
Abstract
In vitro embryo twinning can be used to increase the number of the human embryos available for production of human embryonic stem cell (hESC) lines. The aim of this study was to generate hESCs following the production of the twin embryos by in vitro embryo splitting procedures. In total 21 chromosomally abnormal (three pronuclei) embryos underwent in vitro embryo twinning and were allowed to develop to the blastocyst stage. As a result, 42 twin embryos were obtained, of which 24 developed to blastocyst stage. Using micromanipulation technique, the zona-free blastocysts were recovered and plated onto mitotically inactivated Yazd human foreskin fibroblast (Batch18; YhFF#18) feeder layers in microdrops. After 3 to 5 days of blastocyst culture onto human foreskin fibroblast feeder layers, the hESC-like outgrowths were passaged onto new feeders in microdrops. The initial outgrowths of hESC-like cells were generated, and cells were proliferated, passaged, and some of them expressed hESC and trophoblastic markers; however, no cell lines were established. This might be due to the low cell number and poor quality of inner cell mass within these twin blastocysts. In vitro embryo twinning by increasing the number of the human embryos could be useful in the future for the generation of new pluripotent stem cell lines. However, the challenge remains to optimize the methods.
Introduction
The conventional method for the derivation of human embryonic stem cells (hESCs) is plating the inner cell mass (ICM) on a feeder layer of inactivated mouse embryonic fibroblasts (MEFs) [1]. It was reported that the success rate in hESC derivation depends on human embryo quality, although cell lines have been generated from morphologically poor embryos [2]. One of the challenges for the generation of hESCs is the number of embryos which are donated for the derivation of hESCs.
Due to the totipotent nature of the blastomeres of the early embryos [3], in vitro embryo splitting, using blastomere biopsy from the human cleaving embryos, could be an alternative option to increase the number of the available embryos in assisted reproductive technology (ART) programs. In vitro embryo splitting to two or more embryos would have clinical advantages for women at an advanced age [3] and patients termed as “poor responders” [4], to avoid additional retrieval cycles by providing sufficient embryos for subsequent transfers [5]. Moreover, numerical increases of acceptable embryos for embryo transfer enhance the chance of pregnancy in ART cycles [6]. In addition, in vitro embryo splitting may be able to produce donor embryos for research purposes [7].
Recently, reports suggested that the morphologically viable blastocysts can be obtained following splitting of human cleaved embryos [7 –9], although the quality assessments of the embryos created in such way have been limited. It was shown that twin blastocysts following in vitro splitting were smaller than control group; however, the expression analysis of NANOG by immunofluorescent (IF) staining and confocal microscopy proved the presence of the ICM in these blastocysts at day 6 [8]. Moreover, Noli et al. [7] have detected single NANOG only-positive cells in twin blastocysts, indicating the potential of ICM formation, although the ICM in these blastocysts was considered as poor quality cells.
The present study is reporting the attempts for the derivation of hESCs from twin blastocysts following human in vitro embryo twinning. Derivation of hESCs would confirm the existence of ICM in developed blastocysts following in vitro splitting.
Materials and Methods
Consent and donation of embryos
Embryos were obtained with full informed consent from 17 couples attending a clinic for assisted conception (Ethics Committee reference number: IR.SSU.MEDICINE.REC. 1395. 93/Part I). Triploid zygotes showing three pronuclei (3PN) after fertilization assessment of in vitro fertilization (IVF)/intracytoplasmic sperm injection cycles were donated for this study. These genetically abnormal zygotes were discarded during ART programs. 3PN zygotes were cultured in G-1™ v5media (Vitrolife, Sweden) and incubated with humidified atmosphere at 37°C, 6% CO2 and 5% O2 to six- to eight-cell stages. Good quality cleaved embryos with <10% fragmentation and blastomeres with regular size were selected for splitting.
