
Abstract
Glial cell-derived neurotrophic factor family receptor alpha-1 (GFRα1) plays a crucial role in the self-renewal and maintenance of spermatogonial stem cells (SSCs) from mammals. However, to date, our knowledge about its role in fish SSCs is limited. In the present study, the medaka (Oryzias latipes) gfrα1 duplicate genes, Olgfrα1a and Olgfrα1b, were cloned and characterized. Furthermore, their expression profile and biological activity were investigated. OlGfrα1a and OlGfrα1b predict 524 and 466 amino acid residues, respectively. Both are orthologous to mammalian Gfrα1 by sequence analyses and appear high in spermatogonia by in situ hybridization assay. The knockdown of OlGfrα1a and/or OlGfrα1b via Vivo-Morpholino oligos significantly inhibited the self-renewal and maintenance of SSCs, as evidenced by the decreased proliferation activity of SG3 cells (a spermatogonial stem cell line derived from adult medaka testis) as well as spermatogonia in the testicular organ culture and by the decreased survival rate and expression levels of pluripotency-related genes (klf4, lin28b, bcl6b, and etv5) in SG3 cells. Additionally, our study indicates that OlGfrα1a might function by binding either Gdnfa or Gdnfb (the two medaka Gdnf homologs), whereas OlGfrα1b function by binding Gdnfa not Gdnfb. Taken together, our study indicates that both OlGfrα1a and OlGfrα1b are involved in the self-renewal and maintenance of SSCs by binding Gdnfa and/or Gdnfb, respectively. These findings suggest that the GDNF/GFRα1 signaling pathway might be conserved from mammals to fish species.
Introduction
G
Several studies report that Gdnf and/or Gfrα1 homologs exist in fishes, such as rainbow trout (Oncorhynchus mykiss), Nile tilapia (Oreochromis niloticus), and medaka (Oryzias latipes) [12 –15]. In rainbow trout, Gdnf and Gfrα1 show high expression in almost all type A spermatogonia and dramatically decrease in subsequent developmental stages, implying their potential roles in the spermatogenesis [12,13]. In Nile tilapia, immunohistochemical staining using anti-human Gfrα1 indicated that Gfrα1 might be exclusively expressed in SSCs [15]. Our previous study suggests that two Gdnf homologs (named as Gdnfa and Gdnfb) exist in medaka; moreover, both of them can mediate the self-renewal of SG3, a spermatogonial stem cell line derived from adult medaka testis [14,16]. However, functional study of Gfrα1 in fish SSCs is lacking till now. In the present study, the gfrα1 duplicate genes from medaka fish, namely Olgfrα1a and Olgfrα1b, have been characterized; their expression profile and biological activities in SSCs have been investigated; additionally, their reactivity to Gdnfa and/or Gdnfb has been also initially evaluated.
Materials and Methods
Fish
Medakas were kept in recirculating aerated water tanks at 26°C. Experiments with fish followed the Guidelines for Care and Use of Laboratory Animals prescribed by the Regulation of Animal Experimentation of Chongqing, China.
Complementary DNA cloning
Complementary DNA (cDNA) libraries from the brain and testis of adult medaka were synthesized using the PrimeScript II 1st Strand cDNA Synthesis Kit (Takara, Japan), as previously described [14]. Polymerase chain reaction (PCR) primers specific for the open reading frame (ORF) of Olgfrα1a and Olgfrα1b were designed through searching the GenBank (Supplementary Table S1; Supplementary Data are available online at
Sequence analyses
The protein structure was identified by the PredictProtein Program (
Reverse transcription–PCR
cDNA templates from embryos at different developmental stages, adult tissues including the brain, eye, heart, spleen, liver, intestine, ovary, and testis, and the SG3 cells were prepared as previously described [14]. The intron-spanning primer sets, which generated the correct sizes, were used for PCR amplification. β-Actin was amplified from the same set of cDNA samples as an internal control.
RNA in situ hybridization
RNA in situ hybridization (ISH) was performed as previously described [17]. Adult testes were cryosectioned at 3 μm thickness and stored at −80°C before use. For RNA probe preparation, 1,167-bp gfrα1a and 608-bp gfrα1b cDNA sequences containing T7 promoter were first amplified and sequenced; subsequently, sense and anti-sense RNA probes were synthesized using the digoxigenin (DIG) RNA Labeling Kit (Roche, Switzerland). All synthesized probes were treated with RNase-free TURBO DNase (Ambion). Chromogenic and fluorescent ISH staining was carried out with BCIP/NBT substrates (Roche) and TSA™ Plus Fluorescence Systems (NEN Life Science), respectively. Nuclear staining was carried out with 4,6-diamino-2-phenylindole (DAPI) in the Gold Antifade reagent (Invitrogen).
Cell culture
The SG3 cells were cultured as previously described [14,16]. To test the biological activity of OlGfrα1a and OlGfrα1b, SG3 cells were incubated in ESM4, 5N, or 5N plus medaka Gdnfa (10 ng/mL) or Gdnfb (10 ng/mL) recombinant proteins [14]. ESM4 is the Dulbecco's modified Eagle's medium (DMEM) supplemented with 20 mM HEPES, 100 U/mL penicillin and streptomycin, 15% fetal bovine serum, the nonprotein supplement combination (2 mM
Vivo-Morpholino oligos and reporter assay
The Vivo-Morpholino oligos gfrα1aMo (5′-GCGCATAAACGACGGCAAGAAT
EdU labeling and Annexin V-FITC/PI staining
The SG3 cells were incubated in 5N plus Gdnfa and/or Gdnfb, then gfrα1aMo and/or gfrα1bMo was added, respectively. ConMo with the same corresponding concentration was used as the control. After 48 h of incubation, the proliferative activity and apoptosis of SG3 cells were measured by using EdU (5-ethynyl-2-deoxyuridine) labeling kit (RIBOBIO, Guangzhou, China) and Annexin V-FITC/PI kit (BD) as previously described, respectively [14]. The fluorescence signals were observed under Nikon Ti-S inverted fluorescence microscope.
Real-time PCR
After 7 days of incubation with Vivo-Morpholino oligos, the SG3 cells were harvested to detect the mRNA expression levels of target genes by real-time PCR using delta-delta Ct (threshold crossing value) calculations. Primer pairs used in this study were gene-specific and intron-spanning (Supplementary Table S1). Real-time PCR was performed on the StepOnePlus™ Real-Time PCR System (Life Technologies) as previously described [14].
