
Abstract
Mesenchymal stromal cells (MSCs) in the tumor microenvironment (TME) participate together with tumor cells to suppress antitumor effector cells through the production of immunosuppressive factors, such as transforming growth factor-beta 1 (TGF-β1). Furthermore, TGF-β1 can induce 5′-nucleotidase (CD73) expression in various cell types; this functional activity is associated with the production of adenosine (Ado), which is an immunosuppressive nucleoside. In this study, we provide evidence that coculture of MSCs derived from cervical tumors (CeCa-MSC) with CeCa tumor cells increases CD73 expression in tumor cells and the capacity of these cells to generate Ado in a MSC ratio-dependent manner. Interestingly, the increase in CD73 in the CeCa cell membrane corresponded to an increase in the TGF-β1 expression level in the tumor cells and the TGF-β1 content in the supernatants of the CeCa/CeCa-MSC cocultures. The addition of anti-hTGF-β neutralizing antibodies strongly reversed CD73 expression in the tumor cells. This phenomenon was not exclusive to CeCa-MSCs; coculture of MSCs derived from the normal cervix with CeCa cells produced similar results. These results suggest that the interaction of MSCs with CeCa tumor cells in the TME may condition higher TGF-β1 production to maintain an immunosuppressive status not only through the activity of this cytokine per se but also through its ability to induce CD73 expression in tumor cells and generate an immunosuppressive microenvironment rich in Ado.
Introduction
Mesenchymal stem/stromal cells (MSCs) are a heterogeneous subtype of stem cells that can be obtained from different mature tissues. These cells have the ability to differentiate into pericytes, osteoblasts, adipocytes, chondroblasts, fibroblasts, myoblasts, oocytes, cardiomyocytes, hepatocytes, tenocytes, different epithelial cells (lung, intestine, kidney, and spleen), and even neurons and secrete growth factors with paracrine effects that promote the regeneration of damaged tissues [1 –3]. MSCs have the ability to migrate and settle in sites of inflammation and have been found to be part of the tumor microenvironment (TME). In the latter site they are able to regulate tumor growth and progression through paracrine interactions that have not been fully elucidated [4,5]. MSCs were obtained for the first time from bone marrow (BM) and were characterized phenotypically as presenting the markers, CD105, CD73, CD90, and CD248, expressing low levels of major histocompatibility complex (MHC) class I molecules and not expressing the MHC class II, CD11b, CD14, CD34, CD45, and CD31 molecules [6,7].
MSCs in the TME (T-MSCs) can constitute ∼0.01%–1% of the total number of cells in solid tumor tissues [8]. These cells have been isolated and characterized from ovarian cancer cells [9], giant cell tumors [10], neuroblastoma [11], osteosarcoma, [12], gastric cancer [13], and cervical cancer (CeCa) [14], among others. T-MSCs can support and promote the proliferation of cancer stem cells in addition to promoting the epithelial–mesenchymal transition (EMT) and tumor cell metastasis [9,15]. Likewise, these cells can favor the evasion of immune recognition mediated by cytotoxic T lymphocytes by deregulating MHC class I molecule expression in tumor cells [14] and suppressing the antitumor immune response through either recruitment of immunosuppressive cells, such as myeloid-derived suppressor cells (MDSCs) and regulatory T cells (Tregs) [16], the production of immunosuppressive cytokines, such as interleukin (IL)-4, IL-5, IL-10, and TGF-β [17,18], or the production of immunosuppressive metabolites, such as PGE2, kynurenine, and NO [19].
Recently, our research group reported that MSCs derived from CeCa (CeCa-MSCs) suppressed T lymphocyte responses through the purinergic pathway [20]. In this pathway, the adenosine triphosphate/adenosine diphosphate (ATP/ADP) nucleotides found in high concentrations (50–100 μM) in response to stress signals, such as hypoxia, damage, and inflammation in the TME [21], are hydrolyzed jointly by the CD39 ectoenzymes (ENTPD1, ectonucleoside triphosphate diphosphohydrolase-1, EC 3.6.1.5) to generate the respective nucleotides. Subsequently, the nucleotides are hydrolyzed by the action of 5′-ectonucleotidase or CD73 (EC 3.1.3.5), which converts adenosine monophosphate (AMP) into adenosine (Ado) [22,23]. Most of the extracellular signaling activities of Ado are mediated by adenosine receptors (ARs) on the cell surface, which are coupled to G proteins and divided into four subtypes (A1R, A2AR, A2BR, and A3R) [24 –26].
Transcriptional activation of CD73 within the TME seems to be regulated by the hypoxia-inducible transcription factor (HIF-1)α [27] and the presence of immunosuppressive factors, such as transforming growth factor beta (TGF-β), which can stabilize HIF-1α [28]. Overexpression of CD73 in different tumors has been reported to be associated with a poor prognosis and a greater ability to suppress antitumor immune responses [29 –31].
