
Abstract
Sulfur mustard (SM) exposure, whose symptoms are similar to radiation exposure, can lead to acute injury. Because mesenchymal stromal cells (MSCs) have been used to experimentally and clinically treat acute radiation syndrome, in this study, MSCs were intravenously injected into rats after percutaneous SM exposure. Then, we examined sternum and spleen samples by histopathological and immunohistochemical methods to observe pathological changes. Furthermore, blood samples were taken to test the white blood cell (WBC) count, blood platelet count (BPC), red blood cell count, and the levels of cytokines in the serum. The number of bone marrow karyocytes and the WBC in the MSC + SM group were higher than those in the SM group, and the levels of granulocyte colony-stimulating factor, granulocyte-macrophage colony stimulating factor, monocyte chemoattractant protein-1, interleukin (IL)-1α, IL-5, and interferon-γ in the MSC + SM group remained high at different time points after SM exposure. In addition, the BPC, the level of erythropoietin and the relative weight of the spleen in the MSC + SM group were significantly higher than those in the SM group. Meanwhile, spleens in the MSC + SM group were more hyperplastic and hematopoietic, and had fewer apoptotic cells than in the SM group. Furthermore, rat body weight and locomotion ability in the MSC + SM group were higher than in the SM group. This evidence supports the potential ability of MSCs in immunoregulation and functional improvements to the hemopoietic microenvironment. Intravenous injection of MSCs exerted significant therapeutic effects in rats with percutaneous exposure to SM.
Introduction
Sulfur mustard (bis-[2-chloroethyl] sulfide, SM), a lethal chemical warfare agent, is a highly reactive alkylating vesicant. Due to its high absorbance potential, SM affects various organs, such as skin, eyes, and lungs, and leads to heavy casualties [1 –3]. SM can directly interact with DNA, protein, and glutathione (GSH), among other molecules, which triggers several molecular pathways involved in genotoxic stress, oxidative stress and inflammation, blocks the cell cycle, and induces lipid peroxidation, eventually resulting in apoptosis, necrosis, terminal differentiation, and loss of cellular integrity and function [1,3,4]. The specific mechanism of injury after SM exposure has not been elucidated. Thus, there is no specific and effective prevention or treatment of injury due to SM exposure, and symptomatic and supportive therapy are the main treatment options used in clinics [1,3,5 –8]. However, current therapeutic advances and drug development in the fields of DNA damage repair, oxidative stress, inflammation, and imbalanced calcium homeostasis only relieve symptoms.
Mesenchymal stromal cells (MSCs), usually defined as multipotent MSCs, are a population of cells that can proliferate in vitro as plastic-adherent cells; these cells have fibroblast-like morphology, can form colonies in vitro and differentiate into bone, cartilage, and fat cells. MSCs have been isolated from almost every type of connective tissues, such as bone morrow, adipose tissue, and umbilical cord (UC) [9,10]. Both preclinical and clinical studies have demonstrated the safety of nonimmunologically matched MSCs [11,12]. In addition, given the properties of MSCs such as homing to injury sites, antiapoptosis, immunoregulation, hematopoiesis promotion and tissue repair, the most clinically advanced cellular therapy to date involves the administration of MSCs [10,11]. Results of recently published investigations describe the efficacy of MSCs in a variety of injury and disease models including acute lung injury, diabetes, stroke, and acute radiation syndrome (ARS) [10,13]. The underlying pathology of ARS involves physical and chemical damage to DNA, which affects rapidly dividing cells, and thus, ARS is often subclassified as bone marrow suppression and gastrointestinal (GI) syndrome [13], similar to symptoms of SM exposure. Many investigators speculate that MSC therapy can provoke hematopoietic recovery [10,14], enhance healing of the GI tract [15,16] and skin injury, and ameliorate a variety of additional ARS-associated complications [13]. Studies in animals have indicated that SM exposure also leads to severe bone marrow and GI injury [7,17,18]. Some studies suggest that MSCs are highly resistant to SM and can exert positive effects on patients exposed to SM [19 –21]. Therefore, in this study, to investigate whether MSCs exert positive effects after SM exposure injury, we implanted MSCs into the tail vain of rats exposed to SM. Our results indicated that the intravenous injection of MSCs can dramatically increase the body weight and improve the locomotion ability of rats after SM exposure. Furthermore, we examined the histopathological changes in the bone marrow and spleen. The changes in cell populations and cytokine levels in blood involved in immunoregulation and hematogenesis were also detected.
Materials and Methods
Caution
SM is a highly reactive alkylating vesicant and cytotoxic agent. This agent should be handled only in a well-ventilated fume hood, and the use of gloves and stringent protective measures should be adopted.
Chemicals
SM of ∼97% purity was provided by the Institute of Chemical Defense (Beijing, China).
Animals
Adult male Sprague–Dawley (SD) rats of specific pathogen-free grade were purchased from Laboratory Animal Center, Academy of Military Medical Sciences (Beijing, China). All experiments were conducted in the Evaluation and Research Center for Toxicology, Institute for Disease Control and Prevention of PLA (Beijing, China). All experimental procedures were approved by the Institutional Animal Care and Use Committee at the Institute for Disease Control and Prevention of PLA (approval no: 2015-024) and were in strict accordance with the Guide for the Care and Use of Laboratory Animals published by the United States National Institutes of Health. Every effort was made to minimize the number of animals used and their suffering. Water and food were available ad libitum in the cages.