Testicular organ culture
The testis from each adult medaka was dissected and separated into two parts; one was used for experimental treatment and the other for the control. The separated testicular parts were seeded in 24-well plate and incubated in ESM4 medium. Twenty-four hours later, both gfrα1aMo (2.5 μM) and gfrα1bMo (2.5 μM) were added. After 3 days of onward treatment, the proliferation activity of spermatogonia was measured with EdU incorporation assay. The same concentration of ConMo (5 μM) was added as the control.
Statistical analyses
Statistical analyses were performed with Student's t-test for the comparison between two groups. P < 0.05 or P < 0.01 is considered as significant or very significant differences from the control, respectively.
Results
cDNA sequences and sequence analyses
The Olgfrα1a and Olgfrα1b contain a 1,575-bp and 1,401-bp ORF encoding 524- and 466-amino acid residues (aa), respectively (Supplementary Fig. S1). Both the predicted OlGfrα1a and OlGfrα1b contain 3 cysteine-rich domains with an 18- or 17-aa putative signal peptide, 3 potential N-glycosylation sites, and 31 cysteine residues (Supplementary Fig. S1). A multiple Gfrα1 sequence alignment among representative tetrapods, zebrafish, and medaka reveals that the three cysteine-rich domains (D1–3) are highly conserved across vertebrates (Fig. 1). Additionally, the 10 cysteine residues and 2 triplets (MLF and RRR) in D2 are completely conserved, the latter of which is reported to be important for Gdnf binding in mammals [18] (Fig. 1). OlGfrα1a and OlGfrα1b have identity 52% to each other, and over 52% and 55% to other indicated vertebrates, respectively (Fig. 1).

GFRα1 amino acid sequence alignment. Numbers on the right indicate amino acid positions, dashes indicate deletions, shaded areas indicate shared sequences. The 3 cysteine-rich domains (D1–3), 30 cysteine residues (*), 2 triplets (MLF and RRR) (boxed) and several other motifs (#) critical for GFRa1 binding to GDNF and eliciting downstream signal transduction are highly conserved among examined vertebrates. At the end of the alignment are the percentage identity values of OlGfrα1a and OlGfrα1b to the other corresponding sequences. The GenBank accession number for each sequence refers to Fig. 2. GFRα1, glial cell-derived neurotrophic factor family receptor alpha-1.
On the phylogenetic tree, fish Gfrα1 clustering with tetrapod GFRα1 forms a separate clade, which is distinct from the clades formed by other members of GFRα family, including GFRα2, GFRα3, GFRα4, GRAL (GFRα like), and Gas1 [19]. In the GFRα1 clade, fish-specific Gfrα1a and Gfrα1b diverges and forms two fish-specific subclades, which are consistent with the third round of fish-specific genome duplication after separation between fish and tetrapod lineages (Fig. 2A). A cross-species comparison of chromosome neighboring genes reveals that both Olgfrα1a- and Olgfrα1b-containing regions are syntenic to human Gfrα1-containing chromosome 10, which is the same case for zebrafish (Fig. 2B). Moreover, Olgfrα1a and Olgfrα1b exhibit a similar genomic organization to each other and human Gfrα1, except for one exon less between D1 and D2 in Olgfrα1b (Fig. 2C). These data support that OlGfrα1a and OlGfrα1b are orthologous to mammalian GFRα1.

Phylogenetic tree, chromosomal synteny, and genomic organization.
The mRNA expression profile of Olgfrα1a and Olgfrα1b
Reverse transcription–PCR analyses revealed that the Olgfrα1a and Olgfrα1b transcripts were present in embryos at all tested developmental stages (from morula to fry) (Supplementary Fig. S2). Meantime, both of them were high in the SG3 cells, eye, heart, intestine, and testis and were moderate in the liver, spleen, ovary, and brain (Supplementary Fig. S2).
ISH was performed to examine the cellular type of Olgfrα1a and Olgfrα1b expression in the testis. The medaka testis is composed of seminiferous cysts; spermatogenesis proceeds synchronously at each cyst; spermatogonia are predominantly at the most peripheral region (Fig. 3A1, B1). Both chromogenic and fluorescent ISH indicated that both Olgfrα1a and Olgfrα1b transcripts were high in spermatogonia, moderate in spermatocytes, and barely detectable in spermatids and sperms (Fig. 3A2–3, B2–3). At higher magnification, the Olgfrα1a RNAs were mainly distributed in the cytoplasm, whereas Olgfrα1b in both the cytoplasm and nucleus (Fig. 3A4, B4). These data suggest that Olgfrα1a and Olgfrα1b have similar expression profile except for the cellular distribution in the adult testis.
