
Abstract
The aim of the present work was to determine proliferation capacity, immunophenotype and genome integrity of mesenchymal stromal cells (MSCs) from horse umbilical cord blood (UCB) at passage stage 5 and 10. Passage 4 cryopreserved UCB-MSCs from six unrelated donors were evaluated. Immunophenotypic analysis of UCB-MSC revealed a cell identity consistent with equine MSC phenotype by high expression of CD90, CD44, CD29, and very low expression of CD4, CD11a/18, CD73, and MHC class I and II antigens. Proliferative differences were noted among the UCB-MSC cultures. UCB-MSCs karyotype characteristics at passage 5 (eg, 2n = 64; XY, or XX) included 20% polyploidy and 62% aneuploidy. At passage 10, the proportion of polyploidy and aneuploidy was 21% and 82%, respectively, with the increase in aneuploidy being significant compared with passage 5. Furthermore, conventional GTG-banded karyotyping revealed several structural chromosome abnormalities at both passage 5 and 10. The clinical relevance of such chromosome instability is unknown, but determination of MSC cytogenetic status and monitoring of patient response to MSC therapies would help address this question.
Introduction
M
Equine MSCs are mainly characterized by their adherence to plastic culture-ware, fibroblastoid morphology, and their capability to differentiate into multiple lineages in vitro [7,14,17]. The morphological features, while important, are nonspecific and are also shared by other cell types, such as fibroblasts, which invalidates the use of these parameters alone for the characterization of MSCs. Considering basic criteria established by the International Society for Cellular Therapy (ISCT) to define human bone marrow-derived MSCs [18] and adipose tissue-derived MSCs [19], a panel of positive and negative expression of surface markers (immunophenotyping) have been proposed to identify human MSCs. Positive markers include CD29, CD44, CD90 [20,21], MHC-I [21], and CD105 [13], and negative markers include CD14 [20], CD34 [13,21], MHC-II [20,21], and CD45 [13].
Tessier et al. [15] and De Schauwer et al. [16] have shown equine UCB-MSCs to have high expression of CD29, CD44, and CD90 and have absent or low expression of, MHC-I, MHC-II, CD4, CD8, CD11a/18, and CD73. However, the identity and nomenclature surrounding MSCs remains controversial with some arguing that the term MSC should be abandoned, but this discussion is beyond the scope of the work presented here [22].
At present, several studies have been carried out to investigate the chromosome stability of human MSCs derived from different sources such as bone marrow, adipose tissue, and umbilical cord [15,23 –29]. However, few studies have performed cytogenetics analysis of equine MSCs [30 –32]. Chromosome stability depends on the presence or lack of variations in chromosome number and structure that is species specific. Numerical variations can appear as polyploidy and aneuploidy.
Polyploidy refers to a state in which cells contain more than two homologous (paired) sets of chromosomes deviating the normal diploid chromosome number (ie, 2n), such as tetraploidy (4n), while aneuploidy is a situation in which cells gain or lose a number of individual chromosomes from a homologous pair, either autosomes and/or the sex chromosomes, such as monosomy or trisomy. It is also possible to observe a variation in the chromosome structure of any chromosome in the karyotype, such as deletions, duplications, inversions, and translocations of chromosomal segments.
Changes in amount of genetic material during cell culture can lead to genome alterations and negatively affect cellular functions. However, there are considerable uncertainties concerning the significance of genomic instability during in vitro culture [33 –36]. Ex vivo malignant transformation of MSCs appears to be a rare occurrence, which has only been described in a few reports [25,26]. Genomic alterations such as aneuploidies (without associated malignant transformation) have been more commonly observed during cell culture [27,28,32,36 –38]. For example, cultured equine bone marrow-derived MSCs have been shown to have polyploidy and aneuploidy levels of 2%–7% at passage 3 and 4, while umbilical cord-derived MSCs displayed an average of 22% aneuploidy at passage 3 [30].
Although the clinical implications of these genomic alterations are not well understood, it has been argued that MSCs have low risk of tumorigenesis, as it has been shown that cytogenetic instability leads to a decrease or arrest in cell proliferation and concomitant elimination of affected cell populations [17,23,24,29,35,39,40]. Regardless, it is suggested to monitor genomic integrity of MSCs for use in basic research and clinical trials for cell-based therapy [23,29,41]. The aim of this study was to determine the immunophenotype, proliferative capacity, and genomic stability (ie, cytogenetic analysis) of cryopreserved equine UCB-MSC through passage 5 to 10.
Materials and Methods
Ethics statement
This study was specifically approved by the University of Guelph Animal Care Committee with regard to the procedures of collection of equine peripheral blood lymphocytes and equine UCB (animal use protocols 1756 and 1570). Additional research conducted using specimens of this kind does not require review by the Animal Care Committee (falls under CCAC Category of Invasiveness A). Collection of peripheral blood and cord blood were add-on procedures to the routine care of the horses. No animals were sacrificed during the study.