Isolation, culture, and identification of human UC-MSCs
Human UC-derived MSCs were provided by the Ivy Institute of Stem Cells Co. Ltd. (China). MSCs were cultured in human MSCs serum-free culture medium (Premedical Lab, China) until well-developed colonies of the fibroblast-like cells reached 80% confluence. Then, the cells were trypsinized with 0.25% trypsin-ethylenediaminetetraaceticacid (Invitrogen) and harvested for injection or passage into new flasks for further expansion. Before injection, the harvested cells were rinsed three times in saline and the injection was performed within 2 h of harvest.
For identification, a portion of the harvested MSCs was washed and resuspended in PBS and stained with fluorescein-conjugated antibodies against CD34, CD44, CD45, CD73, CD90, CD105, and HLA-DR (Pharmingen, CA). Then, the cells were analyzed in a flow cytometer using Cell Quest software (Becton-Dickinson, CA).
For osteogenic differentiation, MSCs were plated in osteogenic induction medium [Dulbecco's minimum essential medium (DMEM), 10% fetal bovine serum (FBS), 20 mM b-glycerophosphoric acid, 50 μM vitamin C, and 10 nM dexamethasone]. To induce adipogenic differentiation, MSCs were cultured in adipogenic induction medium (DMEM, 10% FBS, 1 μM dexamethasone, 200 μM indomethacin, and 0.5 μM 1-methyl-3-isobutylxanthine). For chondrogenic differentiation, MSCs were cultured in chondrogenic induction medium (DMEM, 10% FBS, 50 μM vitamin C, 77.5 nM dexamethasone, 1% ITS Plus Culture Supplement, and 10 ng/mL transforming growth factor-β1).
Percutaneous exposure to SM and MSCs injection
After acclimatization for 7 days, SD rats weighing 200 ± 20 g were randomly assigned into four groups (control group, MSC group, SM group, and MSC + SM group) and treated by dermal exposure. After closely clipping the hair (the hair was clipped 12 h before the application), a fresh dilution of SM in dimethylsulfoxide (DMSO) (SM:DMSO = 1:8) was uniformly smeared with a microsyringe on the back of rats in the SM and MSC + SM group to an ∼0.3 cm2 area at an SM dose of 3 mg/kg bodyweight. Pure DMSO was smeared on the rats in the control and MSC groups in the same way. A dressing was applied to the exposed area and left for 4 h.
Rats in the MSC and MSC + SM groups were injected with MSCs (3.5 × 105 cells each) through the tail vein in 600 μL volume saline at 24, 48, and 72 h after exposure; meanwhile, the control and SM groups were injected 600 μL volume saline. In addition, rats in all the four groups were intravenously injected 10% glucose in 600 μL volume saline at 36, 60, and 84 h after exposure.
Body weights were monitored every 2 days since SM exposure. To measure the number of leukocytes, erythrocytes, and blood platelets, venous blood samples were drawn from each rat and tested at 4, 7, 10, 14, 17, and 20 days after SM exposure.
Multiplex immunoassays
Blood samples were collected from the aorta abdominalis at 4, 7, 10, and 14 days after SM exposure. The levels of granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), interleukin-1α (IL-1α), interleukin-5 (IL-5), monocyte chemoattractant protein-1 (MCP-1), interferon-gamma (IFN-γ), and erythropoietin (EPO) in the serum were determined using commercially available high-sensitivity multiplex immunoassays kits according to the manufacturer's protocols (Invitrogen).
Histopathological and immunohistochemical examination
Sternum and spleen samples were isolated from rats at 4, 7, 10, 14, and 20 days after SM exposure and fixed in standard 4% formaldehyde immediately after collection. The samples were dehydrated in an alcohol series and embedded in paraffin. Five-micrometer-thick sections were prepared. After being dewaxed and rehydrated, the sections were stained with hematoxylin and eosin (H&E) and examined. Apoptosis in the spleen was evaluated by TUNEL assay according to the manufacturer's instructions (Roche). In addition, the Ki67 and CD34-positive cells were detected by immunohistochemistry using antibodies (Abcam, United Kingdom). In the “Results” section, we exhibited the representative pathological changes at different times after SM exposure.
Open field test
The locomotion of rats was monitored every 2 days by the open field test. The open field box consisted of a square black box (40 cm × 40 cm × 45 cm). Each animal was placed and monitored in the box for 10 min, during which autonomic activities and the amount of time and distance traveled were measured with video tracking over the last 7 min of the test.
Statistical analysis
All results represent at least three independent experiments and are expressed as the means ± standard error of mean. The data were analyzed with IBM Statistical Package for the Social Sciences (SPSS; version 19) by a single factor analysis of variance, and the differences among the means of all groups were considered significant at P < 0.05 and very significant at P < 0.01.
Results
Characterization of MSCs
MSCs exhibited a spindle-shaped fibroblastic appearance (Fig. 1a). The presence of surface markers of MSCs was examined by flow cytometry. Most of the cultured cells were positive for CD44, CD73, CD90, and CD105 expression but negative for HLA-DR and hematopoietic antigens CD34 and CD45 (Fig. 1b). Osteogenic differentiation was indicated by calcium deposition using Alizarin red staining. Adipogenic differentiation was shown by the accumulation of lipid vacuoles, which were stained by Oil Red O. Chondrogenic differentiation was identified by the observation of Safranin O-stained cartilage tissue (Fig. 1c).