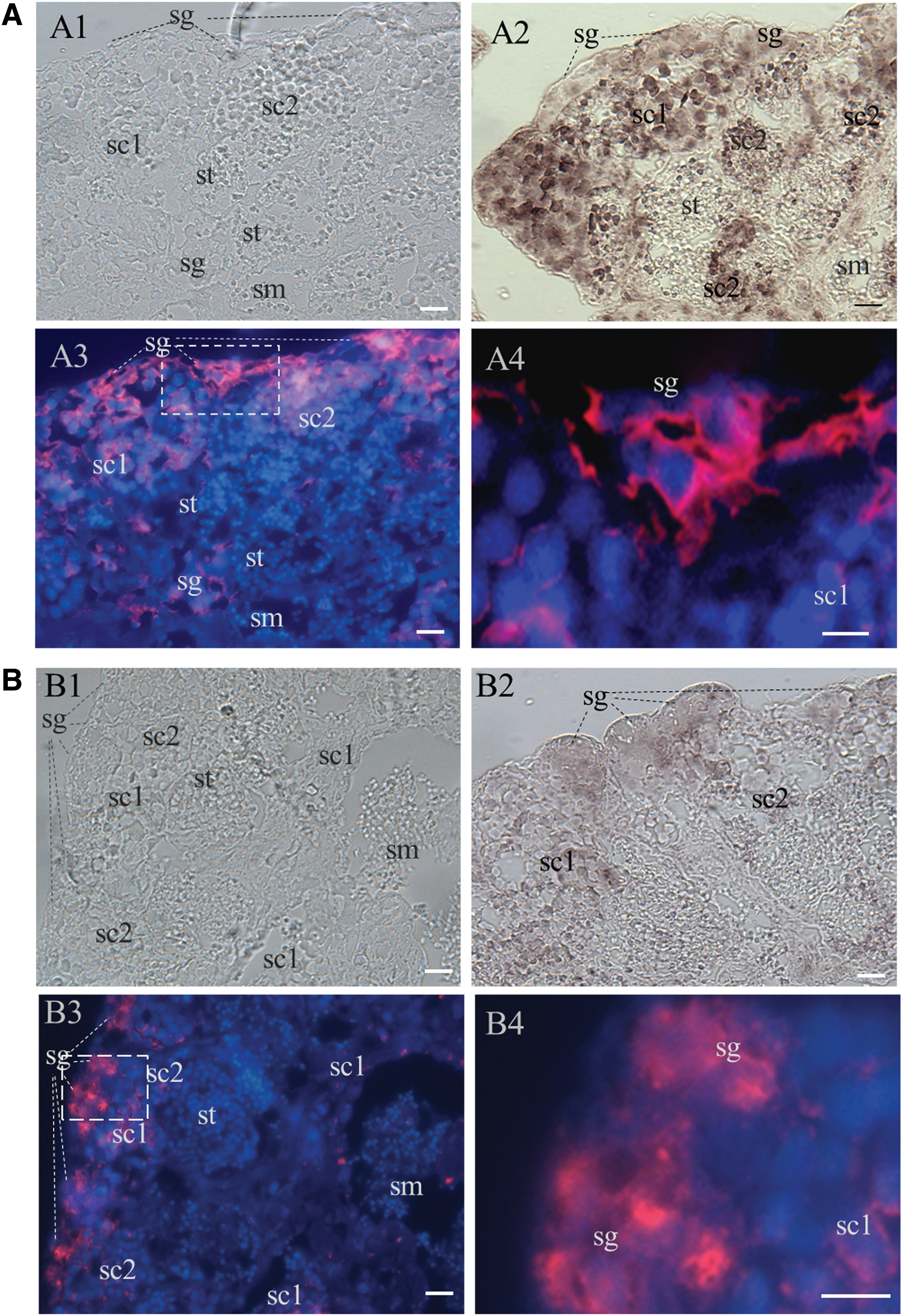
Olgfrα1a
Knockdown of Olgfrα1a and/or Olgfrα1b reduces the proliferation activity of SG3
In medaka, a cell line (SG3) of male germ stem cells spermatogonia has been successfully developed, which shows the intrinsic property of SSCs by self-renewal and differentiation potential [16]. Moreover, our previous study suggests that the proliferative activity of SG3 cells could be significantly promoted by medaka Gdnfa and Gdnfb [14], which makes it an ideal model to evaluate the biological activity of OlGfrα1a and OlGfrα1b.
Due to the absence of antibody specific to medaka Gfrα1a or Gfrα1b, the reporter vectors pCV1a-DsRed and pCV1b-DsRed were used to detect the knockdown specificity of the Vivo-Morpholino oligos gfrα1aMo and gfrα1bMo, respectively. The ConMo and pCV-DsRed were used as the internal controls. Strong fluorescent signal could be observed in the cells transfected with pCV-DsRed, pCV1a-DsRed, and pCV1b-DsRed together with 2.5 μM ConMo treatment, respectively (Fig. 4A). Significantly, 2.5 μM gfrα1aMo treatment reduced over 90% of DsRed signal in pCV1a-DsRed-transfected cells and had no effect in pCV-DsRed- or pCV1b-DsRed-transfected cells (Fig. 4B). Similarly, 2.5 μM gfrα1bMo specifically reduced over 90% of the DsRed signal in pCV1b-DsRed-transfected cells and had no effect in pCV-DsRed- or pCV1a-DsRed-transfected cells (Fig. 4C). These data suggest that gfrα1aMo and gfrα1bMo show high specificity to their target sequence with high efficiency, respectively.
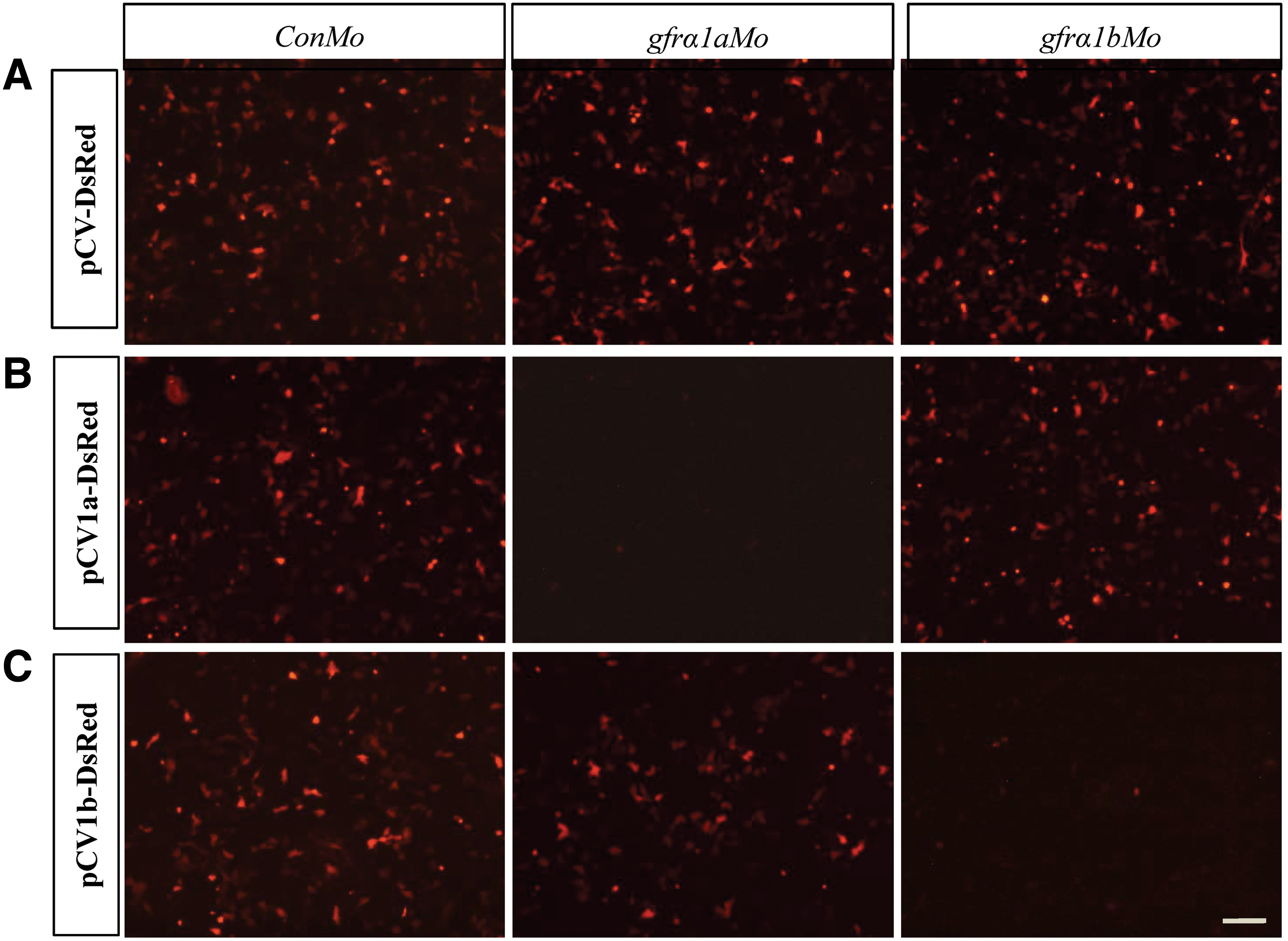
The specificity of gfrα1aMo and gfrα1bMo. The SG3 cells were transfected with the control pCV-DsRed and the reporter vectors pCV1a-DsRed and pCV1b-DsRed by TransIT-2020, respectively. At 2 h post-transfection, 5 μM Vivo-Morpholino oligos were added into the cells, respectively.
