
Abstract
The peritoneum mesothelium lines body cavities and has the same origin as ovarian surface epithelium with probable existence of peritoneum mesenchymal stem cells (PMSCs). In the present research, PMSCs were isolated from peritoneum and differentiated into ovarian cell-like cells using human follicular fluid (HFF) and human cumulus-conditioned medium (HCCM). Anterior abdominal wall and intestinal peritoneum explants were used for cells isolation and cultured in Dulbecco's modified Eagle's medium. After passage 3, purified PMSCs were assessed for morphology, proliferation rate, and cell viability. Then, isolated PMSCs underwent two characterization procedures: (1) differentiation to mesodermal lineage and (2) expression of mesenchymal (CD90 and CD44) and epithelial cell (CK19) markers. The characterized PMSCs were differentiated into ovarian cell-like cells using 10% HFF and 50% HCCM for 21 days, and the expressions of oocyte (Zp3, Gdf9), germ cell (Ddx4, Dazl), granulosa cell (Amh), and theca cell (Lhr) markers were assessed using real-time polymerase chain reaction and immunocytofluorescence assay. Both anterior abdominal wall and intestinal peritoneum mesenchymal stem cells (AP-MSCs and IP-MSCs) showed mesenchymal characters and differentiated to adipocyte and osteocyte. AP-MSCs expressed mesenchymal- and epithelial cell-specific markers more than IP-MSCs and showed an analytically better proliferation rate. The induced AP-MSCs and IP-MSCs were expressed as germ and oocyte cell-specific markers, and this expression increased in the third week of culture. In both groups of AP-MSCs and IP-MSCs, the expressions of Gdf9, Zp3, Ddx4, Dazl, and Amh genes under just HCCM induction showed upregulation significantly on the 21st day of culture compared with day 0. But in protein synthesis of all mentioned genes, both HFF and HCCM had equal induction effect on the 21st day of culture against the 0th day. In addition, LHR was not expressed in any groups. Finally, in both characterization and differentiation procedures, the AP-MSCs respond to inducers better than IP-MSCs.
Introduction
It has been reported that in 30%–40% of couples who seek medical help for the problem of subfertility, the problem was predominantly in females, and in about 30% of cases, the reasons were found in both of them [1]. Female infertility is an assembled disorder and could be a result of a variety of conditions, such as premature ovarian failure, polycystic ovary syndrome, pelvic inflammatory disease, endometriosis, and uterine fibroids [2]. In recent years, there has been much progress in the area of assisted reproduction treatment, and advanced treatments have led to an 80% success rate of reproduction in previously infertile couples.
In the past decade, there has been interest in the use of mesenchymal stem cells (MSCs) for infertility treatment due to their unlimited source, self-renewal, high differentiation potential into mesodermal and non-mesodermal lineage [3], low risk of teratoma formation [4], low immunogenicity, and immunosuppressive effect [5 –7]. These types of cells can be derived from a variety of tissues, including bone marrow, adipose, Wharton's jelly, umbilical cord and peripheral blood, kidney, liver, brain, spinal cord, and mesothelium [8 –10]. The mesothelium has a monolayer of specialized cells known as mesothelial cells that line the pleural, pericardial, and peritoneum cavities and cover most internal organs. The peritoneum mesothelial consists of intraperitoneal or intestinal space, abdominal wall, and omentum. One of the characteristics of peritoneum mesothelial cells is the presence of growth factors (bFGF, VEGF), extracellular matrix (ECM) proteins, cytokines, antigens, and factors that protect from infection, all of which aid in tissue adhesion, prevention/treatment of inflammation, reconstruction, and repair of damage [11]. Additionally, they show many features of epithelial cells including polygonal cell shape, desmosome junction, and epithelial vimentin and cytokeratin filaments [12]. There is now increasing evidence that the peritoneum mesothelial tissue has functional cells with stem cell characteristics [13]. These stem cells are able to differentiate into osteocyte, adipocyte, myocyte, chondrocyte, and nervous cells [14,15]. Furthermore, these cells have fibroblastic-like phenotype, plastic adherence, self-renewal, passaging ability, and MSC surface markers and form colony-forming units (CFUs) in in vitro culture. Moreover, omentum grafts have been used in the treatment of neurodegenerative diseases such as Alzheimer [16]. The omentum's potential to rescue the injured tissues may be a result of its many neurotrophic and angiogenic properties [17].
This study investigated if anterior abdominal wall peritoneum mesenchymal stem cells (AP-MSCs) and intestinal peritoneum mesenchymal stem cells (IP-MSCs) can be differentiated into ovarian cell-like cells (oocyte, germ cell, granulosa, and theca cells) in an appropriate conditioned medium containing various factors, such as human follicular fluid (HFF) and human cumulus-conditioned medium (HCCM).
Materials and Methods
Isolation of peritoneum mesenchymal stem cells
All stages of this study are subjected to the rules of research ethics committee of the Royan Institute (
Cell viability assay
MSCs viability was tested using MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt] assay. AP-MSCs and IP-MSCs (2 × 104 cells in every well) were cultured into a 96-well plate in DMEM/F12. After incubation, the absorption rate was measured in 490 nm using an MTS protocol according to the company's orders after 4, 5, 6, and 7 culture days.
Growth curve
AP-MSCs and IP-MSCs were cultured to the same number (5 × 104) in the 25-cm2 flasks. The cell number was counted as follows. The cell suspension was diluted in DMEM/F12-free serum, 8–10 μL placed on a Neubauer chamber, and the total cells were counted under an inverted light microscope (Olympus CKX41).
