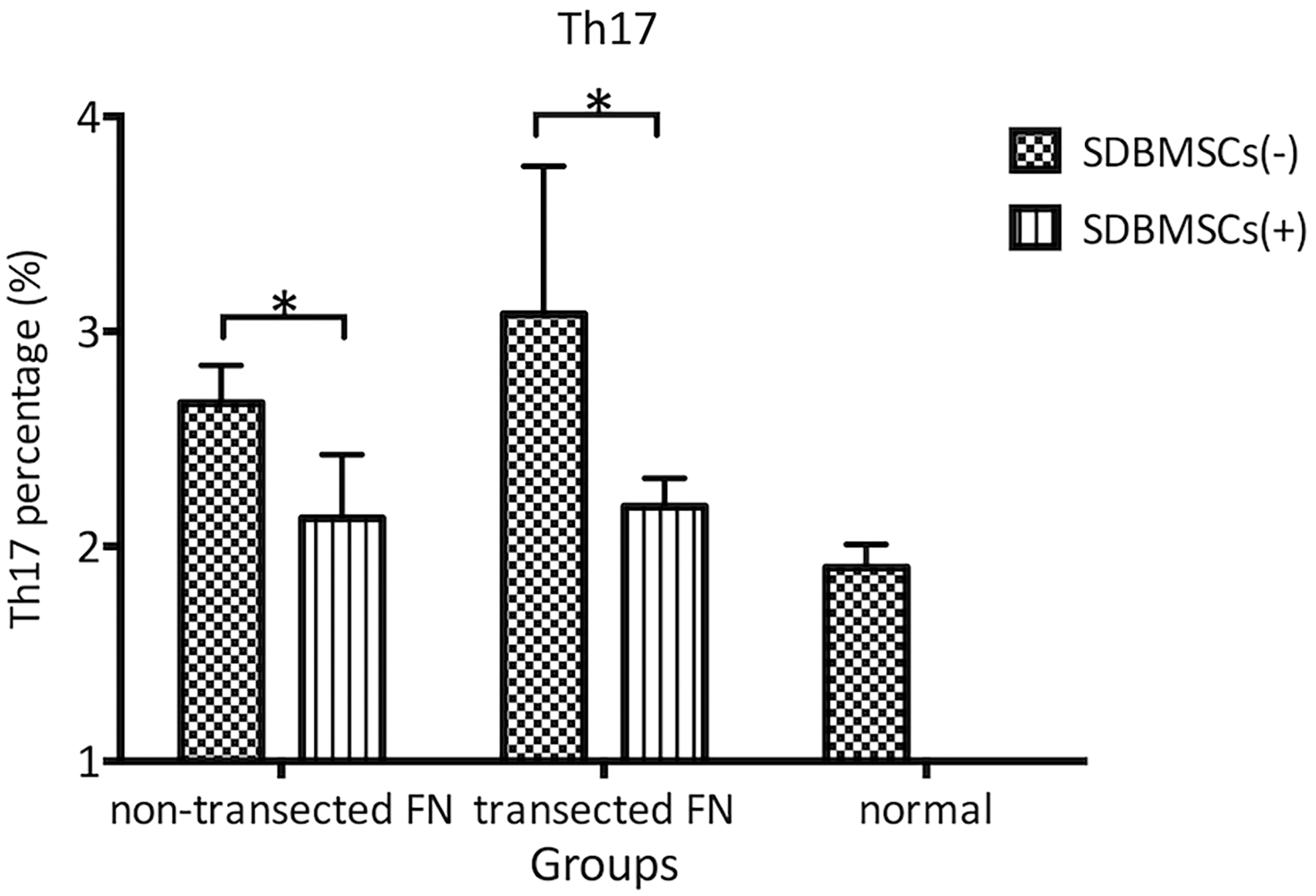
Detection of the Th17 percentages in bilateral cervical LNs with nontransected FN and transected FN using flow cytometry. *P < 0.05. FN, facial nerve; LN, lymph node.
Analyses of the effect of SDBMSCs on the survival of FMNs using the TUNEL staining technique
Compared with FN nuclei in normal SD rats without surgical operation, bilateral FN nuclei in SD rats with bilateral transection of the FN trunk exhibited more obvious apoptosis in FMNs. The apoptosis index of the right side was significantly reduced compared with that of the left side (P < 0.05).