Informed consent was obtained in writing from the horse owners/agents before sampling. The broodmares on the foaling farms are housed in large foaling boxes. The foaling facilities are staffed 24/7 and mares are under constant video surveillance and carrying foaling alarms to allow for observed foaling and assisted delivery if needed. UCB was collected by the farm staff after receiving instruction by Dr. Koch. Instruction included video-review of cord blood collection. Cord blood was collected from an isolated segment of the umbilical cord after the umbilical cord had been clamped and detached from the foal.
Once investigators have an approved animal care protocol from the University of Guelph Animal Care Committee access to these research horses are granted. In this study peripheral blood was collected from three adult mixed-bred horses housed at the Equine Research Station, which is managed by the University of Guelph in partnership with the Ontario Ministry of Agriculture, Forestry and Rural Affairs. The research horses are housed in smaller groups with run-in sheds throughout the year. The horses are on pasture during the summer and during the winter they have access to large paddocks with gravel surface.
Peripheral blood was collected by Dr. Koch. Collection of peripheral blood was collected under mild sedation (Xylazine HCl, 0.35–0.40 mg/kg bwt IV; Bayer, Toronto, ON, Canada) from the jugular vein following which manual pressure was applied for several minutes to aid hemeostasis.
Equine umblical cord blood-derived MSC culture
A total of six unrelated cryopreserved equine UCB-MSC cell cultures were thawed and plated in polystyrene culture flasks and incubated at 38°C with 5% CO2 in a humidified incubator, see Supplementary Table S1 (Supplementary Data are available online at
Cells were harvested at 70% confluency using 0.04% trypsin/0.03% EDTA solution, replated for expansion at a density of 5,000 cells/cm2 and propagated in culture until passage 5 (P5) and passage 10 (P10). Metaphase spread preparations for conventional Giemsa staining and additional GTG-bands karyotyping was performed on all six cultures at P5 and P10. Cell growth was analyzed by direct manual cell counting using trypan blue, and the doubling time was determined. The population doubling time (PDT) was calculated by the formula: TD = t log10(2)/(log10 N − log10 N0), where N = cells harvested, N0 = cells seeded, and t = time of the culture (in days).
Immunophenotype flowcytometry
Equine UCB-MSC cultures and peripheral blood mononuclear cells (PB-MNC) were assessed for expression of MSC-associated cell surface markers (CD29, CD44, and CD90) and hematopoietic markers (CD11a/18, CD4, CD73, MHC-I, and MHC-II) by flowcytometry as previously described [15]. The following antibodies were used to evaluate the cells: APC anti-human CD29 (Clone: TS2116; Bio-Legend), Mouse anti-horse CD44:FITC (Clone: CVS18; Bio-Rad), Mouse anti-rat CD90 (Clone: OX-7; BD Pharmingen), Mouse anti-horse CD4:FITC (Clone: CVS4; Bio-Rad), Mouse anti-horse CD11a/18:FITC (Clone: CVS9; Bio-Rad), PE Mouse anti-human CD73 (Clone: AD2; BD Pharmingen), Mouse anti-horse MHC I:FITC (Clone:CVS22; Bio-Rad), and Mouse anti-horse MHC II:FITC (Clone:CVS20; Bio-Rad). Goat anti-mouse IgG1-FITC (Cat. 97239; Abcam, Toronto, ON, Canada) was used as a secondary antibody against unconjugated primary antibody (CD90). Antibodies and their positive control(s) are listed in Supplementary Table S2.
Cultured UCB-MSC cells were chemically detached with Trypsin-EDTA (Gibco™, Grand Island) and washed with flow buffer before antibody incubation. Incubations were at 4°C in the dark for 15 min, followed by a wash, and secondary antibody [goat anti-mouse IgG1-FITC (Abcam)] incubation carried out at 4°C for 15 min in the dark. All the cells stained with primary/secondary-conjugated antibodies were resuspended in 500 μL of flow buffer and passed through a BD Accuri C6 flow-cytometer (BD Biosciences, San Jose, CA). Negative control samples were cells incubated with only secondary antibody, unstained cells, and cells incubated with isotype-matched nonbinding primary antibody plus fluorescent secondary antibody. 7-AAD (Sigma) was used as a dead cell stain.
A minimum of 20,000 events were acquired for each antibody, run and analyzed on a BD Accuri™ C6 (BD Biosciences). Gates to identify UCB-MSC or PB-MNC populations were maintained consistent throughout all experiments.
