
Abstract
Some types of cells, if not all, that undergo signal exchanges in culture need to contact other cells for various reasons, such as cell-to-cell contact for growth inhibition. However, signal exchanges by cell-to-cell contact before proliferation have never been reported. Using time-lapse recording, we discovered the emergence of several astonishing cell-to-cell contact modes in bone marrow-derived mesenchymal stem/stromal cells (MSCs) before the cells divided. When the cells contacted with another, a huge temporary synapse-like structure formed for molecule exchanges; a cell-tissue particle was taken in by a recipient cell; two cell membranes formed infusion-like structure for a short time; and even a 20-μm long and 5-μm wide cell tail was grafted to another cell. A total of 87% of cells underwent cell-to-cell contact before dividing. After epidermal growth factor–green fluorescent protein (EGF-GFP) vectors were transfected into MSCs and the cells were cocultured with unmanipulated MSCs, the unmanipulated MSCs took in EGF-GFP particles from EGF-GFP expressed MSCs, immediately increased in mitogen genes, and then divided. These results suggest that cells which may lack signal molecules may need to obtain these molecules from other cells through various types of cell-to-cell contact, as mentioned above. Our study provided valuable information to better understand the behaviors of cell-to-cell contact and communication before mitosis.
Introduction
Communication between cells is important to maintaining cell function, to exchanging signal molecules, and to changing the cells' properties. Current studies have demonstrated that PC12 cells can produce a tube-like structure, called a tunneling nanotube, that links two cells in culture [1] and allows molecules and organelles to traffic between the cells. The term plasmodesmata is used to describe gap communications between neighboring plant cells [2]. Spees et al. [3] demonstrated in culture that mitochondria from bone marrow-derived mesenchymal stem/stromal cells (MSCs) were transferred to rescue damaged alveolar epithelia; Islam et al. [4] then demonstrated that the transfer of mitochondria occurred in vivo through connexin 43–containing gap junctional channels. Moreover, ions, DNA, RNA, microRNA, lipids, proteins (including pathogenic proteins), and organelles have been reported to be transported through gap junction or nanotube between two cells [5 –8]. However, whether there are other ways that cells can communicate to exchange molecules, in which one cell may be present in excess and the other may lack, is not known. In this project, we took advantage of the ability of MSCs to generate colonies when plated at low density and time-lapse recording to monitor how MSCs communicate and exchange signal molecules. Our results, presented below, indicate that communication and exchange of materials between MSCs occurred in several ways.
Materials and Methods
This study was approved by Institutional Review Board, Tulane University and Louisiana State University Health Sciences Center.
Bone marrow and MSC preparation
Human bone marrow MSCs were obtained from the iliac crest of normal adult donors. Aspirates of 20 mL were mixed with Hanks' balanced salt solution (HBSS; GIBCO) at 1:1 and layered over about 10 mL of Ficoll (Ficoll-Paque; Pharmacia). After centrifugation, cell layers were separated. The mononuclear cell layer was removed from the interface and suspended in HBSS. The cells were then centrifuged and resuspended in 25 mL complete medium (Minimum Essential Media, 20% fetal calf serum [FCS], and 2 mM
Time-lapse recording
We used a Nikon microscope (ECLIPSE Ti2, inverted) with a motorized stage, computer-controlled lens change, and a built-in incubator which supplied humidified 5% CO2 at 37°C for living cell recording. Human bone marrow MSCs were cultured on a 12-well plate, and this plate was incorporated into the incubator. The computer and ImagePro (5.1) software directed the microscope to take a series of pictures that covered an area of 2 by 2 mm and recorded the coordinates of each picture and montaged the pictures together. The procedure was repeated every 5 min for as long as needed for the experiments. ImagePro can replay these pictures in a continuous mode like a movie.
