
Abstract
Spinal cord injury (SCI), a prominent health issue, represents a substantial portion of the global health care burden. Stem cell-based therapies provide novel solutions for SCI treatment, yet obstacles remain in the form of low survival rate, uncontrolled differentiation, and functional recovery. The application of engineered biomaterials in stem cell therapy provides a physicochemical microenvironment that mimics the stem cell niche, facilitating self-renewal, stem cell differentiation, and tissue reorganization. Nonetheless, external microenvironment support is inadequate, and some obstacles persist, for example, limited sources, gradual aging, and immunogenicity of stem cells. Targeted stem cell gene manipulation could eliminate many of these drawbacks, allowing safer, more effective use under regulation of intrinsic mechanisms. Additionally, through genetic labeling of stem cells, their role in tissue engineering may be elucidated. Therefore, combining stem cell therapy, materials science, and genetic modification technologies may shed light on SCI treatment. Herein, recent advances and advantages of biomaterials and gene manipulation, especially with respect to stem cell-based therapies, are highlighted, and their joint performance in SCI is evaluated. Current technological limitations and perspectives on future directions are then discussed. Although this combination is still in the early stages of development, it is highly likely to substantially contribute to stem cell-based therapies in the foreseeable future.
Introduction
Spinal cord injury (SCI) is a costly and destructive neurological disease with persisting disabilities and devastating psychosocial outcomes [1]. So far, attempts to regenerate neurons and reconstruct neural circuits have been unsuccessful. Stem cell transplantation shows great promise for SCI treatment through spinal cord cell replacement and axonal regeneration to establish a connection in the ruptured neural circuit, yielding desirable results [2 –4].
Although stem cell therapeutics have provided new insight into SCI treatment, some challenges remain to be addressed, including low survival rate, unexpected differentiation, and recovery of functional capacity after cell implantation [5]. The adverse impacts of stem cells are mainly attributed to an unfavorable physicochemical transplant microenvironment [6 –9]. Recently, tissue engineering has sparked remarkable interest among experts. It can create a stem cell niche-like microenvironment to increase stem cell viability while altering stem cell differentiation through biophysical characteristics of materials [10].
Nonetheless, most of the stem cells derived from fetal tissues or embryonic stem cells (ESCs) have demonstrated therapeutic effects but cause ethnical concerns and pose risk of immunological rejection, which hamper clinical translation. As a recent hot technology of genetic engineering, induced pluripotent stem cells (iPSCs), will undoubtedly be able to solve the problem regarding stem cell source [11]. Stem cells can also be labeled by gene manipulation. Compared with traditional histology, the noninvasive tracking of gene-tagged stem cells is far more appealing [12]. The joint use of biomaterials and genetic manipulation can presumably improve stem cell transplant [13], the details of which are discussed in the current article.
Recently, the combination of biomaterials and gene manipulation has been used to facilitate stem cell transplant in SCI mice [14]. In particular, an increasing number of tissue engineering experiments have used iPSCs as seed cells [15,16]. To date, there have been several detailed reviews regarding the incorporation of biomaterials into stem cell-based therapies for SCI [17,18]. However, few reviews have addressed the combination of biomaterials and gene manipulation to promote the transplant of stem cells in SCI. In this study, we first outline the mechanisms of stem cell-based SCI therapy. We then focus on the applications of tissue engineering and genetic engineering-assisted stem cell transplantation in SCI. Furthermore, we propose that combining tissue engineering and genetic engineering to assist stem cell transplantation represents a critical research direction, as well as shed light on the future of SCI treatment.
Stem Cells Hold Great Potential for the Treatment of SCI
Staging of SCI pathophysiology
The spinal cord is a long, thin, tubular structure extending from the medulla oblongata in the brainstem to the lumbar region of the vertebral column and is responsible for communication between the brain and peripheral nervous system. Gray and white matter are the major constituents of the spinal cord. The butterfly-shaped gray matter is located in the center and includes numerous neuronal cell bodies, glial cells, and capillaries. The white matter, containing sensory and motor axons, surrounds and protects the gray matter.
External trauma, which often results in fractured or dislocated vertebrae, is the main cause of SCI. The pathological process of SCI is mainly divided into primary injury and secondary injury and can also be divided into four temporal phases: acute (<48 h), subacute (48 h to 14 days), intermediate (14 days to 6 months), and chronic (>6 months) (Fig. 1). The initial traumatic event (primary injury) leads to damaged neural cells (mainly neurons and oligodendrocytes), disruption of blood vessels, and destruction of the blood/spinal cord barrier. Altogether, these events trigger activation of a continuous secondary injury cascade, which further leads to injury of the spinal cord and neurological dysfunction.
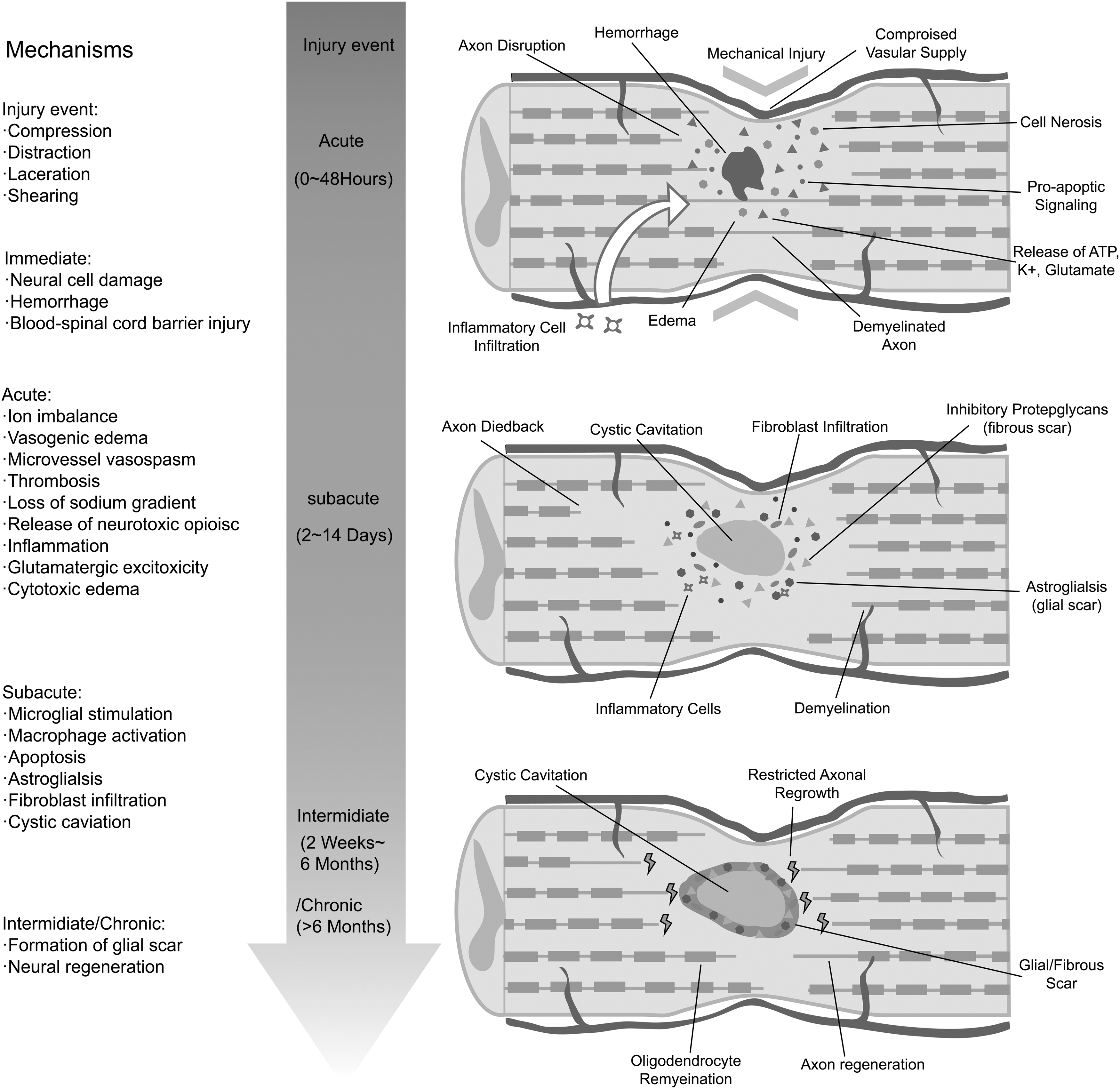
Pathophysiology of SCI in the acute, subacute, and intermediate/chronic stage. Primary and secondary injury determine the final extent of spinal cord damage. The primary injury event starts a secondary injury cascade that unfold in different phases within seconds of the primary injury and lasting for several weeks thereafter. SCI, spinal cord injury.