In the particular case of CeCa, persistent infection caused by high-risk human papilloma viruses (HPV-ARs) plays an important role in cervical carcinogenesis and is strongly associated with the production of immunosuppressive cytokines, such as TGF-β; TGF-β expression correlates directly with the degree of CeCa progression and suppression of the antitumor immune response [32]. TGF-β has been detected in sera and tissues from patients infected with HPV-ARs, in low- and high-grade squamous intraepithelial lesions (SILs) and in CeCa; in addition, the levels correlate directly with the degree of progression of the lesion [33,34] and inhibition of the immune response in patients with SILs [35]. TGF-β is an immunoregulatory cytokine that participates in tumor progression and immunosuppression, and the sources of this factor in the TME include several types of stromal cells in addition to cancer cells [36 –38]. Recently, we reported that CeCa-MSCs induced the expression and secretion of TGF-β in CeCa tumor cells [39]. Based on this background, in this work, we analyze the ability of CeCa-MSCs to induce CD73 expression in CeCa tumor cells through TGF-β production. To determine whether these properties are unique to CeCa-MSCs, we also use MSCs obtained from normal cervical tissues (NCx-MSCs). Finally, to evaluate whether the induction of CD73 in tumor cells is attributable to TGF-β, we employ anti-TGF-β neutralizing antibodies.
Materials and Methods
Cell culture
CaSki (HPV16+) and MS751 (HPV-18+) human cervical carcinoma cell lines used in this study were obtained from the American Type Culture Collection (ATCC). MSCs derived from NCx-MSCs were obtained from tissue samples from three people who underwent a noncancer hysterectomy. The CeCa-MSCs were obtained from biopsies of three patients with stage IIIB CeCa whose histopathological diagnosis was confirmed by the Pathology Department. The local Ethics Committee approved these procedures. The MSCs were obtained by enzymatic digestion and subsequently characterized by their phenotype and ability to differentiate into adipocytes, chondrocytes, and osteocytes as previously described [14]. CaSki and MS751 CeCa cell lines were maintained in RPMI-1640 medium (Gibco Laboratories, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; Gibco, CA), and the MSCs were maintained in low-glucose Dulbecco modified Eagle's medium (Lg-DMEM; Gibco) supplemented with 15% FBS, 100 IU/mL of penicillin, 100 μg/mL of streptomycin (Gibco), and 5 mM
Cell cocultures
Cocultures of tumor cells and MSCs were performed in 50% RPMI-1640 medium and 50% low-glucose DMEM supplemented with 15% FBS. The cells were cocultured for 96 h in transwell chambers (Millicell Chamber; Millipore Corporation, Billerica, MA) at 1:1, 1:2.5, and 1:5 CeCa:MSC cell ratios.
CD73 expression in CeCa cells
CD73 ectonucleotidase expression in the membranes of CeCa tumor cells cocultured for 96 h with MSCs and in the presence or absence of different concentrations (0.5, 10, 20, and 40 ng/mL) of recombinant human-TGF-β1 (rh-TGF-β1) (PeproTech, Inc., Rocky Hill, NJ) was determined by flow cytometry using an anti-CD73-PE monoclonal antibody (BD Biosciences, San Diego, CA). Functional activity of rh-TGF-β1 was determined by using TGF-β-sensitive Mv1Lu cells, as previously described [40]. The analysis was performed based on the acquisition of 25,000 events in a FACS ARIA flow cytometer (BD Biosciences). The results were analyzed and reported as the mean fluorescence intensity ± standard deviation compared with basal CD73 expression in CeCa cells, which was normalized to a value of 1. All experiments were performed in triplicate. Staining with a control isotype antibody was used as a negative control.
CD73 mRNA expression in CeCa cells
To determine the CD73 mRNA expression level in CeCa cells either alone or after cocultivation with the different MSC ratios, RNA was extracted with TRIzol (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. cDNA was synthesized from 500 ng/μL of RNA using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). The polymerase chain reaction (PCR) for CD73 was performed in a real-time Light Cycler 480 PCR system (Roche Diagnostic, Manheim, Germany) using ProbeLibrary Universal Probes. To control variations in the RNA amounts, H6PDH was used as an internal control gene. The primers specific for CD73 and G6PDH were as follows: CD73 (forward: 5′ACTACTACGCCAAGGAGGTCAC3′, reverse: 5′TGCTTGAACTTGTCATAGATTTCG3′) and G6PDH (forward: 5′GCTACGCTCGGATCTTGTTC3′, reverse: 5′CCCAGTGCTTTTCGCTCT3′). The data analysis was performed using the ΔCT method for relative quantification. The thermocycling conditions for each reaction were performed in a final volume of 10 μL according to the protocol provided with the LightCycler 480 Probe Master (Roche Diagnostic).
