
Abstract
Mesenchymal stem cells (MSCs) are a promising therapeutic tool for the treatment of nonhealing diabetic wounds. The pathological nature of the niche microenvironment limits the use of autologous cell therapy in diabetic patients. Prolonged exposure of endogenous MSCs to a pathological microenvironment in vivo reduces their ability to respond to environmental cues. This study investigated the effectiveness of ex vivo antioxidant treatment [N-acetylcysteine (7.5 mM NAC) and Ascorbic acid 2-phosphate (0.6 mM AAP)] to restore the paracrine function of diabetic MSCs. Healthy control [bone marrow stem cells derived from wild-type mice (SCWT)] (source: wild-type C57BL/6J mice) (n = 12) and impaired/dysfunctional [bone marrow stem cells derived from ob/ob mice (SCob)] (source: obese diabetic, B6.Cg-Lepob/J mice) (n = 12) MSCs were isolated. Ex vivo treatment groups (SCWT vs. SCob) were as follows: (1) no treatment (baseline phenotype), (2) stimulated with diabetic wound fluid (DWF) (baseline response), (3) antioxidant preconditioning (preconditioned phenotype), and (4) antioxidant preconditioned with subsequent stimulation with DWF (preconditioned response). The paracrine responsiveness on both the molecular (mRNA expression of 80 cytokines and receptors, quantitative polymerase chain reaction microarray) and protein (23-plex bead-array Luminex assay) level was assessed. At baseline, 31 genes were overexpressed (> × 2-fold) and 39 genes were underexpressed (> × 2-fold) in SCob versus SCWT. In conditioned media, significant differences (P < 0.05) were detected at baseline for two proinflammatory cytokines [tumor necrosis factor alpha (TNFα) and interferon gamma (IFNγ)], four chemokines [keratinocyte chemoattractant (KC), granulocyte colony-stimulating factor (GCSF), Eotaxin, and macrophage chemoattractant protein (MCP1)], and one anti-inflammatory cytokine [interleukin 10 (IL10)]. Following stimulation with DWF, significant differences (P < 0.05) were detected in the secretion of two chemokines [granulocyte macrophage colony-stimulating factor (GMCSF) and Eotaxin], three proinflammatory cytokines (TNFα, IFNγ, and IL9), and four anti-inflammatory cytokines (IL10, IL4, IL13, and IL3). Antioxidant preconditioning significantly dampened the excessive TNFα response observed in SCob and improved the secretion of IL10. Taken together these data suggest that the combined ex vivo treatment of autologous stem cells with NAC and AAP could potentially be an effective strategy to restore the paracrine function of impaired diabetic MSCs before transplantation.
Introduction
O
Mesenchymal stem cell (MSC) therapies aimed at improving wound healing are widely pursued in preclinical trials. This is due to the immune-modulatory, growth-promoting, and anti-inflammatory properties that MSCs possess [3,5 –7]. However, in this respect, the use of autologous stem cells from patients with DM are not as effective in promoting healing as those from nondiabetic controls [8]. Based on the findings from various in vitro studies, it is now known that DM alters the characteristics of endogenous MSCs to such an extent that the changes remain evident, despite removal of these cells to a more optimal in vitro environment [8,9]. Bone marrow stem cells derived from a DM microenvironment have impaired viability, have refractory mobility, and are unable to close an in vitro wound effectively [9 –12]. The mechanism underlying these impairments is, however, not yet clear. Furthermore, it is unknown if these impairments can be reversed using in vitro treatments, and if so, whether preconditioned MSCs will retain a repaired status when reintroduced to the pathological microenvironment in vivo.
Wound healing is a complex process that consists of four main phases: hemostasis, inflammation (destructive/phagocytic), proliferation (extracellular matrix formation, angiogenesis, and reepithelialization), and remodeling (matrix reorganization) [13,14]. The inflammatory phase is essential for the phagocytosis of bacteria, dead cells, and tissue debris. This phase is characterized by immune cell (neutrophil/M1 macrophage) infiltration and the presence of proinflammatory signaling molecules [15]. If the transition from M1 (proinflammatory) to M2 (proregenerative) macrophages does not occur timeously, healing is impaired, and the wound may become chronic. This is the case with diabetic ulcers, where a combination of factors, including an excessive and delayed inflammatory response, dysfunctional supporting cells (fibroblasts, keratinocytes, and epithelial cells), ischemia, impaired angiogenesis, and neuropathy, prevents the wound from transitioning into the proliferative and remodeling phases [5,15,16]. The paracrine properties of exogenously applied allogeneic (healthy) bone marrow stem cells have been shown to restore the wound microenvironment by inducing the proliferative and remodeling phases of healing through anti-inflammatory and proregenerative signaling [7]. Bone marrow stem cells isolated from pathological environments (such as those existing during DM) lose their ability to promote repair. We hypothesize that prolonged exposure of autologous MSCs to a pathological microenvironment in vivo reduces the ability of these cells to respond to environmental cues and thereby effect repair. Assessing whether the paracrine (cytokine) responsiveness of MSCs is skewed toward a prodestructive or proregenerative response could potentially be used as diagnostic parameters to predict the degree of MSC dysfunction in a diabetic patient.
