
Abstract
The commitment of pluripotent stem cells to the cardiac lineage has enormous potential in regenerative medicine interventions for several cardiac diseases. Thus, it is necessary to understand and regulate this differentiation process for potential clinical application. In this study, we developed defined conditions with chemical inducers for effective cardiac lineage commitment and elucidated the mechanism for high-efficiency differentiation. First, we designed a robust reporter-based platform to screen chemical inducers of cardiac differentiation in the mouse P19 teratocarcinoma cell line. Using this system, we identified two natural alkaloids, lupinine and ursinoic acid, which enhanced cardiomyocyte differentiation of P19 cells in terms of beating colony numbers with respect to oxytocin, and confirmed their activity in mouse embryonic stem cells. By analyzing the expression of key markers, we found that this enhancement can be attributed to the early and rapid induction of the Wnt signaling pathway. We also found that these natural compounds could not only supersede the action of the Wnt3a ligand but also had a very quick response time, allowing them to act as efficient cardiac mesoderm inducers that subsequently promoted cardiomyocyte differentiation. Thus, this study offers a way to develop chemical-based differentiation strategy for high-efficiency cardiac lineage commitment, which has an advantage over currently available methods with complex medium composition and parameters. Furthermore, it also provides an opportunity to pinpoint the key molecular mechanisms pivotal to the cardiac differentiation process, which are necessary to design an efficient strategy for cardiomyocyte differentiation.
Introduction
Pluripotent stem cells have the potential to proliferate indefinitely in an undifferentiated state and can differentiate into various cell types under suitable conditions [1]. Thus, these cells provide an important feasible therapeutic alternative for potentially incurable diseases [2]. To take advantage of their enormous potential, it is necessary to develop optimal ways of generating specific cell types in a scalable, cost-effective, and reproducible manner. Several cardiovascular disorders require stem cell therapy as the only feasible therapeutic intervention. Therefore, there is a need to develop efficient protocols of generating functional cardiomyocytes suitable for therapeutic usage.
Several methods have been reported to produce cardiomyocytes from stem cell: treatment with growth factors or cytokines [3 –8], or bacteria-mediated forced delivery of transcription factors to pluripotent cells [9,10]. However, because those methods have limitations such as high production cost [11], risk of bacterial infection [9,10], and low efficiency [11], they cannot be further developed for clinical use. Another promising way to achieve differentiation is to use chemical inducers that mimic physiological cues due to their easy scalability and applicability [11,12].
In the case of chemical-based cardiac differentiation, choice of efficient cardiogenesis-promoting chemicals relied on mimicking the developmental cues that pluripotent cells encounter during differentiation into cardiomyocytes [11,13 –15]. Because cardiac development largely relies on signaling such as activin/nodal [14], bone morphogenetic protein (BMP) [16 –18], and Wnt [11,13,14,18 –21], controlling the signaling results in successful cardiogenesis. However, chemical-based approach also has limitations such as suboptimal efficiency [11] and target specificity [11]. Due to these drawbacks, further development of chemical-based differentiation strategies has been hindered [11]. Therefore, there is a legitimate need to find novel chemical agents that will have defined targets and offer high efficiency of commitment.
In this study, we aimed to find compounds of natural origin that effectively activate cardiogenesis through a well-defined mechanism. For this purpose, we developed an efficient cardiomyocyte-specific readout reporter system based on the mouse embryonic carcinoma cell line P19, which has a pluripotent nature capable of differentiating into three germ layers [22 –25]. In this reporter system, P19 cells express mCherry fluorescent protein driven by the α-MHC (myosin heavy chain) promoter, which is activated upon cardiac induction.
By screening natural compounds using this reporter system, we identified two promising cardiac inducers—lupinine and ursinoic acid—which showed better differentiation capacity than oxytocin, a known inducer [26], and further investigated their effect on cardiogenesis. We found that these chemicals induce cardiomyocyte differentiation by triggering Wnt signaling in the early stage of differentiation. Therefore, these results not only demonstrate that these compounds can be utilized in the development of a simple and efficient cardiogenic differentiation protocol but also confirm the key cellular signaling required for controlling cardiomyocyte differentiation.
Materials and Methods
Cell culture
P19 cells were cultured in αMEM (12571-071; Gibco) supplemented with 10% fetal bovine serum (FBS, 16000-044; Gibco) and 1% penicillin and streptomycin (LS202-02; Welgene) at 37°C in 5% CO2. P19 cells were passaged every 2 days after trypsinizing with 0.05% trypsin-EDTA (LS015-10; Welgene). HEK293T cells were cultured in high-glucose Dulbecco's modified Eagle's medium (DMEM, LM001-05; Welgene) supplemented with 10% FBS and 1% penicillin/streptomycin at 37°C in 5% CO2.
Mouse embryonic stem (mES) cells were cultured in high-glucose DMEM (11995-065; Welgene) supplemented with 15% FBS (SH30919.03; Hyclone), 2 mM
Lentiviral packaging and making stable reporter cell line (Cherry19)
We used the lentiviral SIN18.WPRE vector containing the mCherry fluorescent protein coding sequence driven by the cardiomyocyte-specific α-MHC promoter (Addgene plasmid #21228). This vector also contained the blasticidin S (15205; Sigma-Aldrich) resistance gene under the control of the pluripotency-specific promoter REX so that this vector could be selected in a pluripotent background [27]. α-MHC-mCherry lentivirus particles were produced in HEK293T cells according to previously described methods [27].
Briefly, the α-MHC-mCherry vector, the packaging plasmid pCMV-dR8.2 dvpr (Addgene plasmid #8455), and the envelope protein expression plasmid pCMV-VSV-G (Addgene plasmid #8454) were cotransfected into HEK293T cells in a ratio of 5:4:1 with Turbofect (R0532; Thermo-Fisher). The viral supernatant was collected 48 h after transfection. The viral supernatant was centrifuged and stored at −80°C until future use. For viral transduction, the P19 cells were seeded into 150-mm dishes 24 h before infection. The viral supernatant was mixed with 7.5 μg/mL polybrene (Sigma-Aldrich), added to confluent P19 cells, and incubated overnight. The media were exchanged with fresh media after 24 h. Stable P19 cells expressing α-MHC-mCherry are henceforth referred to as “Cherry19 cells,” are were selected with 10 μg/mL blasticidin S.
