
Abstract
Severe burns are some of the most challenging problems in clinics and still lack ideal modalities. Mesenchymal stem cells (MSCs) incorporated with biomaterial coverage of burn wounds may offer a viable solution. In this report, we seeded MSCs to a biodegradable hybrid hydrogel, namely ACgel, that was synthesized from unsaturated arginine-based poly(ester amide) (UArg-PEA) and chitosan derivative. MSC adhered to ACgels. ACgels maintained a high viability of MSCs in culture for 6 days. MSC seeded to ACgels presented well in third-degree burn wounds of mice at 8 days postburn (dpb) after the necrotic full-thickness skin of burn wounds was debrided and filled and covered by MSC-carrying ACgels. MSC-seeded ACgels promoted the closure, reepithelialization, granulation tissue formation, and vascularization of the burn wounds. ACgels alone can also promote vascularization but less effectively compared with MSC-seeded ACgels. The actions of MSC-seeded ACgels or ACgels alone involve the induction of reparative, anti-inflammatory interleukin-10, and M2-like macrophages, as well as the reduction of inflammatory cytokine TNFα and M1-like macrophages at the late inflammatory phase of burn wound healing, which provided the mechanistic insights associated with inflammation and macrophages in burn wounds. For the studied regimens of these treatments, no toxicity was identified to MSCs or mice. Our results indicate that MSC-seeded ACgels have potential use as a novel adjuvant therapy for severe burns to complement commonly used skin grafting and, thus, minimize the downsides of grafting.
Introduction
S
Many studies, including our previous reports, have successfully used mesenchymal stem cells (MSCs) to promote injury repair in preclinical models and in preliminary clinical case reports [6 –13]. MSCs accelerate closing of wounds, improve neovascularization, and reduce generation of scarring. The mechanism for these benefits are not yet understood, but MSC production of prohealing paracrines has been mooted [14,15]. The lack of engraftment of transplanted MSC into the wound has limited our understanding of the role of MSC lineage differentiation [8], and is a hurdle to be surmounted for therapeutic intervention [16]. Long-term engraftment rates of less than 3% have been reported in injury models of the pancreas [17], heart [18], kidney [19], and liver [20]. Local administration to wound sites shows similar levels of engraftment [8]. Therefore, improved delivery methods may repair MSC injury.
Hydrogels consist of hydrophilic three-dimensional network (3D) that can hold large amount of water or biological fluid, structurally mimic the natural extracellular matrix, and thus are used as 3D scaffolds for MSCs to promote wound healing [21 –27]. Hydrogels fabricated from acellular dermal matrices and other nature tissue components such as collagens and fibrins were successfully used to deliver stem cells to injury sites and showed the promise in promoting repair and regeneration [28 –30]. The problems associated with these tissue-derived natural biomaterials include cost, reproducibility, availability, and the potential risk of disease transmission; and hence, hydrogels fabricated from synthetic biomaterials that could mimic the beneficial structural, mechanical, and chemical properties of skin without these problems. The select hydrogels prepared from synthetic amino acid-based poly(ester amide) polymers (AA-PEAs) and their hybrids with chitosan (Fig. 1) or other clinically used copolymers possess not only these desirable properties but also their unique biological properties that could allow them to be good candidates as the scaffold and delivery carrier of MSCs to heal burn wounds [31].

The chemical structure and specific physical and mechanical properties of hydrogels prepared from UArg-PEA and GMA-chitosan.
AA-PEAs are a relatively new generation of synthetic biodegradable biomaterials that exhibit both protein and nonprotein characteristics due to the presence of amino acid and nonamino acid building blocks (dicarboxylic acids and diols, nontoxic, physiologically existed in tissue) (Fig. 1A) [32,33]. Based on the wide variety of these three building blocks, AA-PEAs have shown enormous design capability and versatility and can be tailored for specific clinical applications. In addition to a wide range of physical, chemical, and morphological properties, the most unique aspect of AA-PEAs is their biological property, particularly muted foreign-body-induced inflammatory response, biocompatibility, and capability to promote the proliferation and/or survival of 11 different types of animal and human cells [31 –40]. Previously published studies have also demonstrated the unique advantages of AA-PEA, which includes reproducible synthetic routes, enzymatic biodegradability via surface erosion mode [32,33]. By selecting different types of amino acids, diols, and dicarboxylic acids and by adjusting the number of hydrocarbons between ester and amide groups, AA-PEA biomaterials are tunable in type of charge (cationic, anionic, or neutral) and density, hydrophobicity, or hydrophilicity, as well as the reactive sites for further chemical functionalization. AA-PEAs have also been engineered into a variety of physical forms ranging from nanofibrous membranes, 3D porous hydrogels, coating for suture, and cardiovascular stent to drug nanodelivery vehicles for multiple biomedical applications [32,33]. A new drug-eluting stent coated with a leucine-based PEA (Slender IDS®) was very recently approved by the European regulatory agency for commercial use in humans. The current research on AA-PEAs has recently been reviewed [32,33].
Among the amino acids for AA-PEA synthesis, arginine (Arg) is the most unique for two reasons: capable of retaining cationic charge over a wide range of pH due to its strong basic guanidine group with an isoelectric point of 10.96 and pKa above 12.5 as well as its biological property. The former character makes Arg suitable to form electrostatic nanocomplex with other negatively charged biomaterials or payloads [41 –44]. Because of Arg's role in wound healing, free Arg supplement to wound patients has been suggested and reported. For example, free Arg therapeutic treatment was applied to diabetic ulcer wounds, but the outcomes were not greatly improved [45 –51]. The Arg-based PEAs themselves or their hybrid hydrogels with either natural polysaccharide biomaterials such as hyaluronic acid and chitosan [31,34 –37] or synthetic biomaterials such as Pluronic diacrylate [38,52] have been reported as the delivery vehicles of bioactive agents such as bovine serum albumin, ovalbumin, hydralazine, insulin, and interleukin-12.
In this report, a biodegradable hybrid hydrogel, namely ACgels, synthesized from unsaturated
Materials and Methods
Preparation of hybrid hydrogels from AA-PEAs and modified chitosan
The hybrid hydrogels, ACgels, were synthesized from two polymer precursors: UArg-PEA and GMA-chitosan according to the published study [31]. Both UArg-PEA and GMA-chitosan precursors have unsaturated double bonds for the purpose of hydrogel formation via intermolecular crosslinking; the double bonds in the UArg-PEA are located in the backbone of the polymer, while the double bonds in the GMA-chitosan are located as the pendant group (Fig. 1A). The number- and weight-average molecular weights of UArg-PEA synthesized by this method are 12.93 and 14.01 kg/mol, respectively. The GMA-chitosan was synthesized by the procedure described in a previous study [58]. The degree of substitution, namely the percentage of methacrylate groups grafted onto the chitosan of GMA-chitosan repeat units, was 37 determined by 1H NMR spectroscopy. At a feed ratio of 20–80, both UArg-PEA and GMA-chitosan precursors were dissolved in deionized water and photocrosslinked in the presence of a photoinitiator Irgacure 2959 (Sigma-Aldrich, St. Louis, MO). The solution was then exposed to UV (365 nm, 100 watts, 30 min) to crosslink the dissolved two polymer components. The resulting ACgels were then soaked in and rinsed with deionized water for 6 h to remove unreacted residues.