The pathogenesis of DM, which contributes to the functional decline of stem cells, is driven by hyperglycemia (advanced glycation end products), chronic inflammation, and oxidative stress [8]. Ascorbic acid 2-phosphate (AAP) and N-acetylcysteine (NAC) are antioxidants that have been shown to preserve the “stemness” of healthy MSCs during prolonged culture, by preventing apoptosis and promoting proliferation, telomerase activity, and increasing intracellular levels of antioxidant enzymes such as superoxide dismutase [17 –20]. It is hypothesized that these protective agents could potentially be used to restore the function of impaired autologous stem cells.
This study therefore investigated whether in vitro antioxidant preconditioning could restore the paracrine function of impaired diabetic bone marrow stem cells. To achieve this, an in vivo murine model of impaired wound healing was established, and diabetic wound fluid collected. The paracrine (secretome; extracellular) and molecular (intracellular) responsiveness of healthy and impaired/dysfunctional stem cells were then assessed at baseline and following ex vivo exposure to diabetic wound fluid. The successful restoration of the paracrine function of impaired/diabetic stem cells ex vivo before use for autologous cell therapy could improve the predictability of cell therapy in diabetic ulcer patients in the clinical setting.
Materials and Methods
This study was approved by the animal research ethics committee (SU-ACUD17-000016) at Stellenbosch University and complied with the South African Animal Protection Act (Act no. 71, 1962). All experimental procedures were conducted according to the ethical guidelines and principles of the declaration of Helsinki.
Induction of full-thickness wounds and collection/analysis of wound fluid
Full-thickness excisional wounds were induced dorsally on obese prediabetic mice (B6.Cg-Lepob/J) (ob/ob) with subcutaneous injections of recombinant neutral endopeptidase (NEP) to (1) prevent wound contraction and (2) more closely mimic diabetic ulcers in patients [21,22].
Seven (n = 7) ob/ob mice (8-week old; weight 43 ± 2 g) were anesthetized using isoflurane (Safeline Pharmaceuticals Pty, Ltd., SA) gas. Dorsal hair was shaved, washed with povidone-iodine and a local anesthetic, and 2% lignocaine (7 mg/kg; Bodene Pty, Ltd., SA) applied before making two identical contralateral full-thickness skin excisions (including the underlying panniculus carnosus) (6 mm2) (Fig. 1). Immediately postwounding and on day 7, NEP (0.3 mg/mL; enzyme activity 50 pmol/h/μg) (SRP6450, Neprilysin/CD10; Sigma-Aldrich, St. Louis, MO) was injected subcutaneously at the wound edges to prevent skin contraction and further delay healing. The enzyme activity of the injected NEP is equivalent to that observed in the foot ulcers of diabetic patients [21,22]. Postsurgery pain (3 days) was managed by adding acetaminophen (300 mg/kg) (GSK, Cape Town, SA) to the drinking water of individually housed animals. Both excisional wounds were covered by a vapor-permeable polyurethane film (Hydrofilm; Paul Hartmann AG, Heidenheim, Germany) and the accumulated exudate (diabetic wound fluid, DWF) harvested by needle puncture [23,24] at 7, 14, and 21 days postwounding. At these time points, DWF was collected by injecting 100 μL sterile saline onto the wounds and then collecting the wound exudate together with the saline. The collected DWF was pooled, sterile filtered, and centrifuged to eliminate all debris and stored at −80°C. Note: on day 14 postwounding, the start of reepithelialization was evident, and wounds were therefore reopened (by removing the epithelial layer) at this time point, following the collection of the DWF.

Full-thickness excisional diabetic wound model.
The total protein concentration of the sterile filtered DWF (9.6 μg/mL) was determined to be threefold less than the total amount of protein in fetal bovine serum (FBS) (27.9 μg/mL) using a standard Bradford assay with BSA (Hyclone BSA; Thermo, Logan) as control (Supplementary Fig. S1; Supplementary Data are available online at
As a parallel control, to assess whether wound closure was truly delayed in our diabetic wound model, acute wounds (without NEP injection) were induced on six (n = 6) healthy control mice (C57BL6/J) (8-week old; weight 25 ± 0.2 g) using the same procedure as described above. Complete macroscopic wound closure and significant healing were evident in the control animals on day 7 (Supplementary Fig. S2).
Determination of the safest, nontoxic doses of NAC and AAP for in vitro antioxidant preconditioning
The dose-response experiment was performed using a mouse MSC line (C3H/10T1/2, clone 8, ATCC ccl-226) (passage 5–7) as part of the refinement criteria of this study to limit the required number of animals. MSCs were cultured in standard growth media (SGM) consisting of DMEM with ultra-glutamine (4.5 g/L high-glucose DMEM; BioWittaker, Lonza, Basel, Switzerland) containing 10% FBS (Biochrom, Berlin, Germany) and 1% penicillin/streptomycin (BioWittaker, Lonza). Cells were maintained at 37°C, in 90% humidified air with 5% CO2. The effect of different doses of NAC (7.5, 15, and 30 mM) (Sigma-Aldrich, St. Louis, MO) or AAP (0.15, 0.3, 0.6 mM) (Sigma-Aldrich, St. Louis, MO) on MSC proliferation (BrdU ELISA; Roche, Basel, Switzerland) was assessed over a period of 24 and 48 h, respectively. This was done to determine the optimum dose of each individual antioxidant. To assess the synergistic effect of the optimum doses, the survival rate of MSCs following exposure to the combined antioxidant treatment was determined over a period of 6 days using Crystal Violet staining (0.01% in dH2O) (Sigma-Aldrich, Steinheim, Germany) [25].