Secondary cellular injury, such as cell death and dysfunction caused by intracellular mitochondrial damage due to tissue ischemia and excitotoxicity, usually occurs during the acute phase and subacute phase [19]. Over the ensuing hours to days, necrotic cells release byproducts, for instance, ATP, DNA, and K+,which can activate microglial cells. In addition, the blood/spinal cord barrier is damaged, permitting an influx of cytokines, peptides, and inflammatory cells, including macrophages, neutrophils, and lymphocytes [20]. Phagocytic inflammatory cells will not only clear cell debris but also release cytotoxic byproducts, such as free radicals, inducing further damage to the spinal cord. At that point, various cells, including astrocytes and fibroblasts, are activated. The inflammatory reaction and the destruction of the vasculature further exacerbate the edema of the spinal cord and contribute to the progression of SCI.
As the acute inflammation subsides, the spinal cord lesion enters the intermediate or chronic phase. A cystic cavity is left behind owing to the overwhelming cell death and degeneration. Moreover, activated astrocytes, along with microglia, meningeal cells, and oligodendrocytes, form a scar around the cavity. In addition to the cellular composition, some extracellular matrix (ECM) proteins are secreted, such as chondroitin sulfate proteoglycans (CSPGs), keratan sulfate proteoglycans (KSPGs), and NG2 proteoglycan, from active microglia, astrocytes, and macrophages in the scar, inhibiting axonal growth [21,22]. The main consequences of SCI include the death of many cells, such as neurons and oligodendrocytes, formation of glial scar, and immune response, all of which obstruct neural regeneration [23,24]. Because of the activation of the secondary injury processes, the final pathological state is usually more severe than that in the first few hours following SCI. The common goal of SCI treatment is the recovery of some sensory and motor functions postcomplete SCI, which involves the following events: regeneration of long ascending and descending axons throughout the lesion site to reconnect the original targets and formation of new neural circuits in the lesion site by newly generated neurons postinjury, which effectively reconnect the transected stumps.
Therapeutic mechanism of stem cells in SCI
Stem cells have three main properties: self-renewal, high proliferative potential, and the ability to differentiate into multiple cell types [4]. Many different kinds of stem cells, including ESCs and various types of adult stem cells (ASCs), such as mesenchymal stem cells (MSCs) and neural stem cells (NSCs), have been transplanted for the sake of promoting tissue regeneration in SCI [25,26]. The therapeutic mechanism of transplanting stem cells after SCI is based on substitution for damaged or lost cells, furnishing of trophic support for neural cells, and modulation of the host environment within the lesion to facilitate axon regeneration, such as suppression of the activation of inflammatory cells and formation of glial scar [27].
Stem cells are transplanted into the lesion and differentiate into neurons and glial cells [28]. Replacement of lost or damaged neurons or oligodendrocytes, which can facilitate myelination of spared axons, is an important goal of stem cell transplantation. It was demonstrated that human ESC-derived oligodendrocyte progenitor transplants were able to restore locomotor function after SCI, which was potentially related to remyelination of spared axons [29].
Transplanted stem cells can also secrete bioactive molecules, such as cytokines and trophic factors, to provide neuroprotection for nerve cells. Many cells, such as MSCs and neural precursor cells (NPCs), can secrete trophic factors to promote the improvement of the injured environment after transplantation, thereby enhancing the survival of host cells, preventing cells from entering apoptosis, supporting blood vessel regeneration, and facilitating the growth of retracted axons across the lesions [30].
The inflammatory response plays an important role in the progression of SCI. Stem cell transplantation benefits regeneration through immunomodulation, stimulating beneficial inflammation, or attenuating detrimental inflammation. An experiment showed that transplanted NSCs could enhance neurological function after SCI and modulate SCI-induced inflammatory responses through reducing infiltrating neutrophils and M1 macrophage activation [31]. In addition, transplanted stem cells improve the host environment by modulation of reactive astrocyte function [32].
Based on the preclinical evidence of efficacy, stem cell transplantation has also been performed in SCI patients to further validate the efficacy. In one clinical trial, significant improvement in motor and sensory function was observed <6 months after autologous bone marrow-derived stem cell transplantation in 6 of 10 individuals. Moreover, in some people, sexual function, bladder control, and walking, at least with support, showed different levels of improvement [33]. A report detailing the clinical response of 277 patients with chronic SCI receiving stem cell transplantation revealed that 1 year after autologous peripheral blood stem cell transplantation, 43% of the patients improved; the ASIA score shifted from A to C in 32 and from A to B in 88 patients [34].
Biomaterials Render Stem Cell Therapy Feasible in SCI
Stem cell-based therapy offers a novel solution for the treatment of SCI, but the principal obstacles to such therapy lie in insufficient viability and uncontrolled differentiation of transplanted stem cells [35]. The physicochemical microenvironment controls the survival and differentiation of stem cells; thus, the key step to overcome the difficulties in stem cell transplantation is construction of a favorable transplant microenvironment. Biomaterial-based scaffolds can provide an environment to facilitate stem cell transplantation by mimicking the stem cell niche in vivo. Both natural and synthetic biomaterials can be used to create scaffolds. In this study, the advantages of biomaterials and the roles of biomaterials combined with stem cells in SCI treatment will be discussed.
Biomaterials used for scaffold fabrication
Scaffolds are, by definition, temporary supporting structures for growing tissues and cells. The biomaterials used for scaffold fabrication can be separated into natural materials and synthetic materials, which can be further divided into degradable and nondegradable materials.
Natural materials, such as fibrin, hyaluronic acid (HA), alginate (AG), and chitosan, are derived from animals or plants. They have good biocompatibility by virtue of their abundant cell-binding domains and natural bioactive motifs, which support the communication between the ECM and cells. Biocompatibility, as well as good biodegradability, is important for maintaining the morphology and function of natural tissues. The scaffolds made of natural materials degrade gradually during cell growth, which can create space for the cells to produce their own ECM and to construct their own scaffolds [36]. However, natural materials also have some shortcomings, such as a fast biodegradation rate and low mechanical strength, and they are usually difficult to modify biochemically because they cannot be designed like synthetic materials from the bottom up. In addition, it is difficult to sterilize natural materials, which often contain contaminants, leading to high immunogenicity and disease transfer [37].
Most of the synthetic materials consist of polymers, such as poly (lactic-co-glycolic acid) (PLGA), poly (glycolic acid) (PGA), and polyethylene glycol (PEG). Synthetic materials can be sterilized, and their key parameters, including porosity, degradability, architecture, and mechanical properties, are easily modified and controlled on the basis of demand [38]. The disadvantages of synthetic materials are poor biocompatibility, which results from the lack of corresponding recognition signals, and potentially toxic metabolites.