Figure 1 shows a representative chemical structure and physical and mechanic properties of ACgels. The ACgel was cut into a cylinder shape (12 × 6 mm, diameter × length) and soaked in 70% ethanol/water for 10 min for sterilization in cell culture hood. The cylinders were cleaned by soaking in 10 mL deionized water for 6 h, then incubated in 10 mL sterile PBS (0.05 M) for 2 h. The cylinders were further cut into disks (5 mm diameter × 2 mm depth) for the following experiments. The key chemicals used in the preparation of ACgels are as follows. L-Arginine (L-Arg), fumaryl chloride, ethylene glycol, and 1,4-butanediol were purchased from Alfa Aesar (Ward Hill, MA). Chitosan (75–85% deacetylated) of molecular weight (MW) 50,000–190,000 Daltons was purchased from Sigma-Aldrich (St. Louis, MO). Glycidyl methacrylate (GMA, 97%) was from VWR Scientific (West Chester, PA). Irgacure 2959 was donated by Ciba Specialty Chemicals Co. More details about the synthesis and characterization of UArg-PEA and GMA-chitosan were presented in the Supplementary Data (Supplementary Data are available online at
Preparation of mouse MSCs
MSCs were isolated from the bone marrow of C57BL/6 J mice (8 weeks old, female, Jackson Laboratory), as described in our earlier publications; more than 95% of isolated MSCs were positive for Sca-1 and CD29 based on flow cytometric analysis [6,7,59]. These MSCs possessed adipogenic and chondrogenic differentiation ability and were cultured in DMEM containing 10% mesenchymal supplement (Stem Cell Technologies, Vancouver, BC, Canada). Cells in passages 3 through 5 were used for experiments.
Seeding of MSCs to ACgel scaffolds and assessment of MSC survival
Each ACgel disk was blotted with a sponge, and then took up a drop (30 μL) of MSC medium with or without the mouse MSCs (1 × 105) by the capillary force when dipping into the drop on hydrophobic waxy Parafilm (Fig. 2). The technique for seeding MSCs to a hydrogel scaffold was developed by Gurtner's group previously [60]. The MSCs seeded into ACgel scaffolds were cultured (37°C, 5% CO2) for the assessment of MSC viability in ACgels (Fig. 2). At day 1, 3, or 6 of the culture, we incubated the MSC-seeded ACgels for 45 min at 37°C with a live/dead cell assay kit consisting of calcein-AM and ethidium-homodimer (ThermoFisher, Waltham, MA). The living MSCs were stained green by calcein-AM, and the dead MSCs were stained red by ethidium-homodimer. After washed with PBS buffer, images of MSCs seeded into ACgels were captured by a confocal microscope (Zeiss LSM510, Zeiss AG, Oberkochen, Germany). The living and dead MSCs were counted. The MSC viability (% of living cells) was then measured. MSCs under conventional 2D culture were used for comparison.
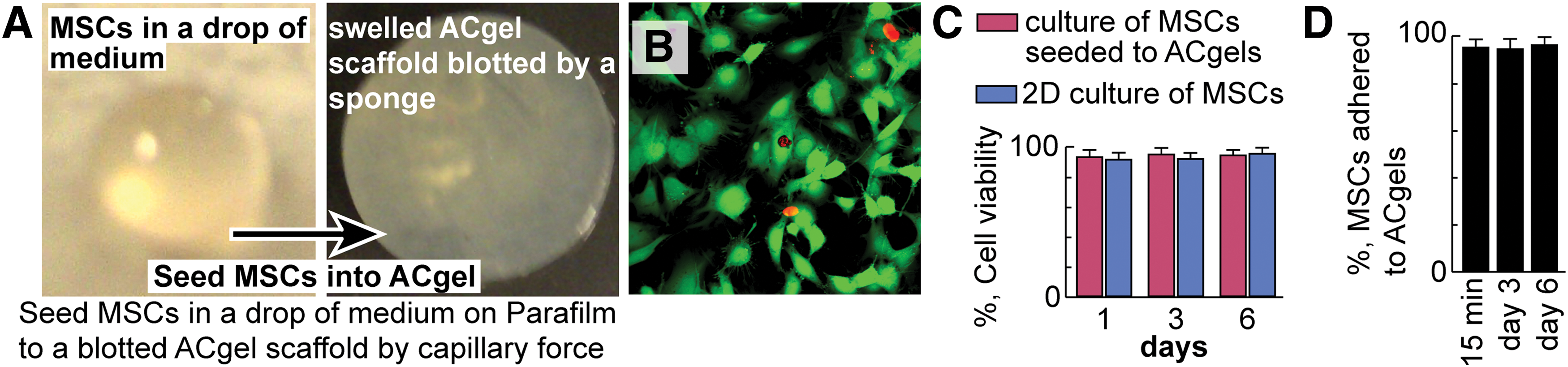
MSCs maintained a high viability after seeded to ACgel scaffolds and cultured.
Because we aimed to study MSCs seeded ACgels in the treatment of burn wounds, we determined the adhesion of MSCs to ACgels after MSCs were seeded to ACgels and cultured for 15 min, 3 days, and 6 days. The MSC-seeded ACgels were washed with medium three times, the media collected from the washing was centrifuged at 2,000 g for 15 min and cells in the pallets were also analyzed with the same live/dead cell viability assay kit [60]. The percentage of MSCs (live and dead) in an ACgel over the total MSCs in the ACgel, and the medium collected from washing is considered as the MSCs adhered to the ACgel.
Mouse model of 3° burn wounds and treatment with MSC-seeded ACgels
We generated third-degree burn wounds and excised burn-created necrotic skin on C57BL/6 J mice (female, 9 weeks old, Jackson Laboratory) following an established burn model [1,53 –57]. The Institutional Animal Care and Use Committee of Louisiana State University Health Sciences Center, New Orleans, approved the protocol. Hair was removed from the dorsal skin of mice by Veet depilating cream. Skin surface was sterilized with 70% ethanol then dried. The 12 mm diameter full-thickness burn wounds were created symmetrically across the midline by loading a 100°C alloy-aluminum-rod (custom-made, 220 g) vertically to each dorsal skin site of mice for 4 s. The rod was preheated to the temperature in boiling water for 5 min and blotted by touching a paper towel for ∼0.2 s immediately before burning the skin. Fluid resuscitation was performed by intraperitoneal injection of 1.7 mL NaCl saline within 1 h postburn and 0.5 mL at 12 h to a ∼24 g mouse.
We excised 5 mm diameter circle of coagulated or necrotic full-thickness skin at the center of each burn wound at 48 h or 2 days postburn (dpb) and filled each excision-generated space with an ACgel disk loaded with C57BL/6 J mouse MSCs (1 × 105) or medium (30 μL) as above and then covered the burn-injured area with an occlusive dressing. The excision mimics the clinical practice of debriding severely burned skin from the burn area and covering wound area with dressings after the debridement. The same type of wounds applied with the media lacking ACgels or MSCs were used as the sham control. The above procedures in vivo were conducted with mice anesthetized by i.p. injection of ketamine–xylazine (80 mg/kg). We also used the MSCs labeled with Dil (Vybrant, red fluorescent, ThermoFisher) to track the implanted MSCs in wounds. MSCs were labeled with Dil by incubation with Dil solution in serum-free culture medium (20 min, 37°C), then washed three times for the removal of untaken Dil according to the manufacturer's instruction.
Postoperative care, wound-closure measurement, and sampling
All wounded areas were covered with Tegaderm waterproof dressings to maintain ACgels in the excision-created skinless areas, to prevent water loss, and to protect the wound areas against unwanted potential damage caused by mouse activities. Mouse behavior was monitored, including foraging, grooming, drinking, and eating. We checked burn wounds daily to ensure that ACgels and dressings stayed in appropriate places.
Wound healing was analyzed by a protocol similar to previously published methods [6,61 –65]. In brief, each wound was photographed with a digital camera (Canon) alongside a thin paper ruler used as a graded scale bar. The wound areas in the photos were calculated using NIH ImageJ software. The wound closure was measured as the percentage of closed wound area relative to the initial burn wound area. After the mice were sacrificed, the burn wounds with 3 mm normal skin rims were excised, fixed in 4% paraformaldehyde (4 h, ∼23°C), soaked in 30% sucrose overnight at 4°C, embedded into OCT (optimum cutting temperature matrix, Tissue-Tec, Torrance, CA), and then stored at −80°C for histological analysis within a month as we did previously [66,67]; in addition, the livers and spleens were collected, weighed, and measured for sizes for the gross assessment of toxicity.