Isolation and characterization of bone marrow-derived SCs
Healthy control (source: wild type, C57BL/6J mice, weight 23.6 ± 0.9 g) (n = 12) and impaired/dysfunctional bone marrow-derived SCs (source: obese diabetic, B6.Cg-Lepob/J mice, weight 37.2 ± 1.7 g) (n = 12) were isolated as previously described [9]. Briefly, following sacrifice of the animals, the femurs were dissected out and the proximal and distal ends of each femur cut open in a sterile environment. Bone marrow aspirates were flushed into tissue culture plates (six-well plates; Nest Biotechnology, NJ) using a 25-gauge needle and syringe containing growth media with 20% FBS (Biochrom). Growth media consisted of DMEM with ultra-glutamine (4.5 g/L high glucose; BioWittaker, Lonza), containing 1% penicillin/streptomycin (BioWittaker, Lonza). Bone marrow aspirates were maintained at 37°C, in 90% humidified air with 5% CO2. After 96 h, nonadherent cells were washed off with phosphate-buffered saline (PBS) and the media replaced with SGM containing 10% FBS (Biochrom). Media were then changed every 3–4 days until the cells reached confluence in passage 0. All subsequent experiments were performed on SCs derived from at least three animals per group (n = 3 biological repeats).
SCs (90% confluent; passage 0) were harvested through trypsinization and resuspended in flow cytometry staining solution (PBS containing 20% FBS). Cell suspensions at a concentration of 1 × 106 cells per 100 μL were co-labeled with rat anti-mouse monoclonal antibodies against the stem cell marker, Ly-6A/E (Sca-1) (PE conjugated; BD Pharmingen, San Diego, CA) and the hematopoietic lineage marker, CD45 (FITC conjugated; BD Pharmingen). Flow cytometry was performed on a BD FACSCalibur instrument using CellQuest software. A total of 15,000 events were recorded before data analysis. Fluorescent compensation settings were established through a compensation experiment using comp beads (BD™CompBead Plus Anti-Mouse Ig, k; BD Biosciences, San Jose, CA). An unstained control sample was used as a negative control for gating purposes. Data analysis was performed using Flow Jo Vx (Treestar, OR) software.
For all subsequent experiments, the isolated bone marrow SCs were expanded in culture under either standard growth media (control) conditions or in the presence of antioxidant preconditioning before stimulation with DWF.
In vitro treatment conditions and study design
Antioxidant preconditioning consisted of combined treatment with NAC (7.5 mM) (Sigma-Aldrich, St. Louis, MO) and AAP (0.6 mM) (Sigma-Aldrich, St. Louis, MO) added to the growth media (SGM), with media being changed every 3–4 days until the cells reached 80% confluence. The inflammatory phenotype and paracrine responsiveness of healthy versus impaired SCs (with and without preconditioning) to environmental cues were assessed following ex vivo exposure to DWF. For the treatment of SCs with DWF, the FBS (10%) in the growth media was replaced by DWF (30%) with an equivalent total protein concentration. The treatment groups were as follows: (1) no treatment (baseline phenotype): SCs expanded in standard growth media (±8 days) and only exposed to growth media, (2) DWF: SCs expanded in standard growth media (±8 days) followed by acute exposure to DWF for a period of 4 h, (3) antioxidant preconditioning: SCs expanded in the presence of NAC/AAP (±8 days), And (4) antioxidant preconditioning+DWF: SCs expanded in the presence of NAC/AAP (±8 days) followed by acute exposure to DWF for a period of 4 h. Refer to Supplementary Fig. S3 for an overview of the study design and treatment groups.
Collection and analysis of conditioned media
After MSCs reached 80% confluence, the growth media (with or without NAC/AAP) were replaced with serum-free media for a period of 24 h. The complete serum-free growth media consists of Stem Pro MSC SFM basal media (No. A10334-01; Gibco, Life Technologies), Stem Pro MSC SFM growth factor Supplement CTS (A10333-01; Gibco, Life Technologies), 2 mM
RNA isolation, cDNA synthesis, and RT2 quantitative polymerase chain reaction microarray
Total RNA was isolated using the RNeasy RNA isolation kit (Qiagen, Berlin, Germany) with on-column DNAse digestion (Qiagen). RNA quantification and quality control were performed using the Eukaryote Total RNA Nano Series II (2100 Bioanalyzer; Agilent Technologies, Inc., Santa Clara, CA). Total RNA (0.5 μg) with an integrity of above 9 (RIN >9) was used as a template for cDNA synthesis, using the RT2 First Strand Kit (Qiagen) that includes a genomic DNA elimination step. cDNA samples for each treatment condition were analyzed with an RT2 Profiler polymerase chain reaction (PCR) inflammatory cytokine and receptor array (96-well plate format; Qiagen) and RT2 SYBR Green Master-mix (Rox PCR Master mix; Qiagen) using an ABI 7900HT Fast real-time PCR system (Applied Biosystems, Life technologies), and SDS software (version 2.3; Life technologies). The web-based PCR Profiler Array data analysis package was used for ΔΔCt-based fold change calculations (Qiagen,
Statistical analysis
Values are presented as mean ± standard error (mean ± SE). Statistical analysis was performed using Statistica software (Statsoft, version 13). Repeated-measures ANOVA with Bonferroni post hoc test was used to assess animal weight and wound closure over time. Factorial ANOVA with Tukey post hoc test was used to assess group, treatment, and group × treatment effects. Genes of interest were identified using a Student's t-test of the replicate 2(ΔCt) values of each gene in the control and treatment groups. The level of significance was accepted at P < 0.05.