In vitro embryo splitting
Before biopsy, to facilitate separation of blastomeres, the embryos were preincubated in microdrops of Ca-Mg-free culture medium (PGD medium; Vitrolife, Sweden) overlaid with mineral oil for 10 min at 37°C. A 35–40 μm hole was made in zona pellucida (ZP) using a 1480-nm wavelength infrared diode laser (OCTAX Laser Shot®; MTG, Germany). Half of the blastomeres were biopsied by aspiration using a micropipette with the inner diameter of 35 μm and inserted one by one into a previously prepared empty ZP. The source of empty ZPs was from the immature oocytes and discarding arrested embryos.
Embryo culture
After splitting, donor blastomere embryos (twins A) and recipient blastomere embryos (twins B) were carefully washed and cultured to G-2™ v5media (Vitrolife) covered with mineral oil under an atmosphere of 6% CO2 in air at 37°C. Development of twin embryos was followed using an inverted microscope (TE300; Nikon, Tokyo, Japan) every 24 h up to blastocyst stage and photographically documented daily.
Derivation of hESCs
Using the biopsy pipettes, the zona free whole blastocysts were aspirated through the hole of the ZPs (Fig. 1) and plated onto the 50 μL microdrops of the mitotically inactivated Yazd human foreskin fibroblast cell line 18 (YhFF#18) feeder layer and cultured with human embryonic stem (HES) medium [1] covered by mineral oil at 37°C at 5% CO2 in air. Under the stereomicroscope and using the pooled glass Pasteur pipette and mouth pipetting, the hESC-like outgrowths were cut into smaller pieces and transferred to the new microdrops of YhFF#18 feeder layer with HES medium. They were cultured at 37°C at 5% CO2 in air.
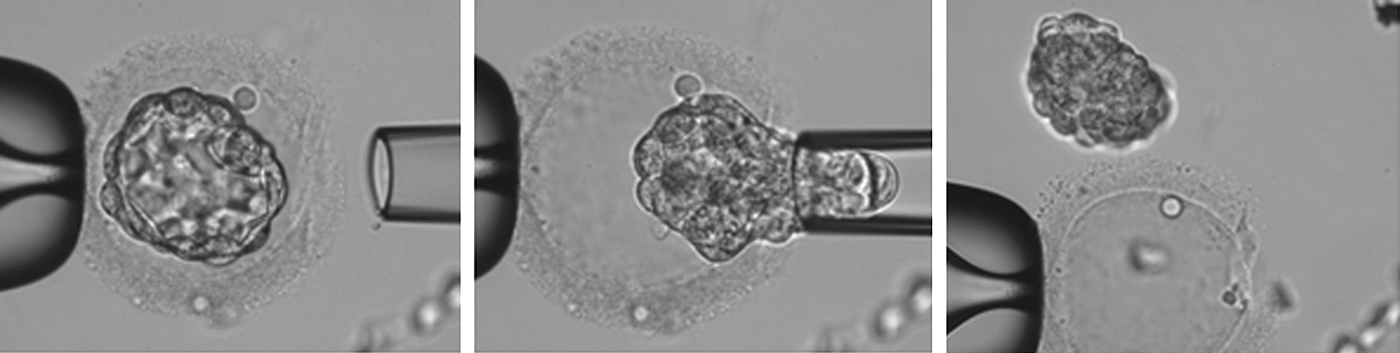
Aspiration of whole blastocyst from ZP using biopsy pipette. ZP, zona pellucida.
IF staining
IF was used to evaluate dual localization for the expression of SSEA1 (sc-21702; Santa Cruz Biotechnology) and TRA-2-49 (ab17973; Abcam, United Kingdom) of the hESC-like outgrowths following first passage (one donor and one recipient). IF was performed as explained elsewhere [10].
Statistical analysis
Statistical analysis was performed using the Statistical Package for the Social Sciences 20 (SPSS, Inc., Chicago, IL). Categorical variables were compared using Chi square. The significance level was 0.05.
Results
In vitro embryo twinning
A total of 21 triploid human embryos were manipulated in this study, of which 17 embryos were split at the 6- to 8-cell stage and 4 were split at the 10- to 12-cell stage (Table 1). After splitting, 42 twin embryos were obtained at cleavage stage. Embryo twinning at the 6- to 8-cell and 10- to 12-cell stage revealed that 55.9% and 62.5% of the embryos developed to the blastocyst stage, respectively (P = 0.38, Table 1; Fig. 2). In total, 24 blastocysts were generated after splitting of 21 embryos that 14 of them were twins (7 pairs).