To overcome some of the disadvantages of single-phase materials and synergize the advantages of different materials, composite biomaterials, which consist of a combination of different kinds of natural and synthetic polymers, are designed. For example, gelatin, with unfavorable mechanical properties, is often used in combination with other substances, such as HA and chitosan [39]. It was demonstrated that the biocompatibility and mechanical properties of chitosan could be modified by blending with PEG [40]. However, because of the increase in the types of available materials, the manufacture of composite biomaterials will require more sophisticated production technology to combine different materials. A more comprehensive description of biomaterials for SCI and their characteristics can be found in Table 1.
Biomaterials Used in Spinal Cord Injury Repair
PCL, poly (ɛ-caprolactone); PEG, polyethylene glycol; PGA, poly (glycolic acid); PLGA, poly (lactic-co-glycolic acid); SF, silk fibroin.
Advantages of the combination of biomaterials and stem cells
Devoid of effective support, transplanted cells alone cannot completely fulfill their intended function, and the cell survival rate is not sufficiently high. The application of biomaterial scaffolds overcomes the shortcomings of weak cell survival and low matrix synthesis capabilities in unitary cell transplantations. Furthermore, the scaffolds can provide temporary mechanical support and biological guidance for the formation of tissues before their degradation and also serve as three-dimensional (3D) templates for subsequent tissue and organ construction. Therefore, the construction of the scaffolds is undoubtedly critical and merits future investigation. Herein, the effect of biomaterials on stem cells is discussed and illustrated in Fig. 2.

Schematic diagram of the effect of the combination of biomaterials on stem cells.
Survival rate of stem cells is higher in the context of biomaterial scaffolds
The key aspect of cell survival could be addressed by providing an environment able to host cells and creating a suitable biomimetic platform where they can proliferate. The scaffold design can mimic the physicochemical features and architecture of the extracellular environment, which is a suitable approach to preserve the active cell phase and improve cell survival. Scaffolds allow stem cells to maintain a shape that closely resembles their natural morphology in vivo. When grown in scaffold, cells are likely to form spheroids and aggregates, which support cell–ECM interactions and cell–cell interactions that mimic the natural environment in vivo. It has been demonstrated that, compared with dissociated cells expanded in monolayer culture, MSCs exhibit better survival when in spheroid form [41]. The higher rate of survival is likely because spheroids do not require stem cell detachment from the ECM, which can provide essential signals for cell survival [42]. Moreover, the 3D environment offered by scaffold allows enhanced cell-to-cell contact, which allows stem cells to form multilayer tissue-like structures, resulting in increased intercellular signaling.
Adhesion molecules are cell receptors responsible for communication between cells and between cells and ECM [43]. As an important step in multiple intracellular signaling pathways, adhesion molecule binding anchors the cell to its environment, and the adhesion molecules themselves connect to the cellular cytoskeleton internally. Thus, adhesion molecules are critical to many cellular activities, including adhesion, mechanotransduction, cytoskeletal organization, and gene expression. Additionally, adhesion molecules participate in the regulatory mechanism of stem cell survival. It has been demonstrated that an irreparable disruption of adhesion molecule signaling can lead to human ESC death [44]. Scaffolds used for SCI treatment are manufactured from a series of natural and synthetic biomaterials. Natural biomaterials are often derived from diverse components of the ECM, including fibrin, HA, and collagen, but can also be based on other naturally derived materials, such as gelatin and alginate. These materials are biocompatible and often have protein sites that bind with adhesion molecules of stem cells with high specificity. In an SCI study, the heterotransplantation of human bone marrow stromal cells carried by an HA-based scaffold in rats resulted in a better survival rate than transplantation with BMSCs in media alone. The promotion of stem cell survival may be caused by interaction between HA, a key component of the ECM, and stem cells through CD44 and RHAMM receptors, which influence stem cell migration and survival [45].
Scaffolds fabricated from synthetic materials may lack sites for cellular adhesion, which can be compensated for by coating with ECM proteins to achieve optimal results. However, immobilization of full-length protein onto the biomaterial is not always necessary to elicit an adhesion molecule binding event. Instead, short polypeptide ligands and recombinant protein fragments, such as the 7th–10th repeat of the fibronectin type III domain, the IKVAV peptide derived from laminin, and the RGD peptide found in vitronectin and fibronectin, are often adequate for adhesion molecule binding [46 –48]. A study showed that, compared with those cultured in a general hydrogel system, NSCs cultured in an RGD-functionalized hybrid hydrogel system exhibit better viability. However, both techniques can result in more stem cells in the active phases at 4 and 8 days than NSC monolayer (two-dimensional) culture [49].
Stem cell fate could be dictated by biomaterial properties
On account of the undirected differentiation of stem cells, they may differentiate into other cells upon transplant into the body, resulting in weaker repair function and even tumor development. Therefore, the regulation of the differentiation of stem cells in vivo is essential. Biomaterials transplanted with stem cells can govern the differentiation of stem cells through their mechanical, topographical, and biochemical properties, as shown in Fig. 3.
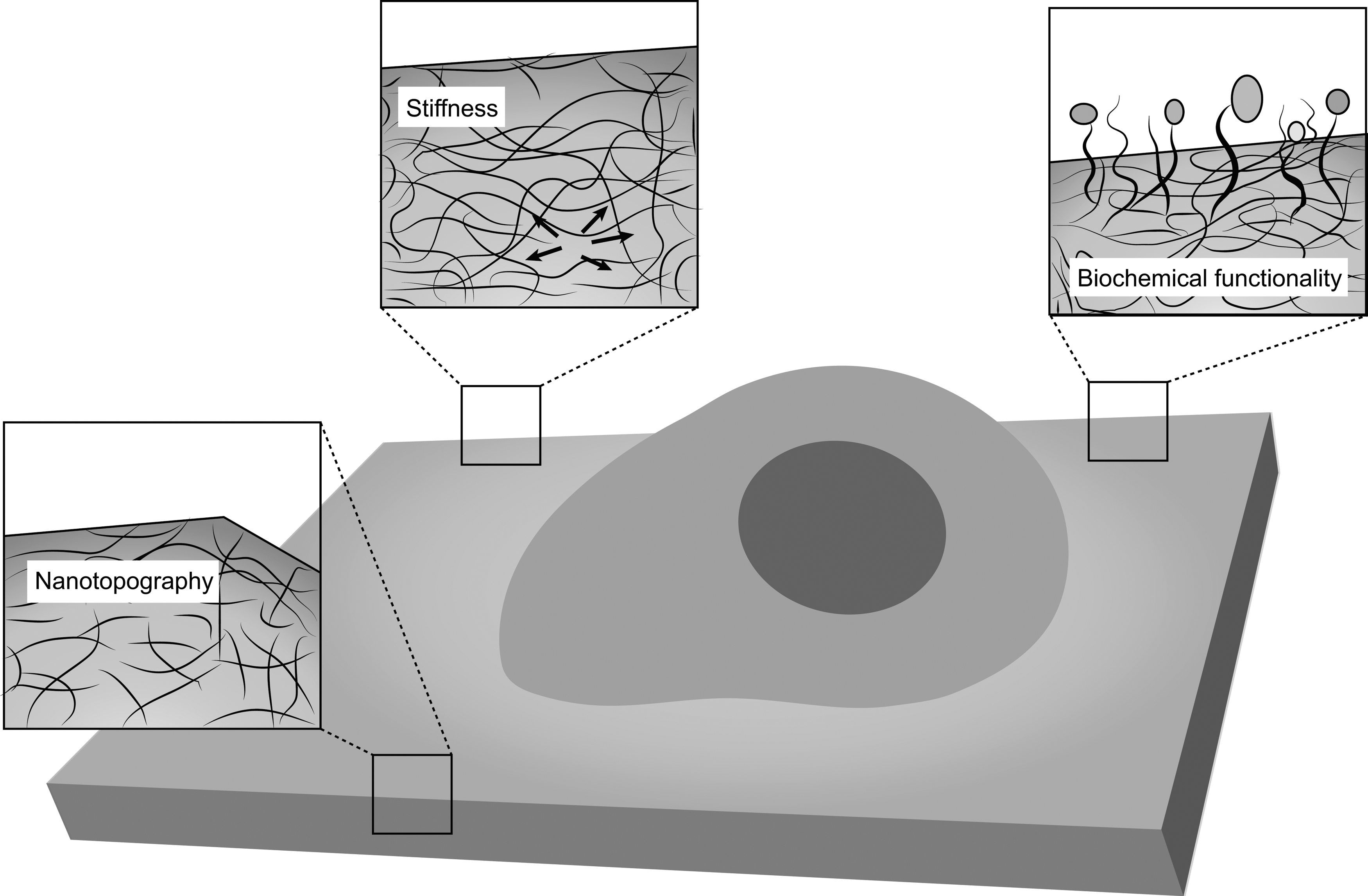
Effects of inherent material properties on stem cells. Stem cell fate can be influenced by inherent properties of materials (exemplified by a two-dimensional polymeric substrate in this schematic) near the material–cell interface, such as stiffness (pictured as force vectors), nanotopography, and biochemical functionality (represented by beads).