Differentiation to osteocyte and adipocyte
AP-MSCs and IP-MSCs were cultured for 3–4 days in a four-well plate until 70%–80% confluency, cultured for 21 days with osteogenic and adipogenic media, and then fixed with 4% paraformaldehyde for 1 h. For detection of bone substrates, the fixed samples were incubated for 2 min with 1% Alizarin Red. For detection of lipid droplets, the samples were incubated for 15 min with 0.5% Oil Red O and observed using an inverted light microscope (Olympus CKX41). These cells were exposed in DMEM/F12 supplemented with 15% FBS, 1% Glutamax, 1% non-essential amino acids, 1% ITS, 1% penicillin/streptomycin as negative control for 21 days. Bone marrow mesenchymal stem cells (B-MSCs) were used as positive control.
Immunocytofluorescence assay for MSC markers
The cells were plated in four-well plate and cultured until 80%–90% confluency, washed thrice with PBS−, and fixed in 4% paraformaldehyde for 20 min at room temperature (RT). Next, the cells were washed with 0.05% PBS−-Tween (PBST). To block unspecific antibodies, the cells were incubated with 10% secondary host serum for 30 min at 37°C. For permeabilization, the samples were incubated with PBS− containing 0.1%–0.5% Triton X-100 for 10 min at RT. The cells were incubated with the primary antibodies (dissolved in 10% BSA and 10% secondary host serum), anti-CD44 (1:100, 550538; BD Pharmingen), anti-CD90 (1:100, 225; Abcam), and anti-CK19 (1:100, 25724; Santa Cruz) for overnight at 4°C. Then, the cells were washed with PBST and incubated with fluorescein isothiocyanate (FITC)-conjugated rabbit anti-rat (1:500, F1763; Sigma), goat anti-mouse (1:500, 6785; Abcam), or goat anti-rabbit (1:500, 6717; Abcam) dissolved in 5% BSA in dark for 1 h at RT. The cells were washed with PBST, incubated for 1–2 min in DAPI followed by three-time washing with PBST, and monitored using a fluorescence microscope (Olympus IX71). The cell numbers were also counted by the ImageJ software to measure the quantity of positive cells. Negative controls were performed by eliminating the primary antibodies. Mouse B-MSCs and skin epithelial cells (SCs) were used as positive control for MSC markers (CD44 and CD90) and epithelial cell marker (CK19), respectively.
HFF preparation
The HFF was procured from healthy women under in vitro fertilization (IVF), from the Royan IVF Center (Tehran, Iran). The HFF was centrifuged (4,500 rpm, RT, 8 min) to remove blood and granulosa cells. The clear supernatant was filtered (0.22-μm pore size). For complement system inactivation, the HFF was incubated for 30 min at 56°C and finally stored at −20°C until assays were performed.
HCCM preparation
Cumulus cells were procured from healthy women under IVF, from the Royan IVF Center (Tehran, Iran), and centrifuged at 1,500 rpm for 5 min. The cell plate was cultured with DMEM/F12 supplemented with 15% FBS, 1% penicillin and streptomycin, 1% Glutamax, 1% non-essential amino acid, and 1% ITS into 12-well plate and were incubated at 97% humidity, 21% O2, and 5% CO2 at 37°C. After 1 week, the cells reached 70%–80% confluence; the clear conditioned medium from primary culture was collected as HCCM and stored at −20°C every other day.
Differentiation induction into ovarian cells (oocyte and germ cell-like cells)
For the derivation and characterization of ovarian cells from peritoneum mesenchymal stem cells (PMSCs), purified AP-MSCs and IP-MSCs were trypsinized, and cell pellets were seeded in a 24-well adherent culture plates. Differentiation medium consisted of DMEM/F12 supplemented with 10% HFF or 50% HCCM, 5% FBS, 1% penicillin and streptomycin, 1% Glutamax, 1% non-essential amino acid, and 1% ITS and was incubated at 97% humidity, 21% O2, and 5% CO2 at 37°C for 21 days. Half of the medium was replaced with fresh medium every 3–4 days. The morphology of differentiated cells from AP-MSCs and IP-MSCs was examined, and images were captured using an inverted light microscope (Olympus CKX41) at 0, 14, and 21 days. Cells larger than 30 μm in diameter were counted. Finally, whole cells were analyzed by reverse transcription quantitative polymerase chain reaction (RT-qPCR) and immunocytofluorescence staining. AP-MSCs and IP-MSCs were cultured in DMEM/F12 supplemented with 15% FBS as negative control.
RT-qPCR analysis
Total RNA was extracted from PMSCs exposed to HFF and HCCM using TRIzol reagent following the manufacturer's directions. After homogenizing the sample with the TRIzol reagent (200 μL/5 × 105 cells) for 5 min, 50 μL chloroform was added and shacked for 5 min. The homogenate was centrifuged (12000 rcf, 4°C, 15 min) to separate a clear supernatant containing RNA in the upper aqueous layer. RNA was precipitated from the aqueous layer, and isopropanol was added it's the same volume and did up-down five to six times and then incubated for an overnight at 20°C. After centrifugation (12000 rcf, 4°C, 15 min), 1 mL of 70% ethanol was added and centrifuged again. RNA plate was solved in 20 μL DEPC water. Then, cDNA was synthesized using cDNA Synthesis Kit (TaKaRa). For quantitative polymerase chain reaction (qPCR), the primers are listed in Table 1. Finally, 4.3 μL of distilled water, 0.5 μL of each forward and reverse primers, 2.5 μL SYBR green, and 0.2 μL ROX were added to 2 μL cDNA (final volume 10 μL). Gene expression in whole cells was determined using the comparative threshold cycle number (2−ΔCt) method.