Cytogenetic analysis
As previously described [6], the UCB-MSCs were seeded into T75-culture flasks and expanded to 60%–70% confluency before metaphase spread preparation. For cytogenetic analysis a cell synchronization protocol was applied [42]. In brief, ∼16 h before cell harvesting, dividing cells were synchronized with Methotrexate (10−7 M final concentration) in fresh culture media.
Cells were released from replication block by replacing the Methotrexate media with fresh culture media. Subsequently, equine UCB-MSCs were treated with 1.25 mM Bromodeoxyuridine (BrdU) for 5 h at 38°C to prevent excessive chromosome compaction and 10 mg/mL colcemid for 30 min to induce metaphase arrest. Cells were then dissociated, suspended in hypotonic solution (75 mM potassium chloride), and incubated at 38°C for 15 min. A few drops of ice-cold Carnoy's fixative (3:1 methanol:acetic acid) were added and mixed by gentle agitation. The cells were pelleted and resuspended in the fixative three times before dropping the cells onto fixative-soaked microscope slides to prepare metaphase spreads.
To determine euploid and aneuploid chromosome constitution, on average 80 and 50 metaphase cells were respectively examined under a Leica DM5500B microscope Retiga Exi II (Q-Imaging) digital camera, after subjecting the chromosome preparations to a standard giemsa