Hydrolytic activity of CD73 in CeCa cells
To determine the enzymatic activity of the ectonucleotidase CD73 in CeCa cells previously cocultured for 96 h with MSCs or in the presence or absence of TGF-β, the cells were washed twice with PBS and then cultured in the presence of 5 mM AMP in a total volume of 100 μL of RPMI with low glucose supplemented with 10% FBS. The FBS was dialyzed previously with a membrane (molecular weight cutoff of 12 kDa). To inhibit the enzymatic activity of CD73, the tumor cells were incubated in the presence of 5 mM adenosine 5′-(α,β-methylene) diphosphate (APCP; Sigma-Aldrich, St. Louis, MO), which is a specific CD73 inhibitor. After 30 min of incubation with the inhibitor, AMP was added to a final concentration of 5 mM. To analyze the generation of Ado in the cell cultures, supernatant samples were collected at the beginning and after 5 h of incubation. Ado was detected by thin-layer chromatography (TLC) using fluorescent plates previously coated with gel (Whatman; GE Healthcare, Freiburg, Germany). For this purpose, 1-μL samples of each supernatant were loaded onto the plates and subsequently eluted for 1 h using a mobile phase composed of a mixture of isobutanol:isoamyl alcohol:ethoxyethanol:water:ammonium in a 9:6:18:9:15 ratio [41,42]. As standards, 1-μL samples of AMP, Ado, and inosine (Sigma-Aldrich) were used at a 5 mM concentration. Finally, the compounds were visualized and photographed under a UV transilluminator (UVP Biodo-H System, Upland, CA).
To quantify the amount of Ado, ultraperformance liquid chromatography equipment (Acquity UPLC; Waters, Milford, MA) was used as a synthetic Ado standard. The quantitative analysis of samples was conducted with the Empower 3 software as previously described [20].
Cytokines and antibodies to TGF-β
rh-TGF-β1 (PeproTech, Rocky Hill, NJ) was used to induce CD73 expression in CeCa tumor cells. After 96 h of culture, CD73 expression in the tumor cells was determined by flow cytometry and real-time polymerase chain reaction (RT-PCR). To neutralize the biological activity of TGF-β1 in the different cultures, a rabbit anti-human neutralizing antibody targeting TGF-β1, -β2, and -β3 was used (anti-TGF-β; R&D Systems, Inc., Minneapolis, MN); the antibody was added according to the recommendations provided by the distributor.
The TGF-β1 levels in the supernatants of the CeCa cell cultures were analyzed with the Human TGF-β1 Quantikine ELISA Kit (R&D Systems, Inc.) according to the instructions provided by the manufacturer.
Statistical analysis
All numerical data are presented as the mean value ± standard error of the mean of three independent experiments. Comparisons were evaluated by Kruskal–Wallis analysis using the statistical software version 7 (GraphPad, Software, La Jolla, CA). The differences were considered significant at P < 0.05.
Results
Coculture of CeCa cells with CeCa-MSCs increases CD73 expression and its ability to generate Ado
In cancer, MSCs play a crucial role in the promotion of tumor progression, since they provide an environment conducive to anchoring tumor cells in the tumor stroma; in addition, they can induce functional changes in tumor cells through secretion of soluble factors or exchange of membrane proteins during their interactions with tumor cells [43,44]. To analyze whether CeCa-MSCs induced changes in CD73 expression in CeCa tumor cell membranes, CeCa cells were cocultured for 96 h in the presence of different CeCa-MSC ratios using transwell chambers. To determine whether these properties were unique to CeCa-MSCs, we also used NCx-MSCs. Upon analyzing CD73 expression, we found that CaSki and MS751 cells exhibited increased CD73 expression in direct relation to the ratio of MSCs present in the coculture (Fig. 1A). CaSki and MS751 cells cocultured with CeCa-MSCs at a 1:5 ratio (CeCa:CeCa-MSCs) increased CD73 expression 2.87 ± 0.70- and 3.04 ± 0.2-fold, respectively, in relation to the basal expression of this enzyme present in the tumor cells. Interestingly, CaSki and MS751 cells cocultured with NCx-MSCs at the same ratio exhibited similar increasing in CD73 expression, 2.95 ± 0.35- and 2.8 ± 0.18-fold, respectively (Fig. 1B). When evaluating relative CD73 mRNA expression after coculture in a 1:5 ratio with NCx-MSCs and CeCa-MSCs, the CD73 mRNA levels in the CaSki cells were increased by 4.78 ± 1.69- and 4.87 ± 1.16-fold, respectively; while, those in the MS751 cells were increased by 13.2 ± 0.8- and 16.8 ± 1.85-fold, respectively (Fig. 1C).