Hematoxylin and eosin staining and scanning microscope
The analysis of reepithelialization and granulation tissue formation of burn wounds was as follows. Serial sagittal cryosections (10 μm thickness) were prepared through the center region of the widest wound-bed on each burn wound specimen using a Leica CM3050 s cryostat microtome and transferred to Superfrost slides. The two sections of each wound representing the widest wound bed or the center of the wound were chosen for the following analysis. The selected sections were stained with hematoxylin and eosin (H&E) as we have done before [6,61,66]. The cryosections were treated sequentially as follows: incubated in PBS (5 min, ∼23°C) to remove the OCT; dehydrated and stained with hematoxylin (2 min, ∼23°C); rinsed with tap water to remove extra hematoxylin; quickly dipped through 1% HCl/alcohol then 0.3% ammonia water to enhance hematoxylin staining and to remove extra hematoxylin; and incubated at ∼23°C in 80% alcohol (10 s) and then in 0.5% eosin (2 min). The sections dipped in 95% alcohol (six times), 100% alcohol (three times), and then xylene (three times) to clean extra eosin, submerged in Crystal mounting media (ThermoFisher), covered with coverslips and dried (∼23°C), and then imaged using an Olympus scanning microscope. The multiple images acquired from the scanning of each burn wound section were integrated to form an image of the whole section and analyzed via Olympus OlyVIA software, and further analyzed via Photoshop software for the measurements of reepithelization and granulation tissue area (mm2). Reepithelialization (%) was calculated as [(distance of reepithelialized neoepithelium from leading edges to wound margins)/(distance of wound bed) × 100%] as previously described. Wound granulation tissue exhibits tight granular morphology between the epidermal layer and the subcutaneous layer and contains new connective tissue and blood vessel capillaries growing from the base of the wound [6,67].
Immunohistochemistry and confocal microscopic analysis
Skin wound cryosections (10 μm thickness) were incubated sequentially in methanol/acetone (1/1, 20 min, −20°C); citrate buffer (20 min, 90–100°C); and blocking buffer [10% BSA and 0.3% trition-X-100; 1 h, ∼23°C]. To measure blood vessels, we incubated the burn wound cryosections with rat anti-mouse CD31 antibody (BD Biosciences, San Jose, CA), followed by Cy3-labeled Donkey anti-rat IgG. Thereby, blood vessels in wound sections were stained with red fluorescence. The vascularity in wounds was presented as CD31+ area (pixel sum) per microscope field.
In parallel, we determined the wound levels of reparative and anti-inflammatory interleukin-10 (IL-10) and M2-like F4/80 Mφs, as well as inflammatory tumor necrosis factor α (TNFα) and M1-like F4/80 Mφs using the immunofluorescent histological procedures outlined below. We incubated the burn wound cryosections with three primary antibodies: (1) rat anti-mouse F4/80 (a mouse macrophages marker) antibody; (2) rabbit anti-mouse CD206 (a marker of M2-like macrophage phenotype) antibody or rabbit anti-mouse iNOS (a marker of M1-like macrophage phenotype) antibody; and (3) goat anti-mouse IL-10 antibody or goat anti-mouse TNFα antibody. These antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The sections were then incubated with three secondary antibodies: Cy3-labeled Donkey anti-rat IgG, Alexa Fluor 488-labeled Donkey anti-Rabbit IgG, and Alexa Fluor 633-labeled Donkey anti-Goat IgG (Life Technologies, Grand Island, NY). After the staining of wound sections with these fluorescently tagged secondary antibodies, the IL-10 or TNFα became fluorescently Magenta; and CD206 or iNOS became fluorescently green; and F4/80 turned into fluorescently red. Nuclei were counterstained fluorescently blue with 4′-6-diamidino-2-phenylindole (DAPI, 1:2,000, 10 min) (Life Technologies). Sections were washed with 1× PBS buffer (10 min, ∼23°C, three times) after each incubation to remove extra staining reagents. The wound sections on glass slides were mounted using mounting media (Vector Laboratories, Burlingame, CA) and examined and imaged using a confocal microscope with a 40× oil immersion objective (numerical aperture: 1.25) and other settings. The images were analyzed by Zeiss Zen-2012 and ImageJ (National Institute of Health, Bethesda, MA), and annotated for presentation using Photoshop. Negative or isotype control staining was conducted using isotype-matched irrelevant IgG of each primary antibody and corresponding secondary antibody to identify potential unspecific staining and background. F4/80+Mφs that were CD206+, iNOS+, IL10+, TNFα+, CD206+IL10+, or iNOS+TNFα+ were counted under the confocal microscope and presented as a percentage of total cells per microscopic field. The tissue levels of IL10 or TNFα were measured as integrated fluorescent intensities per microscopic field.
Statistics
This was conducted using Mann–Whitney ANOVA or t-test. A P-value below 0.05 was considered statistically significant. All data are presented in mean ± standard deviation (SD). The statistical analysis was performed using SAS software (SAS Institute, Cary, NC) and SPSS software (
Results
ACgels maintained MSC viability/survival and adhered to MSCs in culture
To determine the capability of ACgels in sustaining MSC survival in culture, we seeded 1 × 105 mouse MSCs to ACgels by capillary force illustrated as in Fig. 2A. The blotted ACgel took all MSCs in a drop of 30 μL medium. The MSCs scaffolded by ACgels were 93.8%, 95.6%, and 94.9% green at day 1, 3, and 6 of culture, respectively, after the incubation with the live/dead cell assay kit consisted of calcein-AM and ethidium-homodimer (Fig. 2B). This indicated that the MSCs scaffolded by ACgels were 93.8–95.6% viable because only living MSCs can convert calcein-AM to green product, whereas the dead MSCs were red by taking up ethidium-homodimer (Fig. 2C). In comparison, the MSCs of 2D culture had similar viability (92–96%) at day 1, 3, and 6 of culture (Fig. 2C). In addition, the MSCs that were seeded to ACgels adhered to ACgels at 95.9%, 95.2%, and 97.0%, at 15 min, 3 days, and 6 days after the seeding (Fig. 2D). Therefore, ACgels can scaffold and adhere to MSCs and maintain MSC viability and survival in culture without cytotoxicity, similar to previous reports that endothelial cells can adhere well to AA-PEA hydrogels and retain their viability and survival [36,68,69]. A large portion of MSCs on ACgel were polygonal or spindle-like with certain stretching (Fig. 2B), which suggests that the MSCs attached to the ACgel. Few MSCs seeded to ACgels appeared spherical (Fig. 2B).
ACgels sustained the presence of MSCs in the 3° burn wounds of mice
We generated the 3° burn wounds on the dorsal skin of C57BL/6 J mice. The C57BL/6 J mouse MSCs were labeled with red fluorescent Dil and seeded to ACgels. These MSC-carrying ACgels were used to cover the skinless areas generated by the excision of coagulated full-thickness skin of burn wounds at 2 dpb (Fig. 3A). The ACgels sustained the presence of implanted MSCs (red) to wounds at 8 dpb (Fig. 3B), while ACgel fragments were incorporated into wound tissue (Fig. 3C).