The SG3 cells were incubated in 5N, 5N plus Gdnfa (10 ng/mL), and 5N plus Gdnfb (10 ng/mL), respectively, and treated with gfrα1aMo and/or gfrα1bMo. The proliferative activity of cells was measured by EdU incorporation staining. In 5N plus Gdnfa, the proliferation activity of SG3 cells showed a significant decrease with the knockdown of either OlGfrα1a or OlGfrα1b compared with the control (P < 0.05); it further decreased with the knockdown of both OlGfrα1a and OlGfrα1b, and with the concentrations of gfrα1aMo and gfrα1bMo increased (from 2.5 to 5 μM) (P < 0.05) (Fig. 5A). In 5N plus Gdnfb, the proliferation activity of SG3 cells showed a significant decrease with the knockdown of OlGfrα1a in a dose-dependent manner (P < 0.05), whereas no effect with the knockdown of OlGfrα1b (P > 0.05) (Fig. 5B). In contrast, in 5N medium containing no Gdnfa or Gdnfb, the proliferation activity of SG3 cells showed no significant difference with the knockdown irrespective of Olgfrα1a or Olgfrα1b compared with the control (P > 0.05) (Supplementary Fig. S3). Taken together, these results suggest that OlGfrα1a might mediate the proliferation of SG3 cells through binding Gdnfa or Gdnfb, whereas OlGfrα1b through binding Gdnfa not Gdnfb.
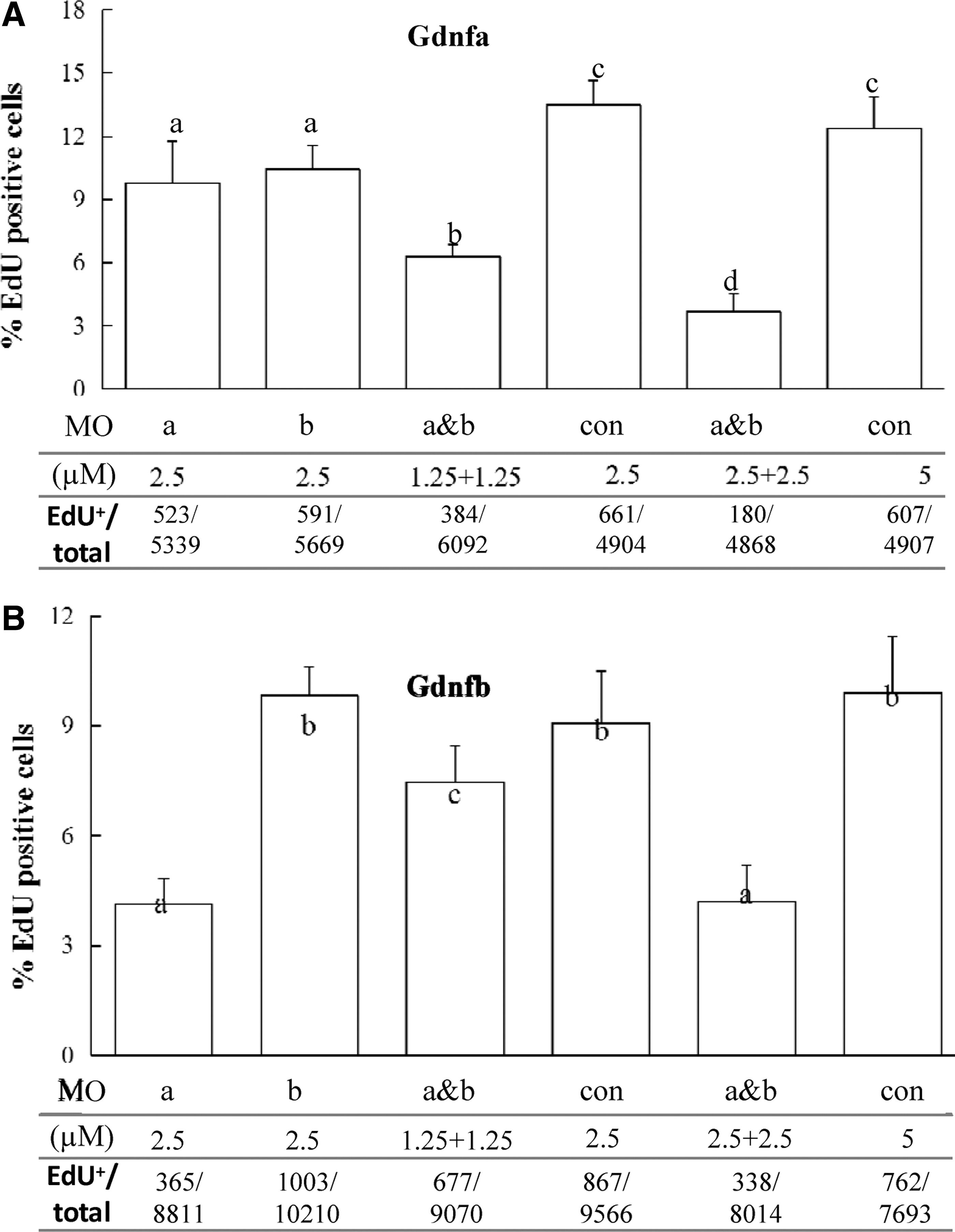
The proliferation of SG3 with the knockdown of OlGfrα1a and/or OlGfrα1b in 5N plus Gdnfa
Knockdown of Olgfrα1a and Olgfrα1b affects the survival of SG3
The SG3 cells were treated with gfrα1aMo and/or gfrα1bMo in 5N plus both Gdnfa (10 ng/mL) and Gdnfb (10 ng/mL) for 48 h and then were measured by Annexin V-FITC/PI staining assay. Unexpectedly, the SG3 cells did not show significant apoptosis with the knockdown of either OlGfrα1a or OlGfrα1b compared with the control (P > 0.05) (Fig. 6). However, with the knockdown of both OlGfrα1a and OlGfrα1b, the cells showed a significant induced apoptosis (P < 0.05) (Fig. 5). These results show that OlGfrα1a and OlGfrα1b are critical for the survival of SG3, and both are functionally equivalent and could compensate for each other.