List of Primers used in qPCR
qPCR, qantitative polymerase chain reaction.
Immunocytofluorescence staining for ovarian cell markers
To confirm more PMSCs exposed to HFF and HCCM at the protein level, cells were washed thrice with PBS− and fixed in 4% paraformaldehyde (20 min, RT). Next, the cells were washed with 0.05% PBST and blocked in PBS− with 10% secondary host serum for 30 min at 37°C. To permeabilize, the cells were incubated for 10 min with PBS− containing 0.1%–0.5% Triton X-100. Then, the cells were incubated with primary antibodies (dissolved in 10% BSA and 10% secondary host serum), anti-GDF9 (1:100, 7407; Santa Cruz), anti-ZP3 (1:100, 48895; Abcam), anti-DDX4 (1:100, 13840; Abcam), anti-DAZL (1:100, 34139; Abcam), and AMH (1:100, 129593; Gentex) for overnight at 4°C. The cells were washed with PBST and incubated with FITC-conjugated rabbit anti-goat (1:500, 2777; Santa Cruz) or goat anti-rabbit (1:500, 6717; Abcam) dissolved in 5% BSA in dark (30 min at 37°C). Then, the cells were washed with PBST and incubated in DAPI for 1–2 min, followed by three times wash in PBST, and viewed using a fluorescence microscope (Olympus IX71). Finally, they were stored in dark at +4°C. The cell numbers were counted using the ImageJ software to measure the number of positive cells. Negative controls were performed by eliminating the primary antibodies.
PMSCs ex vivo culture
For further study and confirmation of differentiation potential of PMSCs, these cells were seeded into the decellularized ECM of the ovarian tissue. Briefly, 2 × 106 cells suspended in 1 mL of DMEM/F12 supplemented with 15% FBS, 1% non-essential amino acids, 1% Glutamax, 1% ITS, and 1% penicillin/streptomycin were injected with an insulin syringe into human ovarian-decellularized ECM pieces (5 × 5 × 1 mm3) and cultured in 12-well tissue culture plates incubated at 37°C in 5% CO2 for 1 week. Finally, PMSCs ability to produce DDX4 protein as germ cell marker was assessed by immunohistochemistry technique.
Statistical analysis
Data are presented as mean ± standard error of the mean. Statistical comparisons were calculated using GraphPad Prism 6.0 software. Analysis of variance using a multiple comparisons test was used to determine the statistical significance, followed by the Tukey test. A significant difference was considered at P-value of <0.05, 0.01, and 0.001 for tests. All experiments were performed in triplicate for technical and biological repetitions.
Results
In vitro culture and characterization of AP-MSCs and IP-MSCs
The peritoneum pieces were collected from the anterior abdominal wall and intestinal regions of 6- to 8-week-old mice. Primary cultures of AP-MSCs and IP-MSCs were established using the peritoneum anterior abdominal wall and intestinal tissue fragments, respectively, in 25-cm2 plastic flasks. Tissue fragments grew adherent to the flask, and AP-MSCs and IP-MSCs started to migrate out from the explants after 3–4 days. These cells exhibited spindle-like shapes along with CFUs and were morphologically similar to B-MSCs in DMEM supplemented with 15% FBS. A few different shape cells were dispersed among the homogenous cells. AP-MSCs and IP-MSCs proliferated over passage 15 by maintaining morphological characteristics. However, in addition to adherent culture, these cells formed spontaneous aggregates in stirred culture by spinner flask at high passages (Fig. 1A). The population doubling time of AP-MSCs and IP-MSCs were ∼37 and 45 h with P < 0.05 (Fig. 1B).

Characterization of mouse anterior AP-MSCs and IP-MSCs.
Our studies showed that AP-MSCs viability by measurement of the mitochondrial activity in the culture for subsequent days is similar to IP-MSCs. The mean viability rates of AP-MSCs and IP-MSCs gradually increased from day 4 to day 7 and significantly different on day 7 AP-MSCs and IP-MSCs from day 4 (P < 0.05), but AP-MSCs and IP-MSCs have not together with the significant difference (Fig. 1C). To characterize multipotent capacity of AP-MSCs and IP-MSCs in passage 3, their potential to differentiate into osteocyte and adipocyte was assessed. Morphological changes were detected corresponding to cells differentiation. However, B-MSCs were used as positive control (Fig. 2). As well as, multipotential capacity were showed by immunocytofluorescence for the expression of CD44 and CD90 MSC markers (Fig. 3A). As shown in Fig. 3B and C, ∼89.0% of AP-MSCs and 78.0% of IP-MSCs were positive for CD44 marker and 89.0 and 82.0 of AP-MSCs and IP-MSCs, respectively, were positive for CD90 marker. Furthermore, 68.0% of AP-MSCs and 71.0% of IP-MSCs were positive for CK19 marker. These results show that AP-MSCs and IP-MSCs have mesenchymal and epithelial cells characteristics.