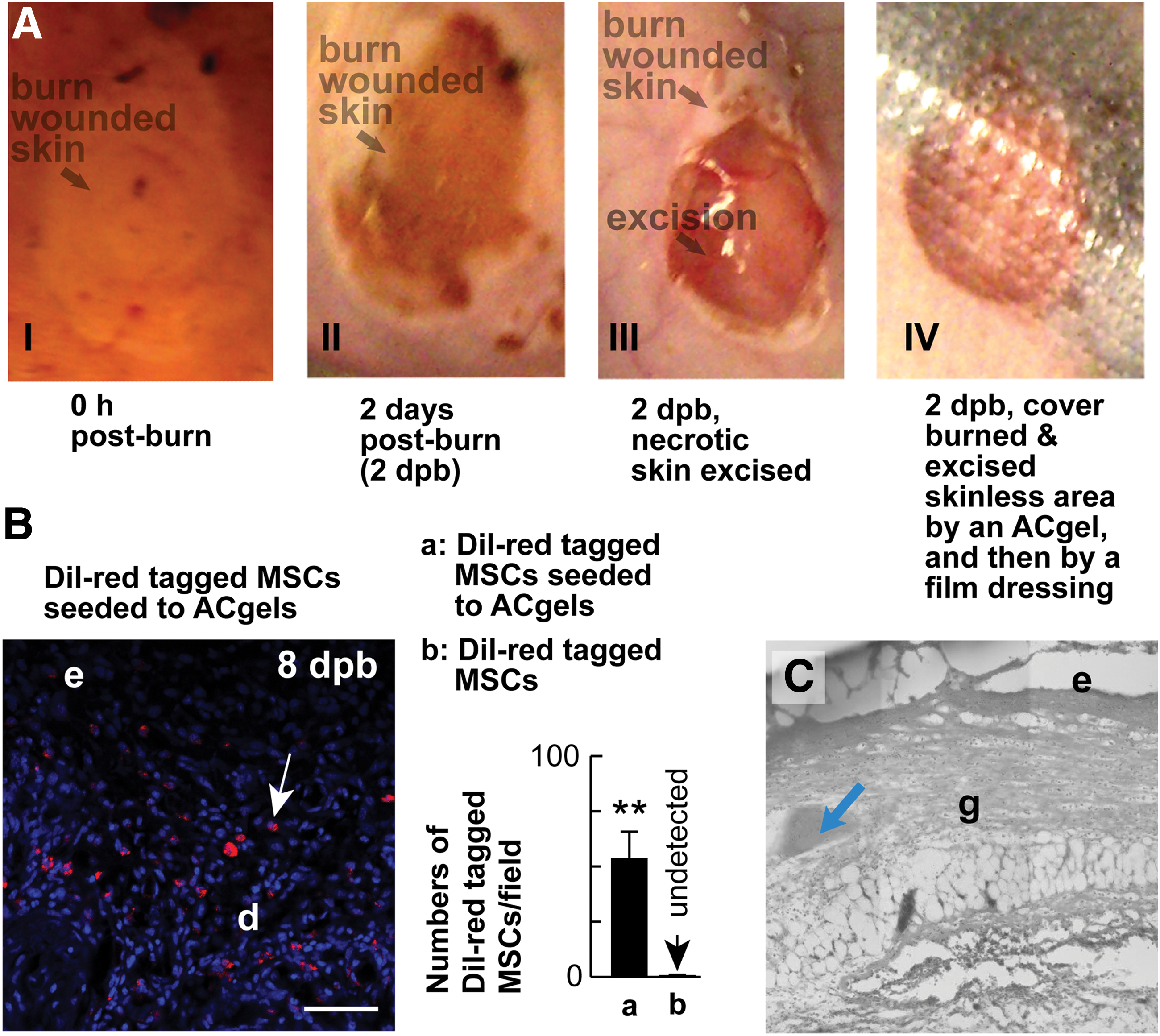
ACgels sustained the presence of MSCs in the 3° burn wounds of C57BL/6 J mice.
MSC-seeded ACgels accelerated the closure of 3° burn wounds
Wound closure is a critical process in the early healing phase that rebuilds the armor against infection and water loss. It cooperates with and promotes other healing processes. The ACgels seeded with syngeneic C57BL/6 J mouse MSCs promoted wound closure of the necrotic skin excised 3° burn wounds of the same strain C57BL/6 J mice by 41% at 8 dpb over the control with medium treatment, whereas ACgels loaded with medium alone did not have substantial effect (Fig. 4). Therefore, MSC-seeded ACgels can significantly accelerate the closure of 3° burn wounds.
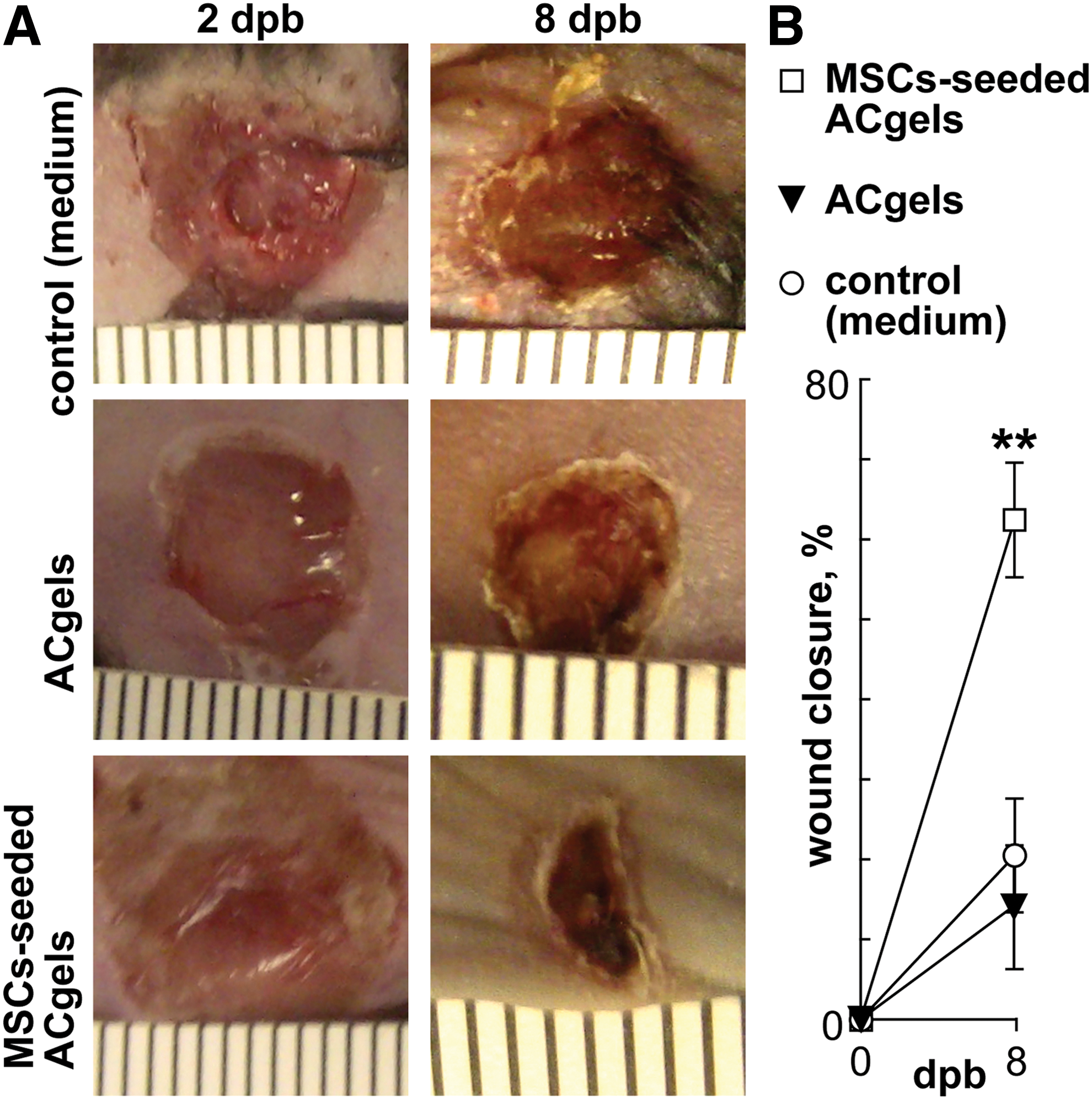
MSC-seeded ACgels accelerated the closure of 3° burn wounds of mice.