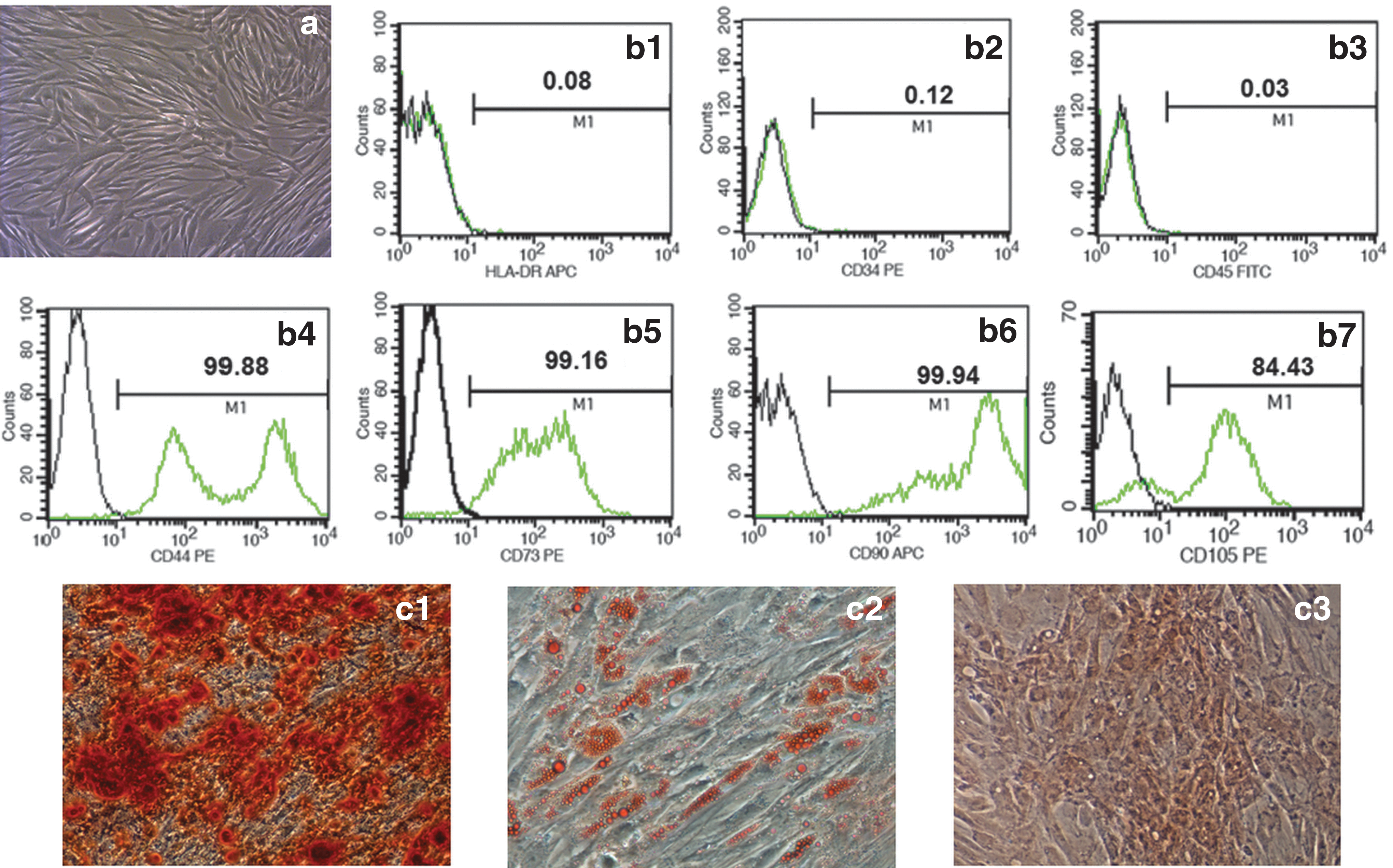
Characterization of MSCs. MSCs exhibit a spindle- and fibroblast-like shape
Influence of MSCs on bone marrow suppression and leukocytes in blood of rats exposed to SM
To observe sternum medulla, we stained sections with H&E after exposure. SM exposure led to a significant decrease in the number of bone marrow karyocytes including hematopoietic cells and leukocytes relative to that in the control group (Fig. 2a). We determined the number of karyocytes in five fields of vision in each rat in each group (n = 4). However, at 4 days after SM exposure, (as indicated by the arrow in Fig. 2a) the karyocyte count in the MSC + SM group was significantly higher than that in the SM group (P < 0.01) (Fig. 2b). At 7 days, the bone marrow showed signs of regaining karyocytes, with organized structure and revitalized proliferation. Eventually, bone marrow made fully recovery at 10 days.