staining and GTG-banding protocol. Representative karyotypes were assembled according to the international standards for the horse karyotype [43], using SmartType software (Digital Scientific UK Ltd.).
Statistical analysis
The results were statistically analyzed using Student's t-test for significance of differences between means among passages (P5 vs. P10). Differences at P ≤ 0.05 were considered significant. The tables indicate the mean and standard deviation.
Results
Immunophenotype flowcytometry and proliferation analysis
During in vitro expansion, all six UCB-MSC cultures maintained a fibroblastoid spindle-shaped morphology although slight differences in appearance were noted with increasing passage number (Fig. 1). Fast-growing and slow-growing MSC cultures were defined based on a comparison between their PDT and total median PDT calculated for cell cultures at both P5 (median = 38.4) and P10 (median = 64.8) separately. Three cell cultures whose PDT were above the median were grouped as low proliferative and three cultures with PDT value below the median were categorized as high proliferative cultures (Fig. 2A). The PDT of UCB-MSCs with high proliferative capacity and low proliferative capacity were respectively 31.2 ± 4.32 and 62.4 ± 18.5 h at passage 5 and 49.4 ± 1.2 and 113.6 ± 23.9 at passage 10 (Fig. 2A). No significant difference was observed between overall PDT of P10 cell cultures (Average PDT: 81.6 ± 38.4) and the overall PDT of P5 cell cultures (Average PDT: 46.56 ± 21.1) (Fig. 2B).