Results
Full-thickness excisional diabetic wound model: collected wound fluid is representative of the in vivo microenvironment of an inflammatory diabetic wound
No weight loss was apparent in the wounded animals (start weight: 43 ± 2 g; end weight: 41 ± 7 g) for the duration of the study (Fig. 1A). Impaired wound closure (macroscopic) was evident in the ob/ob animals over the initial recovery period of 14 days (day 0 to 7: 33% ± 7% wound closure; day 0 to 14: 77% ± 4% wound closure) as well as in the subsequent period after rewounding (day 14 to 21: 48% ± 7% wound closure) (Fig. 1B, C). In contrast, significant wound closure was evident after only 7 days in the WT control animals (Supplementary Fig. S1). Wound fluid was collected and analyzed as indication of the clinical condition of the wound. The DWF collected from the ob/ob animals had a cytokine profile that mimics the clinical characteristics of a nonhealing diabetic wound. Refer to Table 1 for the quantification of cytokine concentrations detected in the DWF. Proinflammatory cytokines (IL1α, IL1β, IL6, IL12p40, IL12p70, IL17A, RANTES, and TNFα) and chemokines (Eotaxin, GCSF, GMCSF, MCP1, MIP1α, and MIP1β) were present in high concentrations, whereas anti-inflammatory cytokines (IL2, IL3, IL4, IL5, IL9, and IL10) had much lower levels within the DWF.
Cytokine Secretion Profile in Response to Diabetic Wound Fluid
Data are presented as mean (SE) in pg/mL. Statistical analysis: factorial ANOVA with Tukey post hoc test. Level of significance accepted at P < 0.05.
Significant difference between SCWT and SCob with preconditioning.
Significant difference in SCob with and without preconditioning.
Significant difference between SCWT and SCob. c *Indicates P = 0.06.
The concentration of these chemokines was above the detection limit as indicated.
GCSF, granulocyte colony-stimulating factor; GMCSF, granulocyte macrophage colony-stimulating factor; IFNγ, interferon gamma; IL, interleukin; KC, keratinocyte chemoattractant; MCP, macrophage chemoattractant protein; MIP, macrophage inflammatory protein; SCob, bone marrow stem cells derived from ob/ob mice; SCWT, bone marrow stem cells derived from wild-type mice; TNFα, tumor necrosis factor alpha.
The safest nontoxic doses of NAC and AAP for in vitro antioxidant preconditioning
Compared to standard growth conditions, of all the concentrations tested (NAC 7.5, 15, and 30 mM; AAP 0.15, 0.3, and 0.6 mM), 7.5 mM NAC had the most significant effect on the proliferation rate of MSCs over a period of 24 h (∼2-fold increase) (P < 0.05) and 48 h (∼2.5-fold increase) (P < 0.05) (Fig. 2A, B). Similarly, 0.6 mM AAP was shown to promote MSC proliferation over a period of 24 h (∼1.3-fold increase) compared to standard growth conditions (Fig. 2A, B). The combined treatments of 7.5 mM NAC and 0.6 mM AAP maintained cell viability over a period of 6 days (Fig. 2C) and were thus identified as the optimum concentrations for preconditioning.

Safest, nontoxic dosages of NAC and AAP.
Characterization of healthy control (source: wild type, C57BL/6J mice) and impaired/dysfunctional (source: obese diabetic, B6.Cg-Lepob/J mice) bone marrow-derived SCs
Flow cytometry surface marker characterization demonstrated that 88% ± 4% of isolated bone marrow-derived cells expressed the stem cell marker, Sca1 (Fig. 3A–C). A percentage of cells did, however, also express the hematopoietic lineage marker, CD45. Immediately, following isolation (passage 0), the bone marrow-derived SCs therefore consisted of a heterogenous population of 60% ± 3% MSCs (Sca1+CD45−) and 39% ± 3% hematopoietic stem cells (Sca1+CD45+) (Fig. 3A–D). There was no difference in surface marker expression in the bone marrow SCs derived from either WT control (SCWT) or ob/ob (SCob) animals (Fig. 3D). SCob had impaired viability and growth with fewer cells visible 96 h postisolation compared to their SCWT counterparts at the same time point (Fig. 3E, F).