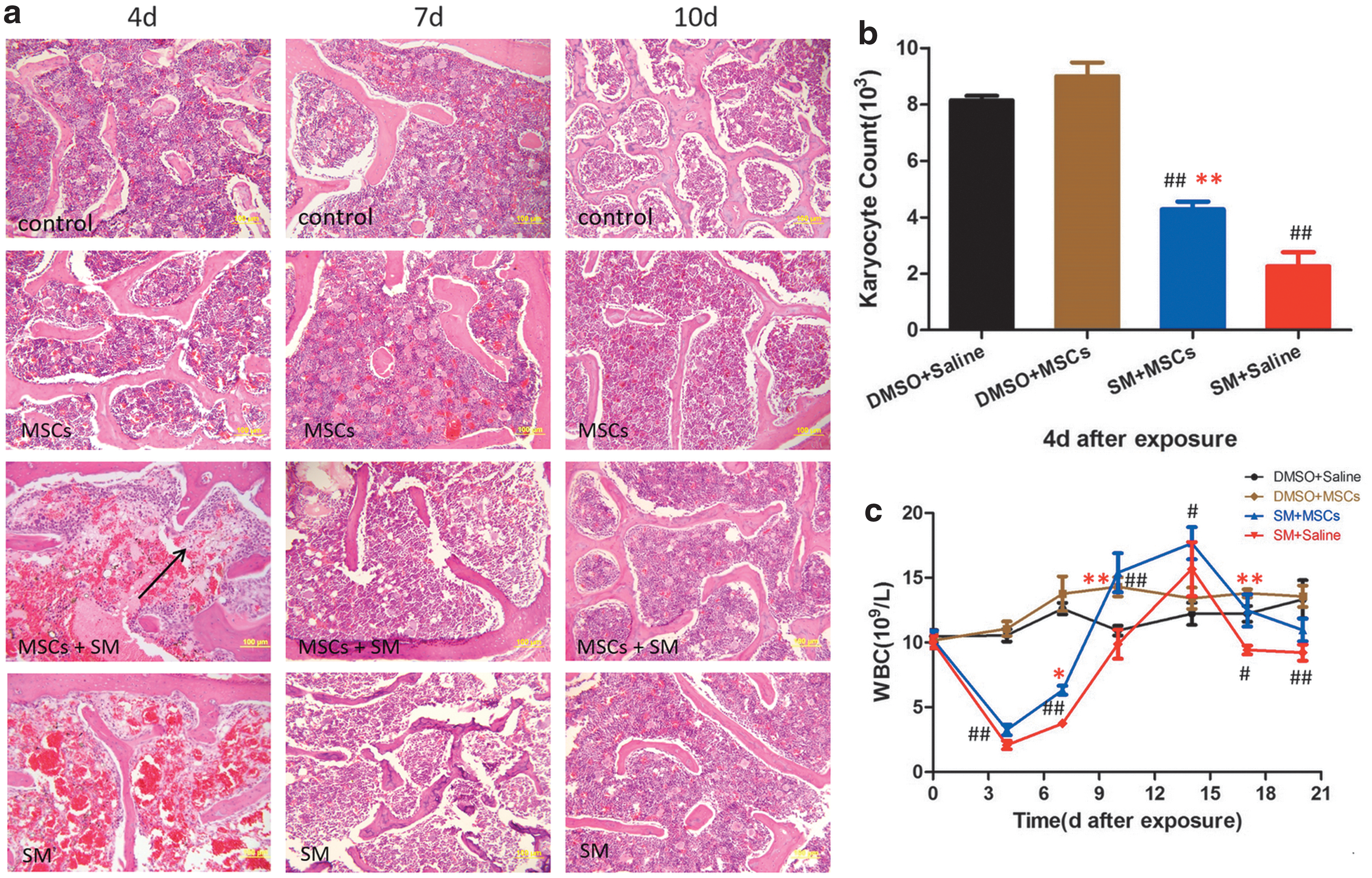
Additionally, the leukocyte counts [white blood cell (WBC) count] in the venous blood of rats were monitored at 4, 7, 10, 14, 17, and 20 days after SM exposure. WBC (n = 6) of the SM and MSC + SM groups was significantly lower than that of the control group (P < 0.01) during the first 10 days after SM exposure (Fig. 2c). However, both the SM and MSC + SM groups demonstrated a gradual improvement from 7 days and caught up with the control group after 10 days, especially the MSC + SM group, which had higher WBC than that in the control and SM groups (P < 0.01) at 10 days. In addition, WBC of the SM group was obviously lower than that of the control group at 17 (P < 0.05) and 20 days (P < 0.01).
Influence of MSCs on the serum levels of G-CSF, GM-CSF, MCP-1, IL-1α, IL-5, and IFN-γ in rats exposed to SM
The levels of cytokines involved in cytothesis and immunoregulation, including G-CSF, GM-CSF, MCP -1, IL-1α, IL-5, and IFN-γ, in the serum of each group (n = 3 or 4, severe hemolysis in some samples caused the differences in the number of samples per group) were detected at 4, 7, 10, and 14 days after SM exposure. As shown in the Figure, the levels of G-CSF (Fig. 3a) and GM-CSF (Fig. 3b) in the MSC + SM group were significantly higher than those in the SM group (P < 0.05) at 7 days. Regarding the level of MCP-1 (Fig. 3c), the MSC + SM group had higher values than the SM group at 10 days (P < 0.05), but at 4 days, the level in the SM group was higher than that in the remaining three groups (P < 0.05). At 4 days, the levels of IL-1α (Fig. 3d) in the SM and MSC + SM groups were significantly lower (P < 0.01) than that in the control group. Thereafter, the level of IL-1α in the SM group remained low, while the level in the MSC + SM group was obviously higher than that in the SM group (P < 0.05) beginning at 7 days. There was no significant difference among the four groups regarding the serum levels of IL-5 (Fig. 3e), except at 10 days, when the value in the MSC + SM group was higher than those in the other three groups (P < 0.05). Nevertheless, the level of IFN-γ in the MSC + SM group was higher at 4 days than those in the other three groups (P < 0.05) (Fig. 3f).