DiD- and DiO-stained cells and cell sorting
DiD and DiO are fluorescent lipophilic dyes that can anchor in a lipid structure on the cell membrane. DiD emits red and DiO emits green fluorescence. DiD and DiO crystals were dissolved in 100% ethanol at 1 mg/mL. One million cells were stained with 1 μL DiD or DiO. After incubation at room temperature for 20 min, the cells were centrifuged down. Supernatant was discarded. The cells were washed with PBS once; DiD-stained cells were then mixed with DiO-stained cells, which were cultured on a six-well plate. At days 0, 1, 2, 3, 4, and 5, the cells were harvested for flow cytometry. Flow cytometer (FACsort; Becton Dickinson) counted red, green, and red-green cell numbers.
Epidermal growth factor–green fluorescent protein vector and expression imaging
Epidermal growth factor–green fluorescent protein (EGF-GFP) coding sequences with Xbal1 and Spe1 were amplified by PCR (Pfu DNA polymerase; Stratagene) from EGF-GFP plasmid (Cat No.: RG230667; Origene) using the primers (TGCTCTAGAGCAATGCTGCTCACTCTTATC; CGGACTAGTCCGTTACTTGTACAGCTCGTC). Then the sequence was installed into AAV2 vectors (Catalog No.: AAV5XXA-1; System Biosciences), which were transferred into Escherichia coli TOP10 cells (C404010; Life Technologies Corp.) for selection. After sequencing, the vectors were purified (QIAprep; Qiagen) and used for MSC transfection. The transfected cells were cocultured with unmanipulated MSCs. The cells were imaged with fluorescent field and bright field at the same site for analysis.
Single cell gene analysis
Using coordinates for cell location, which the software recorded for each picture, the computer could trace the cell in the well at any time. Thirty minutes after the cell took in a primer, which was a small detritus from an axon or membrane, or made contact with another cell, the cell's cytoplasm was collected with a negative-pressure glass patch clamp microelectrode pipette, which was used as a substrate for reverse transcription with random primers [9]. The products from contacted cells were subjected to quantitative PCR to assess alterations of mitogenic genes Ras, ERK1, and ERK2, compared with the genes in the control cells that did not take a primer or make contact with another cell, and measured as fold change in gene alterations.
Statistical analysis
Data are represented as mean ± standard error of the mean. Statistica 13.1 was used for data analysis. The gene changes between the cells that made contact with other cells and uncontacted cells were analyzed with a two-sided, two-sample t-test. When the sample size for each group was 6, analysis of variance (ANOVA) had 80% power to detect an effect size (the variance of the means divided by the within-group variance) of 0.2462 in percentage change, assuming a type I error rate of 0.05.
Results
Cell touch mode
Cell-to-cell contact in culture has been reported by many authors. However, the reason why cells want to contact others is not fully understood. The ability of bone marrow-derived MSCs to expand after plating at clonal densities provides an opportunity to examine the early propagation of individual cells in culture [10,11]. Our observation indicated that MSCs in low density cultures made frequent contact with each other. We observed that apparently a given cell did not touch just any cell, but looked for a specific cell to touch. The cell moved several 100 μm, some moving even up to 2,600 μm, to look for a favorite cell, then touched that cell for 50 min. The two cells then separated, and cell B divided into two cells (Fig. 1A, 1–6).

MSC contact modes in culture.
Sometimes, a cell did not contact another cell directly. As Fig. 1B revealed, a cell (B2) moved toward a primer that was left by another cell (A) from its long projection, and cell B2 engulfed the primer. After 45 min, B2 divided into B2a and B2b (Fig. 1B, 1–6).
Cell-to-cell communication can also have a more sophisticated mode, not just engulfing a molecule, but part of a cell. Figure 1C shows that cell A contacted the tail of cell B with its head part. After this contact, the tail from cell B was implanted and incorporated into cell A. Then, cell A divided into A1 and A2.
Detailed contact procedure
Differential interference contrast (DIC) microscopy can provide better resolution of an image than can phase contrast microscopy. With the DIC lens, the activity of two cells in culture was recorded every 5 min. The images clearly showed the process by which the cell head of one cell came into contact with the tail of another cell (Fig. 2). The vesicle attached on the tail of the cell (black arrow in photograph at time 0:00 to 2:20) eventually moved to the head of the other cell after 2 h, while the vesicles on the head (black arrowhead, white arrowhead, and white arrow) moved to the tail. The tail touched the head, and the main part of the tail that touched the head became thinner over time. At 2:05, this part fused with the head partially, and the tail broke from the cell body, indicated by the white star in the photograph. Then this part of the tail fully fused with the head, and the two cells separated. These findings suggest that a cell tail and vesicles can be exchanged simultaneously between two cells.