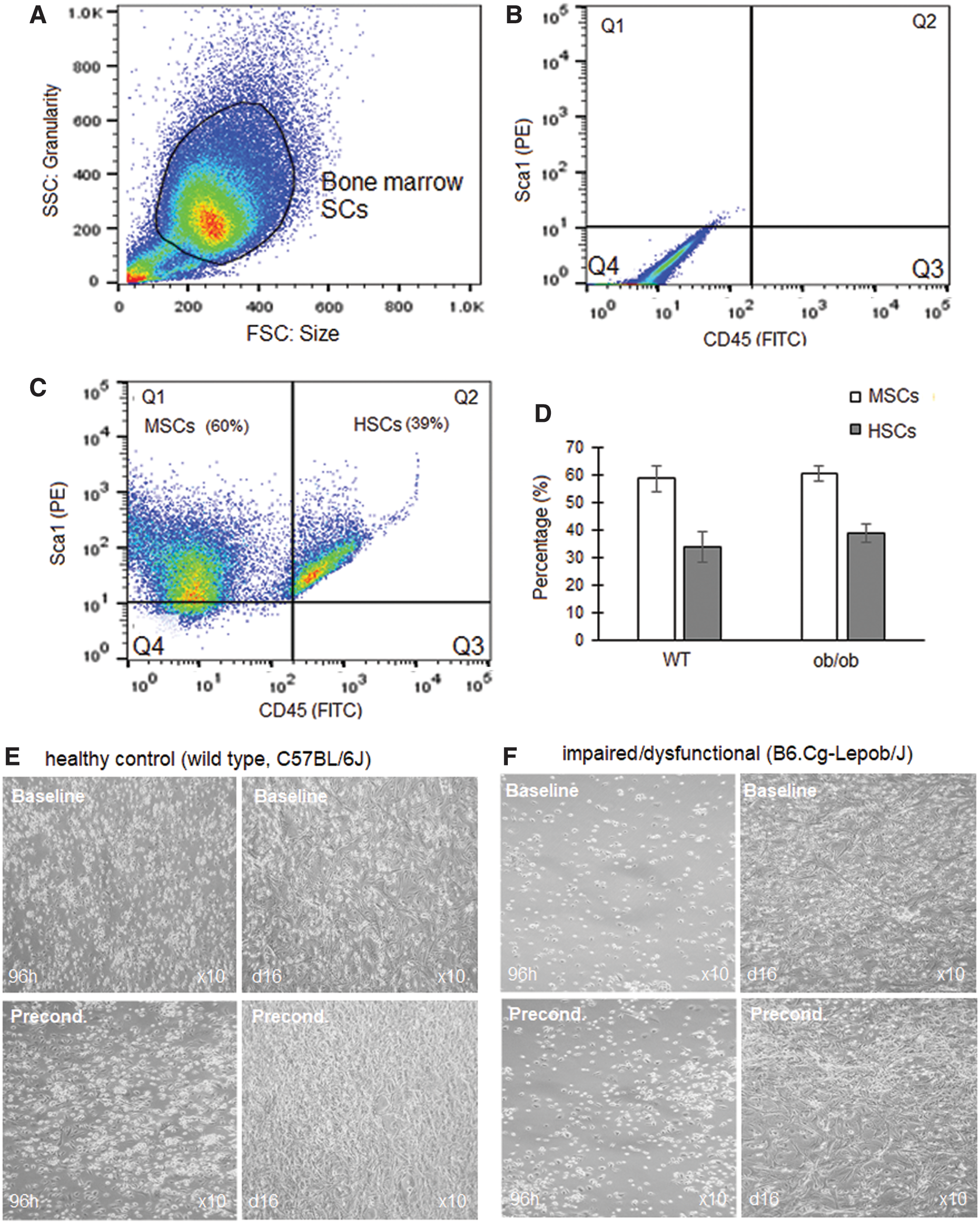
Characterization of healthy control (source: wild type, C57BL/6J mice) and impaired/dysfunctional (source: obese diabetic, B6.Cg-Lepob/J mice) bone marrow-derived SCs.
Secretome differences at baseline: healthy control (SCWT) versus impaired/dysfunctional (SCob)
The secretion of 23 cytokines from SCWT and SCob was assessed at baseline with and without antioxidant preconditioning (Table 2). Of the 23 cytokines assessed within conditioned media, significant differences (P < 0.05) were detected at baseline (without stimulation) for two proinflammatory cytokines (TNFα and IFNγ), four chemokines (KC, GCSF, Eotaxin, and MCP1), and one anti-inflammatory cytokine (IL10).
Cytokine Secretome Differences at Baseline
Data are presented as mean (SE) in pg/mL. Statistical analysis: factorial ANOVA with Tukey post hoc test. Level of significance accepted at P < 0.05.
Significant difference at baseline (control) between SCWT and SCob. a *Indicates P = 0.06.
Significant difference in SCob with and without preconditioning.
Significant difference between SCWT and SCob with preconditioning.
—, not detected.
Proinflammatory TNFα and IFNγ concentrations were significantly higher (P < 0.05) in the conditioned media derived from SCob (TNFα 2.9-fold higher; IFNγ 15 ± 4 pg/mL) compared to SCWT (IFNγ not detected). Preconditioning SCob with antioxidants did, however, downregulate the secretion of TNFα and IFNγ to levels comparable to that of SCWT. The chemokines KC, GCSF, and Eotaxin remained significantly higher (P < 0.05) in the conditioned media derived from SCob (KC 2.2-fold higher; GCSF 6.7-fold higher; Eotaxin 20 ± 7 pg/mL) compared to SCWT (Eotaxin not detected), regardless of preconditioning. Preconditioning did, however, promote MCP1 secretion in SCob (2.5-fold increase) (P < 0.05), whereas only a small increase (1.5-fold, not significant) was evident in SCWT. The anti-inflammatory cytokine, IL10, tended to be higher (2.4-fold) (P = 0.06) in the conditioned media derived from SCob compared to SCWT and was unaffected by preconditioning.
Gene expression (mRNA) differences at baseline and in response to stimulation with DWF: healthy control (SCWT) versus impaired/dysfunctional (SCob)
At baseline, under standard growth conditions, 31 genes were overexpressed (>2-fold) and 39 genes underexpressed (>2-fold) in the SCob versus SCWT (Fig. 4A). The overexpression of colony-stimulating factor (Csf1) (also known as MCSF) (29-fold) was, however, the only significant difference (P < 0.05) between the groups at baseline (Fig. 4A, C). Overall, preconditioning of SCob downregulated the expression of 32 genes and upregulated the expression of 48 genes (Fig. 4B). The expression of Csf1 in SCob was downregulated with preconditioning to levels comparable to that observed in SCWT (Fig. 4C). Preconditioning induced the expression of Ccl8 (also known as MCP2) (P < 0.05), which was previously undetectable in SCob (Fig. 4B, D).