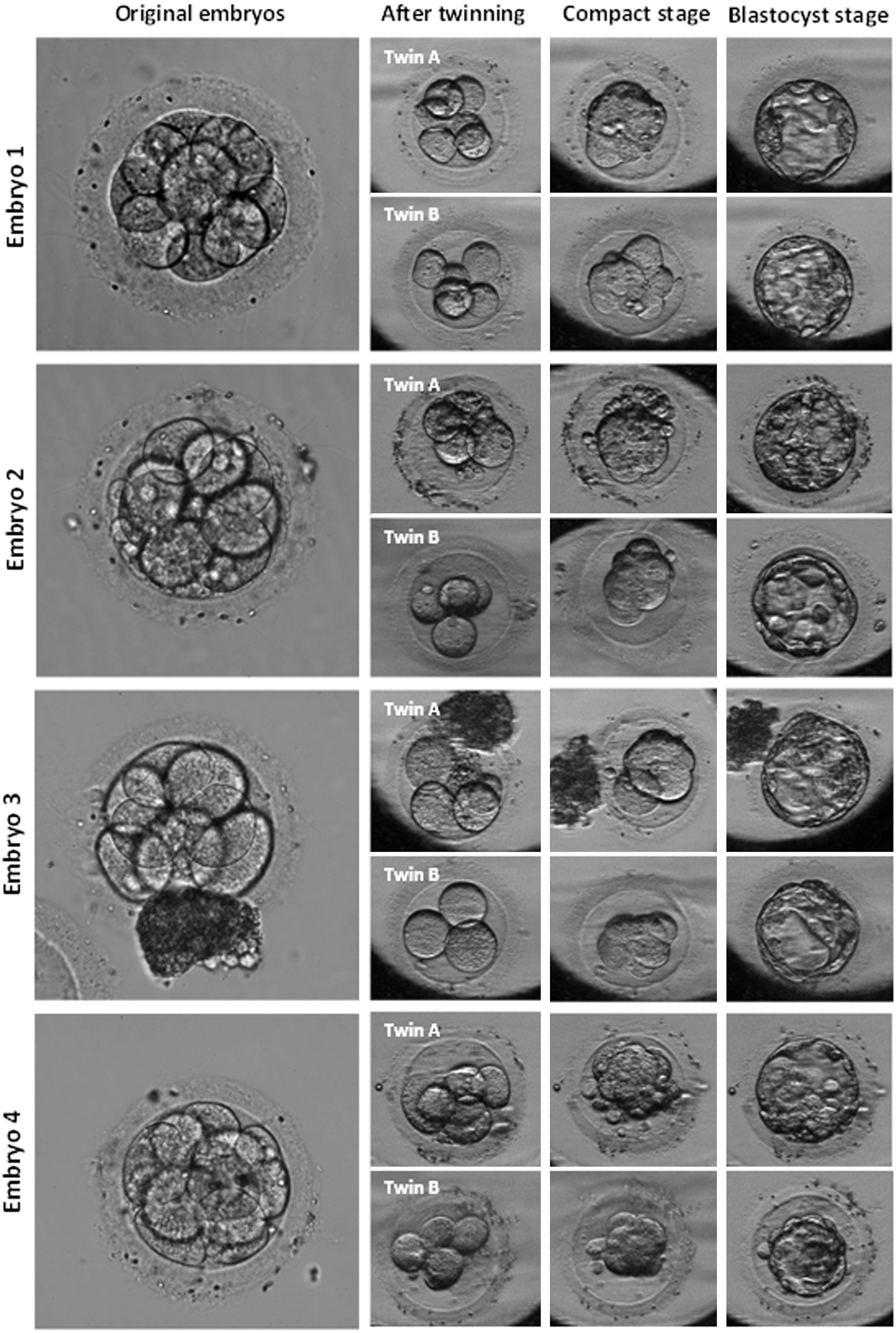
Developmental progress after in vitro twinning of cleavage embryos. Twin A: donor blastomere embryo, Twin B: recipient blastomere embryo.