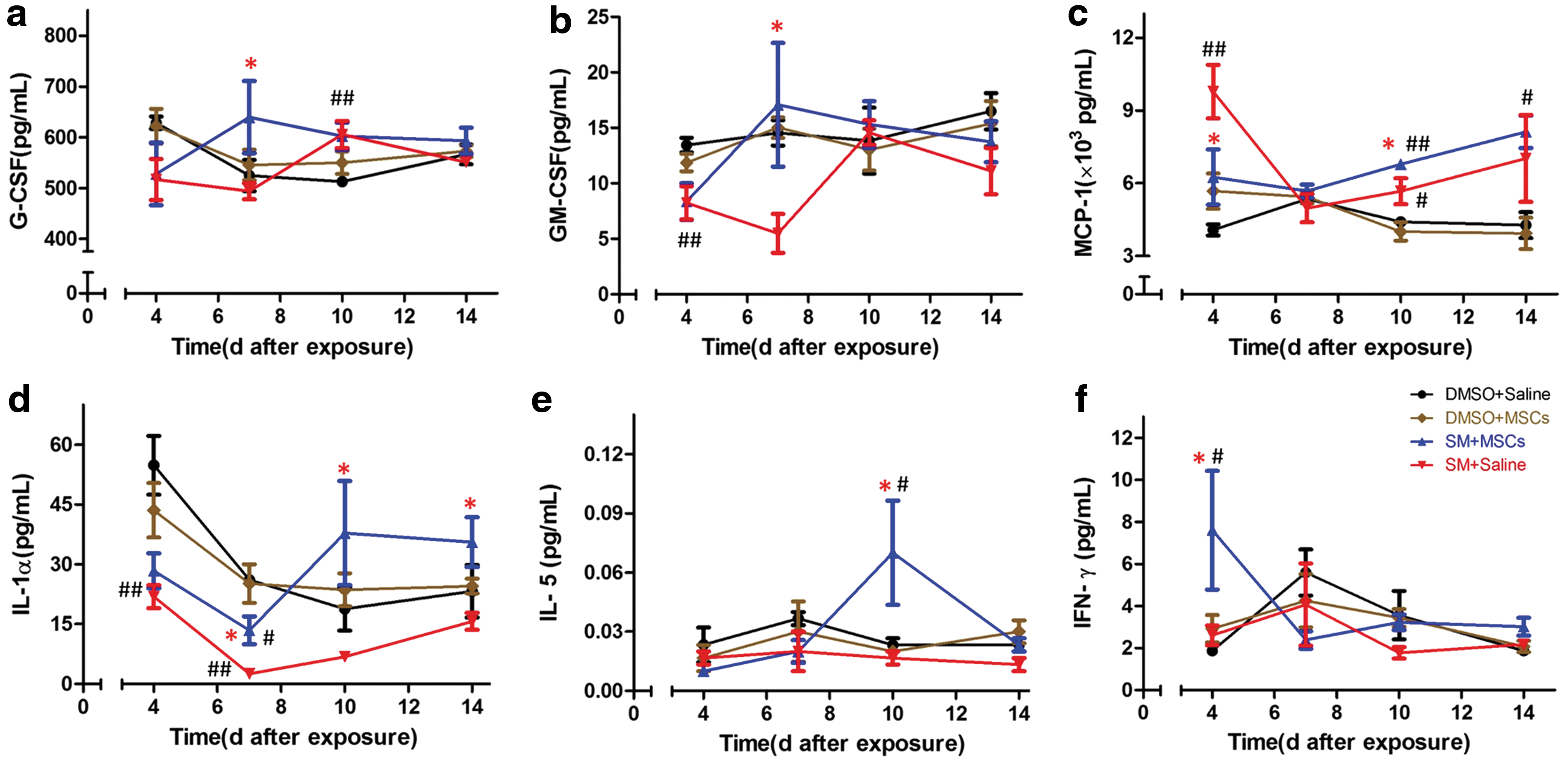
Time-dependent changes in the levels of G-CSF
Influence of MSCs on erythrocytes and blood platelets in rats exposed to SM
At the same time, blood platelet count (BPC) and erythrocyte count [red blood cell (RBC) count] in the venous blood of rats were monitored at 4, 7, 10, 14, 17, and 20 days after SM exposure. The BPC (n = 6) of the SM and MSC + SM groups was significantly lower than that of the control group (P < 0.01) during the first 10 days after SM exposure and was restored later (Fig. 4a). The BPC of the MSC + SM group was significantly higher than that of the SM group (P < 0.01) from 7 days onward. At 10 days, the BPC of the SM group was significantly lower than that of the control group (P < 0.01), at which point, the value in the MSC + SM group had returned to normal. At 14 days, the BPC of the SM and MSC + SM groups was significantly higher than that of the control group (P < 0.01), but both values decreased to that in the control group at 17 days. For the RBC (n = 6), there was no obvious difference between the SM and MSC + SM groups. However, at 17 and 20 days, RBC of the SM group was significantly lower than that of the control group (P < 0.01) (Fig. 4b).

Time-dependent changes in
In addition, the level of EPO (n = 3 or 4, severe hemolysis in some samples caused the differences in the number of samples per group) (Fig. 4c) in the SM and MSC + SM groups was significantly higher (P < 0.01) than that in the control group at 7 and 10 days, but the level in the MSC + SM group was higher than that in the SM group especially at 7 days (P < 0.01). At 14 days after SM exposure, the level of EPO in the SM and MSC + SM groups returned to normal.
Promotive effect of MSCs on splenocyte proliferation in rats exposed to SM
In this experimental model, we stained spleen sections with H&E at 4, 10, and 20 days after SM exposure (Fig. 5a), and monitored the relative spleen weight (spleen weight/body weight × 100%, n = 4) at 4, 7, 10, 14, and 20 days (Fig. 5b) after SM exposure. At 4 days, both the white pulp area and red pulp area and the cellularity of the spleen were decreased in the SM and MSC + SM groups, and the relative spleen weight was significantly decreased (P < 0.01) compared with that in the control group. However, at 10 days, the relative spleen weights in the SM and MSC + SM groups were restored, and it became much higher in the MSC + SM group than in the SM group (P < 0.01). In addition, compared with that in the control and SM groups, the red pulp in the spleen became significantly hyperplastic in the MSC + SM group at 10 days (as indicated by the arrow in Fig. 5a). At 14 days, the relative spleen weight in the SM and MSC + SM groups was significantly higher than that in the control group (P < 0.01), and the value in the MSC + SM group was higher than that in the SM group (P < 0.05). At 20 days, the spleen returned to normal in both the SM and MSC + SM groups.