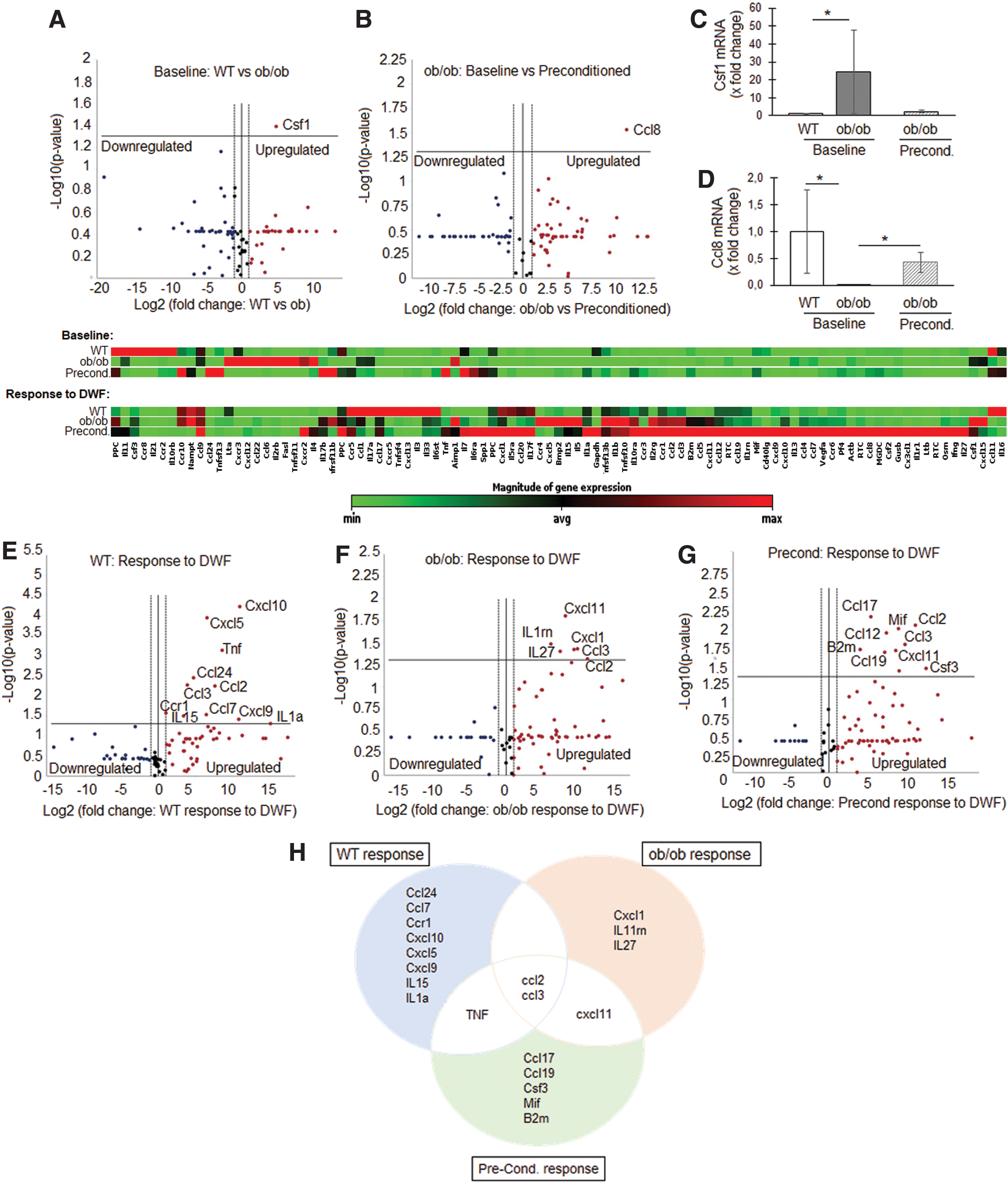
Gene expression differences at baseline and in response to stimulation with DWF: healthy control (SCWT) versus impaired/dysfunctional (SCob).
When exposed to DWF, both the SCWT and SCob responded by significantly (P < 0.05) increasing the gene expression of macrophage chemoattractant proteins, Ccl2 (also known as MCP1) and Ccl3 (also known as MIP1α) (Fig. 4E–H). In SCWT, DWF furthermore induced the expression (P < 0.05) of nine additional genes (Ccl24, Ccl7, Ccr1, Cxcl10, Cxcl5, Cxcl9, IL15, IL1α, and TNF) associated with proinflammatory chemotactic responses. In contrast, SCob responded by increasing (P < 0.05) the expression of only four additional genes (Cxcl1, IL11rn, IL27, and Cxcl11) (Fig. 4F, H). Preconditioning did, however, restore the TNF response (P < 0.05) and induced the expression of Ccl17, Ccl19, Csf3, macrophage inhibitory factor (Mif), and B2m (Fig. 4G, H) in SCob. A summary of the relevant molecular responses (P < 0.05) within these groups is indicated in Fig. 4H.
Differences in the paracrine responsiveness following DWF stimulation: healthy control (SCWT) versus impaired/dysfunctional (SCob)
Following stimulation with DWF, significant differences (P < 0.05) were detected in the conditioned media concentrations of two chemokines [GMCSF and Eotaxin (also known as ccl11)], three proinflammatory cytokines (TNFα, IFNγ, and IL9), and four anti-inflammatory cytokines (IL10, IL4, IL13, and IL3) between groups (SCWT vs. SCob vs. preconditioned SCob) (Fig. 5 and Table 1).