A Summary of Blastocyst Development Outcome of Embryos Cultured Following In Vitro Embryo Twinning
Twin A: donor blastomere embryo; Twin B: recipient blastomere embryo.
P < 0.05; b P = 0.38.
Subanalysis data showed that from 24 blastocysts, 14 blastocysts were derived from donor blastomere embryos (twins A) and 10 blastocysts were derived from recipient blastomere embryos (twins B) (58.3% vs. 41.7%, respectively; P < 0.05). The detailed results regarding the blastocyst development following in vitro splitting embryos are summarized in Table 1.
Generation of hESC-like cells
From the total 24 zona free blastocysts following in vitro splitting, which have been plated onto the microdrops [1] of YhFF#18, 15 outgrowths were resulted, in which 11 of them were looking promising initially (Table 2). The cells morphologically resembling hESCs with high ratio of nuclei to cytoplasm and multinucleolus were observed (Figs. 3 and 4A). After 4 days following passage, a small number of hESC-like cells were observed in the middle of a multinucleus syncytiotrophoblast (Fig. 4B, C). However, from these 11 outgrowths, 6 of them were expanded enough to be able to be passaged further. Nonetheless, no sign of self-renewal was observed after passage 5, and cells become differentiated.

Initial outgrowths from twin embryos following embryo splitting (Twin A: donor blastomere embryos and Twin B: recipient blastomere embryos).
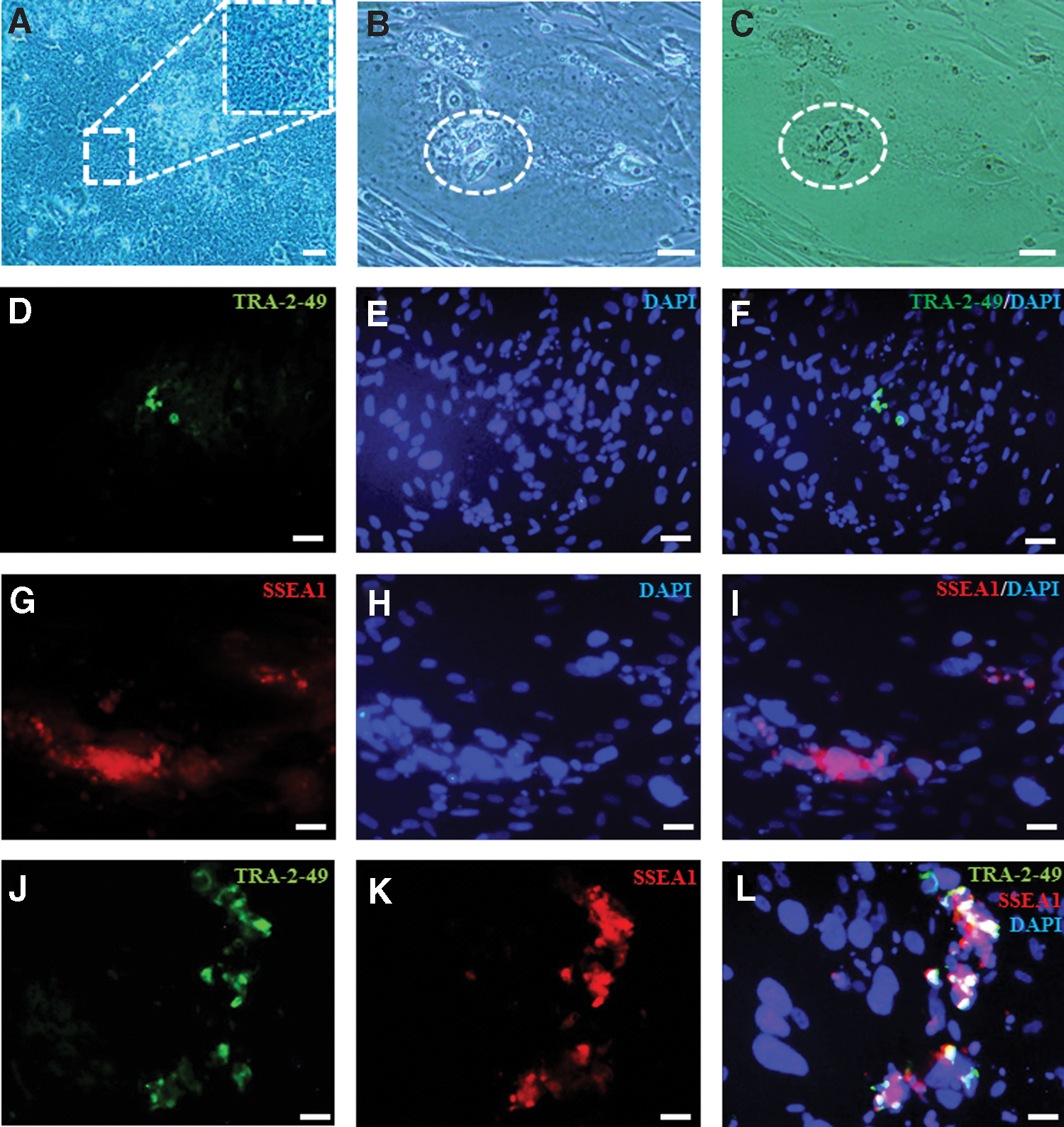
Morphology of hESC-like cells from in vitro twinning embryos
The Summarized Results of the Attempts Made for Generating Human Embryonic Stem Cells Following In Vitro Embryo Twinning
Immunolocalization of TRA-2-49 and SSEA1
While the small numbers of the cells were positive for TRA-2-49 (Fig. 4D–F), the majority of the cells were SSEA1 positive (Fig. 4G–I), which indicated the existence of small number of hESC-like cells and trophoblast cells. One of the outgrowths showed colocalization of both TRA-2-49 and SSEA1 (Fig. 4J–L).
Discussion