Morphology of adherent equine UCB-MSCs after 6 months of cryopreservation.
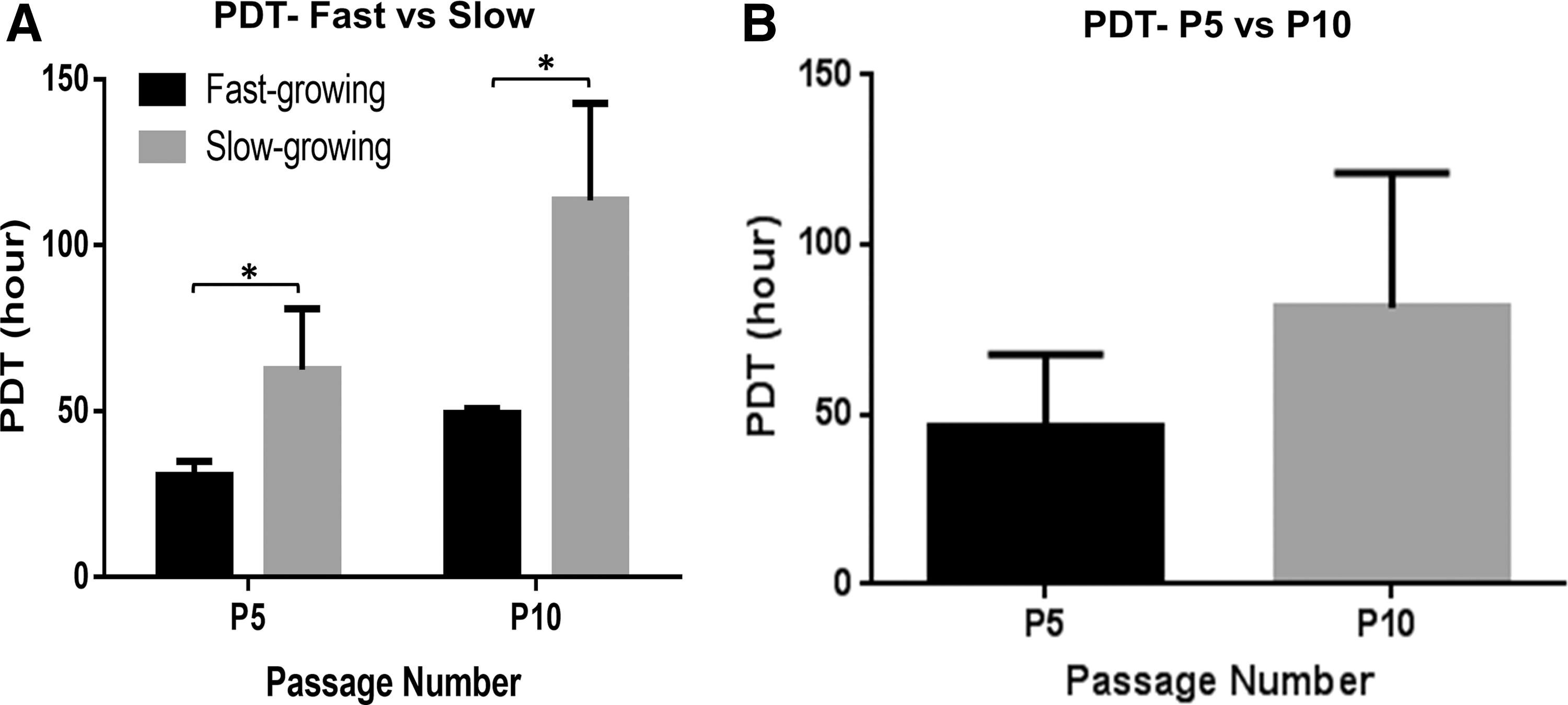
Population doubling time of equine UCB-MSCs.
Flow cytometry analysis showed UCB-MSC cultures with high antigen expression of CD90, CD44, CD29, and low antigen expression of CD11a/18, CD4, CD73, MHC I, and MHC II, which corresponded to the immunophenotypic criteria reported by previous equine MSC studies (Fig. 3). Statistical analysis revealed significant changes in expression of abovementioned surface antigen markers between the UCB-MSC population and PB-MNC fraction (P < 0.05; Supplementary Fig. S1).
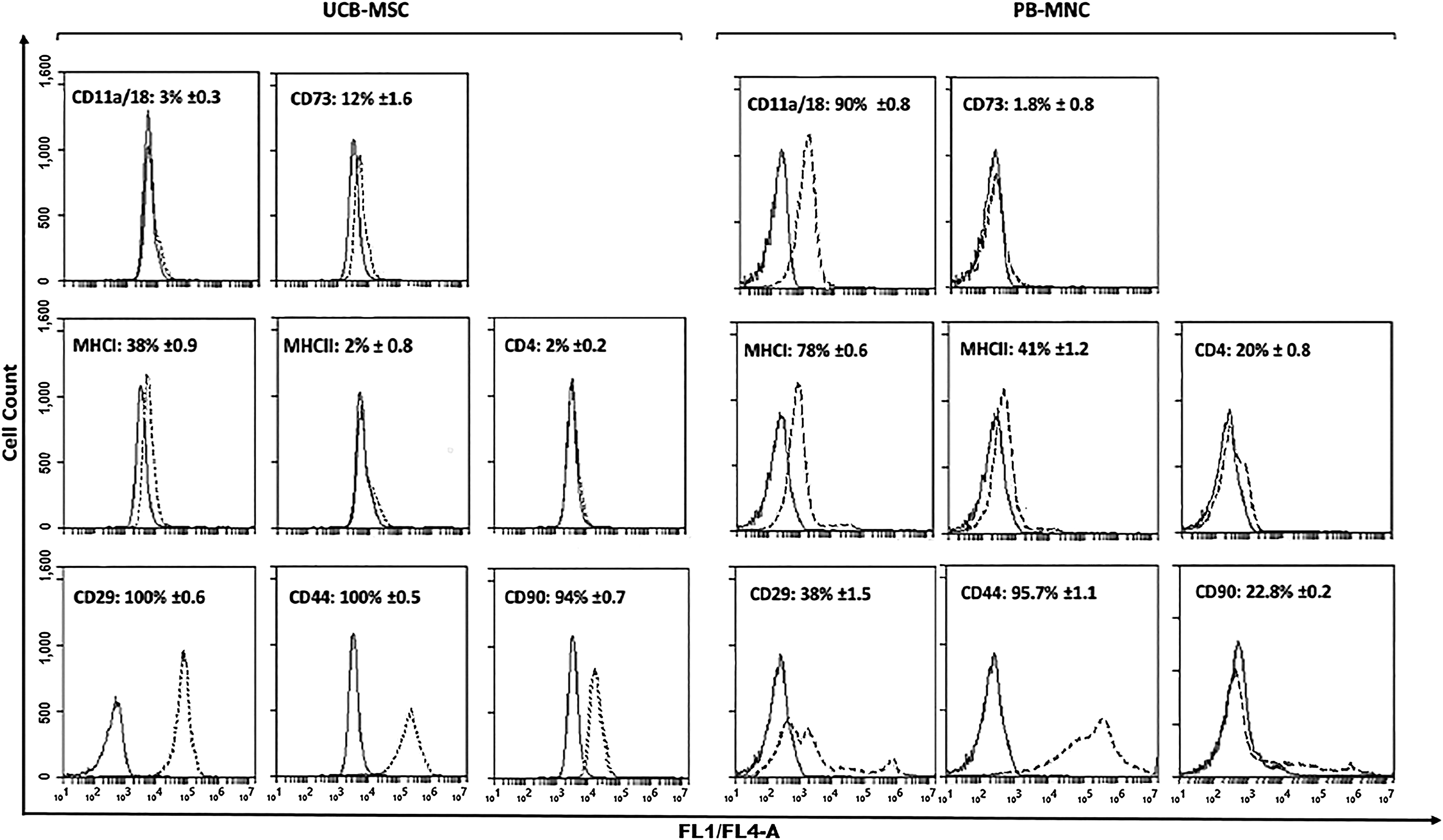
Immuno-phenotypic analysis of equine UCB-MSCs (n = 6) at passage five and equine PB-MNC (n = 3) as biological control. Consistent gates were applied for all cell analyses. Control sample fluorescence is indicated by solid line, fluorescence of samples incubated with specific antibody by dashed line. Numbers in each plot represent the mean proportion (± standard deviation) of cells expressing each antigen. UCB-MSC, umbilical cord blood derived mesenchymal stromal cell; PB-MNC, peripheral blood mononuclear cell.