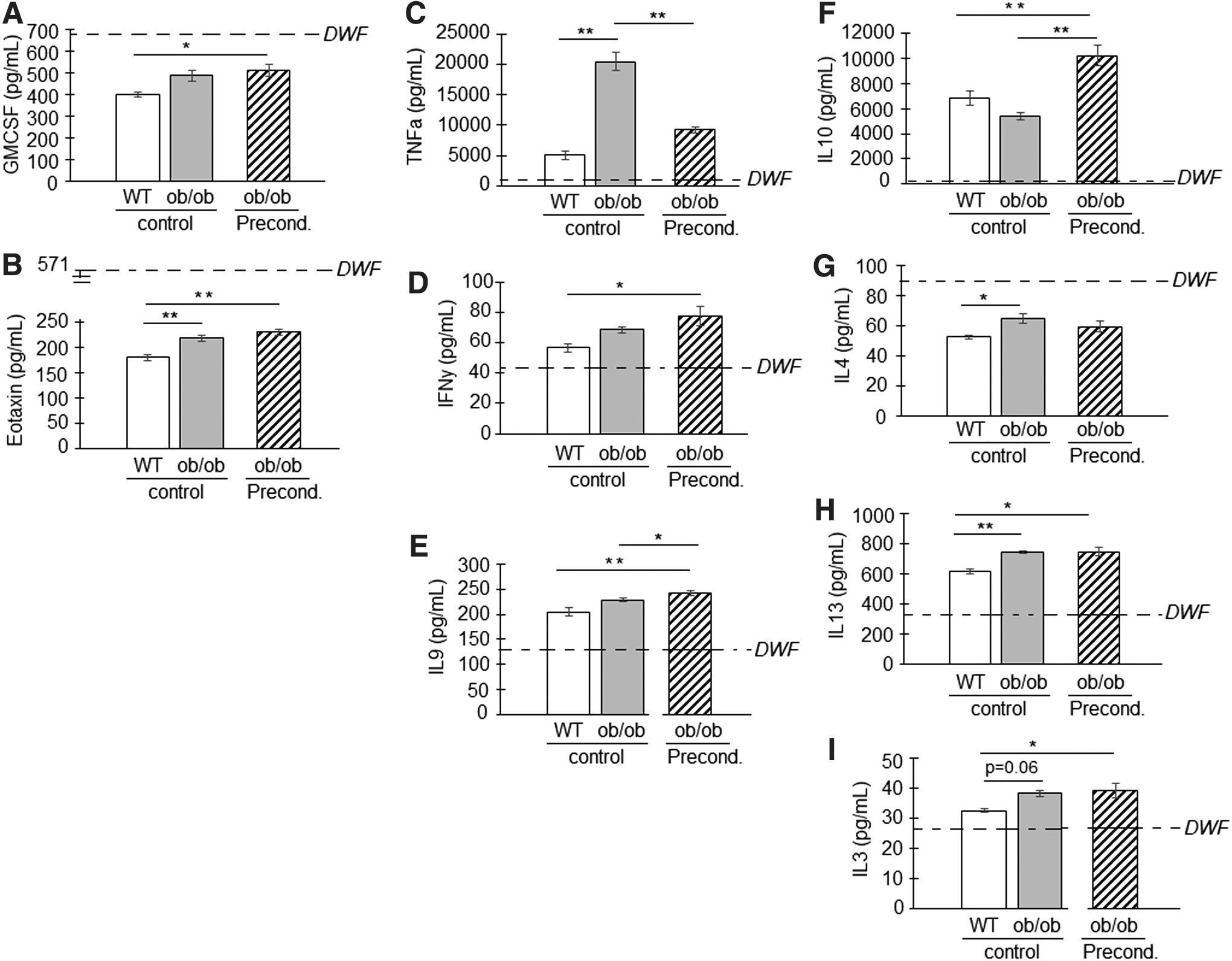
Differences in the paracrine responsiveness following DWF stimulation: healthy control (SCWT) versus impaired/dysfunctional (SCob). The cytokine concentrations in conditioned media poststimulation with DWF were assessed in SCWT, SCob, and preconditioned SCob.
Similar to baseline, the chemokines GMCSF and Eotaxin in conditioned media were detected at higher concentrations in SCob (GMCSF 486 ± 26 pg/mL and Eotaxin 217 ± 6 pg/mL) and preconditioned groups (GMCSF 510 ± 30 pg/mL and Eotaxin 229 ± 9 pg/mL), following stimulation compared to SCWT (GMCSF 399 ± 10 pg/mL and Eotaxin 179 ± 6 pg/mL) (Fig. 5A, B). Despite these differences, the concentration of these chemokines poststimulation was lower than that detected in DWF (Table 1).
Stimulation with DWF induced a much greater proinflammatory TNFα response in SCob (20,441 ± 1,461 pg/mL) than SCWT (5,081 ± 671 pg/mL); this excessive response in SCob was dampened with preconditioning (9,190 ± 487 pg/mL) (Fig. 5C). Preconditioning furthermore increased the release of proinflammatory IFNγ and IL9 following stimulation with DWF in the SCob (IFNγ 77 ± 6 pg/mL and IL9 242 ± 5 pg/mL) compared to SCWT (IFNγ 57 ± 3 pg/mL and IL9 204 ± 9 pg/mL) (Fig. 5D, E).