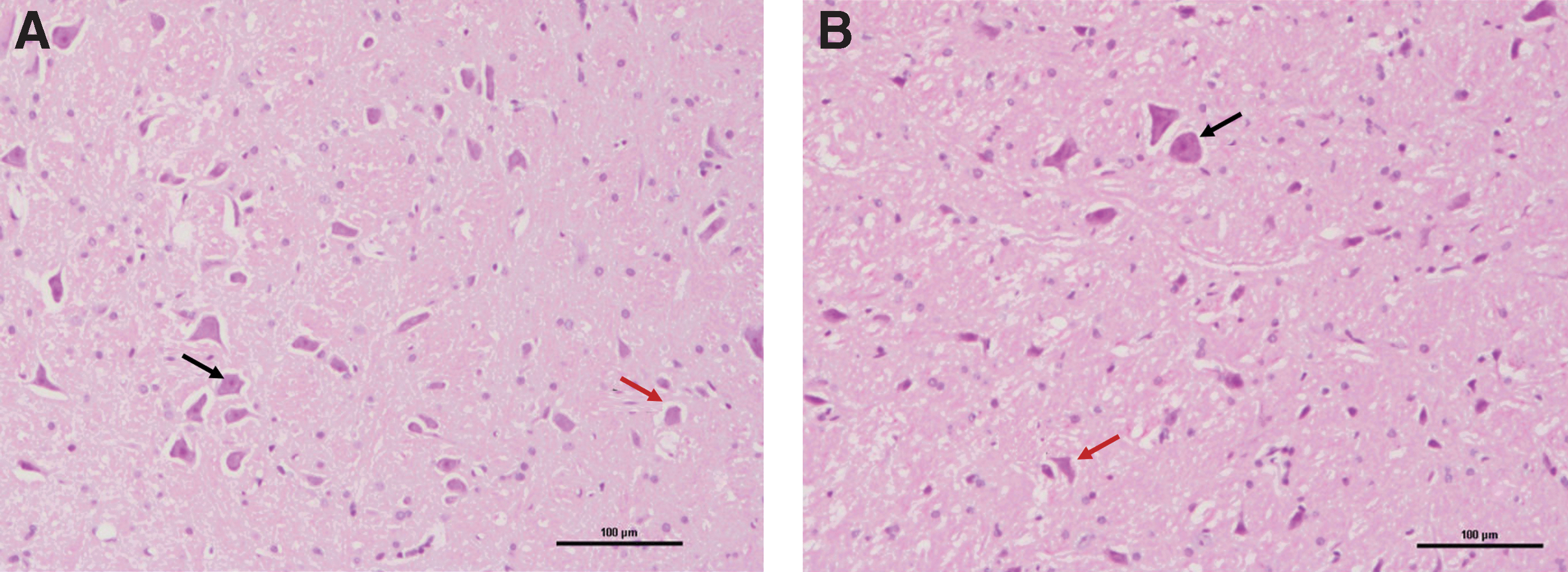
Hematoxylin-eosin staining results
On day 21 after FN injury, the degeneration of bilateral FMNs was significant, and the cytoplasm staining became lighter. Nucleoli were not clear, and some were fragmented (Fig. 3).
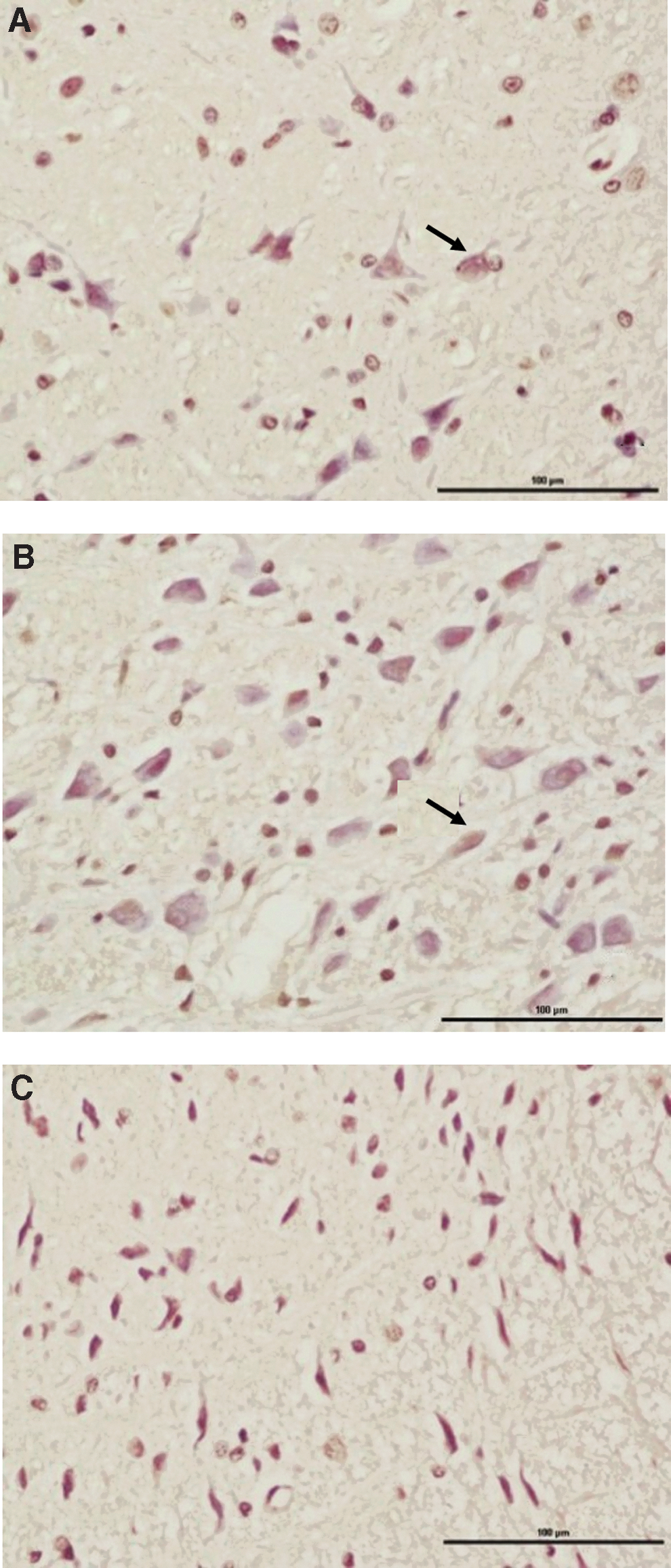
TUNEL staining results
The TUNEL staining results of FN nuclei in normal SD rats without surgical operation was used as the baseline control.
On day 21 after FN injury, the bilateral FN nuclei area contained some TUNEL-labeled neurons. Comprehensive determination was performed combined with assessment of the morphological characteristics of apoptotic cells. Unstained cells became smaller, and the cell membrane was intact. Vacuolation phenomena were observed. Apoptotic bodies were present at the late stage, and attached cells shrank, became round, and detached. Stained cells contained cell nuclei that exhibited slight brown. In addition, chromatin condensation and chromatin margination could be observed under a high-power microscope, and chromatin was split into pieces or had apoptotic bodies (Fig. 4).
Discussion
Effects on cytokines in BMSCs culture
The immune response model of the process of adaptive immunity in nerve injury repair has been elucidated by some studies before. Information on the release of unknown specific antigens during this process after nerve injury is lacking.
Results in this study demonstrated that SDBMSCs cultured in vitro alone or cocultured in vitro with FN both exhibited the ability to autonomously secrete five cytokines, including IL-6, HGF, PGE2, iNOS, and TGF-β1, and the secreted cytokine levels were higher. These findings are consistent with the results of previous studies. The high secretion levels of the aforementioned five cytokines by SDBMSCs might be associated with the inhibitory function on differentiation and production of the Th17 subset in CD4+ T cells reported in various studies. In addition, the cytokine levels in the FN + SDBMSCs group were increased compared with the SDBMSCs group, maybe suggesting that some specific antigens