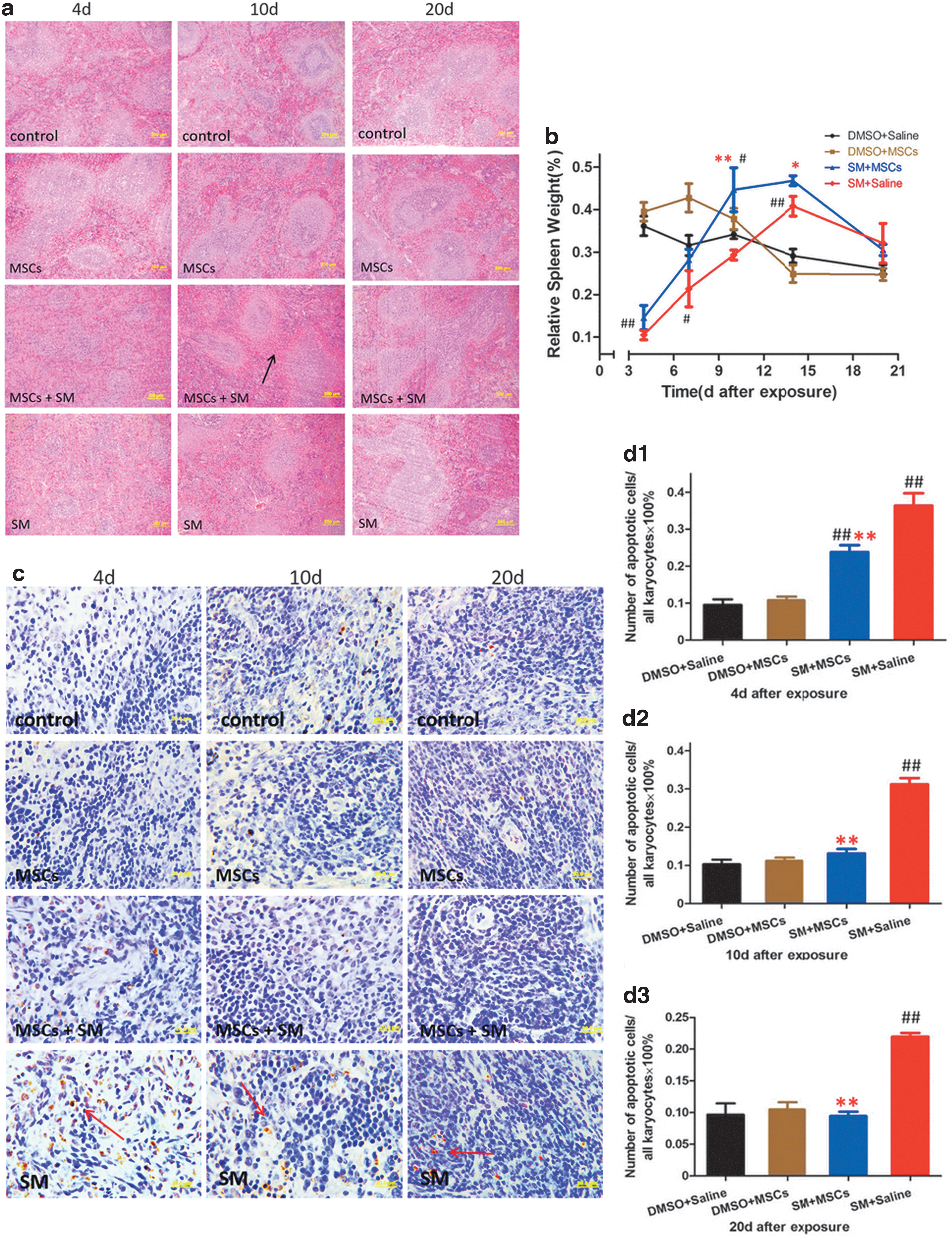
Meanwhile, apoptotic cells were studied on histological slides of the spleen by TUNEL assay (as indicated by the arrow in Fig. 5c). We determined the number of apoptotic cells in five fields of vision and calculated the percentage of apoptotic cells in each rat (Fig. 5d) in each group (n = 4). At 4 days after SM exposure, there were more apoptotic cells in the spleen of rats in the SM and MSC + SM groups than that in control group (P < 0.01), and the value in the SM group was higher than that in the MSC + SM group (P < 0.01). However, from 10 days onward, the number of apoptotic cells in the spleen of the rats in the MSC + SM group returned to normal, while that in the SM group remained high (P < 0.01).We also detected the expression of Ki67 in the spleen (Supplementary Fig. S1; Supplementary Data are available online at
Activation of splenic CD34-positive cells during myelosuppression
In this experimental model, CD34-positive cells were studied on histological slides of bone morrow and spleen by immunohistochemistry. We determined the number of CD34-positive cells in five fields of vision and calculated the percentage in each rat per group (n = 4). At 4 days after the SM exposure, more CD34-positive cells were observed in the spleen (Fig. 6c, d) from the rats in the SM and MSC + SM groups than that in the control group (P < 0.01), and the value in the SM group was lower than that in the MSC + SM group (P < 0.01). Meanwhile, due to the significant decrease in the number of bone marrow karyocytes, the total number of CD34-positive cells in the bone morrow (Fig. 6a, b) in the SM and MSC + SM groups was lower than that in the control group despite the higher relative value in the MSC + SM group (P < 0.01). However, CD34-positive cells in the bone morrow in the SM group continually decrease at 7 days (P < 0.05) and 10 days (P < 0.01) compared with those in the control group, and those in the spleen of rats in the MSC + SM group were greater than those in control groups at 7 days (P < 0.05). Moreover, there was no difference in the number of CD34-positive cells in the spleen between each group at 10 days.