Time-lapse image recorded with DIC microscopy. The pictures were taken every 5 min for vesicle exchanges between two cells (the key pictures are shown here). At 0:00, one vesicle was on the upper cell membrane, indicated by an arrow. One vesicle was on the lower cell, indicated by an arrowhead. At 0:45, the head of the lower cell contacted the tail of the upper cell. At 1:05, another two vesicles appeared on the lower cell membrane, indicated by a white arrow and arrowhead, respectively. The vesicle indicated by a black arrowhead moved toward the upper cell gradually. From 1:40 to 2:20 the vesicle moved onto the upper cell. The two vesicles indicated by white arrows and arrowheads moved onto the upper cell also at 1:40 to 2:00 and were then invisible, suggesting that they merged into the upper cell. At 1:40 to 2:00, the tail membrane of the upper cell was indented, indicated by asterisks. At 2:05, the tail was broken and had fused into the lower cell. The vesicle indicated by a black arrow moved onto the lower cell. DIC, differential interference contrast.
Multiple fusions between two cells
A fusion-like structure between two cells is another mode of cell-to-cell communication. We observed that two cells could form a fusion-like structure at two sites on the cell bodies (Fig. 3). Cell A had a wide head, and cell B had an unobvious head. The two cells moved toward each other. At two places indicated by a white star, the two cell membrane boundaries could not be seen between the cell membranes at these sites. A shallow split formed by the two membranes remained between the two sites. After the two cells were in contact for 50 min, they separated, and then cell B divided into two cells.

Two cell membranes fused together.
DiO and DiD stained cell coculture
To determine whether MSCs making contact with other cells is necessary for cell growth in culture, we used fluorescent lipophilic DiO (green) and DiD (red) dyes, which can anchor in a lipid structure on the cell membrane, to stain two aliquots of the cells, respectively. Then the DiO- and DiD-stained cells were cocultured together. The cells were harvested at days 0, 1, 2, 3, 4, and 5 after coincubation for flow cytometry so that the number of cells with both green and red fluorescence could be counted. At day 0, the cells were cultured together for 2 h; 0.4% of the cells had double color. At day 1, 16% of the cells had double color. At day 5, the number of cells with double colors had increased to 42% (Fig. 4A, B), suggesting that cell membrane materials are frequently exchanged between cells.