Differentiation of P19 and mES cells
The schema for differentiating reporter P19 cells (Cherry19) is depicted in Fig. 1. Briefly, 1 × 104 cells were grown as a suspension culture to form embryoid bodies (EBs) for 3 days in ultra-low attachment culture plates (CLS3473; Corning) to initiate differentiation in the presence or absence of compounds from the chemical library. After 3 days, EBs were transferred to normal tissue culture plates and grown without further treatment.
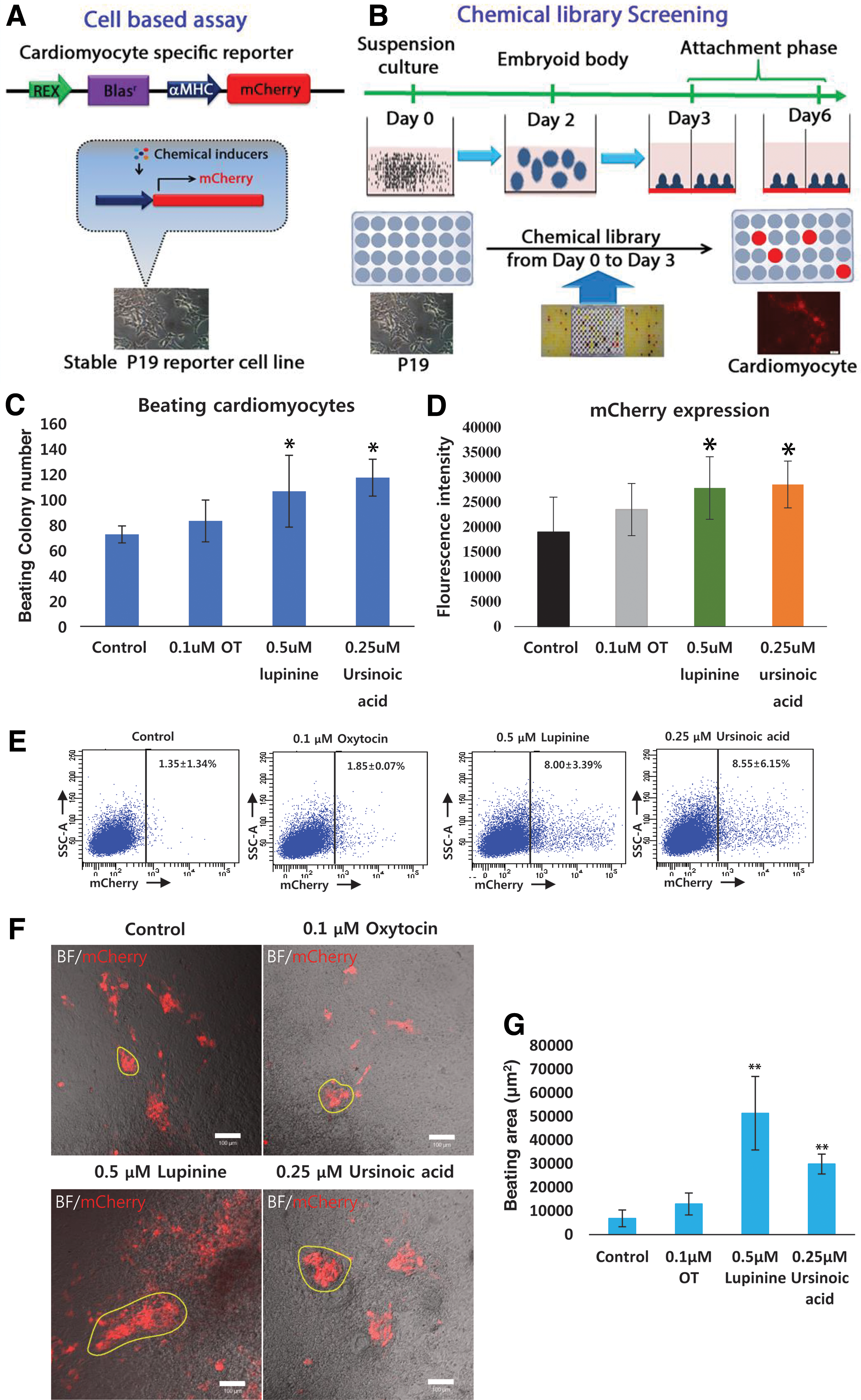
The overall schema of a cell-based assay system for chemical library screening.
Similarly, mES cells were trypsinized and replated on tissue culture dishes for 1 h for preferential removal of MEF feeder cells. Subsequently, 1 × 105 floating mES cells were transferred to low-attachment plates to form EB in the presence or absence of the inducers. After 3 days, EBs were transferred to normal tissue culture plates in fresh media (high-glucose DMEM supplemented with 15% FBS, 2 mL
Natural compound library
A natural product library containing 800 compounds of diverse alkaloids, flavonoids, and sterols (NatProd Collection, MicroSource Discovery System) was diluted to 1 mM in dimethyl sulfoxide (DMSO).
Chemical screening of natural compounds to measure mCherry fluorescence intensity
5 × 102 Cherry19 cells were grown as a suspension culture in low-attachment 96-well plates (3474; Costar) to form EBs for 3 days in the presence of 1 μM compounds from the natural compound library. Subsequently, the EBs were transferred to standard 96-well tissue culture plates. Media were changed every other day. On day 6, the differentiated cells were lysed with 20 μL of 1 × passive lysis buffer (E194A; Promega) with 20 min of gentle rocking. The cell lysate was transferred into 96-microwell optical bottom plates (165305; Nunc) and mCherry fluorescence intensity was measured using multimode microplate readers (TECAN, Männedorf). mCherry fluorescence intensity was subsequently normalized with respect to an empty well and a well containing lysis buffer only. Enhancements of fluorescence intensity were assessed with respect to Oxytocin-treated control P19 cells.