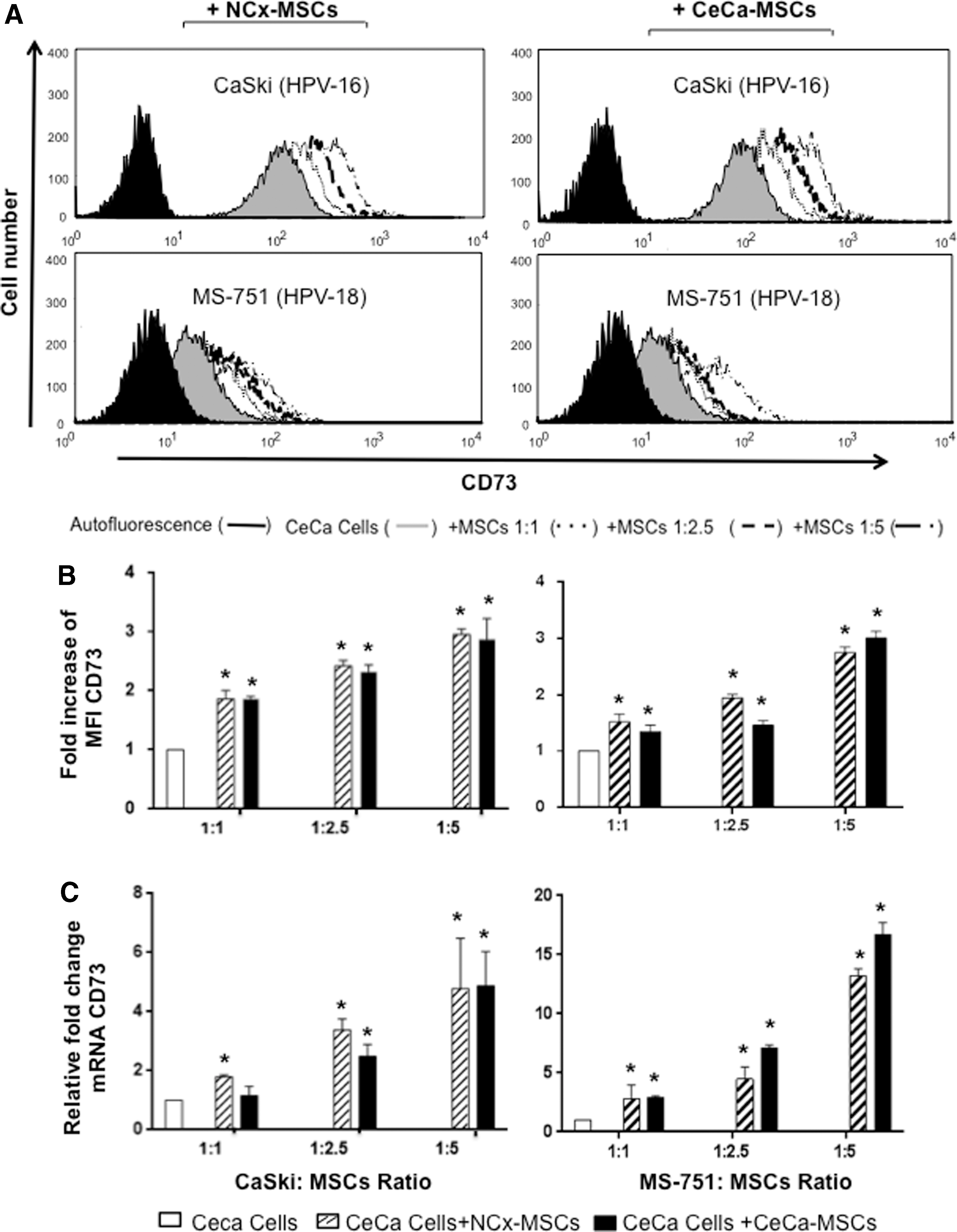
CD73 expression in CeCa cells cocultured with MSCs. CaSki and MS751 cells were cocultured with CeCa-MSCs (n = 3) or NCx-MSCs (n = 3) in 1:1, 1:2.5, and 1:5 CeCa:MSC ratios using transwell chambers. After 96 h, the CD73 expression level in the CeCa cells (25,000 events) was determined by flow cytometry
To analyze whether the increase in CD73 expression in CeCa cells cocultured with MSCs corresponded to an increase in the capacity to generate Ado, CaSki cells previously cocultured with MSCs were cultured in the presence of 5 mM AMP and the presence or absence of 5 mM APCP (a CD73-specific inhibitor). Aliquots of the supernatants were taken at the start and after 5 h of culture, and the presence of Ado was detected by TLC and quantified through UPLC. The capacity of the tumor cells to hydrolyze AMP and generate Ado increased directly with the increase in CD73 expression induced by the MSCs added to the cocultures with the CaSki cells. Apparently, the CaSki cells obtained from the coculture with CeCa-MSCs showed a greater capacity to generate Ado than those cocultured with the NCx-MSCs (Fig. 2A, B). The basal production of Ado by CaSki cells cultured in the presence of AMP was 278.11 ± 46.43 μM, whereas that of the CaSki cells cocultured previously with NCx-MSCs in 1:1, 1:2.5, and 1:5 ratios was 494.185 ± 3.34, 548.055 ± 37.62, and 729.500 ± 107.99 μg/μL, respectively, and that of CaSki cells cocultured with CeCa-MSCs in the same ratios was 515.150 ± 4.90, 681.790 ± 16.83, and 952.950 ± 26.1 μg/μL, respectively. However, the addition of the specific CD73 inhibitor APCP decreased the capacity of CaSki cells to hydrolyze AMP by more than 95% in all cases (Fig. 2B), whereas CaSki cells cultured in the absence of AMP did not produce detectable Ado levels (data not shown). These results provide important evidence of the involvement of CeCa-MSCs in the induction of increased CD73 levels in CeCa tumor cells and an increased capacity to generate Ado.