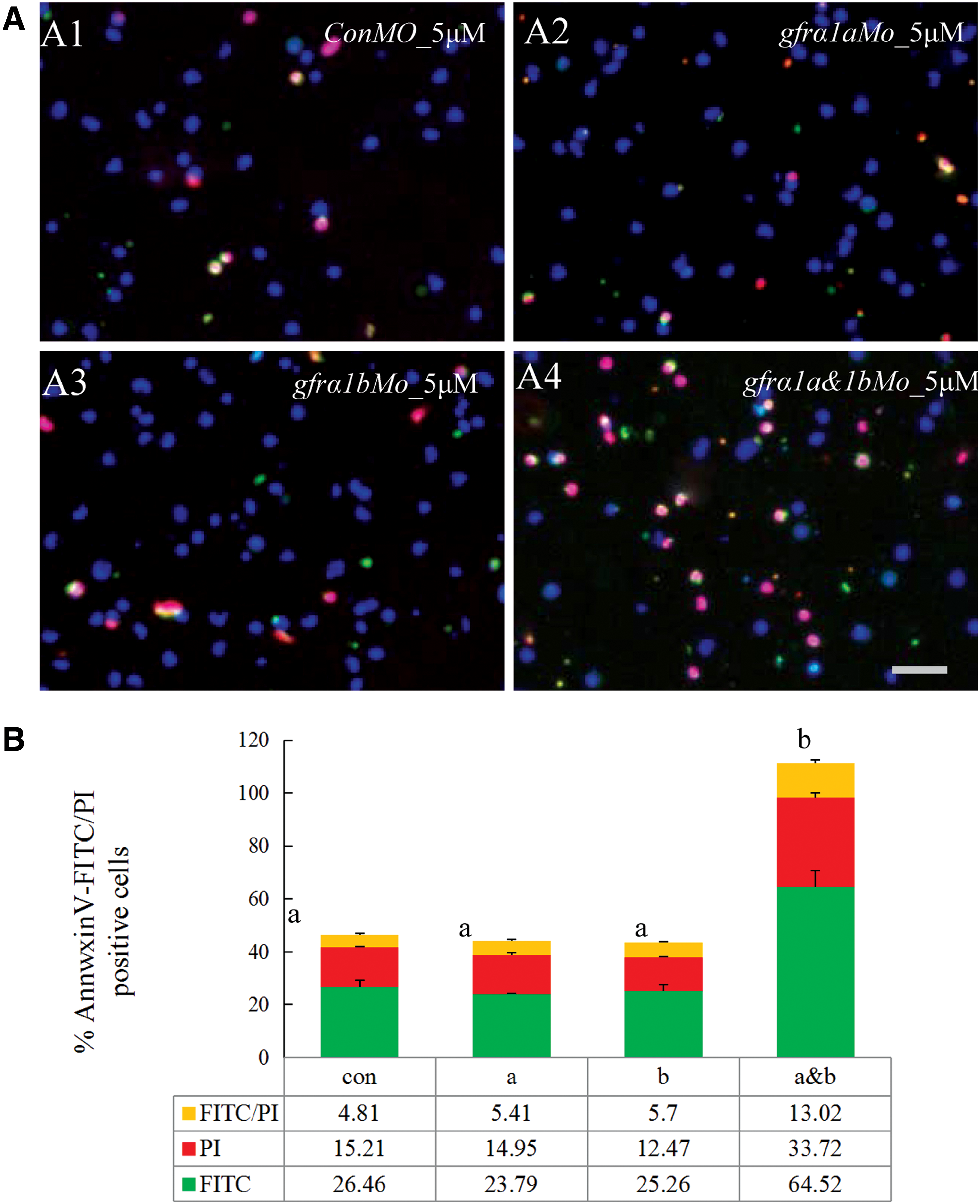
The knockdown of both OlGfrα1a and OlGfrα1b induced apoptosis.
The mRNA expression levels of pluripotency and differentiation-related genes
We used real-time PCR to evaluate the transcriptional regulation with the knockdown of OlGfrα1a and/or OlGfrα1b in SG3 cells. After 7 days of treatment, the expression of pluripotency-related genes including klf4, lin28b, bcl6b, and etv5 was downregulated, whereas the expression of differentiation-related gene c-kit was upregulated (P < 0.05) (Fig. 7). However, the expression of other differentiated markers including sycp3, dmc1, and spata22 was not observed (data not shown). These results suggest that OlGfrα1a and OlGfrα1b might mediate the self-renewal and maintenance of SG3 by upregulating the expression of pluripotency-related genes and inhibiting the differentiation gene.