released after transection of FN had some interactions with SDBMSCs. These unknown interactions might affect the levels of some cytokines secreted by SDBMSCs. In addition, the cytokine levels between the Blank group and the FN group also exhibited certain differences; therefore, the difference between the SDBMSCs group and the FN+SDBMSCs group might be affected by cytokines secreted by in vitro cultured FNs. The specific mechanisms still require further study.
Effects on CD4+ T cell subsets
Current existing studies indicated that FN injury-induced immune responses include peripheral responses at the injury site and central responses that involve neuronal cell body and peripheral glia. The CD4+ T cell-mediated immune response after injury is a complicated multistep process [4].
The study of Serpe et al. [21] in immunodeficient mice that lacked different types of immune cells demonstrated that adaptive immune responses, especially CD4+ T cell-mediated immune responses [22 –28], played an important protective role in FMNs within several weeks of FN injury. Xin et al. [27] studied the developmental trend of all T cell subsets as time progressed after FN injury in mice and classified CD4+ T cells into two groups: the proinflammatory subsets (Th1 and Th17) that promoted FN injury and the anti-inflammatory subsets (Th2, Tr, and Foxp3+ Treg) that protected FN and prevent apoptosis. The numbers of cells in the mentioned two subsets increased after FN injury and peaked on days 7–9 after injury.
The Th17 subset percentage in bilateral cervical draining LNs in groups B and C that received surgical operation were increased compared with the normal baseline control group A, whereas the Th17 subset percentage in bilateral cervical draining LNs in the C group that received FN transection was increased compared with the B group without FN transection. These results indicated that different surgeries in these two groups induced different levels of inflammatory responses in the body of SD rats. Of these responses, Th17 played an important role as one of the proinflammatory subsets.