From the initial screening, we selected six hit compounds (Supplementary Table S1; Supplementary Data are available online at
Flow cytometry analysis
P19 cell-derived cardiomyocytes were dissociated using 0.25% Trypsin/EDTA (LS015-10; Welgene) on the fifth day of differentiation and reattached to the plate as a single layer. One day later (sixth day of differentiation), the cells were dissociated with 0.1% Trypsin/EDTA (Welgene) and resuspended in serum containing media for flow cytometry analysis using a 587-nm laser (BD FACSAria II; BD Biosciences).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from differentiated Cherry19 cells on days 0, 5, 10, and 15 after attachment by the RNeasy Mini Kit (Qiagen). Total RNA (5 μg) was subjected to cDNA synthesis using the RNA to cDNA EcoDry Premix (Oligo dT; Clontech). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using SYBR Green PCR Master Mix (Bio-Rad) on a Bio-Rad Prime PCR instrument. The qRT-PCR conditions included 40 cycles of 30 s at 95°C, 15 s at 60°C, and 15 s at 72°C. The primers used in these studies are provided in Supplementary Table S2.
Preparation of Wnt3a-conditioned media
Wnt3a-producing L cells and control L cells were cultured according to ATCC instructions for producing Wnt3a-conditioned media and control media, respectively. Both cells were cultured in high-glucose DMEM supplemented with 10% FBS and 1% penicillin and streptomycin at 37°C in 5% CO2. After 24 h, conditioned media were collected, filtered, and stored at −80°C until further use.
Immunostaining
mES and P19 cells were grown as suspension cultures for 3 days with and without chemical inducers. P19 cell-derived cardiomyocytes were immunostained using α-sarcomeric actinin (α-SMA) antibody on day 7, Mef2c/MF20 antibodies on day 9, Nkx2.5/α-SMA antibodies on day 10, and cardiac troponin T (cTnT) and cardiac troponin I (cTnI) antibodies on day 15. mES cell-derived cardiomyocytes were immunostained using Mef2C/α-SMA antibodies on day 8 and cTnT/Cx43 (connexin 43) antibodies on day 17.
On each indicated day, cells were washed twice with 1× phosphate-buffered saline (PBS, LB201-02; Welgene), fixed with 4% paraformaldehyde (F-1268; Sigma-Aldrich) for 10 min, permeabilized with 0.25% Triton X-100 (22686; USB Corporation) in PBS for 20 min at room temperature, and washed in PBS thrice for 5 min each. Then, cells were blocked with blocking solution containing 1% bovine serum albumin (0332; Amresco), 22.52 mg/mL glycine (16407; Affymetrix), and 0.1% Tween 20 (T1003; Affymetrix) in PBS for 60 min. Subsequently, the cells were stained with the appropriate primary antibody diluted at a ratio of 1:100 in the blocking solution overnight at 4°C.
The antibodies used were rabbit polyclonal anti-Mef2c (Ab64644; Abcam), anti-Nkx2.5 (AB35842; Abcam), mouse monoclonal anti-sarcomeric actinin (A7732; Sigma-Aldrich), anti-cTnT (AB10214; Abcam), troponin I (TI-1; Developmental Studies Hybridoma Bank), Cx43 (3512S; Cell Signaling), and α-MHC (MF 20; Developmental Studies Hybridoma Bank). After primary antibody incubation, cells were washed in PBS thrice and incubated with either Alexa-488-conjugated goat secondary anti-mouse antibody (A11001; Invitrogen) or Alexa-563-conjugated goat secondary anti-rabbit antibody (A21428; Invitrogen) diluted 1:100 for 2 h at room temperature.
Cells were incubated with 1.0 μg/mL DAPI (D9542; Sigma-Aldrich) for 5 min at room temperature to stain the nuclei. Samples were subsequently visualized using fluorescence microscopy (IX71S1F3; Olympus). Immunofluorescence signal was quantified using Image J software (Institutes of Health, NIH,
Measuring cytosolic Ca2+ concentration
Cells were incubated with 3 μ
Luciferase reporter assay
HEK293T cells were cultured in high-glucose DMEM supplemented with 10% FBS. The 5 × 104 HEK293T cells were seeded into a 24-well tissue culture plate 1 day before transfection. Reporter plasmids, TOPflash (containing T-cell factor [TCF] binding site) or FOPflash (containing mutated TCF binding sites), which drive the expression of the firefly luciferase gene, were transfected along with internal control Renilla luciferase reporter vector (10:1 ratio) to the cells. After 24 h, the cells were transferred to high-glucose DMEM supplemented with 2% FBS and were concurrently treated with oxytocin, natural compounds (lupinine, and ursinoic acid), Wnt activator (Chir99021), and Wnt inhibitor (XAV939). Luciferase activities were measured 48 h after chemical treatment according to the manufacturer's protocol (E1960; Promega). The Topflash/Fopflash luciferase activities were normalized with respect to the Renilla luciferase activity.
Treatment of Wnt inhibitor XAV939
mES cells were cocultured with an MEF feeder layer on gelatin-coated plates in high-glucose DMEM (11995-065; Welgene) supplemented with 15% FBS (SH30919.03; Hyclone), 2 mM
Statistical analysis
Values are presented as mean ± standard deviation as calculated from the average of at least three biological replicates, unless otherwise specified. The statistical significance of variation was analyzed by t-test using Prism 6 software (GraphPad). P < 0.05 was considered to be statistically significant.
Results
Platform for screening novel cardiogenic differentiation inducers
To set up large-scale screening experiments, we used a cardiac-specific α-MHC promoter-driven mCherry reporter cell line. α-MHC is expressed early on during cardiac commitment and therefore can be utilized for screening purposes within a relatively short time frame [27]. Consequently, we established a stable reporter cell line capable of expressing mCherry fluorescent protein upon cardiac induction in mouse P19 cells. A lentiviral reporter construct containing the α-MHC-driven mCherry gene and the pluripotency-specific promoter Rex-driven blasticidin resistance gene was transduced into P19 cells, and reporter-integrated stable cell lines were established by antibiotic selection. Thus, α-MHC-mCherry_Rex-Blar P19 cells (henceforth referred to as “Cherry19 cells”) provided the required platform to perform large-scale high-throughput screening.