MSC transplantation could increase body weight and locomotion ability of rats exposed to SM
After SM exposure, rats were observed every 12 h. The body weights and autonomic activities were monitored every 2 days. In this study, all mortality was observed within 3–5 days after SM exposure (Fig. 7a). The body weights (n ≥ 8, the difference in the number of rats per group was due to death) of rats in the SM and MSC + SM groups decreased dramatically after SM exposure (P < 0.05) and began to recover from 4 days onward (Fig. 7b). Meanwhile, the body weight in the SM group decreased to a lower extent than that in the MSC + SM group, especially at 4 days (P < 0.05). In addition, the average distance traveled by the rats in the MSC + SM group (n = 6) was significantly higher than that traveled by rats in the SM group (P < 0.05) (Fig. 7c), suggesting that MSCs-treated rats had greater locomotion ability and more vitality.
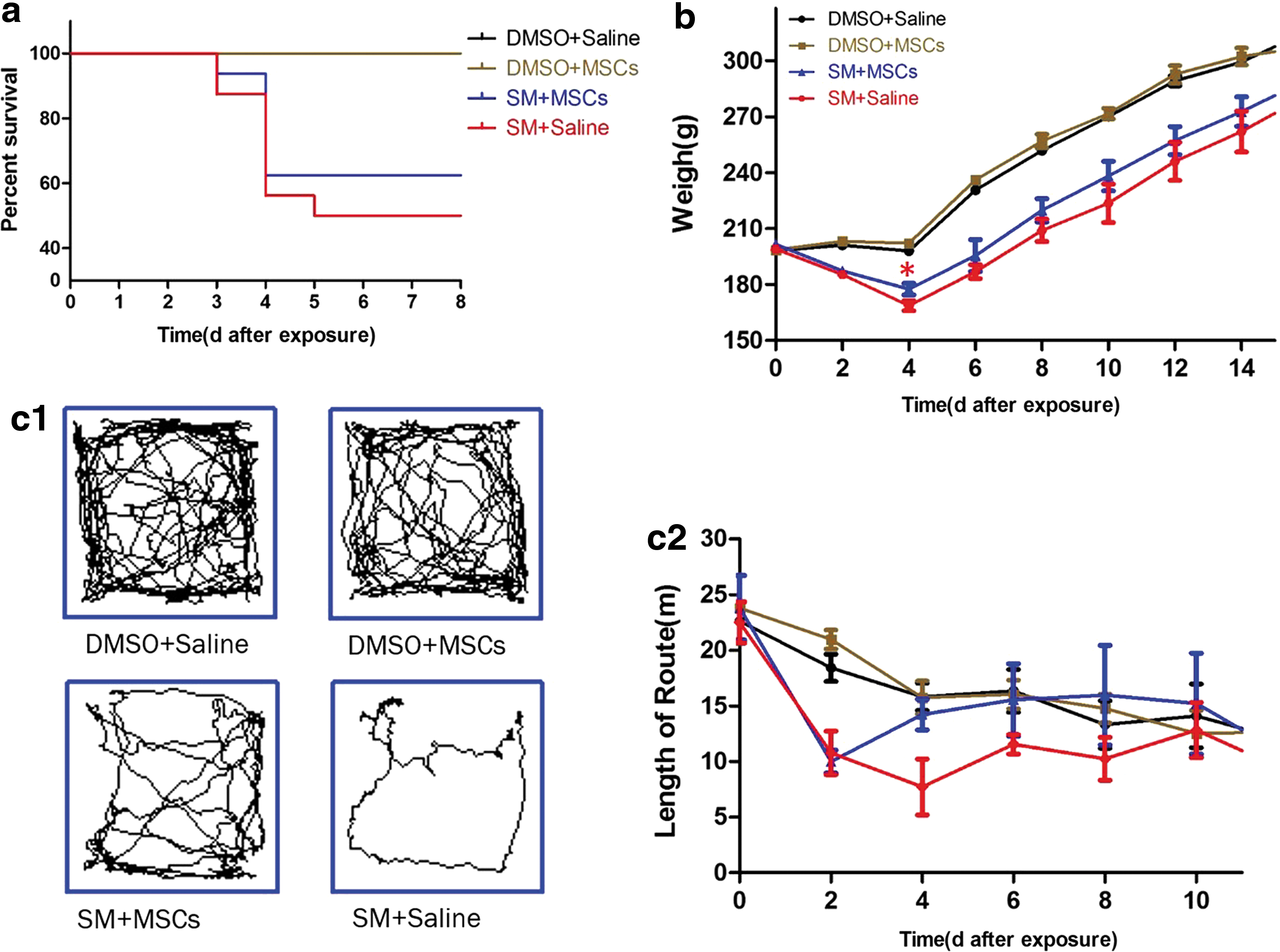
Discussion
The primary aim of this study was to investigate the potential beneficial effects of MSCs on SM exposure injury. In this study, by using a rat model of SM exposure injury, we demonstrated that the administration of MSCs improved body weight and locomotion, indicating that MSCs had positive effects on SM-exposed rats.