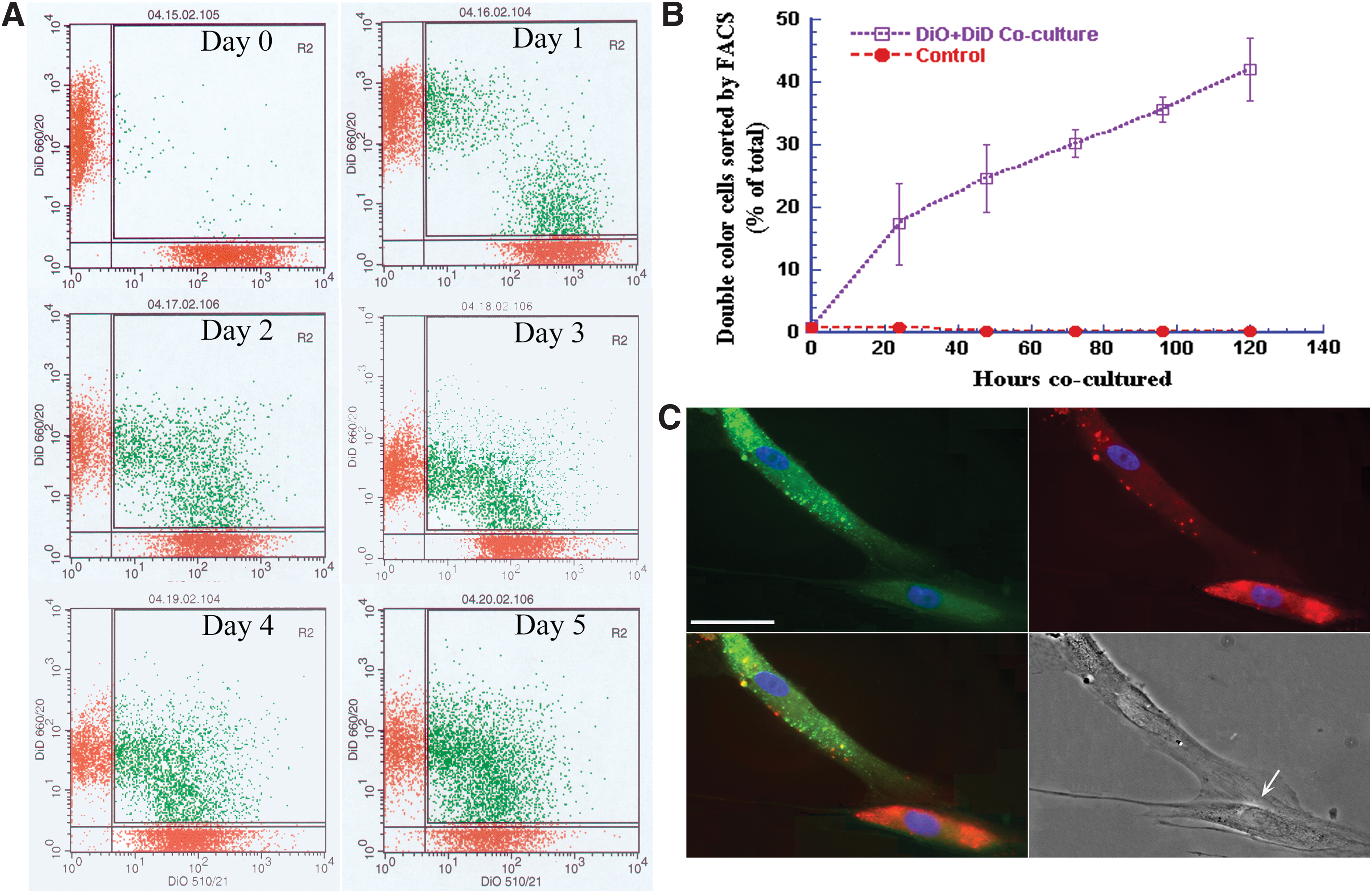
Membrane exchanges between DiD- and DiO-labeled cells in culture. DiD (red) and DiO (green) dyes were used to stain two groups of MSCs. The cells were mixed together for culture.
With fluorescent and DIC bright field pictures, we observed that a green cell tail had come in contact with a red cell body. In the green cell body, several red vesicles were seen; green vesicles were also observed in the red cell. On the merged image, several vesicles had become yellow, suggesting that vesicles had exchanged between the two cells. The DIC picture revealed that a huge synapse-like structure with pre- and postmembrane and narrow luminous space between the pre- and postmembranes had formed between the tail of the green cell and the body of the red cell. A bright luminescent narrow area was shown that could have been a gap between the two cells (Fig. 4C, arrow, bright field), indicating that such a structure can also exchange materials.
We examined 50 cells to determine whether the cells contacted other cells, whether the cells proliferated after contact, and how far the cells moved to find a cell to touch (Table 1). The results suggested that most of the cells contacted one or more cells and that they divided after contact.
Human Mesenchymal Stem/Stromal Cells Contacting Other Cells, Dividing, and Moving
EGF vesicle transferring
An EGF-GFP (EGF-GFP as a fusion protein) vector was used to transfect MSCs with lipofectamine. GFP acted as a visible indicator for EGF expression in the cells. When EGF-GFP-transfected cells were cocultured with unmanipulated cells, a tiled picture at the same area was taken by a fluorescent lens and a bright field DIC lens (Fig. 5). The pictures were then merged for analysis. The fluorescent picture showed a green cell with a vesicle. When the cell grew larger, the green vesicle was moving along its projection. The bright field picture showed that two spindle cells were moving toward each other, then the two cells came in contact with a partial membrane fusion-like structure. After the two cells separated, the vesicle was transferred from the left cell to the right cell. Then the right cell divided into two cells. The left cell had also become round, a sign which indicates that the cell was about to divide. When the fluorescent pictures were merged with the bright field pictures, a green cell (left) and a gray cell (right) had moved closer and then contacted each other. During this contact, the green EGF vesicle was transferred from the green cell to the gray cell; the gray cell then divided into two cells (Fig. 5).