Screening natural compound library
For screening the cardiac differentiation inducers, we used a natural compound library containing 800 active compounds originated from a wide variety of natural sources (Supplementary Fig. S1). Cherry19 cells were grown as a suspension in low-attachment plates to initiate EB formation, and subsequently cultured for 2 days in the presence of each compound. On day 3, EBs were transferred to normal cell culture dishes to continue further differentiation under attachment conditions for another 3 days. On day 6, the cells were visualized for mCherry expression (Fig. 1B).
Based on the enhanced mCherry fluorescence with respect to untreated cells, we found 19 hits that showed the higher fluorescence than the threshold in the first round of screening (Supplementary Fig. S1). Among the initial hits, only six compounds showed significantly enhanced fluorescence with respect to control cells and oxytocin, a known cardiac inducer [5,26], in subsequent differentiation experiments (Supplementary Fig. S2 and Supplementary Table S1). Out of the six compounds, lupinine and ursinoic acid showed most consistent enhancement of mCherry fluorescence in further confirmation experiments, and thus were selected for further characterization (Supplementary Fig. S2A, B).
Characterization of lupinine and ursinoic acid as cardiac differentiation inducers
To identify the optimal dose of each compound, compounds were added at various concentrations and mCherry expression was monitored (Supplementary Fig. S2C, D). Finally, 0.5 μM of lupinine and 0.25 μM of ursinoic acid were chosen for subsequent experiments. The number of beating colonies (Fig. 1C) and the mCherry fluorescence intensity (Fig. 1D) were quantitated as the efficiency of cardiac induction. While oxytocin treatment produced 83 ± 10 beating colonies/104 cells, lupinine and ursinoic acid treatment resulted in 105 ± 25 and 117 ± 15 beating colonies/104, respectively, indicating that 27.7% and 41.0% higher differentiation efficiencies were obtained by the treatment of lupinine and ursinoic acid, respectively, in comparison to oxytocin treatment condition (Fig. 1C).
We also observed a similar trend in mCherry fluorescence intensity: 18.4% and 23.4% enhancement, respectively, in the case of lupinine- and ursinoic acid-treated cells compared to oxytocin-treated cells (Fig. 1D). In addition, FACS-based quantitation revealed that lupinine and ursinoic acid treatment induced 4.3- and 4.6-fold enhancement of the mCherry(+) cell population, respectively, with respect to the oxytocin control (Fig. 1E). Consistently, lupinine- and ursinoic acid-derived cardiomyocytes had a significantly larger synchronous beating area in comparison to oxytocin-derived cells (Fig. 1E, F). Lupinine treatment resulted in the largest beating area of 51,000 ± 15,000 μm2 and ursinoic acid treatment resulted in a beating area of 30,000 ± 4,000 μm2 (Fig. 1E, F and Supplementary Movies S1–S4), which were significantly larger than the areas generated by either oxytocin-treated cells (12,000 ± 4,000 μm2) or untreated cells (10,000 ± 3,000 μm2).
Effect of lupinine and ursinoic acid on cardiac-specific marker expression
To further validate activation of the cardiac-specific program at a molecular level, we quantitated the transcript levels of various cardiac lineage-specific markers by qRT-PCR (Fig. 2). We examined several cardiac markers corresponding to early, middle, and late stages of cardiomyocyte development and determined their expression at different time points (ie, 5, 10, and 15 days; Fig. 2). Early- and middle-stage markers, including GATA4, Mef2c, and Nkx2.5, showed an increased expression on day 10. Transcript levels of GATA4 and Nkx2.5, in case of both lupinine and ursinoic acid treatment, gradually increased up to day 10, after which they were significantly reduced (Fig. 2). This trend might be due to the fact that the natural compound-treated cells advanced to the next stage of maturation where these markers were no longer relevant.
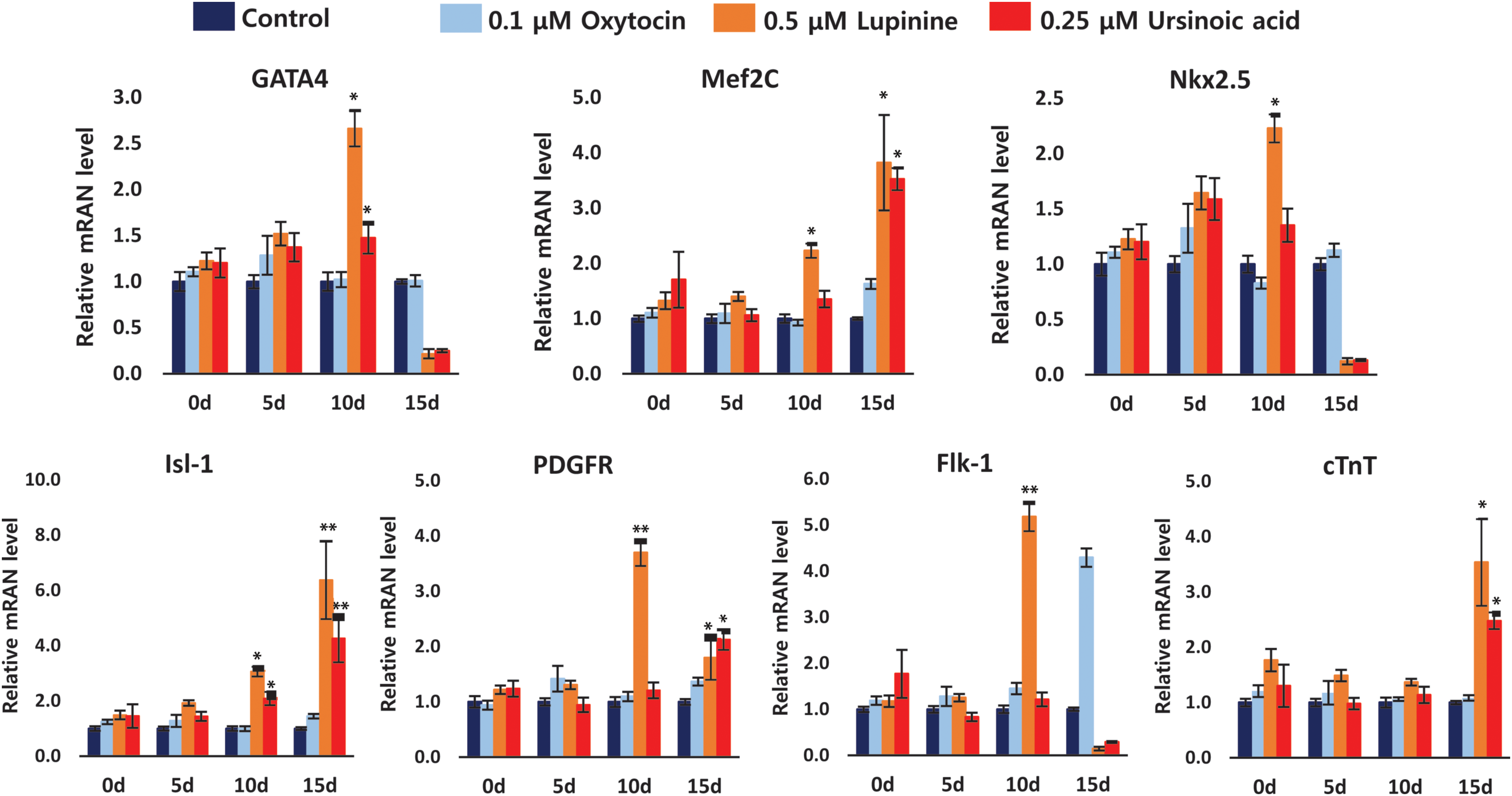