Cytogenetic analysis
The cytogenetic analysis of equine UCB-MSCs showed that these cells had a large amount of numerical (euploid and aneuploid), and structural chromosome abnormalities. At P5 ∼ 105 metaphase MSCs of each cell cultures were screened for euploidy and on average 20% of them revealed polyploidy chromosome constitution (Fig. 4). Similarly, ∼80 metaphase cells were screened for aneuploidy, with 60% of MSC cultures displaying aberrant karyotype appearing in different forms of monosomy, trisomy, and nullisomy. Monosomy, trisomy, and nullisomy of chromosome 13 were detected at higher frequency; furthermore, nullisomy occurred also for sex chromosomes pairs. Structural abnormalities of chromosomes were also observed in the MSC cultures at passage 5 (Table 1, Fig. 5, and Supplementary Fig. S2).
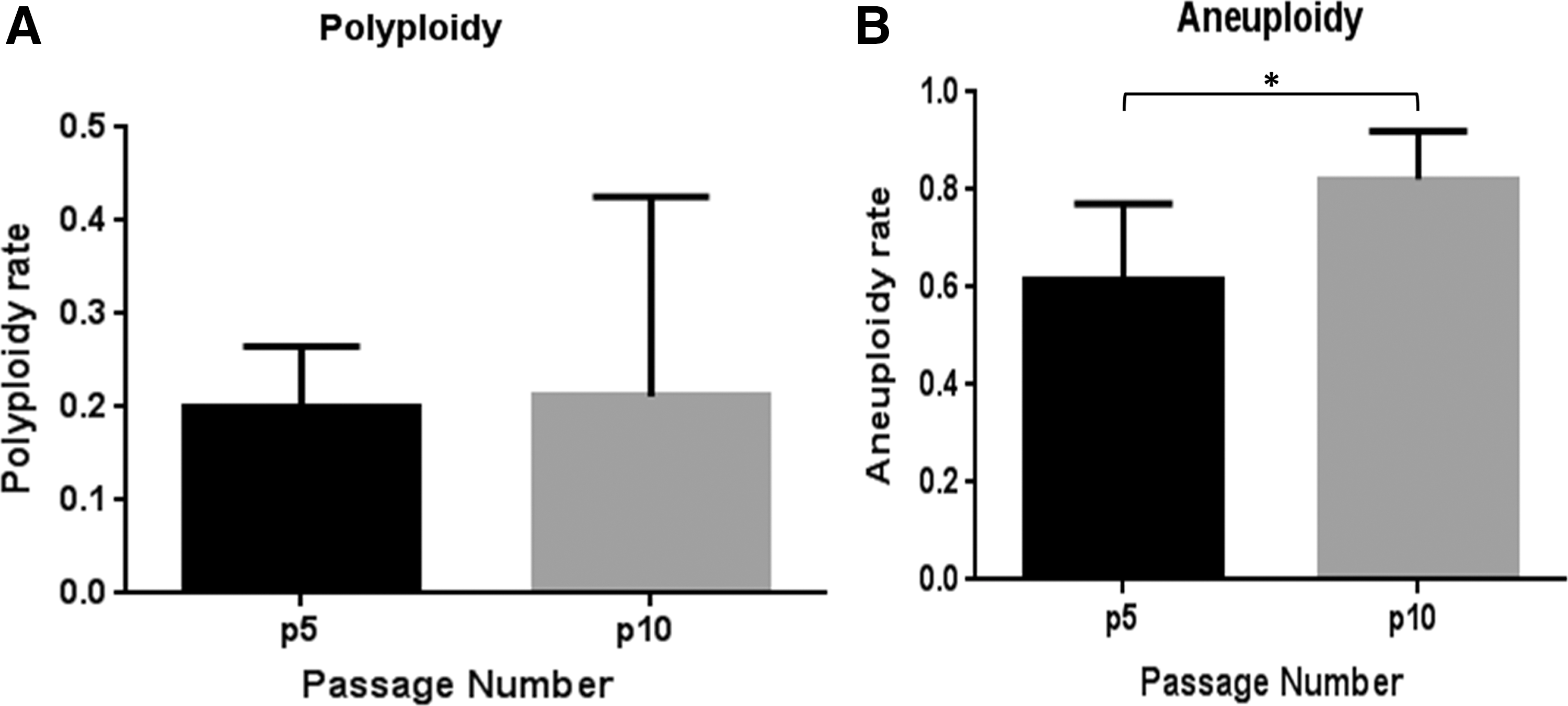
Percentage of chromosomal abnormalities in equine UCB-MSCs at P5 and P10.
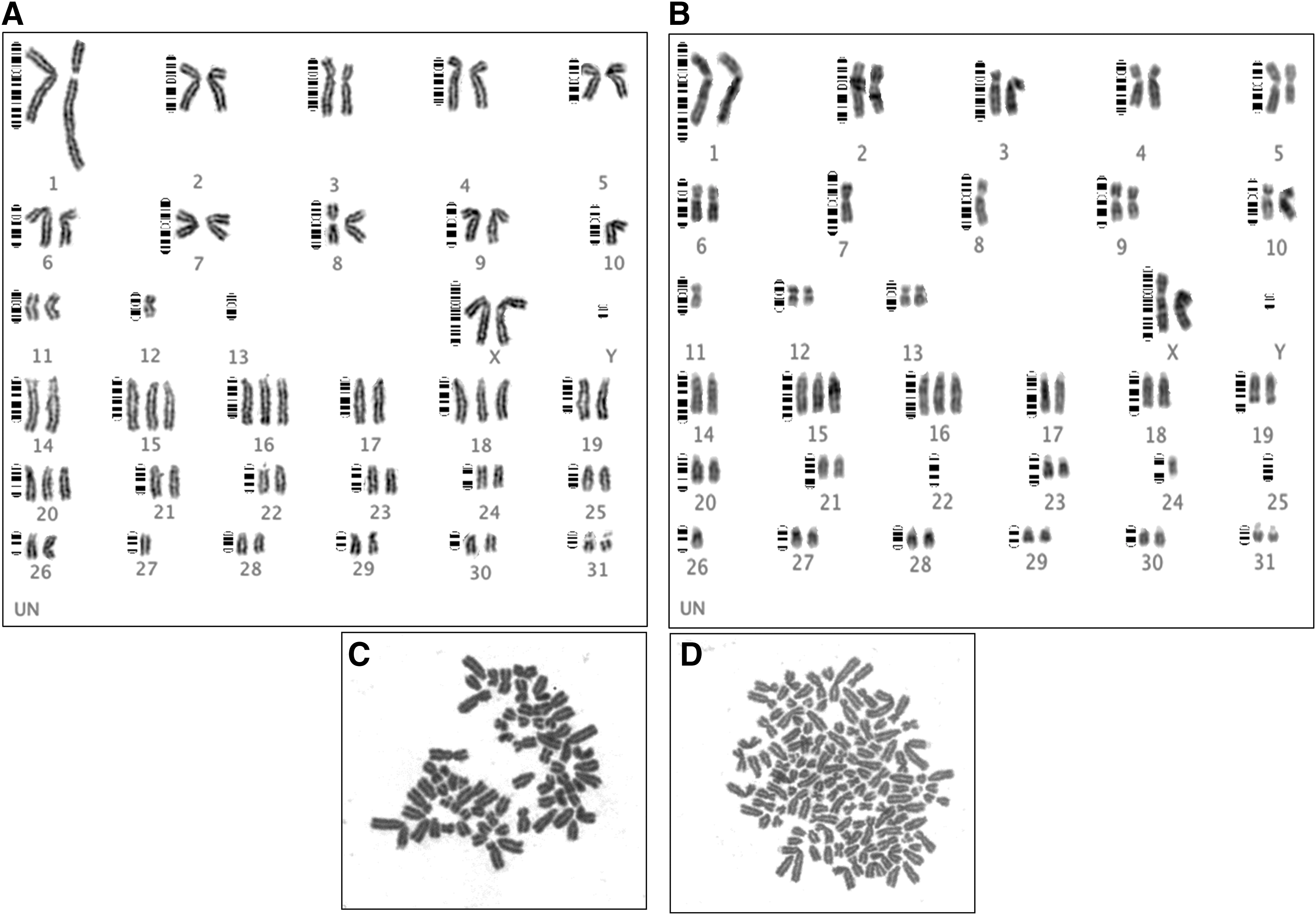
Cytogenetic analysis of equine UCB-MSCs:
Cytogenetic Profile of Equine UCB-MSCs at P5 and P10
UCB-MSC,
On average 80 metaphase MSCs per cell culture were subsequently screened at P10 for euploidy in which 21% revealed polyploidy (Fig. 4). Screening ∼42 metaphase cells for aneuploidy, 82% of MSC cells displayed such chromosomal abnormality in the forms of monosomy, trisomy, and nullisomy.