Hydrolytic activity of CD73 in CeCa cells cocultured with CeCa-MSCs. CaSki cells were cocultured for 96 h with CeCa-MSCs (n = 3) or NCx-MSCs (n = 3) in 1:1, 1:2.5, and 1:5 CeCa:MSC ratios using transwell chambers. Subsequently, the cells were separated from the coculture, washed, and cultured for 5 h in the presence of 5 mM AMP and the presence or absence of APCP, which is a specific CD73 inhibitor. The presence of Ado was detected by TLC
TGF-β1 produced in the CeCa/CeCa-MSC coculture strongly participates in the induction of CD73 expression in CeCa cells
TGF-β1 induces CD73 expression in different cell types and increases their capacity to generate extracellular Ado [45,46]. In addition, we previously demonstrated that CeCa-MSCs were capable of significantly increasing TGF-β1 expression and secretion in CeCa cells [39]. In a previous work we assayed the effect of rh-TGF-β1 on Mv1Lu cell proliferation and found that at concentrations higher than 10 ng/mL it was inhibited [40]. To corroborate this activity, in this work we used the CeCa cell lines CaSki and MS751 in the presence of different rh-TGF-β1 concentrations (5–40 ng/mL). After 96 h of culture, CaSki and MS751 cells cultured in the presence of 5, 10, 20, and 40 ng/mL of rh-TGF-β1 had increased CD73 expression in a dose-dependent manner (Fig. 3A), with increases of 1.19 ± 0.03-, 1.32 ± 0.05-, 1.86 ± 0.18-, and 2.06 ± 0.20-fold; and 1.28 ± 0.04-, 1.76 ± 0.15-, 2.86 ± 0.09-, and 3.68 ± 0.2-fold in relation to the basal CD73 expression in the CaSki and MS751 cells, respectively, which were normalized to a value of 1 (Fig. 3B, C). Similar results were observed when evaluating relative CD73 mRNA expression. Using the same rh-TGF-β1 concentrations, the increases in the CD73 mRNA levels in the CaSki cells were 1.18 ± 0.38, 1.92 ± 1.69, 3.12. ± 1.16, and 9.33 ± 3.8; and in the MS751 cells were 1.31 ± 0.28, 2.36 ± 0.79, 5.02. ± 0.62, and 8.73 ± 1.36, relative to the basal CD73 mRNA expression level in the CaSki and MS751 cells, respectively, which were normalized to a value of 1 (Fig. 3C).

TGF-β1 increased CD73 expression in CeCa cells in a dose-dependent manner. CaSki and MS751 cells were cultured for 96 h in the presence of different concentrations (5, 10, 20, and 40 ng/mL) of rh-TGF-β1. After 96 h, the CD73 expression level in the CeCa cells (25,000 events) was determined by flow cytometry
To analyze the hydrolytic activity of CD73 expressed in the CeCa cells, 1 × 106 CaSki cells previously cultured for 96 h with different rh-TGF-β1 concentrations (5–40 ng/mL) were washed and cultured for 5 h in the presence of 5 mM AMP. The amount of Ado generated by the tumor cells corresponded to the increase in CD73 induced by TGF-β1 in the cell cultures (Fig. 4). The Ado concentrations in the supernatants of CaSki cells cultured with 5, 10, 20, and 40 ng/mL of rh-TGF-β1 were 290.215 ± 9.76, 318.34 ± 15.11, 512.87 ± 8.44, and 753.20 ± 32.77 μM, respectively, whereas the Ado concentration in the cells cultured in the absence of rh-TGF-β1 was 199.00 ± 9.00 μM. Importantly, the addition of APCP completely inhibited the ability of CaSki cells to generate Ado (Fig. 4B), suggesting that TGF-β1 increased the ability of CeCa tumor cells to generate Ado through induction of CD73 expression.