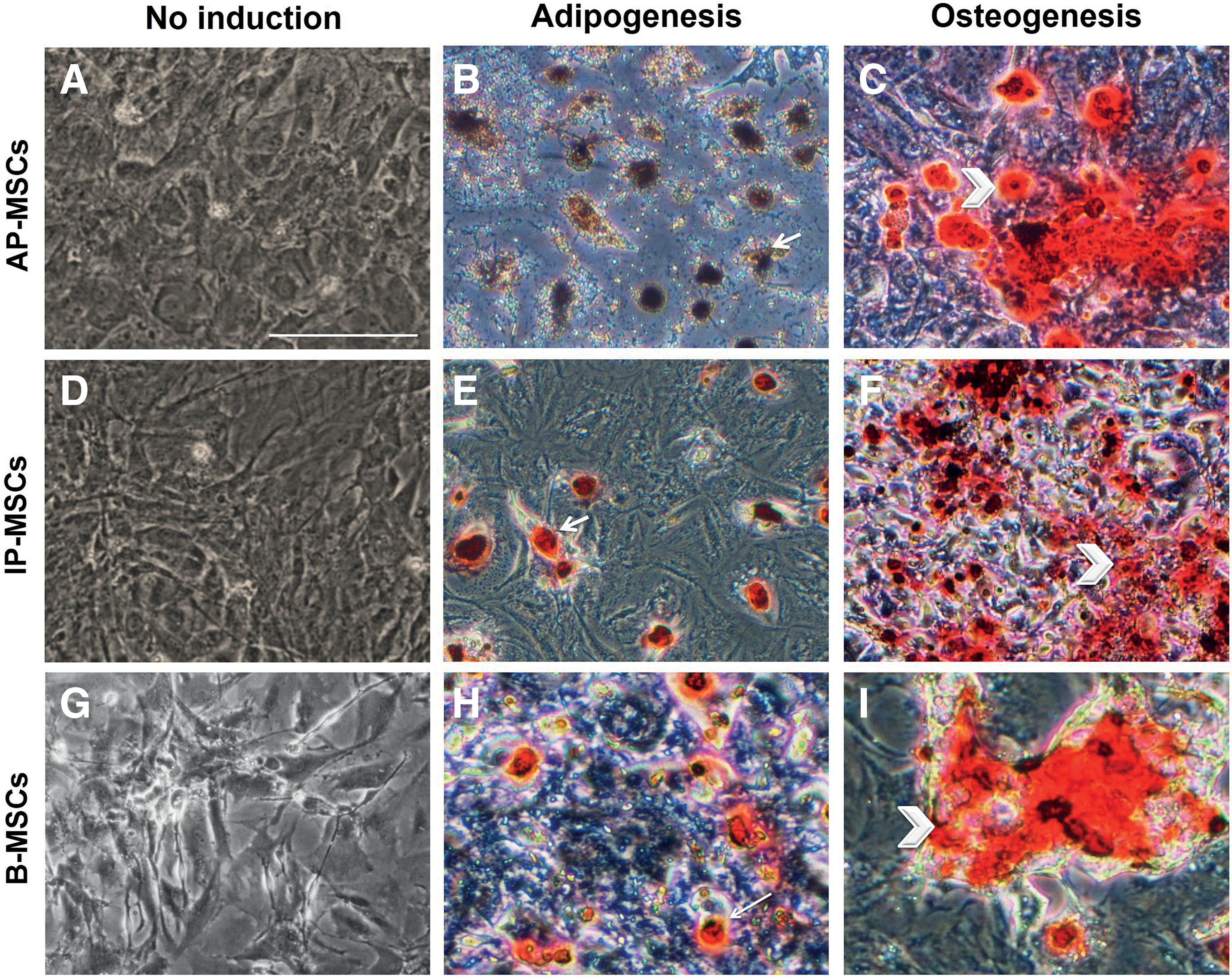
Adipogenesis and osteogenesis differentiation of PMSCs.

Cultured peritoneum mesenchymal stem cells expressed both mesenchymal and epithelial characteristics by immunocytofluorescence.
Morphological changes of AP-MSCs and IP-MSCs by HFF and HCCM induction
A one-step procedure was used to differentiate the passage 4 mouse AP-MSCs and IP-MSCs into ovarian cell-like cells. These cells were exposed to 15% FBS as control, 10% HFF + FBS 5% and 50% HCCM by adherent culture. In 10% HFF or 50% HCCM groups, some subpopulations of the cells morphologically changed after 4 days of differentiation. These cells began to grow into spherical shape and then gradually became morphologically similar to immature oocyte in size. A few of these cells remained small in diameter, but many of them reached up to 30 μm and were called germ cell (GC)-like cells (GLCs). The rest of the cells grew to over 50 μm and were called oocyte-like cells (OLCs). Furthermore, AP-MSCs can formed a few blastocyst-like structures when exposed to 10% HFF for 21 days (Fig. 4I). The morphological changes were observed least in 15% FBS. The cell count results showed that the AP-MSCs and IP-MSCs with diameter over 30 μm did not significantly differentiate in 10% HFF and 50% HCCM groups, but both groups analytically increased from day 0 to day 21 in the 15% FBS as control group (Fig. 4).