Aneuploidy of chromosomes 13, 15, 16, and 19 were respectively observed at 50%, 66%, 50%, and 50% of MSC cell populations (Table 1). On average, at P5 MSCs displayed 20% and 62% of metaphase polyploidy and aneuploidy, respectively, while at passage 10 the rates of polyploidy and aneuploidy were 21% and 82%. Although there was no significant difference between polyploid MSCs at passage 5 and 10, aneuploidy of passage 10 MSCs was significantly higher than aneuploidy of passage 5 MSCs (Table 1 and Fig. 4). Screening of UCB-MSC cultures with high or low proliferation capacity at P5 and P10 showed no significant difference (P < 0.05) with regard to polyploidy and aneuploidy (Supplementary Fig. S3).
Structural chromosome abnormalities were observed in karyotypes from cell of MSC cultures studied at P10 (Table 1, Fig. 5, and Supplementary Fig. S2). Individual GTG-banded karyotypes demonstrated obvious derivative chromosomes as evidences that some structural rearrangements have occurred, although it was not feasible to define the chromosomal segment involved in the exchanges the presence of derivative chromosome was evident, since chromosome pairing following the standard guidelines for equine metaphase karyotyping lead to these observations.
The occurrence of numerical and/or structural chromosome aberrations was not related to the sex of the MSC donor as those occurred in both male and female MSC cell cultures that were cultured for P5 and P10 passages. Aneuploidies appeared in the forms of monosomy, trisomy, and nullisomy. Trisomy of chromosomes 13, 15, 16, and 19 were detected at higher frequency and nullisomy occurred in both autosomes and sex chromosomes (Table 1).
Discussion
In this study, we assessed the cell identity, proliferation potency, and cytogenetic status of cryopreserved UCB-MSCs isolated and expanded in vitro. Although there is currently no consensus on cell characteristics for classification of equine MSCs, our UCB-MSCs showed some basic properties defined for the human MSCs; they were plastic-adherent, displayed a fibroblast-like morphology, displayed high expression of CD29, CD44, and CD90, and low expression of CD11a/18, CD4, CD73, MHC I, and MHC II.
Like other cytogenetic reports on stem cells [44,45], conventional cytogenetic analysis of UCB-MSCs used for cell therapy identified a high rate of chromosomal alterations during culture in vitro. High prevalence of aneuploidy of UCB-MSC at P5 and P10 was observed. At P5, aneuploidy was detected in 62% of the cells, but this increased to 82% at P10. Our finding of aneuploidies in UCB-MSC cells is consistent with a previous study in which equine umbilical cord MSCs were observed with 22% aneuploidy at P3 of in vitro cultivation [30]. In contrast, equine bone marrow-derived MSC at P3 and P4 passages of in vitro cultivation were found with an aneuploidy level of 2%–7%, which was similar to the level of karyotype variability in equine peripheral blood lymphocytes [30].
We further found UCB-MSC cells with trisomies for chromosomes 13, 15, 16, and 19 at high frequency, suggesting that equine UCB-MSCs in the current culture and handling system are prone to acquire a unique set of chromosomal aberrations. This phenomenon has been reported for some human multipotent stem cell lines [35]. Trisomy for human chromosomes 13, 18, and 21 has been reported by different studies [46 –49].
Our observed differences over time could be due to acquired or accumulated changes during culture expansion. Culture condition such as oxygen tension, presence of antioxidants, length of culture period, and number of freeze/thaw cycles may affect the chromosome stability of cells [23,30,35,50,51]. There may also be shortening of telomere length during culture expansion that may further contribute to chromosome instability [50].