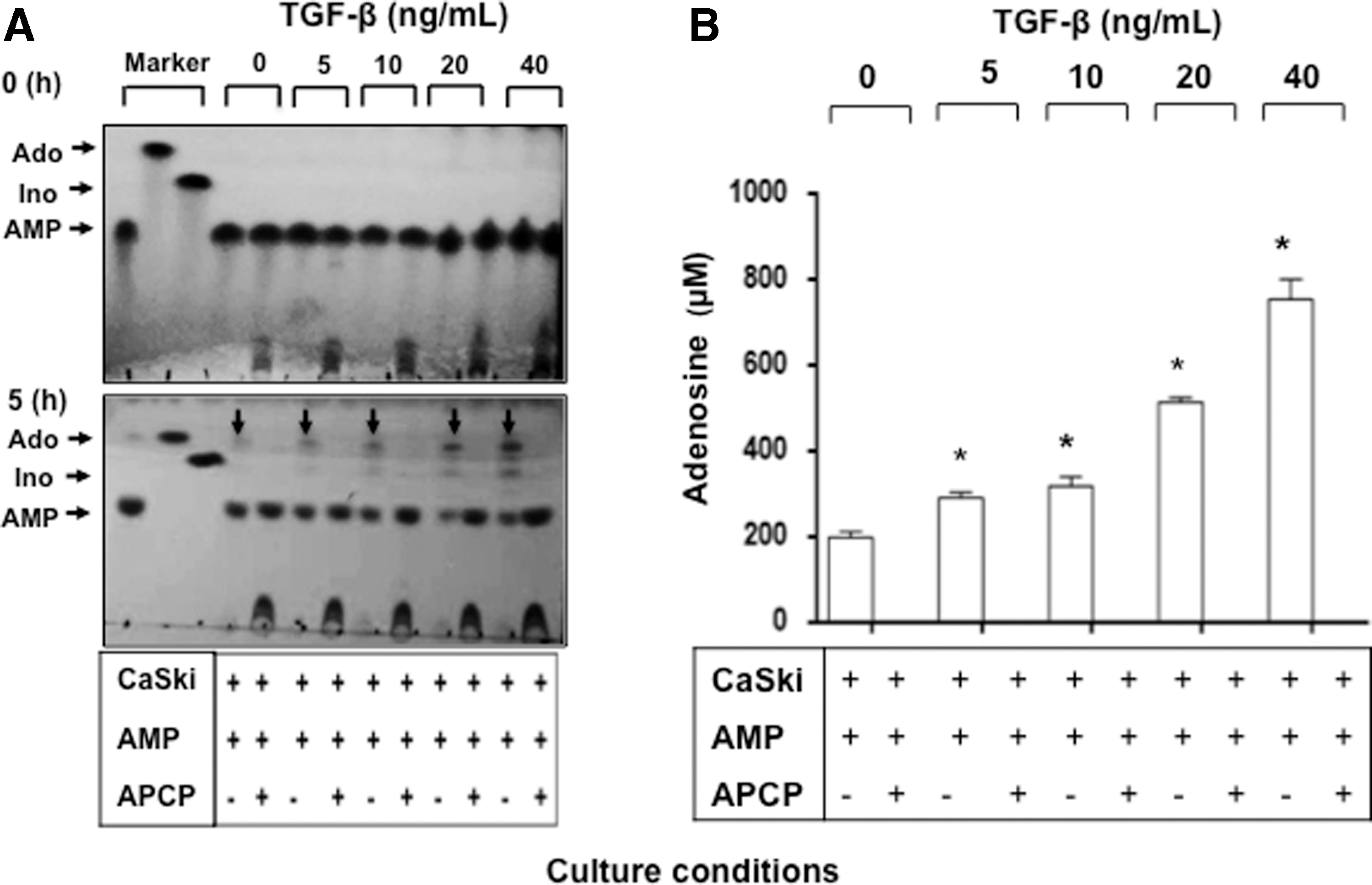
Hydrolytic activity of CD73 in CeCa cells cultured in the presence of TGF-β1. CaSki cells were cultured for 96 h in the presence of different concentrations (5, 10, 20, and 40 ng/mL) of rh-TGF-β1. Subsequently, the cells were separated from the coculture, washed, and cultured for 5 h in the presence of 5 mM AMP and the presence or absence of APCP, which is a specific CD73 inhibitor. The presence of Ado was evidenced by TLC
On the other hand, when analyzing the TGF-β1 content in the supernatants from the CaSki and MS751 cells and MSC cocultures, we observed that the concentrations of this cytokine increased in direct relation to the proportion of MSCs added to the cocultures (Fig. 5A). The TGF-β1 concentrations in the supernatants of the CeCa cells and CeCa-MSC cocultures were significantly higher (P < 0.05) than those in the cultures to which the NCx-MSCs were added. The TGF-β1 contents in the CaSki cells and CeCa-MSC cocultures in 1:1, 1:2.5, and 1:5 ratios were 560.355 ± 48.29, 828.803 ± 69.57, and 1042.09 ± 144.49 pg/mL