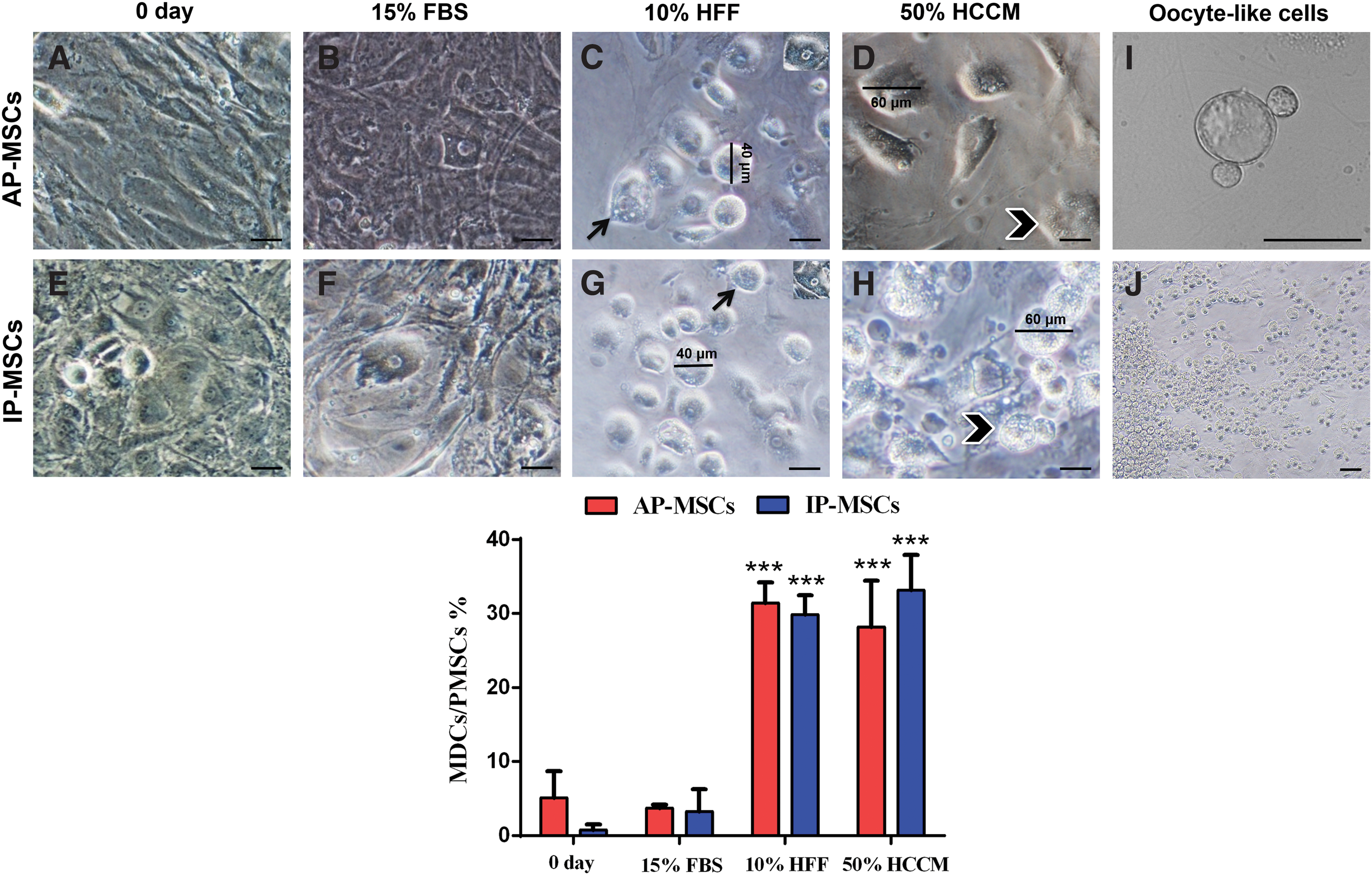
Morphological characterization of oocyte-like cells and germ cell-like cells differentiated from PMSCs.
RT-qPCR analysis of ovarian lineage genes
To verify whether the OLCs, GLCs, granulosa, and theca cells were identified based on morphological changes, and the ovarian cells gene expression was studied by RT-qPCR for oocyte (Zp3 and Gdf9), germ (Ddx4 and Vasa), granulosa (Amh), and theca (Lhr) cells in AP-MSCs and IP-MSCs at 0, 14, and 21 days of induction. Each amplified product was classified into differentiated and control cells. Once the products specificity was established, temporal changes were evaluated in the mRNA levels of these markers in the form of 2−ΔCt and Gapdh was a housekeeping gene. As shown in Fig. 5 for Gdf9, AP-MSCs showed increasing trend until the 21 days of culture with all FBS, HFF, and HCCM inductions, whereas IP-MSCs showed increase until 14 days of culture, but a sharp decrease occurred from the 14th to 21st day except in the presence of 50% HCCM. The HCCM group had better results in Gdf9 in both AP-MSCs and IP-MSCs special compared with the 0th day. For the Zp3, in AP-MSCs, HCCM and HFF groups showed increasing trend, but FBS showed a reduction, whereas in IP-MSCs, HFF and FBS did not show any increase, but the HCCM group significantly increased from the 14th to 21st day. However, the HCCM group in both AP-MSCs and IP-MSCs was better than the others and showed significant increase from the 0th day. For Ddx4, in both cell types, HCCM had the best induction from 14 to 21 days and again showed significant rise compared with the 0 day. For Dazl in both cell types and in total inductions, significant surge occurred from 14th to 21st day except for FBS in AP-MSCs and HFF in IP-MSCs. Here as well, the HCCM group showed the best performance. Finally, FBS induction did not have addictive effect on Amh expression in both cell types, but HCCM had powerful effect in AP-MSCs and showed significant increase compared with the 0th day (Fig. 5).