In addition, Ghannam et al. [17] and Qu et al. [18] demonstrated that BMSCs could inhibit the differentiation and production of Th17. We also demonstrated that the Th17 subset percentages in the right cervical draining LNs in groups B and C that received SDBMSC injection were reduced compared with the left cervical draining LNs without SDBMSC injection, suggesting that local injection of SDBMSDCs in the FN injury site could significantly reduce the Th17 subset percentage in cervical draining LNs at that side. In addition to the in vitro experimental results, BMSCs could also inhibit the Th17 subset in vivo. These findings were consistent with the existing in vitro study results [29]. Besides some studies have indicated that IL17 has such effect as we described, administration of blocking antibody to neutralize IL17 is also important and valuable to further clarify the function of IL-17 on FN injury. Because of the unavailability of reliable IL-17 blocking antibody in this research, IL-17 blocking experiment has been scheduled in the future study.
Effects on FMNs
After FN injury, the distal broken end of the axon degenerates and disappears. Macrophages phagocytose debris and Schwann cells proliferate. In addition to changes surrounding the peripheral nerve trunk, numerous studies demonstrated that FN nuclei in the CNS at the ipsilateral side will exhibit neuronal apoptosis, which peaks on day 15 after injury [30]. The influence of FN transection injury on FMNs is associated with the distance between the injury plane and FN nuclei. When the distance is closer, the injury of FMNs is more obvious.
Jones et al. [4] and Marzo et al. [31] studied the influences of the peripheral immune system on FMNs after FN injury. FMNs are basically divided into three groups in terms of numbers. Ten percent of neurons died immediately after FN injury. Fifty percent of neurons could survive for a longer period after FN injury. The numbers of the aforementioned two groups were essentially unaffected by immune responses. The other 40% of neurons depend on the involvement of the peripheral immune systems and/or neurotrophic factors. The distribution of these three groups of FMNs in six FN nuclei was not significantly different.
Various types of MSCs secrete a series of factors in the injury area to increase the survival rate of motor and sensory neurons. The in vivo study of Reid et al. [32] demonstrated that incorporation of adipose tissue-derived SCs into nerve conduits for repair of nerve defects could effectively reduce neuronal apoptosis after injury in peripheral nerves. The study of Wang et al. [33] and English et al. [34] on sciatic nerve injury demonstrated that BMSCs exhibited significant protective function on spinal cord anterior horn neurons.
Compared with FN nuclei in normal SD rats without surgical operation, we found that bilateral FN nuclei in SD rats with bilateral transection of the FN trunk exhibited FMN apoptosis after 21 days of surgery. The apoptosis indexes between the left and right side were significantly different. The apoptosis index of the right side was significantly reduced compared with that of the left side. These results suggested that local injection of SDBMSCs at the FN injury site significantly reduced FMN apoptosis in FN nuclei.
Summary
Based on our study results and previous results in the literature, we propose the following hypothesis. SDBMSCs injected in the local FN injury site secrete high levels of IL-6, TGF-β1, HGF, PGE2, and iNOS. IL-6 and TGF-β1 promote the differentiation and proliferation of Th17 subset cells. In addition, Th17 subset cells secrete IL-17 and promote PGE2 secretion by BMSCs. High PGE2 levels subsequently inhibit IL-17 secretion by Th17 subset cells. During this process, IL-17 expression is reduced. In flow cytometry, IL-17 is typically used to stain and label Th17 cells; therefore, the presentation involves the reduction of the percentage of Th17 cells.
Finally, we observed a more complete repair process after FN injury. Antigens are released after FN injury to induce immune responses. SDBMSCs secrete IL-6, HGF, PGE2, iNOS, and TGF-β1 cytokines, and these cytokines can regulate the balance among all subsets of CD4+ T cells. Collectively, these cytokines can protect FMNs and inhibit FMN apoptosis to further promote repair after FN injury.
In summary, BMSCs play a significantly active role in the repair process after FN injury. Specifically, BMSCs play important roles in CD4+ T cell-mediated adaptive immune responses after FN injury. The results of this study verified the conclusions of some previous studies. We performed in vivo experiments based on previous in vitro experiments and the obtained results were consistent. In addition, a hypothesized model regarding the mechanisms underlying the regulation of the Th17 subset percentage by SDBMSCs was proposed (Fig. 5). Further studies are required to elucidate the comprehensive immunomodulation mechanisms of SCs on FN injury.

A hypothesized model of the mechanisms underlying the regulation of the Th17 subset percentage by SDBMSCs.
Footnotes
Acknowledgments
This research was supported by National Natural Science Foundation of China (no. 81470698), Beijing Natural Science Foundation (no. 7172176), National Key Research and Development Plan of China (nos. 2016YFA0101000 and 2016YFA0101002), and Scientific Project of Young and Middle-aged Researcher of PUMCH (no. pumch-2013-007) to H.Y.
Author Disclosure Statement
No competing financial interests exist.