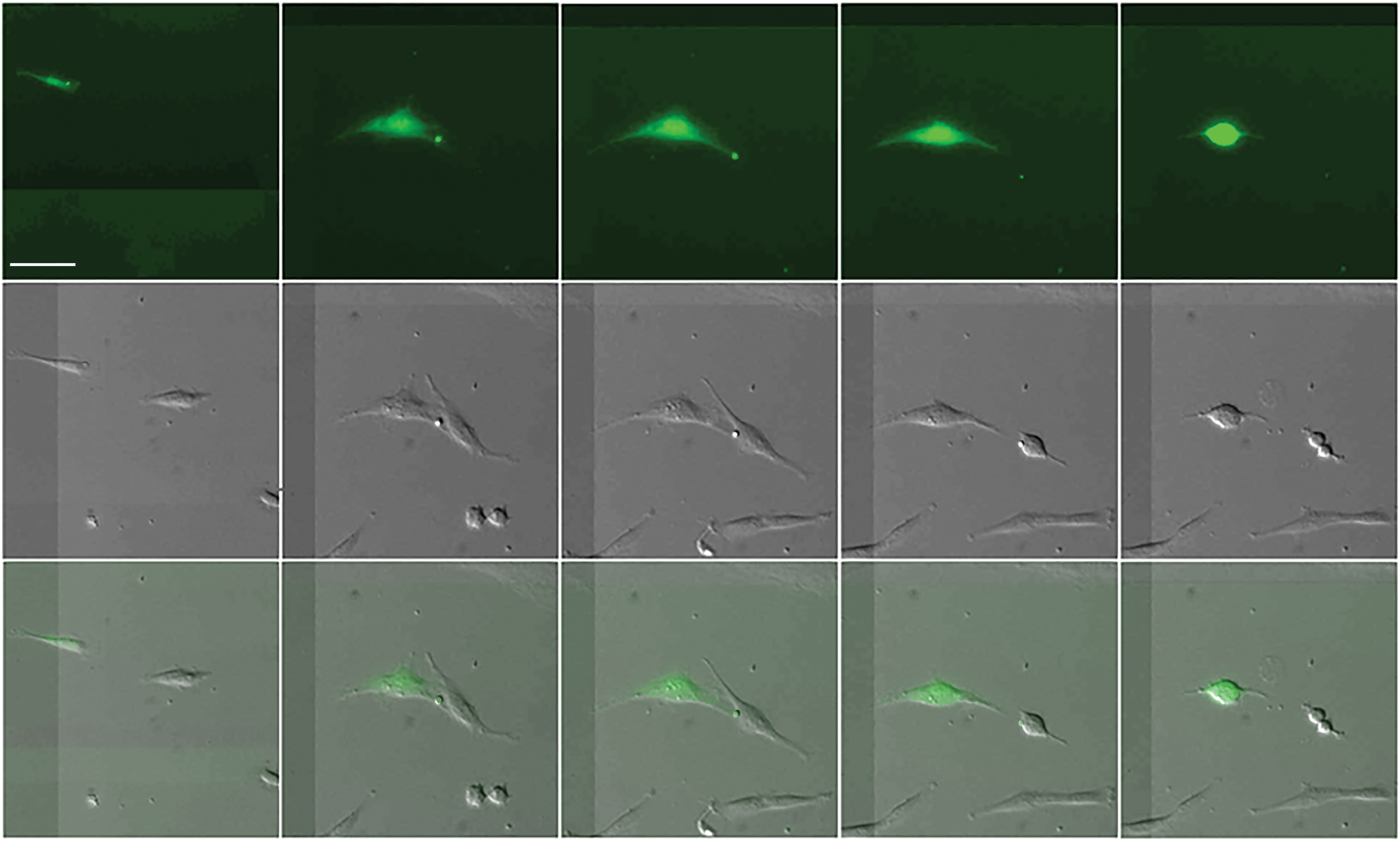
A cell that expressed EGF-GFP contacted an untreated cell. After EGF-GFP transfection, the cell was cocultured with a normal MSC. The images at the same location were obtained with a fluorescent field and with a DIC bright field and were then merged. In the fluorescent field image, a cell was green and carried a green vesicle. In the bright field images, two large long cells were displayed. On the merged image, one cell was green, and the other was gray. The green cell moved toward the gray cell. Then the two cells contacted tightly, and the green vesicle, which was carried by the green cell, was transferred to the gray cell. After a while, the gray cell divided into two cells. The green cell also rounded up, indicating that the cell was about to divide. These findings suggest that the gray cell received EGF and then divided. Scale bar represents 30 microns. EGF-GFP, epidermal growth factor–green fluorescent protein. Color images available online at
Single cell gene analysis
We observed that a cell divided after it took in the vesicle or exchanged material from another cell. Using coordinates for cell location, which was recorded by computer, we traced the cell that took in the vesicle or made contact with another cell; then we collected the cell's cytoplasm 30 min later with a negative pressure glass patch clamp microelectrode pipette, which was used as a substrate for reverse transcription with random primers. With the same method, we traced the cells that did not touch other cells and then collected the cells as a control for gene changes. We then used quantitative PCR to assess alterations of mitogenic genes Ras, ERK1, and ERK2, compared with the genes in control cells. The results suggested that Ras, ERK1, and ERK2 were significantly increased after the cells took in the vesicles (Table 2). EGF binding to the EGF receptor results in protein tyrosine kinase activation to activate and increase Ras. Ras belongs to a GTPase family, which further activates ERK1 and ERK2 to enhance cell proliferation. The asterisks in the Table 2 indicate increased gene fold significance (*P < 0.05, one-way ANOVA, two sample t-test).