Because cells in different types of tissue survive, perceive, and respond in different mechanical environments, they prefer an environment similar to their native environment for growth. Stem cells present in various tissues experience different stiffness, which affects their fate, including adhesion, migration, proliferation, and differentiation. The softer the biological material, the higher the differentiation of stem cells into neural cells [50]. For example, a harder material promotes stem cells to differentiate into osteoblasts, whereas a softer tissue promotes differentiation into neural cells. Leipzig and Shoichet cultured rat neural stem/precursor cells (NSPCs) on gel with different stiffness and proved that gel stiffness affects the differentiation profile of stem cells. Softer (0.1 kPa) methacrylamide chitosan hydrogels predominantly promote NSPC differentiation into neurons and astrocytes, whereas harder hydrogels predominantly promote their differentiation into oligodendrocytes. Furthermore, NSPCs cultured on gels with moderate stiffness (3.5 kPa) result in the greatest amount of proliferation [51]. In response to extracellular mechanical signals, stem cells regulate the contraction of the cytoskeleton and maintain intracellular mechanical homeostasis by modulating the affinity to focal adhesions and the aggregation and depolymerization of adhesive proteins [52]. These adhesion complexes and interacting cytoskeletal components can activate mechanical response signals, such as the RhoA and MAPK transactivators, leading to the activation of downstream mechanical pathways that ultimately upregulate nuclear gene expression [53].
Because cells are able to sense micro- and even nanoscale topographical cues from their environment, changes in the nanotopography of biomaterials can direct stem cell fate. For example, nanofibers of various diameters could affect the differentiation potential of NSCs. Rat hippocampus-derived adult NSCs were cultured on laminin-coated electrospun polyethersulfone (PES) fibers with average diameters of 1,452 ± 312, 749 ± 153, and 283 ± 45 nm. Fibers of the latter diameter increased the differentiation of oligodendrocytes, whereas greater neuronal differentiation was observed in culture of NSCs on fibers of 749-nm diameter [54]. The mechanism that influences stem cell proliferation and differentiation may involve the different nanotopographic cues of the biomaterial, affecting cytoskeletal organization and structure of cells. After further investigation, it was determined that nanotopology is likely to regulate stem cell growth or differentiation through ERK and Jnk signaling pathways [55].
To encourage stem cell–scaffold interactions, many immobilized biological components are often used to decorate materials. In addition to the nature of the scaffold material, protein modifications, including epidermal growth factor (EGF), neurotrophin-3 (NT-3), and glial cell-derived neurotrophic factor (GDNF), can be applied to the scaffold to regulate stem cell fate [56,57]. Many experiments have also employed antibodies to regulate stem cell differentiation. Anti-EFGR antibody was tethered to a collagen scaffold to promote NSC differentiation into neurons for SCI repair. The decorated scaffold induced higher production of nascent neurons from transplanted and endogenous NSCs, which facilitated reconstruction of the neuronal connections with host neurons or between NSCs to transmit electrophysiological signals and promote functional recovery [58]. Our laboratory loaded catalpol, an active extraction of traditional Chinese medicine, into carbon nanofibrous scaffolds to guide human adipose tissue-derived stem cells (hASCs) to differentiate into neurons. We found that hASCs within the catalpol-loaded scaffold had markedly upregulated expression of Nissl and βIII-tubulin, markers of neurons, as determined by reverse transcription/polymerase chain reaction (RT-PCR), western blot, and immunocytochemistry assays [59].
Combinatorial strategies of biomaterial-supported stem cell transplantation to reconstruct functional neural connections
Stem cell-based therapy provides a potential source of cells to repopulate the injured spinal cord and assist in functional recovery by promoting regeneration of host axons, replacing damaged circuits, and secreting related factors. Scaffold decorated with ECM adhesion proteins, bioactive factors, or peptides can not only improve stem cell therapy by providing a defined microenvironment during transplantation but also promote host cell regeneration by supplying a favorable framework to bridge the neural gap in SCI lesions. To synergize stem cells and scaffold, a combinatorial strategy involving stem cells and scaffold is applied in SCI treatment. After SCI, the reconstruction of functional neural connections provides the structural foundation for functional recovery. In this section, we introduce the role of stem cells combined with scaffolds in the reconstruction of functional neural connections in SCI treatment. More comprehensive information regarding biomaterial-supported stem cell transplantation for SCI can be found in Table 2.
The Combination of Biomaterials and Stem Cells in Spinal Cord Injury Treatment
AGs, alginates; CSPG, chondroitin sulfate proteoglycan; DPSCs, dental pulp stem cells; ECM, extracellular matrix; EGFR, epidermal growth factor receptor; ESC, embryonic stem cell; ESNPCs, embryonic stem cell-derived neural progenitor cell; GDNF, glial cell-derived neurotrophic factor; GFs, growth factors; HBDS, heparin-binding delivery system; iNSCs, induced neural stem cells; MEP, motor evoked potential; MSCs, mesenchymal stem cells; NPCs, neural precursor cells; NSCs, neural stem cells; NSE, neuron-specific enolase; NSPCs, neural stem/precursor cells; NT-3, neurotrophin-3; PCLA, poly (e-caprolactone)-block-poly (
Gap at the lesion epicenter could be bridged by scaffolds combined with stem cells
After SCI, with the death of nerve cells and formation of glial scar, a cavity gradually develops, which hinders axonal regeneration. To promote regeneration, it is necessary to fill the cavity. Scaffolds of specified architecture transplanted into the hole provide a platform, which can act as a “bridge” for injured tissue, to support the function of stem cells and host cells, including axon regrowth, neuronal relay formation, and myelin regeneration, resulting in significant structural and behavioral recovery.