Assessment of transcript levels of key cardiac-specific markers in the presence of inducers. Transcript levels of various cardiac-specific markers corresponding to early (GATA4), mid-early (Nkx2.5, PDGFR, and Flk1), and mid-late stages (Mef2c, Isl1, and cTnT) in the lupinine- or ursinoic acid-treated cells were quantitated using real-time PCR and compared to those of cells treated with oxytocin or no inducer. For optimal signal assessment, marker expression was determined on days 0, 5, 10, and 15. Fold changes were calculated with respect to the signal in cells treated with no inducer. Data here represent the mean ± SEM (n = 3). * and ** indicate P < 0.05 and P < 0.01, respectively, compared to oxytocin. cTnT, cardiac troponin T; PCR, polymerase chain reaction. Color images available online at
Therefore, the suppression of these genes is required for successful progression to the next stages of development. Interestingly, we observed a dramatic increase of the markers of cardiac progenitor cells (CPCs), including PDGFR and Flk, in the lupine-treated condition, suggesting that expandable CPC can be obtained during the differentiation process. On the other hand, ursinoic acid treatment resulted in only a slight upregulation of PDGFR (Fig. 2). Consistently, we observed the upregulation of middle- and late-stage markers, including Mef2C, Isl1, and cTNT, on day 15.
Next, we examined the protein levels of pan muscle and cardiac markers to further assess the extent of differentiation in various conditions. The number of cells expressing pan muscle markers such as α-SMA, MHC, and Mef2c was significantly higher when lupinine or ursinoic acid was used (Fig. 3A.I and II) in comparison to the cases with oxytocin or no inducer. To eliminate the chance of detecting immunopositivity from noncardiac cells, we monitored the expression of cardiac-specific markers Nkx2.5, cTnT, and cTnI (Fig. 3A.III and IV). Both lupinine and ursinoic acid treatment increased the number of cells that were immunopositive for cardiac-specific markers. These results support that both compounds significantly activated the cardiac-specific program.

Assessment of protein levels of key cardiac-specific markers in the presence of inducers.

Next, we examined the capacity of these two compounds as cardiac differentiation inducers using mES cells. We also confirmed that lupinine and ursinoic acid enhanced the cardiogenesis of mES cells by observing an increase in the number of immunopositive cells for Mef2c, α-SMA, cTnT, and Cx43 upon treatment of mES cells with natural compounds (Fig. 3B.I–IV). The extent of maturation in these cells was assessed by characteristic subcellular structures of cardiomyocytes marked by α-SMA, cTnT, and Cx43 antibodies. Prominent subcellular structures were more pronounced in the cells treated with natural compounds compared to oxytocin or no treatment (Fig. 3B.II–IV). The enhanced cardiomyocyte differentiation upon natural compound treatment was also validated by quantitating immunostaining signal intensity of all the antibodies used above (Fig 3C, D).
Physiological validation of cardiomyocyte differentiation induced by lupinine and ursinoic acid
To access mechanical properties such as beating amplitude and frequency of the differentiated cells, we measured intracellular calcium transients [Ca2+]i in cardiomyocytes differentiated from mESs under different conditions. We observed spontaneous [Ca2+]i signals in all conditions, but lupinine-treated cells showed the highest amplitude, followed by ursinoic acid- and oxytocin-treated cells. Noninduced cells showed the most compromised signal amplitude among all the samples (Fig. 4A, C). Upon pacing the cells at a frequency of 1 Hz, we observed a similar trend in amplitudes with improved signals, compared to without pacing (Fig. 4B, C).

Calcium transient measurement.
Enhanced cardiac induction occurs through activation of Wnt signaling
For optimization of the differentiation efficiency and formulating rationally designed protocols, it is necessary to understand the mechanisms of enhanced cardiac induction upon compound treatment. Cardiomyocyte commitment from pluripotent stem cells has been broadly conceived as a biphasic process consisting of cardiac mesoderm generation followed by cardiomyocyte commitment [6,28], which are controlled by Wnt signaling positively and negatively, respectively [13,29,30]. Because our screenings of natural compounds were performed in the EB phase, we reasoned that the cardiac mesoderm commitment might be affected by compound treatment by Wnt activation.