Increased Mitogeneic Genes After Cell-to-Cell Contact
The asterisks indicate increased gene fold significance (P < 0.05, one-way analysis of variance, two sample t-test).
These results indicate that material exchange between cells has multiple contact modes and that these exchanged materials can enhance cell proliferation and possibly even alter cellular properties.
Discussion
Evidence suggests that cell-to-cell communication happens in most kinds of cells, if not all. Such communication occurs not only between cognate cells but also between dissimilar cells [12]. In this study, we focus on cell-to-cell communication before mitosis between human bone marrow-derived MSCs, which possess some of the properties of stem cells and have the potential to repair tissue damage and to treat various diseases [13 –16]. Gaining an understanding of the significance and mode of MSC communications would advance our knowledge in the potential applications of MSCs for disease treatment.
Exchange of molecules between cells has been reported. Exchanged molecules may be used for signals that a cell needs for a specific purpose. As mentioned in the Introduction, these signal molecules can be DNA, RNA, ions, and organelles. However, whether molecular exchanges occur between two cells randomly is not clear from previous studies. Our time-lapse recording revealed that a MSC did not contact another cell randomly. The MSC moved in culture and passed by many cells; even when the cell encountered other cells head-to-head, the cell did not make contact until it found a specific cell (Fig. 1A), and then the two cells touched each other. In a similar case, a primer lay in culture as several cells passed by and did not take it in, but a cell came to the primer and took it in. After a while, the cell divided into two cells (Fig. 1B). We observed a MSC that moved 2,600 μm to find a specific cell to contact (Table 1). Although we currently do not know what it is about a cell that attracts another cell, we speculate that something such as a molecule attracts the cell or a few molecules attract the two cells mutually. This will be discussed further below.