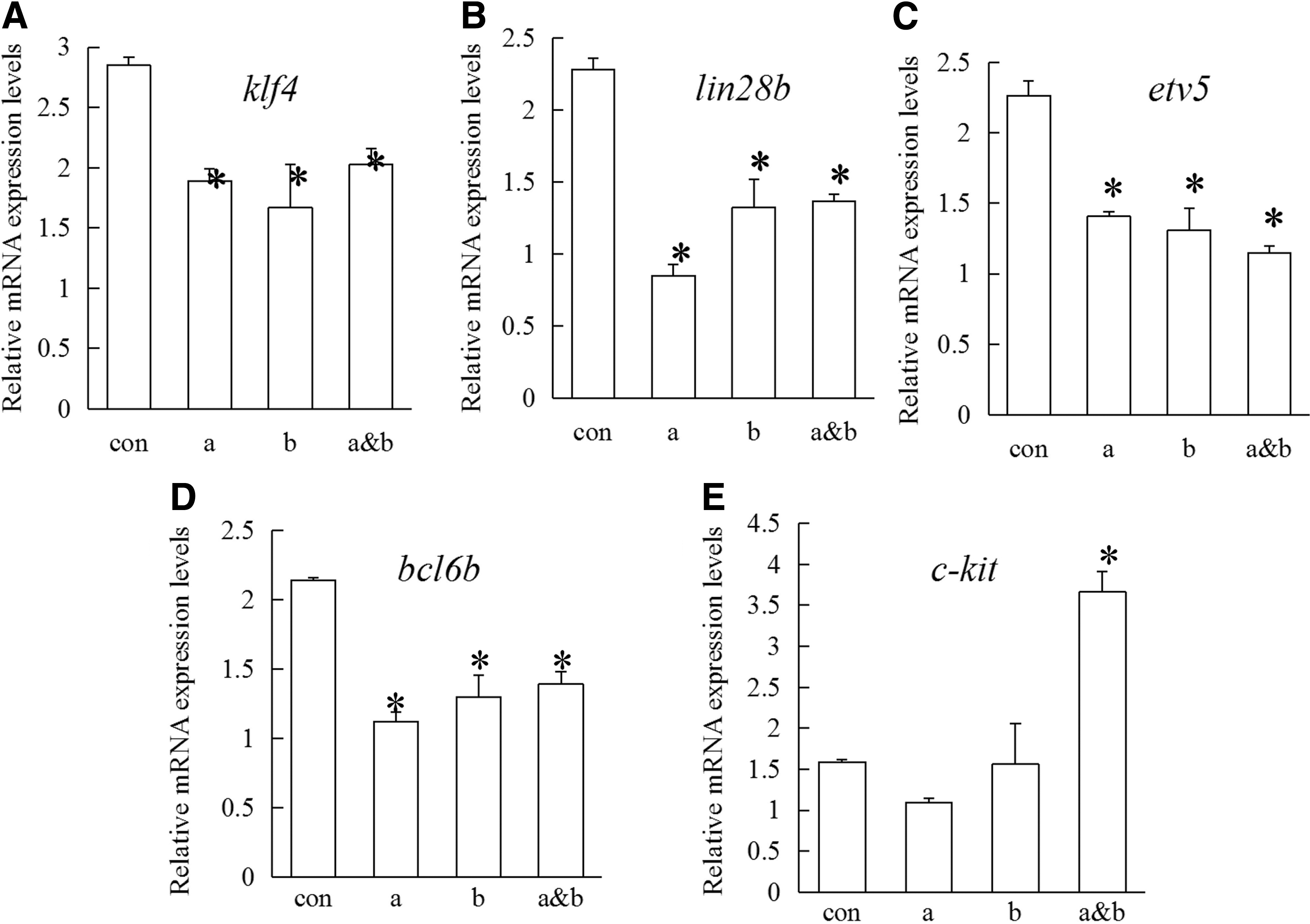
The mRNA expression levels of SG3 with the knockdown of OlGfrα1a and/or OlGfrα1b in ESM4. SG3 were treated with gfrα1aMo and/or gfrα1bMo in ESM4 medium, and then, total RNAs were harvested after 7 days of treatment for quantitative reverse transcription–polymerase chain reaction analyses. The mRNA expression levels of klf4
Knockdown of Olgfrα1a and Olgfrα1b affects the proliferation activity of spermatogonia in testis
In the control treatment, the percentage of EdU-positive spermatogonia was about 66.2%. Meanwhile, the percentage of EdU-positive spermatogonia reduced to 27.8% with the knockdown of both OlGfrα1a and Olgfrα1b (Fig. 8). Thus, our results show that the knockdown of Gfrα1a and Gfrα1b can inhibit the proliferation of spermatogonia in the adult testis.