To test this hypothesis, we treated cells in the EB phase for 72 h with lupinine or ursinoic acid and quantitated the Wnt3a transcript level at different time points. We observed that the Wnt3a transcript level at 24 h was significantly higher in the cells treated with the natural compounds compared to the untreated cells or oxytocin-treated cells (Fig. 5A). This rise in Wnt3a level was more pronounced when cells were grown in Wnt3a-conditioned media (Fig. 5A). This observation might be due to a synergistic effect of the Wnt3a ligand and the natural compounds working in the same signaling axis.
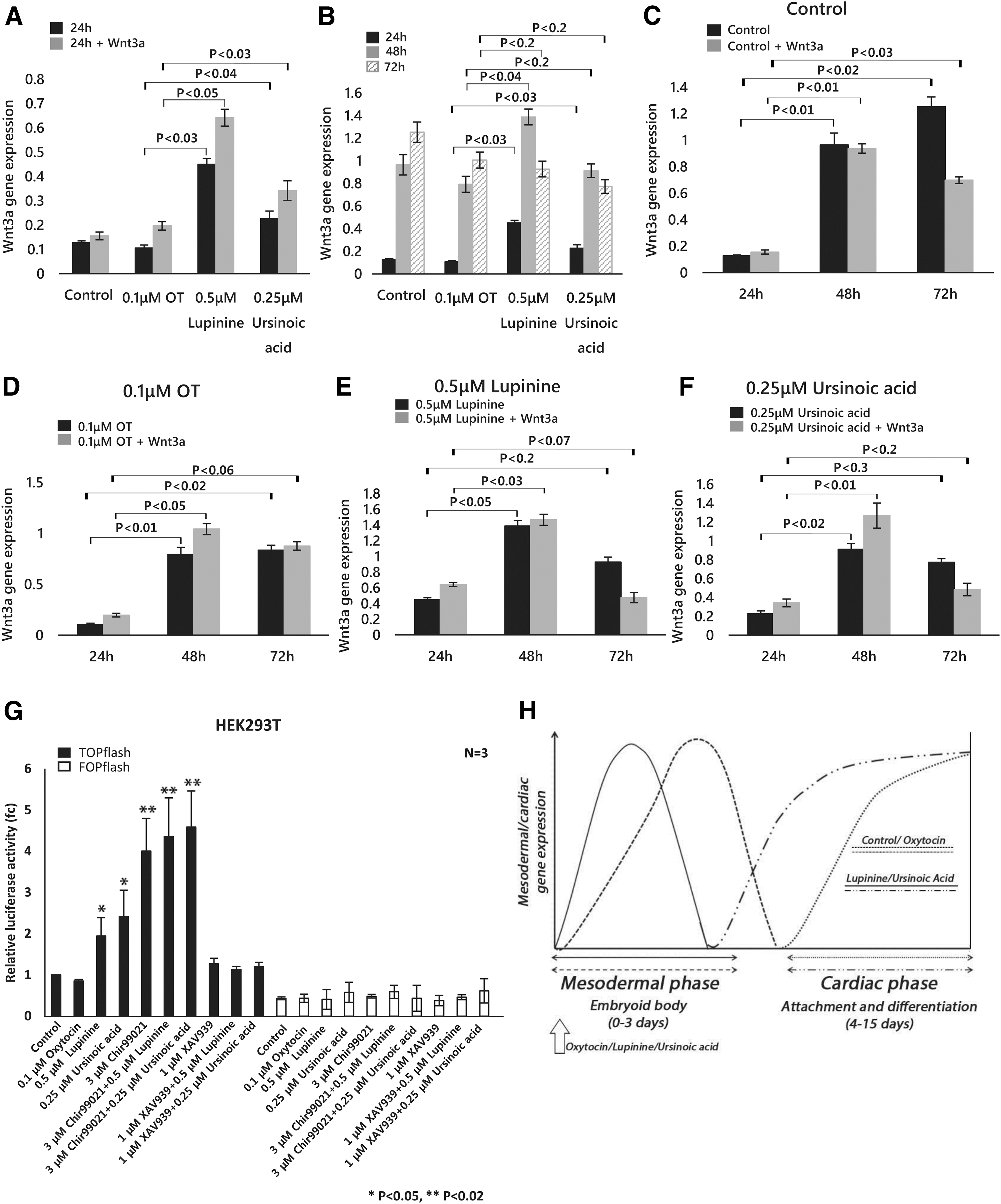
Wnt3a transcript level upon compound treatment.
We also noticed that Wnt 3a transcripts increased up to 48 h, but started decreasing at 72 h (Fig 5B) when either lupinine or ursinoic acid was added, whereas there was a continuous increase of Wnt3a transcript level until 72 h in the control and oxytocin-treated cells (Fig. 5B). These results indicate that natural compounds induce the early activation and subsequent suppression of Wnt signal. Because Wnt signaling is required for efficient cardiac mesoderm during the EB phase and also undergoes suppression for the transition from mesoderm to the next phase of cardiac commitment [13,30], it was hypothesized that the natural compounds in this study enhance the cardiogenesis by controlling the Wnt signaling pathway.
This hypothesis was further supported by observing the synergistic cooperation between the natural Wnt3a ligand from conditioned media and all conditions, including lupinine and ursinoic acid, and this synergy persisted only up to 48 h of treatment (Fig. 5C–F). After that time point, the cooperation was decoupled and Wnt3a transcript levels in cells grown in Wnt3a-conditioned media were dramatically suppressed, possibly through a negative feedback mechanism. This Wnt3a suppression was more dramatic in cells treated with lupinine and ursinoic acid (Fig. 5E, F), accounting for enhanced cardiac differentiation by these natural compounds.
To further consolidate our view on modulation of Wnt signaling by each compound, we quantitated the expression of Wnt target genes, including Lef1, Tcf1, and Auxin2, at different time points after treating with chemicals (Supplementary Fig. S3A). All three genes showed an enhanced expression upon chemical treatment in the EB phase. The time-dependent variation of their expression pattern was consistent with that of Wnt3a, indicating that chemical-induced cardiac differentiation is the consequence of Wnt signaling activation (Supplementary Fig. S3A).