The disruption of neural connections is the primary cause of sustained dysfunction following SCI. The major goal of repairing damaged connections is to promote retracted axon sprouting or regeneration. It was demonstrated that stem cells combined with scaffolds transplanted into SCI lesions could promote axon regrowth, which may be due to the growth platform provided by the scaffolds. A study showed that in SCI rats, the sensory and motor spinal axons extended through the human NPC/PGA complex scaffold [60]. In addition to promoting the extension of retracted axons, transplanted stem cells combined with scaffold can promote the repair of neuronal connectivity through formation of neuronal relays. An experiment has shown that stem cells embedded within scaffold grafting can promote the formation of synapses between transplanted stem cells and host neurons. Compared with those in BMSC-treated SCI rats, higher NF-positive signals, which indicated the regeneration of axons, were found in SCI rats treated with BMSCs and scaffold [61].
Loss of myelin and apoptosis of oligodendrocytes are observed early in SCI. Demyelination results in poor metabolic shuttle between axons and oligodendrocytes and impairs the conduction of electrical signals; both are related to functional recovery after injury. Grafts of stem cells within scaffolds can differentiate into oligodendrocytes and strengthen myelin regeneration. It was demonstrated that, compared with stem cells transplanted alone, stem cells combined with scaffolds could differentiate into more oligodendrocytes. In a comparative study between scaffold- and intralesional injection-based transplantation of MSCs for acute SCI, rats were subjected to standardized SCI, followed by transplantation of allogeneic MSCs, either through chitosan scaffold (IC group) or PLGA scaffold (IP group) or through intralesional injection (IL group). The results showed a higher success rate of MSC engraftment in the scaffold groups compared with the IL group. The transplanted MSCs mainly differentiated into oligodendrocytes in the scaffold groups, whereas the predominant differentiated cells detected in the IL group were astrocytes. Moreover, the functional improvement was most notable in the IC group, which corresponded to the features of the biomaterials used for scaffold fabrication [62].
Growth environment could be improved by the combination of scaffolds and stem cells
The lack of neurotrophic factors and glial scar formation, together with inflammatory reaction at the lesion, is regarded as the main mechanism resulting in the failure of the reconstruction of functional connections [63]. In addition to supporting the growth of stem cells and endogenous neuronal cells by bridging the gap, the scaffold can cooperate with stem cells to provide a favorable growth environment for establishment of functional neural connections.
It was proven that cotransplantation of scaffolds and stem cells can regulate the inflammatory response. In one experiment, it was found that the scaffold could efficaciously deliver CCL2 secreted by human MSCs, which plays a role in not only recruiting macrophages but also driving their transformation to an M2 neuroprotective phenotype, to preserve motor neurons and myelin in the lesion. Moreover, mice treated with hMSCs loaded into hydrogel showed a significant improvement in locomotor performance compared with the untreated group, which may be due to the protective effect of CCL2 on neurons and myelin [64].
Glial scarring impedes axonal regeneration; thus, reducing the formation of glial scar is thought to have a positive effect on axonal regeneration and functional recovery. Transplantation of stem cell-seeded scaffolds can not only diminish fibrosis from the secondary injury process of SCI [65], but also attenuate secondary glial scar formation. In a chronic SCI dog model, transplantation with the combination of MSCs and scaffold reduced secondary glial scarring after resection of the initial scar. In chronic SCI dogs, the scar tissues were surgically resected 2 weeks after spinal surgery [66]. Subsequently, MSCs seeded in scaffolds were transplanted into the lesion, and it was found that locomotor function was significantly enhanced. Moreover, compared with that in the control group, CSPG deposition (composition of glial scar) in the treated group was lower. In addition, many newly myelinated axon fibers and synapse formation were noted in the lesion at 1 year postinjury. Thus, combination transplantation may be a potential therapy for chronic SCI that reduces glial scar formation, promotes neuronal regeneration, and ultimately improves locomotor recovery.
Modifying different proteins, such as neurotrophic factors, on the scaffold can directly provide a favorable environment for stem cells and host cells. A composite treatment system, consisting of an alginates/silk fibroin/glial cell line-derived neurotrophic factor (AGs/SF/GDNF) scaffold and human umbilical cord MSCs (hUCMSCs), was used in SCI rats. Compared with GDNF alone, AGs/SF scaffold, or AGs/SF/GDNF scaffold, the combination of hUCMSCs and GDNF loaded on AGs/SF scaffold showed better repair and therapeutic effects in the SCI rats, such as higher neuron survival and preferable functional improvement of locomotor activity [56]. Nevertheless, scaffolds loaded with active substrates do not always promote the repair of SCI. A study demonstrated that growth factor mimetics loaded on mesoporous silica particles (MesoMIM) as well as human spinal cord NSPCs (hscNSPCs) supported sensory axon regeneration in mice with dorsal root avulsed spinal cord. However, when MesoMIM and hscNSPCs were transplanted together, sensory axon regeneration failed. It is possible that coimplantation of MesoMIM prevented stem cell migration, which promoted sensory axons to grow through the newly established glial scar [67]. In another study, to limit the effect of the inhibitory environment surrounding the transplanted cells, anti-inhibitory molecules and ESC-derived progenitor motor neurons (PMNs) were seeded within scaffolds together for implant in the lesion. Nonetheless, the combination of sustained delivery of anti-inhibitory molecules and PMNs led to reduced cell survival, which may be related to an enhanced inflammatory reaction in the lesion [68]. While combination therapies have potential for effective treatment of SCI, further work is needed to improve the effects.
Merging Biomaterials and Gene Manipulation May Improve Stem Cell Implant in SCI
Although biomaterials can improve the viability of stem cells and promote their differentiation in SCI, the support from the external microenvironment is inadequate, and many issues surrounding stem cells have yet to be resolved, such as limited sources and immune rejection after transplantation. Genetic manipulation directly modifies stem cells by different gene technologies, which can directly improve their performance. Furthermore, reporter gene-modified stem cells allow tracking of cell fate in vivo over a long period and help clarify the mechanism of SCI treatment. Therefore, the combination of biomaterials and gene manipulation can better promote the therapeutic effect of stem cells on SCI. In this section, the application of genetically modified stem cells for treatment of SCI and the combination of gene manipulation and biomaterials to facilitate stem cell implant in SCI will be discussed, as shown in Fig. 4.