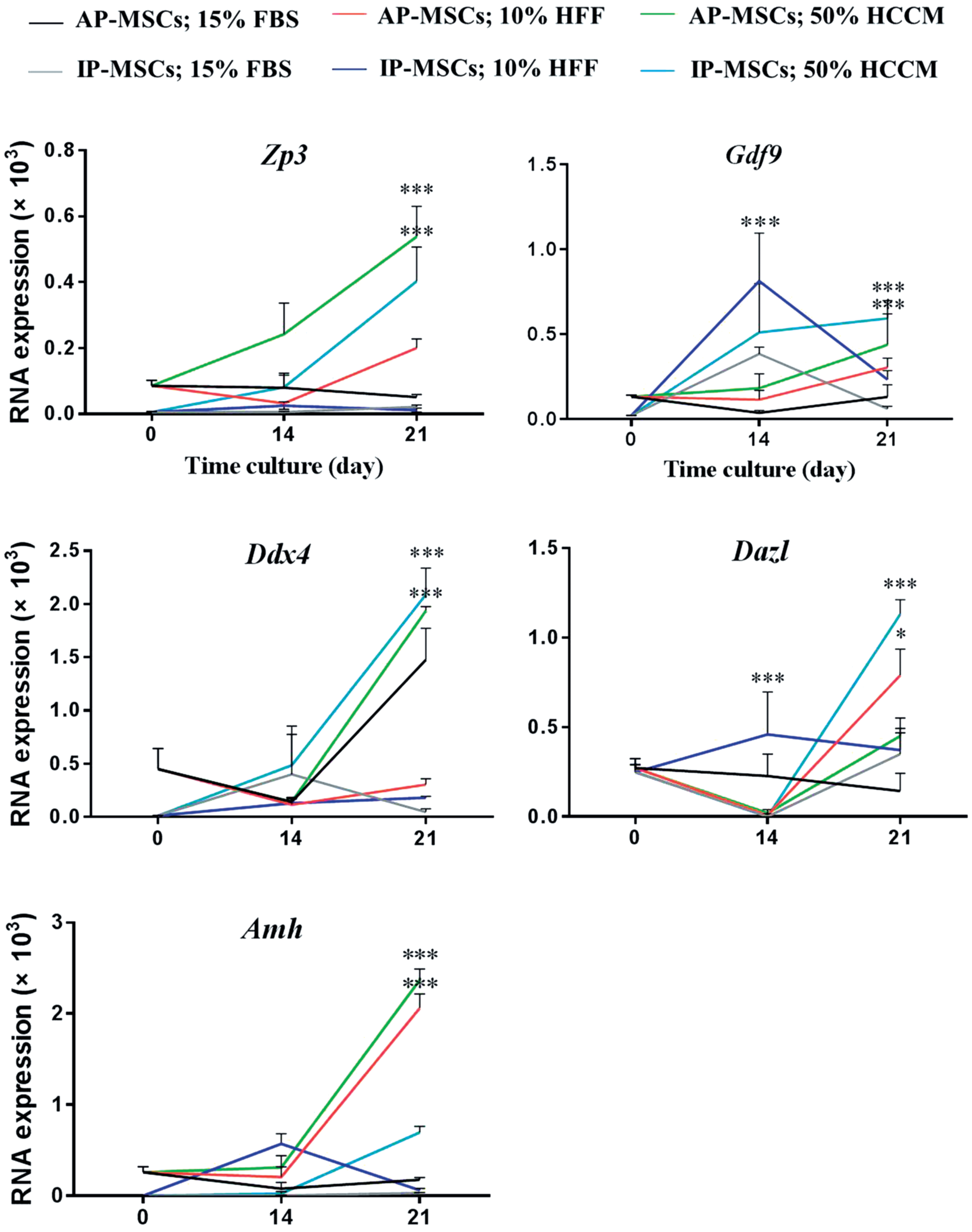
Gene expression analysis. Oocyte cell (Zp3, Gdf9), germ cell (Ddx4, Dazl), and ovarian cell (Amh) markers in oocyte-like cells, germ cell-like cells, and other ovarian cell-like cells expression by RT-qPCR. (*P < 0.05 and ***P < 0.001). RT-qPCR, reverse transcription quantitative polymerase chain reaction. Color images are available online.
Immunocytofluorescence for the expression of oocyte, germ cell, and granulosa cell proteins
To further characterize ovarian cell-like cells, immunocytofluorescence staining was applied to oocyte (ZP3 and GDF9), germ cell (DDX4 and DAZL), and granulosa cell (AMH) at 0 and 21 days of induction. ZP3, GDF9, DDX4, DAZL, and AMH proteins in both cell types and in HCCM and HFF groups showed increase compared with the 0th day while weak signal was detected in the controls with FBS. Furthermore, Amh protein was detected in granulosa-like cells with less fluorescent intensity than the other proteins (Fig. 6).

Immunocytofluorescence staining of oocyte, germ, and granulosa cell markers in differentiated PMSCs (Diff Cells). Images and graphs related to detection of
PMSCs differentiation potential into germ cell-like cells ex vivo
PMSCs are able to transdifferentiate into ovarian specialized cells in vitro. For this purpose, first, we showed that PMSCs were exposed to HFF and cumulus cells-conditioned medium able to differentiate into ovarian cell-like cells in vitro. To confirm this differentiation potential, PMSCs were injected into the decellularized ECM of the ovarian tissue. This differentiation potential was demonstrated with immunohistochemistry staining for DDX4 as a primordial germ cell marker. Immunohistochemistry staining result demonstrates that PMSCs by expressing DDX4 in ex vivo culture have potential differentiation to specific cell lineages of ovarian tissue (Fig. 7).