Generally, two pronuclei form after fertilization of an oocyte by sperm followed by normal diploid cleavage stage. However, abnormal PN occurrences, including 0PN, 1PN, and 3PN, have been seen in the IVF programs, of which 3PN zygotes with a rate of 2%–9% have been the most common PN abnormality [11]. Routinely, the chromosomally aberrant fertilized zygotes were discarded or donated for research due to their abnormal development to full term and correlation with early pregnancy loss [12]. In contrast, some studies mentioned that 3PN zygotes can give rise to hESC lines [13,14]. In the present study, 3PN embryos were used as an easy source of human embryos for donation for research study to evaluate the possibility of generation of hESCs following human in vitro embryo splitting.
There are limited reports of successful in vitro embryo splitting in primates [15] and humans [7 –9], which have left open the question of developmental competence of human in vitro embryo splitting. Our study has demonstrated on the developmental potential of the human twin embryos created by in vitro embryo splitting from human embryos at the cleavage stage. However, the blastocyst formation rate of donor embryos was higher than the recipient embryos (Table 1), and the difference was significant. According to the literature, the monitoring of the human 3PN zygotes showed normal in vitro development until the first two or three cell cycles, then arrested or abnormally developed by exhibition of unequal sized blastomeres, fragmentation, and multinucleation. This may be the reason that ∼10% of the triploid zygotes reached the blastocyst stage [16].
Hence, in the current study, good quality triploid zygotes were selected for the twinning procedures, which successfully developed to 8–10 blastomere stage. So, the blastocyst formation rate was 57.1% from 42 twined embryos. In fact, embryos with higher number of blastomeres seem to be more potent to progress to blastocyst after in vitro embryo splitting procedure (Table 1). Similar to our findings, Illmensee et al. indicated that the splitting of six- to eight-cell embryos yielded superior rates of twin embryos developing to blastocysts compared to two- to five-cell embryos [9]. Moreover, it was shown that the size of the twin blastocysts is correlating with number of starting blastomeres [7].