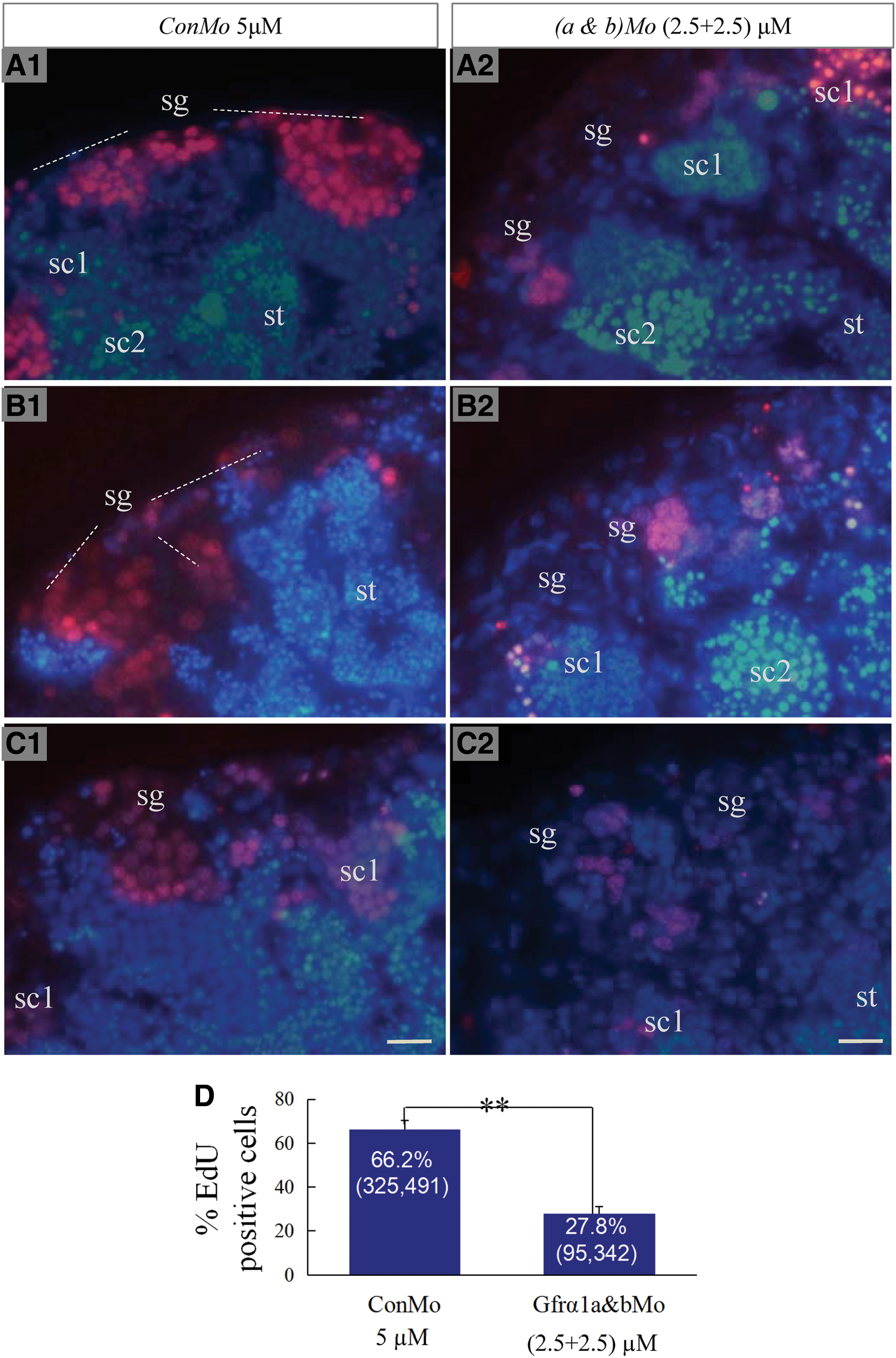
The proliferation of spermatogonia with the knockdown of OlGfrα1a and OlGfrα1b in adult testis. The testicular tissues were treated with gfrα1aMo
Discussion
In this study, we have shown that both Olgfrα1a and Olgfrα1b, the gfrα1 duplicate genes from medaka, are homologous to mammalian GFRα1 not only by sequence and chromosome synteny but also by expression and function. Four independent lines of evidences support this notion. First, the predicted OlGfrα1a and OlGfrα1b show high sequence identity to GFRα1 from the other vertebrates examined and contain the sequence characteristic of GFRα family members, including a putative signal peptide, three conserved cysteine-rich domains, and a C-terminal hydrophobic domain separated by a hydrophilic linker region for glycosylphosphatidylinositol linkage. Second, on the phylogenetic tree, OlGfrα1a and OlGfrα1b are clustered with other GFRα1 to form a separate clade, suggesting their ortholog to mammalian GFRα1 rather than to the other members of GFRα superfamily (Fig. 3A). A cross-species comparison of chromosome locations further supports this. Both medaka chromosome 15 in Olgfrα1a-containing region and 19 in Olgfrα1b-containing region are syntenic to human chromosome 10 in the Gfrα1-containing region, suggesting that Olgfrα1a and Olgfrα1b are the duplicate genes diverged from the teleost-specific whole-genome duplication events [20]. Third, both Olgfrα1a and Olgfrα1b show high expression in spermatogonia in adult testis, which are similar to the mammalian counterparts. Finally and most importantly, both are involved in the self-renewal and maintenance of SSCs. Taken together, these data suggest that both OlGfrα1a and OlGfrα1b might have conserved function as mammalian counterpart in SSCs.
About 350 million years ago, teleost fish experienced a third round of whole-genome duplication event, which resulted in a large number of duplicate genes [20]. The duplicated genes might lead to nonfunctionalization, functional redundancy, subfunctionalization, or neofunctionalization [20]. In the present study, the medaka Gfrα1 contains two homologs (ie, OlGfrα1a and OlGfrα1b). Sequence analysis implies that OlGfrα1a and OlGfrα1b might be functionally redundant. The two homologs share identity 52% to each other and contain common features characteristic of the Gfrα1 family. Particularly, both contain multiple conserved residues and motifs critical for GFRα1 binding to GDNF and eliciting downstream signal transduction, such as 207NN208 or 148NN149 (152NN153), R319 or R260 (R259), 375SNS377 or 316SSS318 (316SNS318) in the OlGfrα1a or OlGfrα1b (mouse Gfrα1), the two triplets (MLF and RRR) in both central regions (Fig. 1) [19,21 –23]. In mice, site-directed mutagenesis and functional assays showed that 152NN153, R259, and 316SNS318 in GFRα1 is crucial not only for GFRα1 and RET binding but also for activation of the receptor complex [23]. Similarly, the MLF and RRR triplets are highly conserved among members of the GFRα1 family and critical for the interaction of GFRα1 with GDNF, which have complementary properties to the receptor binding surface identified in GDNF with hydrophobic and negatively charged residues [19,22 –24].
Accumulative studies have demonstrated that GFRα1 is a conserved marker of undifferentiated spermatogonia in a wide spectrum of mammals, including human, monkey, pig, mouse, rat, and hamster [6,25 –29]. A recent study reports that GFRα1-positive SSCs could be accumulated owing to stage-specific ectopic expression of GDNF [30], suggesting the crucial role of GDNF/GFRα1 signaling pathway in the self-renewal of SSCs. In the present study, both Olgfrα1a and Olgfrα1b were strongly expressed in spermatogonia with differential cellular distribution, implying their biological activity in the self-renewal/differentiation process of spermatogonia. By the way, the moderate mRNA expression of Olgfrα1a and Olgfrα1b was detected in spermatocytes, not in spermatids and sperms (Fig. 3). In chicken, Gfrα1 was observed in spermatogonia, spermatocytes, and round spermatids [31]. The differential expression profiles of GFRα1 between species suggest that it might be involved with the different stages of spermatogenesis.