respectively; in the presence of NCx-MSCs, the TGF-β1 contents were 396.171 ± 28.81, 482.621 ± 39.16, and 666.34 ± 59.13 respectively, whereas the TGF-β1 concentration in the supernatant of CaSki cells cultured in the absence of MSCs was 273.299 ± 10.88. While, the TGF-β1 contents in the MS751 cells and CeCa-MSC cocultures in 1:1, 1:2.5, and 1:5 ratios were 520.7 ± 40.13, 702.81 ± 38.5, and 854.83 ± 72.57 pg/mL respectively; in the presence of NCx-MSCs, the TGF-β1 contents were 256.45 ± 18.73, 496.75 ± 20.18, and 652.66 ± 49.13 pg/mL, respectively. Whereas, the TGF-β1 concentration in the supernatant of MS751 cells cultured in the absence of MSCs was 218.56 ± 21.31 pg/mL. On the other hand, the TGF-β1 mRNA expression level also increased in the tumor cells mainly when the proportion of CeCa-MSCs was increased (Fig. 5B). These results suggested that the presence of CeCa-MSCs in the TME could condition the tumor cells to produce higher TGF-β1 levels.
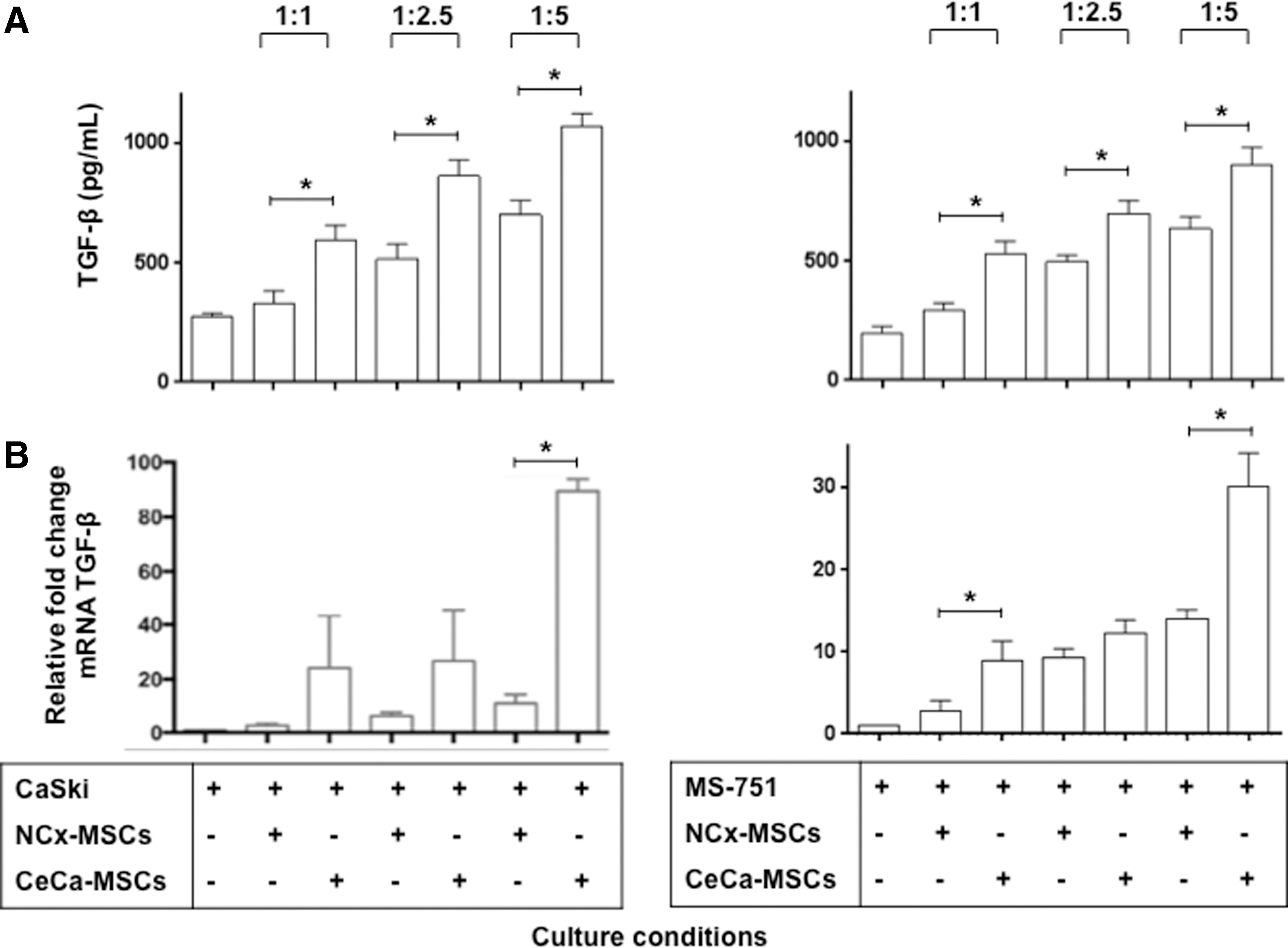
CeCa cells were induced to increase the expression of TGF-β1 in coculture with CeCa-MSCs.