The cell-to-cell communication for signal molecule exchange revealed by our time-lapse imaging occurred in the following contact modes: (1) A cell contacts another cell, head-to-head or head to body (Fig. 1A); (2) A cell engulfs a primer left from another cell (Fig. 1B); (3) A cell tail grafts to another cell body (Fig. 1C); (4) The membranes of two cells form a fusion-like structure for molecular exchange at one site or multiple sites (Figs. 2 and 3); and (5) A synapse-like structure forms between two cells (Fig. 4C). The membrane fusion-like structure and a part of a cell graft for molecular exchange have never been reported before. These contact modes can transfer large amounts of RNA, ions, proteins, and organelles, suggesting that after material exchange, the properties of this cell may be altered. Cancer cells may spread by these approaches to turn healthy cells to tumor cells.
Why do cells need to make contact with other cells? The purpose of this contact may be to obtain molecules that the cells lack. This speculation comes from the finding that cells divided after making contact with other cells. With DiO- and DiD-stained cells in coculture, 42% of the cells displayed double colors, suggesting that these cells had exchanged their membrane components (Fig. 5A). When MSCs were cocultured with renal tubular cells, cytoplasm and organelle transfer between the two kinds of the cells was also observed [17].
To confirm that a MSC needs an extracellular signal to promote proliferation, EGF-GFP-labeled MSCs were cocultured with unmanipulated MSCs. With bright-field images and fluorescent images at the same imaging frame, the EGF-GFP vesicle was taken in by the MSC. After that, the cell divided into two cells. The result confirmed that EGF molecules may attract a cell that needs extracellular EGF to enhance cell proliferation, as one of the cell-to-cell contact mechanisms. Of course, extracellular EGF molecules are not the only molecules that can promote proliferation; many other types of molecules can also enhance cell proliferation, such as fibroblast growth factors, insulin-like growth factors, and transforming growth factors [18 –20]. However, an EGF-containing vesicle is much larger than an EGF molecule, and it seems impossible for such a vesicle to bind to an EGF receptor. Whether the role of the EGF vesicle when it is taken in is similar to that of an EGF molecule when it binds to an EGF receptor is not clear. With a single-cell PCR technique, we selected cells that took in a vesicle or made contact with other cells for mitogenic gene analysis. Our results confirmed that mitogenic genes Ras, ERK1, and ERK2 were significantly increased in the cells that took in an EGF vesicle or made contact with other cells, compared with cells that did not take in a vesicle or did not make contact with other cells (Table 2). These results suggest that the cells that took in a vesicle or make contact with other cells significantly increased mitogenic genes to promote cell proliferation.
The cycle of MSC proliferation is normally 7–16 h, depending on plating density and the donor's age [21]. Our observation suggests that cell-to-cell contact for molecule exchange occurs most at 1 to 2 h before the cell divides (Figs. 1 –3). The time corresponds to the later G2 phase, prophase, and mitotic phase of mitosis [22], although it has been stated that growth factors regulate cell proliferation during the G1 phase [23]. This discrepancy needs to be further studied.
The transient cell contact before mitosis observed in this study suggests that the early replication and differentiation of MSCs in culture require cell-to-cell signaling similar to the cell-to-cell signaling that determines cell fate in embryonic development and elsewhere. The cell-to-cell signaling after extracellular molecular attraction further involves the membrane-bound ligands Notch, Hedgehog (Hh), Wnt, [24,25] connexins, and actins, which we did not plan to study in this project. We focused on what attracted two MSCs to contact before mitosis. Complexity of cell-to-cell communications between MSCs is far beyond our understanding at current stage. As our results clearly indicated, even part of a MSC can be grafted into another cell. If a part of a tumor cell is grafted to a healthy cell, can the normal cell turn into a tumor cell? Tumor RNA and organelles that have a specific structure, such as mitochondria [26], exist in the transferred part; therefore, the tumor's RNA and organelles may have the ability to turn the healthy cell into a tumor cell. If so, this could be favorable to tumor spread.
Footnotes
Acknowledgments
The authors thank Dr. Darwin Prockop for his precious time to read and to comment on this article. They also thank Ms. Tamara Locke for her editing assistance. This project was supported by LSU transitional grant and Helen Buchanan and Stanley Joseph Seeger Research Endowment to Dr. Abdi.
Author Disclosure Statement
The authors have no potential conflicts of interest.