The GDNF/GFRα1 signaling pathway ensures the survival and self-renewal of SSCs in mammals; a long-term in vitro culture system of SSCs in rodents including mouse, rat, and hamster has been developed by supplementing with GDNF in the culture media [1,2,5,10]. However, whether the GDNF/GFRα1 signaling pathway in SSCs is functionally conserved among vertebrate phyla is still far from clear. Our knowledge about fish SSCs is very limited. Several reports together with our previous work indicate that Gdnf widely exist in fish [13,14,32,33]; moreover, the medaka Gdnfa and Gdnfb can mediate the self-renewal and maintenance of SG3 [14]. However, what their receptors are and how they work in fish SSCs still remain to be elucidated.
In this study, Vivo-Morpholinos were chosen to investigate the function of OlGfrα1a and OlGfrα1b in SSCs. A Vivo-Morpholino is composed of a Morpholino oligo with a unique covalently linked octa-guanidine dendrimer in a conformation effective to penetrate cell membranes, which is shown to effectively enter and function within cultured cells and a wide variety of tissues [34]. A large number of reports have demonstrated that Vivo-Morpholinos are a promising tool for gene silencing in vitro and in vivo, especially for those of which gene knockout is lethal during early embryo development [34 –36]. For instance, it reports that the knockout of GFRα1 resulted in the death of newborn mice within 24 h due to uremia [37]. Another advantage of Vivo-Morpholinos is that simultaneous inactivation of two or more genes can easily be accomplished, thus offering the opportunity to study redundant and/or combined effects of interesting genes. Additionally, gene silencing of ex vivo cultured tissues from the same individual allows for direct and strict comparisons between the Morpholino-specific and the control effects.
In SG3, the Vivo-Morpholino against OlGfrα1a or OlGfrαb could specifically knockdown their protein expression, as shown by over 90% reduced DsRed signal of the reporter vectors pCV1a-DsRed and pCV1b-DsRed, respectively (Fig. 4). Importantly, the knockdown of OlGfrα1a and/or OlGfrα1b expression by Vivo-Morpholinos resulted in the dramatically decreased proliferative activity of SSCs, as evidenced by the reduced percentage of proliferative (EdU-positive) cells in SG3 cells and spermatogonia in the testicular organ culture (Figs. 5 and 8). Moreover, our data suggest that both OlGfrα1a and OlGfrα1b are critical for the survival of SG3, as evidenced by the highly significant induced-apoptosis with the knockdown of both OlGfrα1a and OlGfrα1b (Fig. 6). By the way, no apparent apoptosis occurred with the knockdown of either OlGfrα1a or OlGfrα1b, which might be attributed to the functional redundancy in 5N plus Gdnfa and Gdnfb (Fig. 6). Consistently, in mouse, the knockdown of GFRα1 expression by small-interfering RNAs resulted in the suppression of SSC proliferation, a phenotypic differentiation, and a decrease in RET phosphorylation [11]. Collectively, our data indicate that both OlGfrα1a and OlGfrα1b are involved in the proliferation and survival of the medaka SSCs, suggesting that the GDNF/GFRα1 signaling pathway might be functionally conserved in SSCs among vertebrate phyla.
Studies show that KLF4 as well as LIN28b are RNA-binding pluripotent stem cell factors, which are strongly expressed in SSCs and critical for their self-renewal and maintenance [38,39]; BCL6b and ETV5 are important regulators in the self-renewal of SSCs [5,40 –44], and the Bcl6b- or Etv5-null mouse displayed an impairment of spermatogenesis, resulting in a Sertoli cell-only phenotype and aspermia [43 –46]; the C-KIT is used as a marker for differentiating spermatogonia and considered to be critical for the type A1–A4 spermatogonia divisions [47 –49]. Our previous study indicates that the expression level of bcl6b was upregulated after 4 h of Gdnf treatment in SG3 cells, whereas etv5 appeared no change [14]. In the current study, the knockdown of OlGfrα1a and/or OlGfrα1b in SG3 for 7 days resulted in the downregulation of klf4, lin28b, bcl6b, and etv5, whereas upregulation of c-kit (Fig. 7). These data imply that the OlGfrα1a and OlGfrα1b might mediate the self-renewal of SSCs through promoting proliferation, maintaining pluripotent property, and inhibiting differentiation (Fig. 9) The expression of other differentiated markers including sycp3, dmc1, and spata22 was not observed (data not shown), suggesting that the SG3 cells did not differentiate into late spermatocytes or spermatids.

The Gdnf/Gfrα1 signaling in the self-renewal and maintenance of SSCs in medaka. By binding Gdnfa and/or Gdnfb, the Gfrα1a and Gfrα1b might enhance the proliferation, survival, and maintenance of SSCs and upregulate the expression levels of klf4, lin28b, bcl6b, and etv5 (the solid line arrow). Meanwhile, the Gdnf/Gfrα1 signaling might inhibit the differentiation of SSCs and downregulate the expression level of c-kit (the dotted arrow). SSC, spermatogonial stem cell.
In 5N medium, the proliferation activity of SG3 cells showed no significant difference with the knockdown OlGfrα1a and/or OlGfrα1b (Supplementary Fig. S3), which provides an ideal model to investigate the reactivity and specificity of ligand receptor in 5N plus Gdnfa or Gdnfb. With the knockdown of OlGfrα1a, the proliferation activity of SG3 cells showed significantly decrease in 5N plus Gdnfa or Gdnfb, suggesting that OlGfrα1a could function by binding either Gdnfa or Gdnfb (Fig. 9). Meanwhile, with the knockdown of OlGfrα1b, the proliferation activity of SG3 cells showed significant decrease in 5N plus Gdnfa but not Gdnfb, suggesting that OlGfrα1b could function only by binding Gdnfa not Gdnfb (Fig. 9). Nevertheless, further study needs to be carried out to investigate the reactivity of Gdnf/Gfrα1 in vivo.
In conclusion, our study first demonstrates that the two homologs of medaka Gfrα1 are predominantly expressed in spermatogonia, and their knockdown could greatly inhibit the self-renewal and maintenance of SSCs. This study provides insight into the self-renewal and maintenance of SSCs in lower vertebrates just like fish.
Footnotes
Acknowledgments
This work was supported by grants XDJK2017B007, XDJK2017D102, and XDJK2017D103 from Fundamental Research Funds for the Central Universities (Ministry of Education of China), grants 31630082, 31502147 and 31572597 from the National Natural Science Foundation of China, and grant cstc2016jcyjA0529 from the Natural Science Foundation Project of CQ CSTC.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.