The second part of this work focused on the attempts to generate hESCs following in vitro embryo splitting from 3PN embryos. As was mentioned earlier, diploid hESCs can be derived from triploid zygotes, so they can serve as an alternative source of hESC line derivation [17]. Studies demonstrated that triploid embryos have been cultured on a MEF feeder layer and give rise to hESC lines successfully [13,14,18]. In the present study, microdrops [1] of YhFF#18 at passages 15 to 19 were used as feeder layer after mitotical inactivation. However, YhFF#18 was used to culture Yazd hESC lines (Yazd1–3) at the similar passage number, but the type of feeder and its passage number may have the impact on the success of hESC derivation.
The novelty of the present study, other than using human foreskin fibroblast (HFF) feeder layer to derive hESCs from abnormal embryos, is the attempt to derive hESCs from twin embryos following in vitro splitting. hESC-like outgrowths were generated from these twin abnormal embryos on HFF feeder layer in microdrop. However, there were difficulties in keeping them growing in culture for more than three to five passages.
In this regard, three main points should be considered: first, the embryos that were used in this study originated from genetically abnormal (triploid) embryos. Chen et al. reported that unlike diploid zygotes, successful derivation of hESCs from triploid zygotes had poor efficiency. They also showed that the ratio of driving triploid and diploid hESC lines is 1:3 [18]. Second, embryo quality served as one of the key factors that affected on successful derivation of hESC lines from blastocyst [17]. Moreover, recent reports suggested that hESCs are derived more efficiently from high quality embryos [19,20]. In contrast, it is difficult to obtain the good quality blastocysts following splitting for hESC derivation. Likewise, the blastocysts formed from the isolated blastomeres have reduced cell numbers [5,7]. It has shown that most of the cells in the ICM of blastocysts following in vitro twinning express markers of both ICM (NANOG) and trophectoderm (CDX2) [7].
Based on these findings, we hypothesized that one probable reason for nonestablishment of the hESC line from twin embryos is that cells in the outgrowth may still express both markers [7] and they could be closer to “naive” hESCs, which require different derivation and maintenance conditions. However, it was difficult to check these markers due to risk of losing them for further expansion and establishment of cell lines, following the first passage of outgrowths from two embryos (one donor and one recipient); the expression of TRA-2-49 (hESC marker) and SSEA1 (human trophoblast cell marker) revealed expression of these markers in some of the cells separately (Fig. 4D–I) and in one outgrowth colocalization of markers (Fig. 4J–L) similar to putative human embryonic germ cells (hEGCs). Considering the limitations at these early stages before cell line establishment, it is very difficult to interpret.
In contrast, following in vitro embryo twinning, the cell numbers of the blastocysts reduced, specially the ICM. It was shown that treatment of human embryos with the TGFβ inhibitor SB431542 increases NANOG-positive epiblast cells [21]. Moreover, adding other factors such as CHIR99021 (CH), a GSK3β inhibitor, and ROCK inhibitor Y27632 (Y) may increase the derivation efficiency of hESCs [22] from twin embryos following in vitro splitting. As a final point, in this study, late passaged HFFs were used in microdrops as the feeder layer for the derivation of hESCs, which might have an impact on the derivation success rate.
In summary, human blastocysts were obtained successfully following in vitro splitting embryos from 3PN zygotes, and first step of hESC generation was succeeded with formation of outgrowth. The morphology of colonies clearly looked like hESCs; however, no hESC line resulted. In the successful derivation of hESC line from twinned 3PN embryos, it would be expected that hESCs revert to a normal diploid characterization. Generation of hESCs following in vitro embryo splitting will lend itself for the future clinical grade stem cell production and may be replaced with cord blood stem cell banking to preserve stem cells for the next generations.
Footnotes
Acknowledgments
The authors are very grateful for Prof. Harry D. Moore and Prof. Peter W. Andrews for their critical comments. The authors are very grateful for Dr. Ali-Mohammad Abdoli for his support as a manager of the Yazd Reproductive Sciences Institute. This work was supported by grants for Miss Marjan Omidi's PhD project from the Yazd Reproductive Sciences Institute and the Research Deputy of the Shahid Sadoughi University of Medical Sciences.
Author Disclosure Statement
No competing financial interests exist.