To further confirm the activation of Wnt signaling by the natural compounds, we assessed the Wnt-enhanced transcriptional activity by the TOPflash reporter assay. In alignment to our observation of Wnt activation by the natural products, we observed that the luciferase activity was highest in the presence of the natural compounds, in comparison to the activity of the control or the activity under the condition of oxytocin treatment (Fig. 5G). Next, we tested the effect of a Wnt activator (Chir99021) and a Wnt inhibitor (XAV939) on the Wnt activity enhanced by the natural compounds (Fig. 5H). While Chir99021 further enhanced the Wnt activity, XAV939 completely abolished the activity. We also compared the cardiogenic potential of the natural products to that of the Wnt activator Chir99021 (Supplementary Fig. S3B, C).
From this experiment, we observed that the effect of the natural compounds on cardiac cell differentiation was comparable to that of Chir99021. The chemical-treated cardiogenesis was further enhanced when the natural compounds were treated with Chir99021 (Supplementary Fig. S3B, C). These results support that both natural compounds are Wnt signaling activators.
Optimization of cardiac differentiation by modulating Wnt signaling
Our results clearly support that Wnt signaling activation upon natural compound treatment in the EB phase is the main cause of enhanced cardiomyogenesis. Therefore, we further hypothesize that adequate suppression of Wnt signal in the condition of natural compound treatment can further facilitate cardiac differentiation. To test this possibility, we explored the effect of XAV939, a well-characterized Wnt inhibitor [20], on the natural compound-derived cardiogenesis by treating XAV939 right after activating the Wnt signal with natural compounds (Fig 6A).
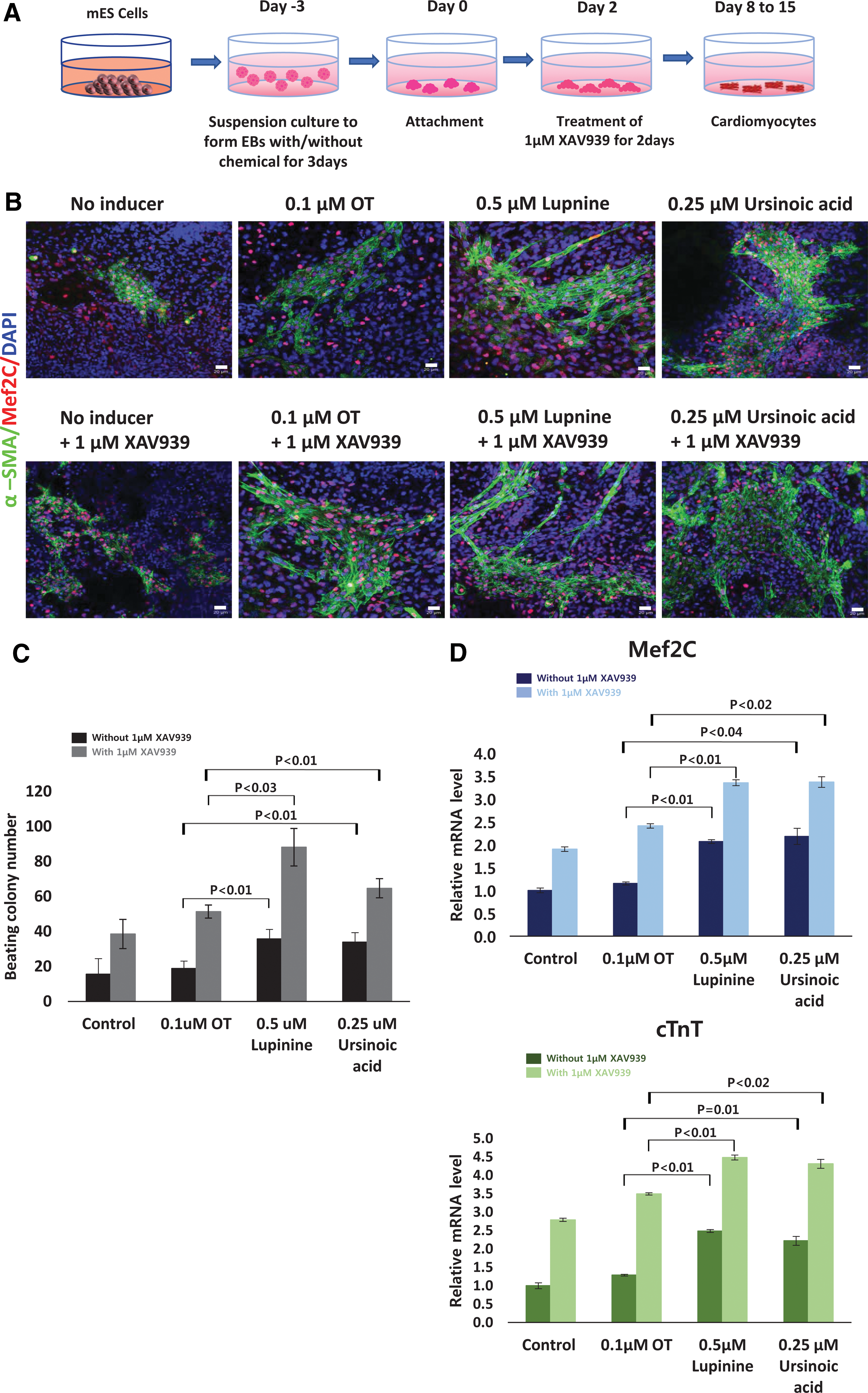
Cardiac differentiation using natural compounds and a Wnt inhibitor.
Although natural compounds alone enhanced the efficiency of cardiogenesis, the efficiency was enhanced dramatically when combined with XAV939, as depicted in immunostained cardiac colonies (Fig. 6B). This phenomenon also aligned with the observation that at least twofold higher beating colonies were obtained when XAV939 was cotreated with natural compounds compared to the condition without XAV939 treatment (Fig 6C). Among various treatment conditions, lupine in combination with XAV939 led to the highest number of beating colonies, which were 4.5-fold higher than those differentiated in the condition of oxytocin alone (Fig 6C).