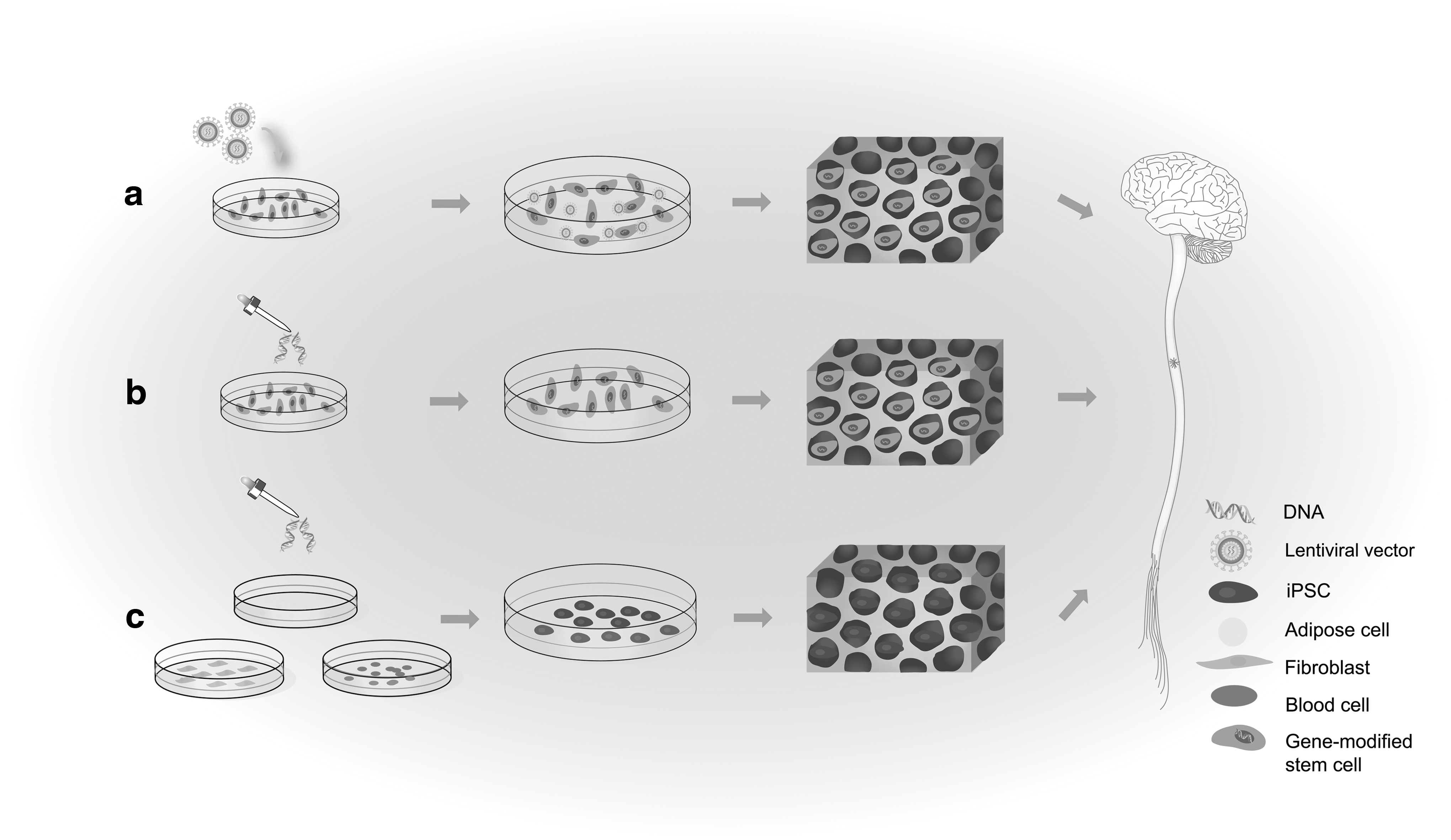
Schematic diagram of the effect of the combination of tissue engineering and gene manipulation on stem cells in SCI.
Genes encoding detectable proteins in stem cells combined with biomaterial scaffold
To ensure stem cell therapies are effective and successful in the treatment of SCI, it is crucial to track the survival, migration, and differentiation of transplanted cells, their capabilities of reconstructing brain function, and their biological role. Histology, as the traditional method of evaluating stem cell outcome, is the golden standard in preclinical cell monitoring [69]. However, this requires the sacrifice of experimental animals, and histology provides no longitudinal or whole-body monitoring, hampering clinical translation. Therefore, noninvasive methods have been gradually developed and used widely for investigation of SCI [70,71]. These methods enable direct visualization of stem cell delivery with an indication of the location of the transplanted cells and their survival over time.
Genes encoding detectable proteins in stem cells
Reporter gene labeling is a noninvasive cell tracking technique in which a detectable protein gene that can be expressed by progeny is integrated into the cell's chromosome through viral or nonviral vectors. Thus, the proliferation and death of cells can be detected by changes in signal following cell administration [72].
The most widely used imaging reporter gene in SCI is Fluc, which encodes luciferase protein that catalyzes oxidation of
Although BLI is a valuable and widely used tool for preclinical cell monitoring, it cannot be translated to a clinical setting; BLI can penetrate only a few centimeters of tissue. Therefore, different positron emission tomography (PET), single-photon emission computed tomography (SPECT), and magnetic resonance imaging (MRI) reporter genes have been developed, such as MRI reporter gene ferritin and radionuclide reporter gene D2R [75]. Multiple imaging allows for the full positioning of stem cells, as shown in Fig. 5. For example, the combination of PET and computed tomography (CT) is commonly used for longitudinal monitoring in SCI [76]. Multimodal noninvasive imaging reporter genes can also be developed for use with different imaging technologies to obtain sufficient information regarding the biological behavior of stem cells. Cao developed a novel triple-fusion (TF) reporter gene that can be detected by PET, BLI, and fluorescence and transduced it into ESCs. They observed that the reporter gene was stably expressed in ESCs, and the survival, migration, and proliferation can be dynamically monitored long term [77]. However, there are some problems with the technology of multiple reporters. First, fusion protein genes of a large size are often difficult to construct. Additionally, fusion proteins lose some bioactivity during gene fusion and protein expression. These problems must be addressed before cell tracking technologies can be applied to clinical treatment.
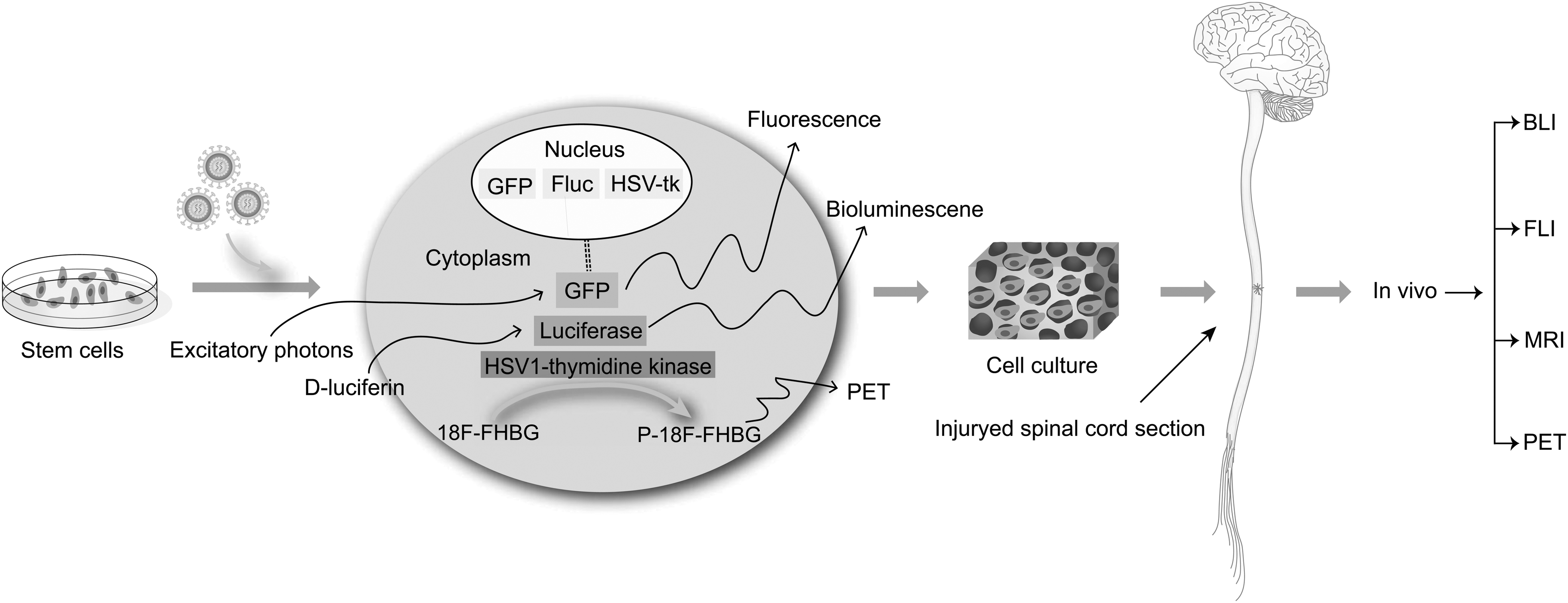
Multimodal imaging can be applied to track stem cell behavior. A workflow diagram for labeling of stem cells and subsequent transplant in vivo is presented: first, cells are labeled using a marker for BLI, MRI, FLI, or PET. Second, stem cells are cultured within biomaterial scaffold in vitro and transplanted into the injured spinal cord. Finally, stem cells are tracked in vivo with a scanner or camera. BLI, bioluminescence imaging; FLI, fluorescence imaging; MRI, magnetic resonance imaging; PET, positron emission tomography.