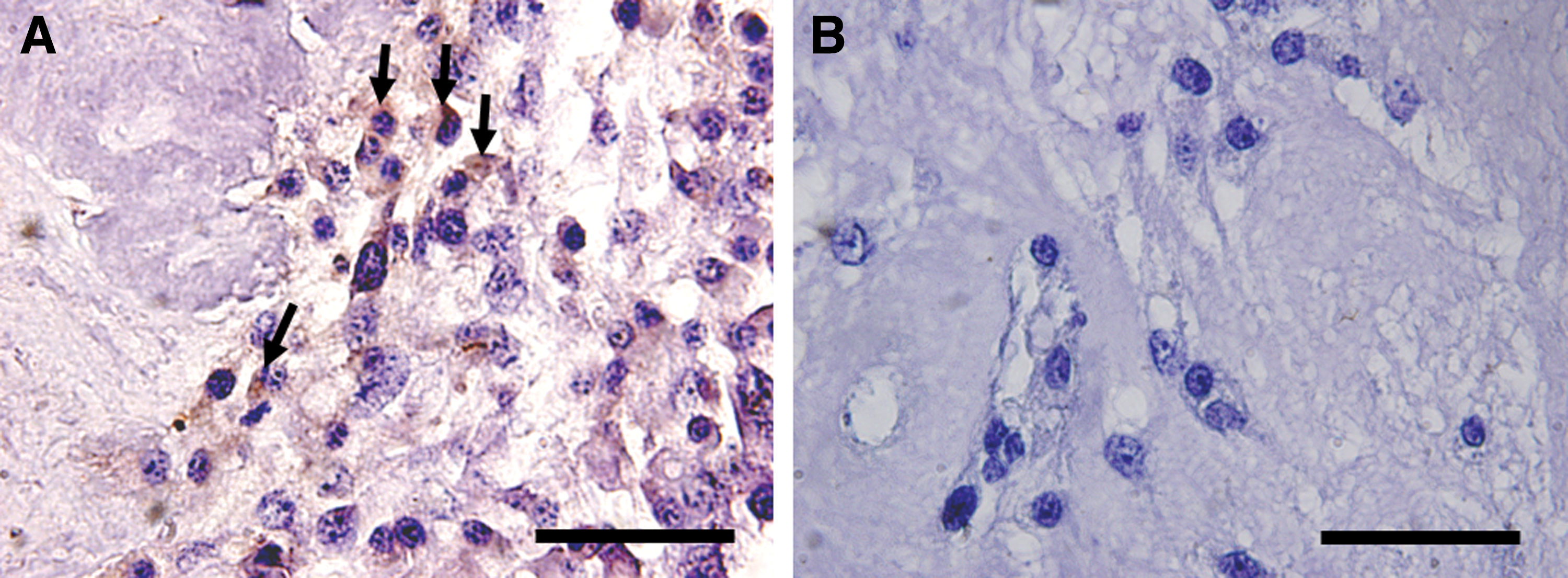
PMSCs ex vivo differentiation. Immunohistochemical staining of PMSCs seeded into ovarian scaffold for antibody against DDX4 as a germ cell marker.
Discussion
There has been, in recent times, a surge of interest in the use of MSCs in regenerative medicine and tissue engineering [18]. Recently, researchers have used these cells in novel infertility therapies [19]. Although some sources of MSCs like bone marrow are easy to use, their derivation is invasive due to pain and possibility of bone fracture and infection. MSCs derived from the other tissues like peritoneal mesothelium, which is lining organs and anterior abdominal wall, are interesting because of their basic function and their secretion of some essential growth factors. Although the peritoneum mesothelium is of mesenchymal origin, it shows epithelial characteristics such as surface expression of cytokeratin, suggesting that the tissue has some undifferentiated cells. The peritoneum mesothelium can potentially repair injured tissues because of its MSCs remnants [20].
The peritoneal mesenchymal stem cells are easily obtainable and can be harvested from the patient's omentum, without rejection to autotransplantation. However, studies have showed that peritoneum mesothelial cells bear markers similar to those expressed on B-MSCs [21]. In this study, we have shown for the first time, the differentiation potential of the anterior abdominal wall and intestinal peritoneal mesenchymal stem cells (AP-MSCs and IP-MSCs) into ovarian cell-like cells. For this purpose, the peritoneum mesothelial tissue explants were cultured in vitro, and then, the cells migrated from the edge of the tissue. In primary culture, these cells showed the characteristics of plastic adherence, colony formation, and heterogeneous morphology. These cells could be cultured until passage 15 without loss of multipotential properties, such as homogeneity, spindle-shaped, and colony-forming morphologies similar to B-MSCs. Also, these cells proliferated in both static (adherent) and dynamic (suspension) cultures.
According to the cellular growth pattern, AP-MSCs show a higher proliferation rate than IP-MSCs and were similar to B-MSCs [22]. Dauleh et al. cultured mouse peritoneum mesothelial cells and were able to culture up to 30 passages without observing cellular aging and exhibited clonogenicity as MSCs [22]. By contrast, Ksiazek et al. (2006) showed an increase in the size of human mesenchymal cells and downregulation of KI67 indicating low proliferation rate by passaging [23]. MTS assay showed that in vitro survival rates of both AP-MSCs and IP-MSCs were not comparable to B-MSCs. To prove the multipotential capacity of AP-MSCs and IP-MSCs, in addition to the biological assays, adipogenesis and osteogenesis potentials were assessed.
Differentiations to adipocyte and osteocyte were demonstrated by the accumulation of lipid droplets in the cells cytoplasm and calcium phosphate production in ECM, respectively. Shen et al. showed that mouse peritoneum mesothelial cells have MSC characteristics potential to differentiate into adipocyte, osteocyte, myocyte, and neurons in vitro, as well as differentiation into osteocyte in vivo [24]. Also, Dauleh et al. showed in murine species that adipogenic and osteogenic differentiations decrease passage by passage, which could be due to changes in the expression level of some of the stem cell markers. Therefore, high differentiation rate by increasing passage represents to a powerful stemness capacity.