MSC-seeded ACgels promoted reepithelialization, granulation tissue formation, and vascularization in 3° burn wounds
Reepithelialization and granulation tissue growth are also crucial processes in the early healing phase of skin wounds resulting from burn injuries. To determine whether these processes were promoted by wound-covering MSC-seeded ACgels, we conducted serial sagittal cryosection through the center region of the widest wound bed on each burn wound. The sections representing widest wound bed were stained with H&E, and then analyzed using Olympus scan microscope. We found that the reepithelialization for the treatment with MSC-seeded ACgels at 8 dpb was significantly higher compared to medium treatment control (22.1% ± 0.7% vs. 16.3% ± 1.9%, P < 0.05, Fig. 5A, B). In addition, the wounds treated by MSC-seeded ACgels yielded larger granulation tissue area than that of medium control, whereas the granulation tissue area from ACgel treatment did not vary from the medium control (Fig. 5A, C). Therefore, the combination of ACgel and MSCs promoted granulation tissue formation in burn wounds [65].
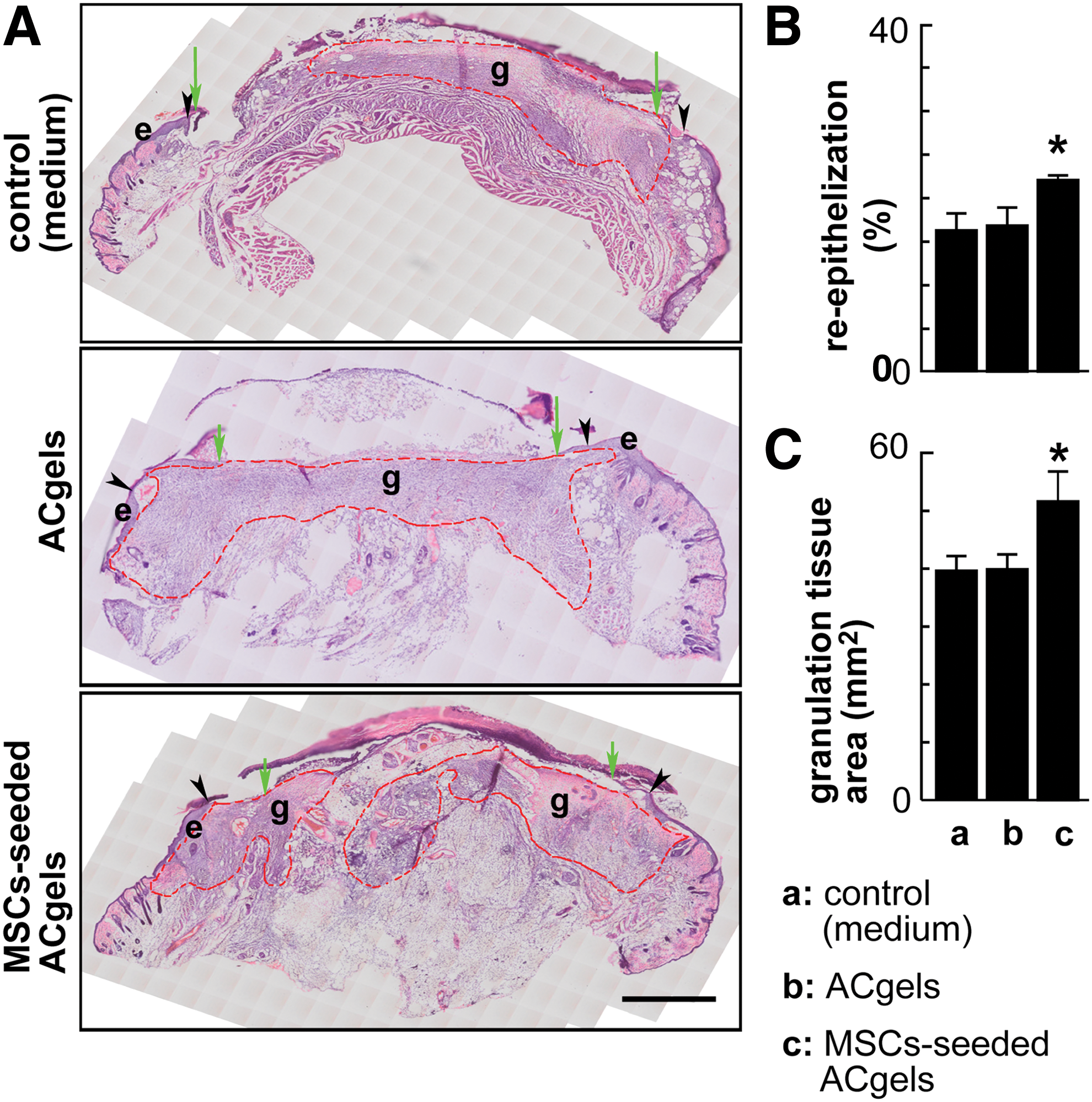
ACgels seeded with mouse MSCs promoted reepithelialization and granulation tissue formation in the 3° burn wounds of mice.
Vascularization regenerates blood vessels in burn wounds; thus, it is essential for wound healing. We studied whether MSC-seeded ACgels or ACgel alone promotes wound vascularization. The wound vascularity was determined by histological measurement of CD31+ area in wound sections. The wounds treated by MSC-seeded ACgels showed markedly highly vascularity compared with medium control and ACgels alone at 8 dpb. The treatment with ACgels alone also resulted in higher CD31+ vascularity compared to medium control (Fig. 6). Moreover, the MSC-seeded ACgels yielded markedly higher vascularity than ACgels alone, suggesting that MSCs significantly contribute to the angiogenic bioaction of MSC-seeded ACgels.

The blood vessel regeneration in the 3° burn wounds at the late inflammatory phase and early proliferative phase was enhanced by ACgels seeded with mouse MSCs or ACgels alone. The burn wound model and treatment were conducted on C57BL/6 J mice as in Figure 4.
Mechanistic insights: induced reparative, anti-inflammatory cytokine IL-10 and M2-like macrophages, and reduced inflammatory cytokine TNFα, M1-like macrophages at late inflammatory phase of burn wound healing
Chronic inflammation is detrimental to wound healing. It is critical for wound repair that macrophages switch from inflammatory M1-like phenotype to reparative anti-inflammatory M2-like phenotype and produce reparative anti-inflammatory factors, including IL-10, in the late inflammatory phase or proliferative phase of healing. To test whether MSC-seeded ACgels affect these mechanistic processes, we determined the burn wound levels of IL-10, M2-like Mφs, and IL-10 expression Mφs. Both MSC-seeded ACgels and ACgels alone induced higher levels of IL-10 than medium control (3945 ± 264 or 2901 ± 233, respectively, vs. 1247 ± 311 intensity summation of IL-10+ pixels per microscopic field of wound sections) at 8 dpb; In addition, MSC-seeded ACgels were more effective in this induction than ACgels alone (Fig. 7A, B). Both MSC-seeded ACgels and ACgels alone increased the percentages of IL-10 expressing F4/80+ macrophages (IL10+F4/80+) compared with control (16.1% ± 2.8%, P < 0.01 and 12.5% ± 1.9%, P < 0.05, respectively, vs. 5.6% ± 1.1%); and MSC-seeded ACgels increased more than ACgels alone did although the difference was not significant (Fig. 7A, C). MSC-seeded ACgels and ACgels alone led to higher percentages of M2-like CD206+F4/80+ macrophages among total cells (with DAPI-blue nuclei) in burn wounds than medium control did (25.3% ± 5.1%, P < 0.01 and 19.7 ± 3.1, P < 0.05, respectively vs. 10.1% ± 4.8%), whereas MSC-seeded ACgels did not make a significant difference with ACgels alone (Fig. 7A, D). Furthermore, the percentages of IL-10 expressing M2-like CD206+F4/80+Mφs (IL-10+CD206+F4/80+Mφs) in burn wounds descended from MSC-seeded ACgels (13.1% ± 1.9%, P < 0.01 vs. control), ACgels alone (7.8% ± 1.0%, P < 0.05 vs. control), to medium control (4.7% ± 0.4%), which indicated that MSC-seeded ACgels increased the portion of IL-10 expressing M2-like Mφs among the cells of 3° burn wounds. Therefore, MSC-seeded ACgels or ACgels alone can markedly induce the wound levels of anti-inflammatory IL-10, IL-10 expressing Mφs, M2-like Mφs, and IL-10 expressing M2-like Mφs in the late inflammatory phase of burn wounds. Moreover, MSC-seeded ACgels were markedly more potent in this aspect compared with ACgels alone.