Genes encoding detectable proteins in stem cells combined with scaffold
After combining stem cells with tissue engineering to treat SCI, it is also necessary to track the fate of stem cells in the scaffold material to evaluate the therapeutic potential of stem cell-based tissue engineering strategies. One experiment demonstrated that luciferase-labeled stem cells can still be traced in vivo after binding to the scaffold in spite of reduced fluorescence intensity [78]. Different characteristics of the scaffold have various effects on stem cells. BLI can be used to assess the effect of potential confounding factors of the scaffold on stem cells. In one study, BLI was used to compare the differences between agarose and RGD-functionalized alginate hydrogel scaffold. The number of stem cells was strongly correlated with the intensity of fluorescence of the different scaffolds. Furthermore, alginate constructs exhibited twofold greater BLI counts than agarose constructs at comparable hMSC doses. Meanwhile, transplantation of low cell doses resulted in better survival than high cell doses within an alginate matrix, which may be because the cells did not compete for nutrition [79]. In another study, the proliferation of human NSCs incorporated within hydrogels with different elastic properties was evaluated by BLI in vivo. Compared with the cells within the stiff hydrogel, NSCs within the soft hydrogel proliferated more vigorously [80].
Limited research has been performed on the effects of gene-labeled stem cells combined with biomaterial scaffolds for spinal cord regeneration. However, the combination is used in a similar environment brain tissue. In a corticectomized rat model, BLI was used to determine the survival of transplanted human NSCs seeded in a 3D biocompatible scaffold. Compared with 8 days for the cells alone, the bioluminescence signals from the combination of stem cells and scaffold were maintained for 14 days. The image-guided approach proved that the viability of stem cells could be supported by scaffold [81]. Long-term survival of transplanted stem cells is a basic requirement for functional recovery, and scaffolds can enhance the survival rate of stem cells. In vivo imaging techniques are useful for monitoring stem cell transplantation because they allow longitudinal observation of stem cell survival and migration. It is safe to assume that when in vivo imaging techniques for stem cells combined with scaffolds are applied in models of SCI, the survival, distribution, and even differentiation of transplanted stem cells can be easily and dynamically monitored.
Despite these successes and remarkable potential, many issues exist in reporter gene labeling technologies, such as attenuation of signal, security of transfected stem cells, and limited capability of revealing cell functionality. In the current research, most experimental subjects are small rodents, which are considerably different from humans, thereby limiting the direct applicability of the preclinical research to clinical practice. Larger animal models used in preclinical experiments could bridge the gap between humans and rodent models, but are expensive and complicated. More importantly, current tracking techniques can only supply information related to spatiotemporal location, migration routine, and viability of transplanted stem cells. For clinical trials, it is necessary to visualize the differentiation and even functionality of transplanted stem cells. Moreover, signal attenuation due to the binding of the labeled stem cell and scaffold could influence the monitoring. Therefore, more experiments are needed to improve reporter gene technologies to trace stem cells more precisely in SCI.
Genes regulating expression of signal transduction-related proteins in stem cells combined with biomaterial scaffold
Signal transduction is the process by which an extracellular signal is transmitted through a cell as a series of molecular events, which eventually results in a cellular response. Proteins responsible for detecting signals are generally termed receptors. As members of the signal transduction-related proteins, changes in both the signal molecules and receptors can influence the signaling transduction, thereby regulating the cellular response. Signal molecules normally presenting during development, which promote cell differentiation and survival, are insufficient in the adult spinal cord lesion. Nakamura et al. showed that in a favorable environment, such as injured neonatal spinal cord, transplanted NPCs can survive, integrate with host tissue, and differentiate into neurons and oligodendrocytes, which result in a return of lost function [82]. Stem cells could increase favorable signals by increasing the expression of secreted proteins to improve the inhibitory posttransplantation microenvironment. Moreover, when expression of stem cell receptors changes, cell responses, which are related to changes in signal transduction, can be regulated. Thus, gene transfection technology to regulate the expression level of signal transduction-related proteins and biomaterial scaffolds to improve the effect of cells are acceptable strategies to enhance the cellular functions of stem cells in SCI tissue repair.
Genes regulating expression of secreted proteins in stem cells combined with scaffold
Growth factors modulate neuronal survival, neurite outgrowth, synaptic plasticity, and neurotransmission. Many in vivo and in vitro studies have shown that growth factors, such as GDNF, brain-derived neurotrophic factor (BDNF), basic fibroblast growth factor (bFGF), nerve growth factor (NGF), NT-3, NT-4, and NT-5, can promote the proliferation, differentiation, and migration of NSCs [83]. Since the half-life of the trophic factor is short, it is difficult to introduce it alone into the central nervous system. The introduction of trophic factor genes into stem cells by means of transgenics can promote the continuous release of trophic factors at a physiological dose and achieve a long-term effect.
The GDNF gene was utilized to improve the therapeutic outcome of stem cells in SCI. Differentiated BMSCs without genetic modification or GDNF-overexpressing BMSCs (BMSCs-GDNF) were injected into rats on the third day post-SCI [84]. Compared with the BMSCs, the BMSCs-GDNF showed better results in terms of BBB score and axon counting. This result suggests a novel strategy to promote the effect of stem cells through combination with gene therapy, which can improve the transplantation base therapies for SCI. In another study, GDNF was transfected into placenta-derived MSCs (PMSCs) and BMSCs to compare the effects of different sources of stem cells transfected with GDNF in the treatment of SCI [69]. However, there was no significant difference between transfected PMSCs and BMSCs based on immunohistochemistry, flow cytometry, kinesiology, and BBB locomotion score.
When the genetically modified stem cells are transplanted together with the scaffold, the scaffold can improve the cell survival rate and gene expression, thereby further expanding the therapeutic effect of the stem cells. Gao and his group introduced the BDNF gene into MSCs through liposomal vector and used the MSCs in conjunction with adhesive peptide-modified hydrogel scaffold for SCI treatment. Compared with the untransfected stem cell transplantation group, genetically modified stem cells could significantly inhibit the formation of glial scar, weaken the immune response, and promote axonal regeneration and tissue integrity. Moreover, these effects were significantly attenuated compared with those in the transplantation of genetically modified stem cells alone. Therefore, the combination of scaffolds and gene modification with growth factors can significantly improve the effect of stem cells on SCI [14]. In another study, NT-3-overexpressing NSCs or NSCs embedded within poly (ɛ-caprolactone) (PCL) scaffolds were transplanted into a rat hemisection model. NT-3-overexpressing NSCs showed better migration and survival and more frequently differentiated into oligodendrocytes and neurons compared with normal NSCs. In addition, rats with NT-3-overexpressing NSCs seeded within scaffolds showed the best locomotor recovery and were the only group with recorded motor-evoked potentials. The positive outcome may be related to the increased level of NT-3 [85].