A slight difference in the anti-inflammatory IL3, IL4, and IL13 response to DWF was evident between SCob (IL3 38 ± 1 pg/mL; IL4 65 ± 3 pg/mL; and IL13 745 ± 7 pg/mL) and SCWT (IL3 33 ± 1 pg/mL; IL4 53 ± 1 pg/mL; and IL13 617 ± 14 pg/mL) (Fig. 5F–I). IL4 in conditioned media poststimulation was, however, below the concentration detected in DWF (Table 1). Preconditioning significantly improved the secretion of anti-inflammatory IL10 poststimulation in the SCob group (SCob 5,415 ± 317 pg/mL; Precond 10,207 ± 812 pg/mL; SCWT 6,823 ± 575 pg/mL) (Fig. 5F).
Discussion
Tissue repair and regeneration using endogenous stem cells represents the ultimate goal in regenerative medicine; however, the pathological nature of the in vivo microenvironment has hampered the use of autologous cell therapy in diabetic patients. This study demonstrated that bone marrow stem cells derived from an obese diabetic microenvironment have a skewed inflammatory phenotype; this may contribute to their inability to perform the imperative function of restoring the localized (wounded area) conditions needed for normal healing. Wound fluid is generated by enhanced capillary leakage and local activity of a variety of resident and migratory cell types that react to a breach in the skin barrier. The composition of wound fluid is therefore broadly assumed to reflect the clinical condition of a wound [26,27]. In the context of this ex vivo study, wound fluid therefore represented the extracellular fluid space within the microenvironment of a diabetic wound. Exposing healthy and impaired/dysfunctional bone marrow stem cells to DWF in vitro enabled an investigation into the paracrine responsiveness of these cells on both molecular and protein level. This study was furthermore the first to demonstrate the efficacy of antioxidant preconditioning in restoring the paracrine function of impaired/dysfunctional stem cells toward that seen under control nonpathological conditions.
Although the ob/ob animal model has been used in numerous studies investigating diabetes-related impaired wound healing [28 –31], the endogenous stem cells derived from these animals have not yet been characterized. At baseline, the secretion of proinflammatory cytokines (TNFα, IFNγ) and chemokines (KC, GCSF, and Eotaxin) was more pronounced in the impaired/dysfunctional bone marrow stem cells. These factors all function to amplify the first stage of a wound (inflammatory response), by recruiting neutrophils, lymphocytes, eosinophils, and keratinocytes to the wounded area [5,13,32]. At the mRNA level, a similar pattern was observed; however, only the overexpression of Csf1 was significant between healthy versus dysfunctional stem cells and corresponded with higher chemokine (GMCSF) levels detected in the conditioned media. These and other chemokines were already present within the wound microenvironment (DWF) at very high concentrations and the additional release thereof is thus unwanted. Antioxidant preconditioning was able to reduce the secretion of proinflammatory TNFα and IFNγ at baseline, but did not reduce chemokine secretion.
In the context of cell therapy, the paracrine responsiveness of stem cells to the wound microenvironment will determine their effectiveness in promoting healing [3,6,7]. The bone marrow stem cells (healthy and dysfunctional) responded by upregulating the gene expression and protein secretion of macrophage chemoattractants MCP1 (Ccl2) and MIP1α (Ccl3). In chronic diabetic wounds, a significant delay in macrophage infiltration related to a deficiency in ccl2 has been demonstrated [33]. Wood et al. [33] furthermore indicated that acute ccl2 treatment could restore the macrophage response and promote healing. This is supported by Yin et al. [34], indicating that the use of scaffolds that actively release MCP1 promoted macrophage infiltration and improved wound healing in a mouse model of streptozotocin-induced diabetes. MCP1 is also known to influence the effector state of macrophages [35], to mediate the angiogenic effect of transforming growth factor beta (TGFβ) by recruiting vascular smooth muscle cells to the endothelium [36], and to recruit/improve the migration of bone marrow MSCs [37]. In this study, preconditioning further enhanced the MCP1 response of all the bone marrow stem cells.
Dysfunctional bone marrow stem cells did, however, have an excessive TNFα response to DWF. TNFα is a potent proinflammatory cytokine that amplifies inflammation through NFκB signaling [38]. Persistent TNFα signaling is known to occur in diabetic wounds [39] and prevents the phenotype switch of macrophages form phagocytic (M1) to proregenerative (M2). Ashcroft et al. [40] demonstrated that targeting TNFα in a mouse model (secretory leukocyte protease inhibitor null mice) of severely impaired wound healing-blunted leukocyte recruitment reduced NFκB signaling, altered the M1/M2 macrophage balance, and improved healing. In this study, antioxidant preconditioning of dysfunctional bone marrow stem cells blunted the TNFα response and improved the secretion of anti-inflammatory IL10.
Taken together, these data suggest that the combined ex vivo treatment of autologous stem cells with NAC and AAP could be an effective strategy to restore the paracrine function of impaired bone marrow stem cells before transplantation. This study was, however, done with acute exposure to the pathological microenvironment and future studies should assess whether these cells will retain their restored ability over longer periods of exposure.
Footnotes
Acknowledgments
This research was supported by grants from the National Research Foundation (NRF) (grant no. 105921) and the Harry Crossley foundation. YMA received bursaries from the NRF and the Faculty of Medicine and Health Sciences, Stellenbosch University. We would like to thank Prof. K.H. Myburgh for her insight and role as mentor.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.