Alternatively, the changes could be due to clonal selection in an adaptive ecosystem. In this scenario, clonal populations with chromosomal abnormalities might have gained an advantage allowing them to eventually become the dominant population within a given cell culture in vitro [52]. Clonal selection of trisomies 8 and 11, promoting cell proliferation, has been described in different mouse embryonic stem cell lines [53 –55]. It was also reported that chromosomal aberrations, accumulated in human mesenchymal stem cell cultures, took over the culture rapidly within seven passages. It suggests that multipotent stem cells may obtain beneficial chromosomal aberrations enabling them to dominate the cell populations in in vitro cell cultures [43].
We found increased PDT over time, which is a common observation in primary cultures [17,24]. Whether the increase in PDT and abnormal karyotype rates are related is still undetermined. It has been reported that cytogenetic disorders were accompanied by senescence initiating in a significant proportion of cells with no morphological alterations. However, these cells were not able to propagate in culture, representing for instance that high levels of aneuploidy in human MSCs may be associated with a low proliferative capacity [51]. Andraszek et al. [44] made similar observations that the presence of polyploidy leads to termination of cell division, promotion of cellular senescence, and apoptosis. Our findings are consistent with the observations of those two prior reports [44,51].
In contrast to those studies reporting a reduced proliferation as a result of the presence of aneuploidy during culture, Gaztelumendi and Nogués [52] indicated that mouse embryonic stem cells carrying chromosome abnormality (double trisomy of chromosome 8 and 11) promoted cell proliferation, leading to a faster clonal selection [52]. Occurrence of genomic diversification during cultivation may also lead to abnormal functioning of the genome including oncogenic transformation such as inactivation of tumor suppressor genes and upregulation of oncogenes [45].
The relevance of these chromosomal abnormalities on cell function relating to properties of MSCs such as tri-lineage differentiation and lymphocyte modulation in vitro is undetermined. The same UCB-MSC cell cultures evaluated in this study has been used in other studies within our lab evaluating tri-lineage and lymphocyte suppression properties (unreported). In these studies, the UCB-MSC cultures at higher passages showed suppressed chondrogenic and lymphocytic potency as expected (unreported).
Safety of MSC therapy is a major focus of basic research and is under clinical evaluation in a wide range of medical fields. Initial clinical trials of human MSCs have not reported the occurrence of transformation and tumorigenesis (ie, malignant transformation) [24,25,30,35]. Roemeling-van Rhijn et al. [23] indicated that development of other genomic abnormalities is not associated with the occurrence of aneuploidy in human MSCs [23]. Murine and macaque MSCs appear susceptible to transformation and capable of sarcoma formation while human MSCs do not seem to share these features [24,56]. This may be a reflection of fundamental difference among MSCs of different species or it could be due to species independent differences among the investigated MSC cultures.
Some form of autophagy, a catabolic process by which cells arrest intracellular lipids, proteins, and organelles and deliver them to lysosomal compartments where they are degraded, may reduce chromosomal instability in MSC cultures [57]. MSC populations with low percentage of chromosomal abnormalities have been noted to remain stable during several passages until senescence occurred upon which the cells lost the autoregulatory capacity to degrade oncogenic protein substrates. This subsequently resulted in higher frequencies of chromosomal abnormalities [23]. It remains to be determined if such autophagy regulation occurs in UCB-MSC cell cultures and if so, at what time point it would be exerted and chromosome instability would start to accumulate.
In conclusion, plastic adherent cells derived from UCB express markers consistent with a MSC phenotype and do not express hematopoietic markers. Several chromosomal abnormalities were identified in the examined UCB-MSCs. The influence of culture conditions and multiple freeze-thaw cycles needs to be investigated before generalized conclusions can be made regarding UCB-MSC chromosomal stability during propagation and handling in vitro. It appears advisable to further investigate the best conditions to expand equine MSCs to minimize genomic abnormalities in vitro.
The clinical relevance of such chromosome instability is unknown, but determination of MSC cytogenetic status and monitoring of patient response to MSC therapies would help address this question.
Footnotes
Acknowledgments
This project was supported by the Equine Guelph and Partners (T.G.K.), NSERC-DG (T.G.K.), CFI-LOF (T.G.K.), MRI-ERA (T.G.K.), and the Dean's Office, Ontario Veterinary College (A.H.A, PhD scholarship). The authors sincerely thank Dr. Sarah Lepage, University of Guelph for her diligent proofreading and constructive criticism of the article.
Author Disclosure Statement
A.H.A., R.B., D.A.F.V., and W.A.K. declare no competing financial interests. T.G.K. acts in a volunteer capacity as nonexecutive Director, Scientific Affairs (ex officio) of eQcell therapies, Inc., Aurora, ON, Canada, a company for which Dr. Koch's research laboratory provides equine stem cell isolation and storage services. Dr. Koch holds a minor noncontrolling share in eQcell therapies, Inc.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.