Although transplantation of scaffolds seeded with genetically modified stem cells could create a favorable microenvironment to protect the cells and boost axon extension of host neurons, gene-modified stem cells may present many unpredictable problems. Moreover, when the secreted proteins and stem cells (without gene modification) are loaded on scaffolds that are subsequently transplanted into SCI lesion, they can also promote tissue repair. In fact, a series of sophisticated bioinspired systems have been developed to control the quantity, timing, and location of protein release [86]. Thus, more experiments are needed to compare these two ways from various aspects, such as maneuverability, durability, and security, which may facilitate selection of a more efficient method to treat SCI.
Genes regulating expression of receptors in stem cells combined with scaffold
Myelin is an important neural circuit structure whose regeneration remains a challenging issue in SCI. Axon myelination facilitates neuronal performance by increasing the speed of nervous impulse, and therefore, damage to myelin and oligodendrocytes can cause severe dysfunction. A study showed that a tissue-engineered neural scaffold-containing Schwann cells overexpressing NT-3 and NSCs overexpressing tyrosine kinase receptor type 3 (TrkC), a high-affinity receptor of NT-3, coseeded in gelatin sponge (GS) scaffold transplanted into SCI lesions resulted in better myelination [87]. In the study, Lai and his group first built a tissue-engineered neural scaffold in vitro and then transplanted it into a complete T10 spinal transection rat model. Subsequent results revealed that many myelins were formed, and the myelin sheath showed distinct multilayered lamellae formed by the seeded cells based on transmission electron microscopy. In addition, it was found that certain parts of the myelin sheath that wrap the axon were formed by NSC-derived myelinating cells.
Although many in vivo and in vitro studies demonstrated the neuronal properties of cells derived from MSCs under different induction environments, MSC transdifferentiation into neurons remains controversial. One major concern is that the MSC-derived cells fail to connect with the host neuronal network. Zeng et al, integrated MSC-derived neural-like cells into a neural network by genetically engineering MSCs overexpressing TrkC after rat spinal cord transection [88]. They cocultured MSCs overexpressing TrkC with NT-3-overproducing Schwann cells in 3D GS hydrogel for 14 days in vitro to predifferentiate MSCs into cells exhibiting neuronal features. Then, they transplanted MSC-GS or GS into an acute SCI rat model. After 8 weeks, MSC-derived neuron-like cells formed similar connections with host tissue, and c-fos was expressed only in the lumbar and grafted spinal cord cells when stimulated by motor cortex electrical signal; the BBB score of the MSC-GS-treated group was significantly higher than that of controls receiving GS.
These findings provide a basis for the future application of the combination strategy to repair SCI. However, Schwann cells overexpressing NT-3 also secrete many types of trophic factors, such as bFGF, BDNF, and NGF as well as adhesion molecules and ECM. Further study is needed to explore the concrete mechanism by which genetically engineered stem cells interact with NT-3-overproducing Schwann cells in SCI repair.
iPSCs combined with biomaterial scaffold for SCI treatment
iPSCs-derived from somatic cells by transduction with transcription factors have revealed characteristics comparable to those of ESCs, without supply constraints and ethical concerns, and have thus emerged as a potential treatment option for repairing SCI.
The application of iPSCs in SCI treatment has also produced favorable results. Nori and his colleagues investigated the potential of transplanted human iPSCs (hiPSCs) for treatment of SCI. They observed that grafted hiPSC-derived neurospheres (hiPSC-NSs) mainly differentiated into the three neural lineages (neurons, astrocytes, and oligodendrocytes), with no tumor formation within the lesion. Compared with the vehicle-treated control animal, more significant functional recovery was observed in the hiPSC-NS group [89]. The efficacy of three different types of stem cells, including spinal fetal cell-derived NPCs, hiPSCs, and BMSCs, in the treatment of SCI was compared. Among these cells, the iPSC-NPCs provided the most beneficial effect, preserving host tissue, reducing astrogliosis, increasing axonal sprouting, and promoting motor functional recovery [90].
The combination of scaffold and iPSCs has emerged as a viable therapy for SCI. iPSCs as seed cells circumvent the issues with ethics and insufficient supply. Meanwhile, the scaffold can improve the low survival rate of iPSCs caused by the adverse environment of SCI and promote iPSC differentiation into neural cells. In one study, iPSC-derived NSCs (iNSCs) encapsulated within a 3D gelatin methacrylate (GelMA) hydrogel were transplanted to promote tissue regeneration after SCI. iNSCs encapsulated in the 3D GelMA hydrogel survived and differentiated better in vitro and promoted more axon regeneration by decreasing inflammation and inhibiting glial scar formation in vivo than the control group [91]. In another recent experiment, myelomeningocele in a fetal lamb model was treated with both iPSC and scaffold therapies. NSCs derived from hiPSCs (iPSC-NSCs) were seeded within nanofibrous hydrogen and then transplanted into an animal with SCI. Thirty days after surgery, posttransplant immunohistochemistry demonstrated that hiPSC-NSCs survived, differentiated into the neuronal lineage, and integrated within the spinal cord axonal elements of the host [92].
One of the major challenges in iPSC transplantation is teratoma formation. A study observed that injectable hydrogel attenuates long-term teratoma formation of iPSC-derived oligodendrocytes in an SCI model [93]. The combination of iPSC-derived oligodendrocyte progenitor cells and peptide-modified hydrogel composed of methylcellulose and hyaluronan was transplanted into a rat model of SCI. Compared with transplantation in media, oligodendrocyte progenitor cells in scaffolds have a higher survival rate and exhibit better integration. In addition, the formation of teratomas was attenuated when the cells were seeded within hydrogel, which may be related to the increase in differentiation. Thus, the combination of scaffold and iPSCs is a promising therapeutic strategy for SCI repair.
Although iPSCs embedded in scaffolds have great potential for SCI treatment in autologous transplantation, some obstructions, such as tumorigenicity and feasibility, should be overcome before clinical application. About 3 months are required to generate hiPSCs from somatic cells of patients. Then, more than a year is required to evaluate the quality of the produced cells, such as their tumorigenic potential. It was demonstrated that the optimal time for treatment of SCI through cell transplantation is the subacute phase (48 h to 14 days after SCI) [94]. Therefore, it is difficult to transplant autologous hiPSCs into patients with SCI within this timeframe. It has been proven that scaffolds can promote stem cell differentiation and reduce tumor formation, but scaffolds cannot destroy tumors. More research is needed to solve these issues to apply iPSCs combined with scaffolds early in clinical therapy.
Perspective
The combination of biomaterials and gene manipulation in stem cell-based therapy promotes neurological tissue renovation and presents several promising and exciting avenues in the area of regenerative medicine. The combination can be used not only in SCI but also in many other conditions, such as wound healing and chondrogenesis [95,96].
Biomaterial scaffolds provide a transplanted microenvironment, mimicking the stem cell niche in vivo, to influence the fate of stem cells through biophysical signals from materials and further facilitate the renovation of damaged tissues. Genetic manipulation directly modifies stem cells by different gene technologies, which can directly improve their performance. Genetic modification of the immunogenicity of stem cells is a unique tactic to overcome graft rejection [97]. Transfection of the human telomerase reverse transcriptase (hTERT) gene into BMSCs could combat cell senescence [98]. There are no studies of weakening cell immunogenicity and combating cell senescence using scaffolds in SCI repair, representing possible directions for future experiments to verify their effect.
Given the promising results of the mentioned studies, it is speculated that the joint use of biomaterials and genetic manipulation will positively impact stem cell transplant in SCI. To date, few experiments on biomaterials combined with gene-assisted stem cell transplantation in SCI have been performed, which provides an opportunity for future studies, including but not limited to the aspects mentioned above.
Footnotes
Acknowledgment
This research was funded by the Chinese National Natural Science Foundation grant numbers 81271412 and 81471308.
Author Disclosure Statement
No competing financial interests exist.