We further checked the transcript levels of cardiac-specific markers, Mef2C (middle-stage marker) and cTnT (mature marker), to confirm the effect of XAV939 (Fig. 6D). Consistently, inclusion of XAV939 to the cardiac differentiation protocol using the natural compounds resulted in a higher expression of the markers (Fig. 6D). All of these data suggest that the best strategy of cardiogenesis is the facilitation of efficient mesoderm commitment driven by natural compounds, as Wnt pathway activators, followed by differentiation to cardiac cells through the suppression of Wnt signal.
Discussion
The limited regeneration potential of adult cardiomyocytes reduces their capacity to heal from many cardiovascular disorders that require tissue regeneration. To develop cost-effective, scalable, and highly efficient differentiation strategies to produce cardiomyocytes for use in stem cell-based therapies, we screened a natural compound library and identified chemical inducers that can enhance cardiomyocyte differentiation. For this purpose, we created a cardiac-specific screening platform in mouse pluripotent P19 cells (Cherry19 cells), which provided us with a reliable and efficient readout system for determining differentiation efficacy.
Use of this reporter-based cell line not only allowed us to forgo cumbersome immunostaining-based determination of cardiac commitment in the screening stage but also enabled us to reduce the batch-to-batch variance of differentiation capacity depending on the potency of the starting cells. We validated this system by identifying several candidate molecules that have the capability of enhancing the differentiation of P19 and mouse ES cells into cardiomyocytes.
Among the hit compounds, lupinine and ursinoic acid were chosen for further functional study, and their efficacy was accessed using various robust methods. Most importantly, the number and size of synchronously beating colonies induced by lupinine or ursinoic acid treatment were significantly enhanced in comparison to colonies produced by oxytocin treatment or without treatment. In agreement with this observation, treatment of each compound resulted in enhanced expression of cardiac-specific markers in both Cherry19 and mES cells.
Lupinine, an alkaloid naturally occurring in the seeds of Lupinus luteus L. and other Lupinus species, is known to reduce the effect of ethanol anesthesia [31,32]. Lupinine also inhibits cholinesterases [33,34] and immunoactive constituents in blood [35]. Ursinoic acid naturally occurs in the roots of Angelica ursine [36], but has not been annotated for any therapeutic applications. Because the effect of these two compounds on promoting cardiomyocyte differentiation was previously unknown, our results attributed a new role for lupinine and ursinoic acid in cardiomyogenesis and may help develop a new efficient protocol for cardiomyocyte differentiation.
When we performed the chemical screening, we concentrated on identifying chemicals that could induce the first phase of differentiation, which entails cardiac mesoderm commitment. We reasoned that efficient mesoderm commitment would not only help achieve efficient cardiac differentiation but also open up the possibility of effectively generating the cells similar to CPCs. By virtue of stemness, CPCs are capable of self-renewing and thus can give rise to a large pool of cardiac cells, which are beneficial for transplantation studies [37]. Accordingly, we observed a dramatic enhancement of the CPC markers, PDGFRa and Flk1 [37], especially upon lupinine treatment.
It was previously reported that initiation and maintenance of CPCs require complicated culture conditions [37,38], for example, BMP4, Wnt agonists, activin/nodal, or a combination of Lif and Wnt agonists [37] at the optimized concentrations in serum-free media. However, in this study, we confirmed that a single compound, lupinine, can induce high levels of CPC markers. Although it is necessary to perform a more extensive study to characterize the self-renewal properties, maintainability and expandability of the cells generated in the presence of lupinine, our study provides an example of a strategic approach that can be utilized to obtain more efficient conditions for obtaining CPCs. Therefore, this study not only demonstrates that a single chemical inducer can replace complicated media containing a variety of cytokines and signaling modulators to achieve efficient cardiac commitment but also provides enormous prospects for obtaining cardiac cells with progenitor properties.
The intrinsic question in this study is how these compounds contribute to enhancing cardiomyocyte differentiation. We observed efficient induction of Wnt activity as well as enhancement of Wnt3a transcripts in the cells treated with the natural compound inducers compared with those in the control cells (no treatment) and oxytocin-treated cells. The Wnt signaling pathway is one of the most important pathways for the primitive streak and subsequent cardiac mesoderm generation, which explains why we obtained efficient cardiac commitment upon chemical treatment. Interestingly, we also observed that our compounds not only rapidly upregulated Wnt3a but also suppressed Wnt3a expression earlier than control cells and oxytocin-treated cells (Fig. 5). This result suggests that Wnt3a suppression, which is necessary for proceeding to the next phase of cardiac specification, happens in oxytocin-treated or untreated cells at a later stage than the lupinine- or ursinoic acid-treated cells.
Based on these observations, we concluded that lupinine- or ursinoic acid-treated cells initiate early cardiac mesoderm commitment and also proceed to the next phase of cardiac commitment earlier. We also concluded that regulation of Wnt signaling in the order of time is thus crucial for efficient differentiation, as proposed in a working model explaining the overall mechanism for the natural compound-induced enhancement of cardiomyocyte differentiation (Fig. 5H). Based on our observation and the proposed working model of the compounds, we rationally designed an optimized cardiogenesis protocol consisting of the initial Wnt activation stage with our natural compounds and the subsequent Wnt suppression stage by using XAV939 (Fig 6). This optimized strategy has been verified by the significantly enhanced cardiogenesis efficiency that we observed compared to the protocol with a natural compound alone.
The next generation of cell differentiation protocols will largely rely on strategically designed methods for efficient lineage generation. The study presented here provides important and effective ways for modulating initial phases of mesoderm commitment. Thus, this study not only confirms the significance of Wnt signaling in mesoderm and cardiac commitment but also provides valuable information for designing an efficient strategy for cardiac generation from pluripotent cells. In another aspect, this study also suggests that a chemical-based differentiation strategy can be a simpler alternative to existing complicated protocols for cardiac differentiation, which usually depend on the inclusion of multiple cytokines and pathway modulation.
Footnotes
Acknowledgments
This study was supported by National Research Foundation of Korea (NRF) grants (2017M3A9B4065374 to K.K.K. and 2015R1D1A1A01058188 to D.D.), and KRF program (2015H1D3A1062463) to K.K.K. and D.D.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.