In addition, by analyzing surface markers of AP-MSCs and IP-MSCs, we phenotypically characterized a CD44+/CD90+/CK19+ subpopulation. AP-MSCs and IP-MSCs dominantly expressed CD44- and CD90-like B-MSC. Similar morphology and expressions of cell surface antigens indicate the origin and function of these cells. Shah et al. displayed the expression of CD44 and CD90 but not CD45 expression in buffalo peritoneum mesothelial stem cells [25]. Chue et al. also showed that peritoneum mesothelial stem cells isolated from dog express surface markers similar to B-MSCs. In addition to the mesenchymal features, peritoneum mesothelial stem cells exhibit epithelial features with CK19 production. In the current study, the existence of CK19 in AP-MSCs and IP-MSCs was shown by comparing with skin epithelial cells as positive control.
However, since ovarian surface epithelium (OSE) cells are derived from peritoneum mesothelial cells in embryonic period, and also because many researchers believe that OSE is one of the main sources of oogonial stem cells, it seems that PMSCs potentially could differentiate to ovarian cells. We have shown, for the first time, the induction of AP-MSCs and IP-MSCs to generate OLCs and GLCs. We have shown that PMSCs can differentiate into ovarian cell-like cells by HFF and HCCM. Earlier researchers have shown that these types of cells have potential to differentiate into mesodermal and non-mesodermal lineages. Garcia-Gomez et al. reported high proliferation rate of human peritoneum mesothelial stem cells, which express CD34, bFGF, and VEGF, and differentiate into endothelial cells after transplantation [9]. Lowery et al. have also shown that the mouse omentum stem cells do not express CD34 and CD45 markers and, in specific differentiation medium, secrete the CCSP protein produced by Clara cells (the lung bronchioles epithelial cells) [26]. In addition, Keshteli et al. differentiated rat omentum stem cells into neurons [15]. In the present research, AP-MSCs and IP-MSCs were differentiated into ovarian cell-like cells in HFF and HCCM for 21 days. Morphological changes and marker expression profiles at the mRNA and protein levels were evaluated in differentiated PMSCs. Twenty-one days after induction with the FBS group as control, there were rare ovarian cell-like cells. However, considerable number of spherical cells was observed in HFF and HCCM, which shows the existence of cells subpopulation in peritoneum mesothelial tissue with inherent characteristics like ovarian cells.
Studies have shown that OLCs and GLCs derived from different MSC sources induced with FF, CCM, gonadotropins, and some growth factors. Danner et al. observed that the aggregates derived from mouse pancreatic stem cells spontaneously differentiate into OLCs [27]. Dyce et al. achieved oocyte from porcine embryonic stem cells in 5% FF and 5% FBS [28]. Shah et al. (2017) differentiated buffalo embryonic stem cells in 20%–40% CCM into OLCs and GLCs, and they showed the most expression of meiosis and germ cells genes in 20% CCM and the most expression of gamete genes in 40% CCM [25]. In the current investigation, we have shown that peritoneal mesenchymal stem cells from abdominal wall (AP-MSCs) or viscera surrounded peritoneum (IP-MSCs) have the potential to differentiate into OLCs and GLCs in terms of oocyte and germ cell markers expression under HFF and HCCM induction. Additionally, AP-MSCs and IP-MSCs were differentiated into granulosa-like cells with ability of AMH (granulosa cell marker) expression, but they could not present LHR as theca cell marker.
This study has shown that HCCM and HFF are more effective than FBS for expression of ovarian cell-specific markers. Furthermore, HCCM has the greatest effect on the production of OLCs and GLCs. However, in the production of granulosa-like cells, the effects of HCCM and HFF were approximately equal, but neither had an effect on the production of theca-like cells. Both AP-MSCs and IP-MSCs had similar differentiation potential into OLCs, but AP-MSCs had greater ability than IP-MSCs in production of GLC markers.
It is understood that some of AP-MSCs and IP-MSCs would support oogenesis process. However, we did not observe any folliculogenesis or follicular formation even at primordial stage, and in fact, the size of the OLCs was not as large as natural oocytes, AMH production suggests this spontaneous capacity in these cells. Furthermore, PMSCs are able to differentiate into ovarian cell-like cells not only when exposed to follicular or cumulus cells factors but also when seeded and cultured in the bed of ovarian ECM. Appearance of DDX4-positive cells, 1 week after ex vivo culture, within ovarian ECM gathered with PMSCs showed a good potential of peritoneum mesothelial stem cells to differentiate and then produce germ cells proteins in the niche of ovary.
Conclusions
It has been demonstrated that PMSCs are differentiated to osteoblasts, neuronal cells, and in damaged arteries begin to endothelialization, and also differentiated into ovarian cell-like cells. Thus, this type of cells has high plasticity, and it is possible that they show different behavior under different conditions. Our investigation offers a small step in the large area of stem cell therapy in infertility treatment. Although there are some restrictions in human-like sample privation, or unable to track them in vivo, PMSCs can be isolated from peritoneum mesothelium and differentiated into the ovarian cell-like cells and may help to reintroduce the fertility ability.
Footnotes
Acknowledgments
This study was supported by the Royan Institute, the Iranian Council of Reproductive Biomedicine Research Center (Tehran, Iran), and we are thankful for all of our colleagues especially in embryology and stem cell departments.
Author Disclosure Statement
No competing financial interests exist.