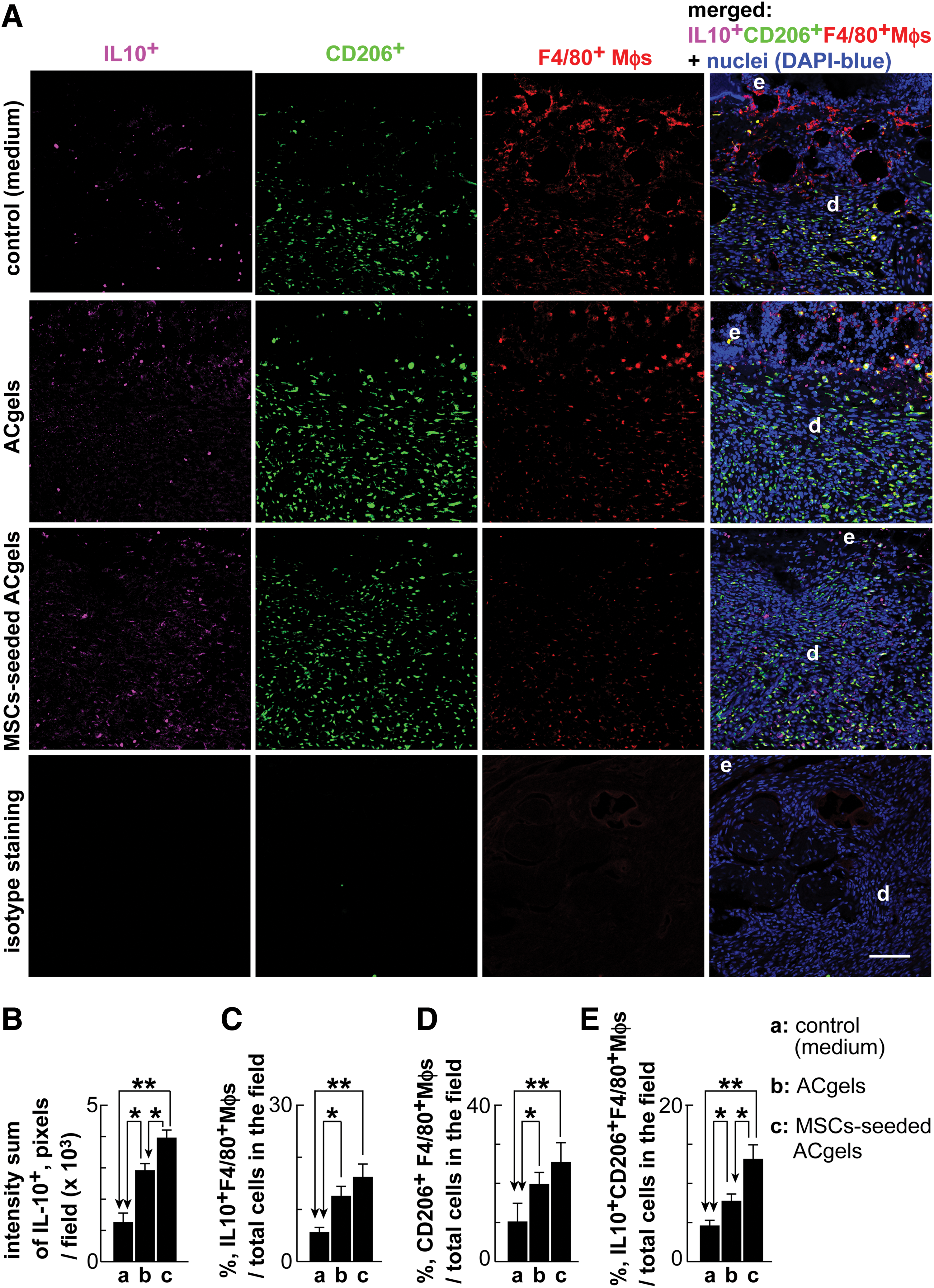
Augmented levels of reparative, anti-inflammatory IL-10 and M2-like Mφs at the late inflammatory phase of burn wounds by MSC-seeded ACgels or ACgels alone.
We further determined burn wound levels of inflammatory TNFα, M1-like Mφs, and TNFα expressing Mφs. Our immunohistological analysis showed that MSC-seeded ACgels or ACgels alone reduced the wound levels of inflammatory TNFα at 8 dpb in the late inflammatory phase of healing compared with the control (1293 ± 239, P < 0.01 and 2935 ± 458, P < 0.05, respectively; versus control 4251 ± 316, the intensity summation of TNFα+ pixels per microscopic field of burn wound sections). MSC-seeded ACgels reduced more TNFα than ACgels alone (P < 0.05) (Fig. 8A, B). Since Mφs are the key cells regulating the late inflammatory phase of healing, we determined Mφ expression of TNFα in wound sections. MSC-seeded ACgels or ACgels alone reduced the percentages of TNFα expressing F4/80+Mφs, that is, TNFα+F4/80+Mφs, among the total wound cells counted by DAPI-blue nuclei, compared with medium control (12.1% ± 4.3%, P < 0.01 and 15.4% ± 2.7%, P < 0.05, respectively; vs. 25.8% ± 3.8% of medium control) (Fig. 8A, C). The treatment by MSC-seeded ACgels or ACgels alone markedly decreased the percentages of M1-like iNOS+F4/80+Mφs among the total DAPI-blue cells in burn wounds compared with medium control at 8 dpb (10.9% ± 5.2%, P < 0.01 and 13.3% ± 4.5%, P < 0.05, respectively; vs. medium control 22.8% ± 3.1%) (Fig. 8A, D). To delineate mechanistic insights further, we determined treatment effects on TNFα expressing M1-like Mφs (ie, TNFα+NOs+F4/80+Mφs) (Fig. 8A, E). The results revealed a trend consistent with the percentages of M1-like iNOS+F4/80+Mφs shown in Fig. 8A, D, and E. The percentages of TNFα+NOs+F4/80+Mφs among the total wound cells counted from DAPI-blue nuclei of wound sections were 6.1% ± 2.7%, 12.9% ± 3.1%, and 21.8% ± 4.2% for MSC-seeded ACgels, ACgels alone, and medium controls, respectively. Statistical analysis of these data indicated that MSC-seeded ACgels and ACgels alone attenuated the wound levels of TNFα+iNOs+F4/80+Mφs (P < 0.01 and P < 0.05, respectively, compared with control) at 8 dpb of late inflammatory phase of healing, and that MSC-seeded ACgels were more effective than ACgels alone in the attenuation (P < 0.05). In aggregate, MSC-seeded ACgels or ACgels alone can significantly reduce the wound levels of inflammatory TNFα, TNFα-expressing Mφs, M1-like Mφs, and TNFα- expressing M1-like Mφs in the late inflammatory phase of burn wounds. MSC-seeded ACgels were more effective than ACgels in these actions.