Similar to the symptoms of radiation exposure, SM exposure caused severe myelosuppression in our experimental model. At 4 days after exposure, karyocyte count in the bone marrow decreased significantly, while erythrocytes were maintained as shown by cytometry and the H&E staining of sternum sections. Moreover, the white pulp area of the spleen in the SM and MSC + SM groups observably decreased at 4 days. At the same time, leukocyte count in the venous blood of SM-exposed rats also dropped rapidly and began to rise from 4 days onward. Many clinical trials have demonstrated that MSC-treated subjects experienced rapid recovery in total leukocyte count [10,13,14], and the results of our experiments support this notion. More karyocytes were observed in the H&E-stained sternum sections at 4 days in the MSC + SM group, and WBC in the venous blood of rats in the MSC + SM group was restored faster and to higher levels than those in the SM group, especially at 10 days. To explain this phenomenon, we detected the cytokine levels of G-CSF, GM-CSF, MCP-1, IL-1α, IL-5, and IFN-γ, which are involved in immunoregulation and hematogenesis in the serum of all rats at 4, 7, 10, and 14 days after SM exposure, and our results suggested that MSCs could promote the increase in cytokine levels in serum. There are several studies that have indicated that MSC injections could induce significant elevations in serum G-CSF and GM-CSF levels, which are involved in cell proliferation, immunoregulation, tissue repair [22], and regeneration [23,24]. Additionally, IL-1α, described as the first proinflammatory cytokine, can be produced after stimulation by many kinds of cells such as macrophages, keratinocytes, monocytes, myeloid cells, and natural killer cells. IL-1α performs multiple functions as follows: (i) as an intracellular modulator of transcription, (ii) as a local immune adjuvant in its membrane-associated form on myeloid cells, and (iii) as a secreted, extracellular alarmin that can act in a long-range fashion to impact innate and adaptive immune responses [25,26]. In recent years, many published investigations have shown that IL-1α can induce the production of IL-1α, G-CSF, and GM-CSF by MSCs [13,23], which may explain why IL-1α, G-CSF, and GM-CSF levels in the MSC + SM group were higher than those in the SM group in our experimental model. Moreover, many investigations have demonstrated that IL-1α and MCP-1 are critical for leukocyte recruitment and homing to sites of injury. In fact, MSCs can themselves migrate toward MCP-1 [13,27]. In this study, the level of MCP-1 also increased after MSC transplantation at 10 days, and this may help MSCs and other immunological cells migrate to the sites of injury to promote tissue healing. IL-5 is known as an important cytokine that can stimulate eosinophil proliferation, which is a major type of leukocyte [28]. IFN-γ orchestrates leukocyte attraction and directs growth, maturation, and differentiation of many cell types in addition to enhancing natural killer cell activity and regulating B cell functions [29]. In our study, the levels of both IL-5 and IFN-γ increased to a certain degree at different times after MSC transplantation. High levels of IL-5 and IFN-γ indicate that MSCs may be involved in leukocyte generation and immunoregulation. In either case, immunoregulation of MSCs in the rat model with percutaneous exposure to SM may be achieved through paracrine interactions.
Based on our results, the BPC and RBC in the SM and MSC + SM groups decreased at 7 days. Although BPC in the SM group was higher than that in the MSC + SM group at 4 days, the valley point in the MSC + SM group was higher at 7 days, suggesting that rats in the MSC + SM group suffered less damage to the hematopoietic system than did rats in the SM group. Consistent with the change in the BPC of the SM and MSC + SM groups at 10 and 14 days, the red pulp of the spleen in the SM and MSC + SM groups was more hyperplastic than that in the control group, and the relative spleen weight in the SM and MSC + SM groups was significantly higher than that in the control group. Additionally, the level of EPO in the serum of rats in the MSC + SM group was increased at 7 and 10 days; similarly, the levels of G-CSF and GM-CSF were also increased at 7 days, which may be related to the increase in the BPC and hyperplasia of spleen. As shown in this study, MSCs may enhance the hyperplasia of the red pulp and decrease apoptosis in the spleen to support the reconstruction of the hematopoietic system by regulating the level of EPO in serum. In addition, the change in the number of CD34-positive cells in the spleen and bone marrow may suggest that MSCs help the spleen activate more quiescent CD34 cells to compensate for hemopoiesis weakened by the subsequent myelosuppression that occurred on and after 4 days. Similar to the spleen, the liver has some hematopoietic functions, and thus, we examined the histopathology of the liver (Supplementary Fig. S2). However, we observed no obvious injury to the liver similar to the spleen from SM in our experimental model, and MSC therapy exerted no significant effect on it.
The mechanism of SM exposure injury remains unclear, but the main hypothesis is that SM reacts with cellular macromolecules to lead to toxic effects, especially DNA alkylation, which is responsible for triggering apoptosis [4 –7]. Therefore, in this study, we observed that the rapidly dividing cells such as hematopoietic stem cells appeared to be more noticeably affected by SM. Furthermore, SM led to a significant decrease in the number of bone marrow karyocytes and severe myelosuppression in between 3 and 5 days after exposure. The turnover time of karyocytes in the bone marrow is approximately 5–7 days, during which blood cells are differentiated from hematopoietic stem cells. Our results thus indicated that, to some extent, stem cell injury leads to a reduction in the number of bone marrow and blood karyocytes. Because both in vitro and in vivo studies support the role of MSCs in promoting the restoration of the intestinal epithelium and small intestinal villi via the nurturing of the growth of residual surviving crypt stem cells [10,13,30 –32], whose mechanism might be that MSCs enhance local secretion of growth factors and chemotactic signals that attract progenitor cells to sites of tissue damage, we think MSCs might play a role in protecting or replacing stem cells in bone marrow and blood; however, further studies are needed to support this view.
In summary, we have demonstrated that the injection of human MSCs via the tail vein exerted significant therapeutic effects in a rat model of percutaneous exposure to SM through immunoregulation and improvement of the hemopoietic microenvironment. To the best of our knowledge, this is the first experimental animal model of rats with percutaneous exposure to SM that evaluates the efficacy of human MSC therapy for SM exposure. This study may provide preclinical data for the treatment of SM exposure or myelosuppression using human MSCs.
Footnotes
Acknowledgments
The authors thank Zhuang Wang and Ping Sun from Ivy Institute of Stem Cells Co. Ltd. (Beijing, China) for isolation, culture, and identification of human UC-MSCs.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.