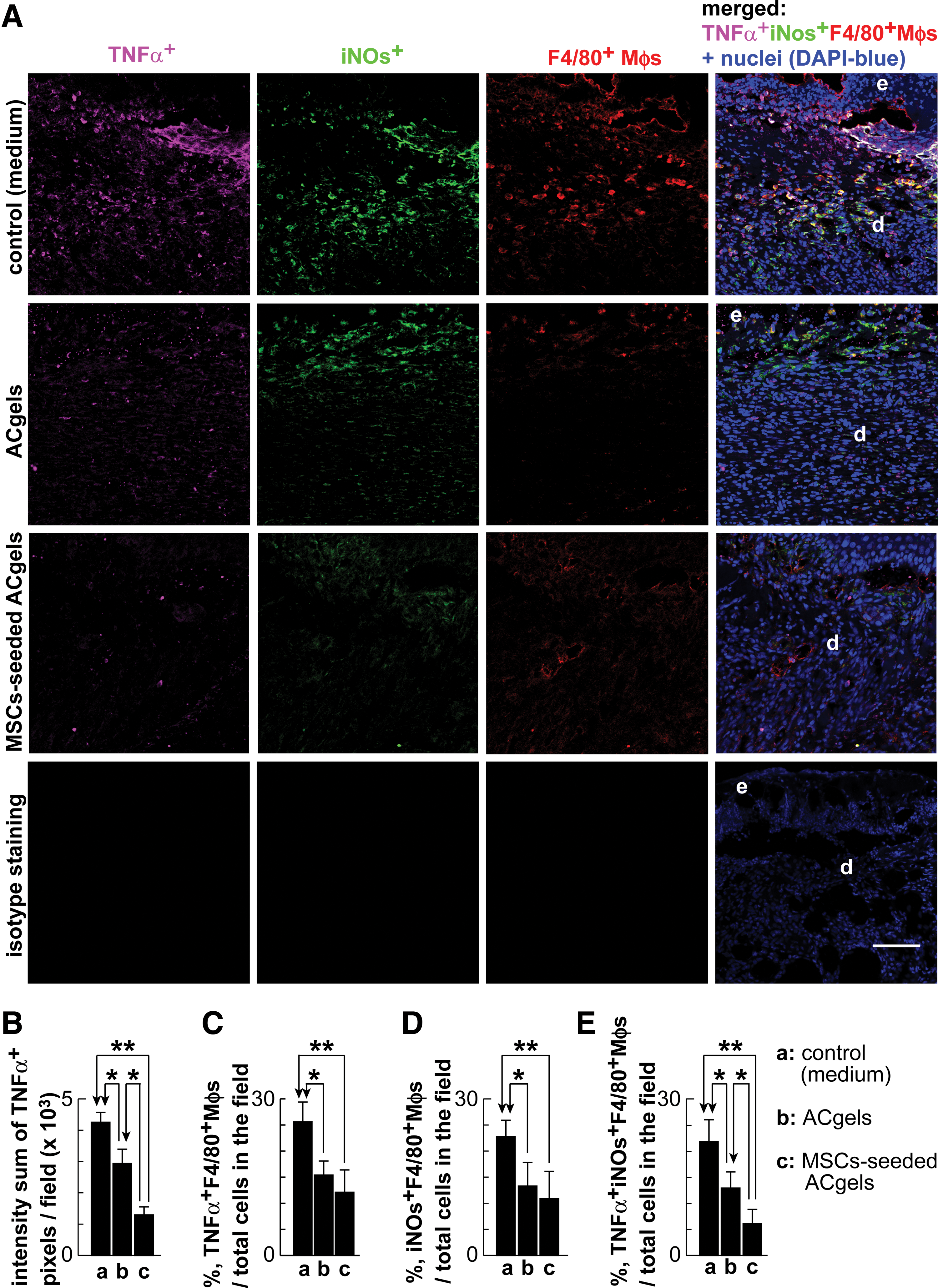
Attenuated levels of inflammatory TNFα and M1-like Mφs at the late inflammatory phase of burn wounds by MSC-seeded ACgels or ACgels alone.
There were no observable difference among the groups of MSC-seeded ACgels, ACgels, and control on mouse behavior (including foraging, grooming, drinking, and eating), neither on the weights nor sizes of mouse livers and spleens, suggesting that the treatments were safe to the mice.
Discussion
The use of hydrogel scaffolds, mostly fabricated from natural biomaterials, as a delivery vehicle for stem or progenitor cells has proven beneficial for cell-homing enhancement in previous studies that examined injury models of the heart [29], ischemic hindlimb [70], diabetic wounds [71], and burn wounds [2,54,72,73]. Amino acid-based poly(ester amide) polymers, or AA-EPAs, are a relatively new class of synthetic biomaterials that are biocompatible, biodegradable, nontoxic, and possess many merits over natural biomaterials and other synthetic biomaterials [32,33]. The hydrogels synthesized from arginine-containing AA-PEA and polysaccharide chitosan, that is, ACgels, exhibit excellent properties, thus are a promising candidate for use as the scaffold and delivery carrier of MSCs and wound coverage for the treatment of severe burn wounds. This utility of ACgels was explored here on the 3° burn wounds of mice. We found that MSC-seeded ACgels markedly promoted burn wound closure, reepithelialization, granulation tissue growth, and blood vessel regeneration. The results suggest that ACgels may enhance MSC functions and the cellular processes of healing [74].
Spontaneous reepithelialization of severe burn wounds is a slow process. It usually takes many days before the wound is resurfaced. To promote the reepithelialization and other healing process, excision of burn-generated necrotic skin and skin grafting are usually conducted for the treatment of severe burns [5]. In this exploratory investigation of the potential therapy using ACgel-carried MSCs, we focused the early stage of burn wound healing, including the inflammatory and proliferative phases. Our results demonstrated the effectiveness of ACgel-carried MSCs and ACgels in the promotion of the critical healing processes, including wound closure, reepithelialization, granulation tissue growth, and/or blood vessel regeneration. Future studies need to evaluate the effectiveness of MSC-seeded ACgels in wound remodeling outcomes, including collagen deposition, scarring, and wound breaking strength.
ACgels incorporated with MSCs were more effective than ACgels alone in promoting reepithelialization, blood vessel growth, and the wound levels of IL-10 and M2-like Mφs and reducing the wound levels of TNFα and M1-like Mφs at late inflammatory phase of 3° burn wounds although ACgels alone demonstrated most of these actions. Based on previous studies of ours and others, at least 5 × 105 MSCs are needed to be implanted to wounds to manifest MSC promotion of wound healing when injected alone (with culture medium or saline as the vehicle [2,73,75,76]); or at least 2.5 × 105 MSCs are needed if MSCs are carried by a pullulan-collagen hydrogel bioscaffold [60]; that is, 1 × 105 MSCs are not enough to have the actions under these conditions. In contrast, 1 × 105 MSCs carried by ACgels here was markedly effective in these actions, and are significant better than ACgels alone. Therefore, ACgels significantly promoted MSC functions in burn wound healing. Moreover, an adequate increase of MSC numbers would promote more healing of burn wounds. However, there may be a “ceiling” for the effect, beyond which increasing the MSC number may not be able to increase further the promotion of healing. Reversely, using less MSCs is likely to result in little to no promotion of burn wound healing.
The good adhesion of MSCs to ACgels (Fig. 2D) positively correlates with the high viability of MSCs seeded to ACgels (Fig. 2C), suggesting the potential contribution of adhesion to MSC viability [77,78]. The pendant guanidine groups in Args of ACgels (Fig. 1A) exist in positively charged guanidinium form in physiological conditions (∼pH 7.4). Thus, ACgels are positively charged in vivo and in vitro. Many hydrogels rely on tethering via arginylglycylaspartic peptides (RGDs) for cell attachment [79,80]. In contrast, positively charged and protein-mimicking ACgels (Fig. 1A) provide excellent attachment to cells (negatively charged) without RGD ligands [31,40,68,69,81 –83]. The appropriate attachment of MSCs could be also attributable to arginine containing amino acid based polyester amide that could mimic RGD ligands in certain aspects, similar to other biopolymer hydrogels reported [77] in addition to the positive charge effects of ACgels.
The results of MSC adhesion to ACgels and survival on ACgels are consistent with our results on MSC presentation in burn wounds at 8 dpb (Fig. 3B). The trophic property of ACgels on MSCs provides the basis for us to use ACgels as the scaffolds or carriers for MSC implantation to wounds. The augmentation of MSC viability through ACgel delivery is in turn associated with the effective promotion of burn wound healing and blood vessel regeneration. Defining the contribution of MSC adhesion on ACgels to the MSC viability will need an array of ACgels with different capabilities to adhere to MSCs, which is meaningful but beyond our scope that focuses on testing the effects of the combination of ACgels and MSCs on severe burn wounds and associated macrophage phenotypes.
Based on our results in this report, the positive zeta potential +14.15 mV of ACgels used in this report is adequate so that it does not cause any cell toxicity. In general, some cationic polymers, such as pure arginine-based polypeptides, are cytotoxic. However, ACgels are not pure Arg-based polypeptides or chitosan because the Arg moiety and chitosan in the polymers backbone are considerably diluted by non-Arg and non-chitosan building blocks; hence, their cationic charge densities are much lower than a pure Arg-based biopolymer or chitosan alone. All our earlier published Arg-PEA biopolymers in a variety of physical forms from 3D microporous hydrogels, films, micro/nanospheres, micelles, and fibrous membranes, have shown excellent cell biocompatibility in vitro and in vivo [32,33]. The 11 types of cells studied for cell biocompatibility of AA-PEA biopolymers include mouse aortic smooth muscle cells, NIH 3T3 fibroblasts, bovine aortic endothelial cells, rat macrophages, porcine aortic valve smooth muscle cells, mouse preosteoblastic MC3T3-E1 cells, human type like Detroit 539 fibroblast primary cells, human umbilical vein endothelial cells, human coronary artery endothelial cells, human prostate cancer cells, and Hela cells. The recent in-depth reviews of our AA-PEA biopolymers and their hybrids, including their biocompatibility, are available in references [32,33].
This ACgel seeded with MSCs is for both artificial skin tissue filling (ie, replacing the severe wounded skin) and wound coverage. The MSCs are gradually released from ACgels to wounds, as suggested by the presence of Dil-tagged MSCs in wounds (Fig. 3B) and ACgel residues after biodegradation (Fig. 3C) at 8 days postburn. The MSCs injected to wounds all at once without ACgels, however, were not able to survive in burn wounds for such long time [77,84], therefore, were not tested in this study (with an additional concern that MSCs alone applied multiple times would be not comparable to the ACgel and MSC combination since the hydrogel and MSC combination is only applied to the wound once). MSCs are not exactly immobilized by ACgels. Instead, ACgels just provided temporal scaffolds and housing to MSCs so that MSCs are not added to wound all at once, which may reduce MSC survival and prohealing functions. The MSCs that temporally lived in ACgels and gradually migrate to wounds should also produce paracrines that can be released to wounds and promote angiogenesis, anti-inflammation, and tissue regeneration. In the future, we should decipher the actions of these paracrines produced by MSCs in ACgel scaffolds on the wound healing process.
Most studies have shown that MSCs promote the healing of chronic wounds and severe burns mainly by paracrine actions, that is, by producing reparative factors. Only a very small percentage of implanted MSCs differentiate into skin cells [14,15]. Here, we targeted chronic inflammation and Mφs in burn wounds, since chronic inflammation in chronic wounds is the key culprit that impairs the healing [2,54,72,73] and Mφs are the key player in wound healing and chronic inflammation [85,86]. Burn injury markedly increased TNFα levels compared with nonburn wounding in mice at 3 days postburn in infiltrating cells, wound fluid, and skin from burn wounds [87]. High levels of TNFα are associated greatly with pathologies in wounds [88]. It is reported that levels of anti-inflammatory reparative IL-10 are reduced in burn wound cells [89] and MSCs induce IL-10-producing Mφs in vitro and in vivo [90]. IL-10 modulates immune cells toward wound healing. IL10 is also involved in endothelial activation and leukocytes recruitment in wounds. We found that ACgel-carried MSCs markedly increased wound levels of IL-10 cytokine and M2-like Mφs, but reduced the levels of inflammatory TNFα cytokine and M1-like Mφs, suggesting a novel mechanism for the reparative actions of MSC-seeded ACgels [91,92]. Our results indicate that ACgel scaffolds facilitate MSC functions in the immunomodulation that reduces the late phase inflammation, and switches Mφs to M2-like phenotype in the 3° burn wounds. Consistent with this notion, ACgel-carried MSCs increased IL-10 expressing Mφs and reduced TNFα expressing Mφs (Figs. 7D, E, and 8D, E). ACgels may also promote MSC production of other prohealing growth factors and cytokines and modulation of other cells, including karatinocytes, fibroblasts, endothelial cells, dendritic cells, and master cells important in burn wound repair, which needs to be studied in the future.
Successful healing of burn wounds involves well-coordinated cellular and molecular processes. Generally, chronic inflammation associated with high levels of TNFα and M1-like Mφs and low levels of IL-10 and M2-like Mφs correlates with impairment of wound healing [65,74,93,94]. Moreover, wound vascularization in the early phase of wound healing positively correlates with wound healing [95 –97]. However, for the specific situation of ACgels, some of these correlations did not manifest since the treatment on burn wounds with ACgels alone did not accelerate the wound healing (closure, reepithelialization, and granulation tissue growth) (Figs. 4 and 5), although ACgels alone had similar actions as ACgels seeded with MSCs on the modulation of macrophages and promotion of wound vascularization (Figs. 6 –8). These results suggest that some factors we have yet to study may cause these noncorrelations. It will be meaningful to decipher these mechanisms underlying the noncorrelations, since it may provide a new target to improve the healing of severe burn wounds.
Diabetic wounds and severe burn wounds share several key comment pathological features, including inflammation [1 –5,98]. Our results on severe burn wounds are consistent with the report by Chen et al. showing that the hydrogel treatment alone inhibited the chronic inflammation, and hydrogel-MSCs promoted the formation of granulation tissues in diabetic wounds [99]. ACgel-MSCs may also promote the proliferation and remodeling, similar to their hydrogel-MSCs [99]. It is likely that ACgels can promote MSC production of angiogenic and/or regenerative growth factors, cytokines, and chemokines in wounds, including VEGF, IGF, HGF, IL-10, and so on, similar to their hydrogels [99]. In the future, we should also determine the expression levels of wound healing growth factors, cytokines, and chemokines in wounds that are treated with ACgels seeded with MSCs and ACgels alone to delineate the functions of ACgels on MSCs. The gene analysis by qPCR should be conducted as well to provide more information on how ACgels support the function of MSCs.
Skin grafting is usually required to provide the essential coverage of severe burn wounds. However, it needs the excision of skin from donor sites, which creates new wounds, pain, and complications, and is costly [1,2,4,75,76,100,101]. Many skin substitutes were developed for wound coverage; each has its own functional drawbacks and is costly. MSCs possess promising regenerative ability [2,73,75,76]. Although MSCs has been vigorously investigated in wound repair, more technological development and mechanistic studies are needed to allow reliable MSC usage in clinical applications. Our results suggest that the ACgel wound coverage incorporated with MSCs could be used provisionally to shield severe burn wounds with no or reduced requirement of skin grafting, thus it could avoid or minimize the problems associated with skin grafting. It can also promote the regeneration of skin from burn wounds. This is the first study on the feasibility of ACgel-carried MSCs for severe burn treatment using the mouse model of 3° burns. The results have provided the basis for future optimization of the ACgel-MSC system and evaluation of its translational value for ideal therapy of severe burn wounds.
Footnotes
Acknowledgments
Many thanks to Ms. Lindsey A. McGehee for editing. This work is supported by grants from the National Institutes of Health (R01-DK087800 to S. Hong and C. C. Chu) and the Research to Prevent Blindness, New York, NY. The authors thank the Becky Q. Morgan Foundation for the research funds to C. C. Chu. This work made use of the Cornell Center for Materials Research Facilities supported by the National Science Foundation under award no. DMR-1120296.
Author Disclosure Statement
No conflicts of interest, financial or otherwise, are declared by the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.