To corroborate that the TGF-β1 contained in the supernatants of the CeCa cells and MSC cocultures participated in the induction of CD73 in the CeCa tumor cells, CaSki and MS751cells were cultured in a 1:5 ratio with MSCs in the presence or absence of anti-TGF-β neutralizing antibodies. Interestingly, the addition of human anti-TGF-β neutralizing antibodies to the CeCa cells and CeCa-MSC or NCx-MSC cocultures reversed CD73 expression by more than 50% in the CaSki cells and between 30% and 40% in the MS751 cells (Fig. 6A). Notably, CD73 expression in CeCa cells cultured in the presence of rh-TGF-β1 was partially reversed when anti-TGF-β neutralizing antibodies were added, which suggested that other factors produced in these cultures autocrinously by CeCa cells cultured in the presence of rh-TGF-β1 or paracrinously (generated in the coculture with MSCs) participated in the induction of CD73 in the tumor cells.

Anti-TGF-β strongly reverses CD73 expression in CeCa cells cocultured with CeCa-MSCs and diminishes their capability to generate Ado. A total of 5 × 104 CaSki and MS751 cells were cocultured with CeCa-MSCs (n = 3) or NCx-MSCs (n = 3) in 1:5 ratios using transwell chambers and in the presence or absence of 2 μg/mL of anti-TGF-β neutralizing antibodies. After 96 h, CD73 expression was analyzed by flow cytometry.
To determine whether inhibiting the induction of CD73 in CeCa cells through the addition of anti-TGF-β neutralizing antibodies reduced the capacity of the CaSki cells to hydrolyze AMP, CaSki cells previously cultured in a 1:5 ratio with MSCs in the presence or absence of anti-TGF-β neutralizing antibodies were washed and cultured for 5 h in the presence of 5 mM AMP, and in the presence or absence of APCP. Interestingly, the addition of anti-TGF-β neutralizing antibodies significantly reduced the capacity of the CaSki cells to generate Ado (Fig. 6B). The Ado concentration in the supernatant of CaSki cells cultured in the absence of MSCs was 256 ± 45.6, whereas the Ado concentrations in the supernatants of CaSki cells previously cultured in the presence of NCx-MSCs and CeCa-MSCs were 761.65 ± 107.4 and 964.48 ± 28 μM, respectively; and those when anti-TGF-β neutralizing antibodies were added, were 505.10 ± 8.4 and 542.18 ± 28.5 μM, respectively (Fig. 6C). Importantly, the addition of APCP strongly inhibited the ability of CaSki cells to generate Ado (Figs. 6B, C).
These results suggest that during the intercommunication of tumor cells with MSCs in the TME, TGF-β1 production is an important factor in maintaining an immunosuppressive status not only for the activity of this cytokine but also for its capacity to induce CD73 expression in the tumor cells and generate an immunosuppressive microenvironment rich in Ado.
Discussion
MSCs are a major component of the TME and play a key role in the promotion of tumor progression through differentiation into carcinoma-associated fibroblasts and/or tumor-associated fibroblasts, pericytes, and prefibroblasts by providing tumor cells with a survival advantage either by promoting their proliferation [47,48] or creating an immunosuppressive microenvironment [49]. They also provide support and a framework for anchoring tumor cells in the form of a tumor stroma [44]. However, the precise mechanisms by which MSCs exert their effector functions have not been fully elucidated. In this context, several studies have shown that MSCs exert paracrine activity on neighboring cells by secreting various bioactive factors that can radically change their cellular functions, including survival, apoptosis, maturation, and differentiation [50,51]. For example, MSCs promote the EMT in tumor cells and can promote metastatic colonization through TGF-β1 production in the TME [52,53]. Similarly, TGF-β1 has been reported to contribute to the creation of an immunosuppressive TME rich in Ado by inducing CD73 expression in Tregs and MDSCs [54,55]. Based on these findings and our recent report that CeCa-MSCs were able to significantly increase TGF-β1 expression and secretion in CeCa tumor cells [39], in this study, we analyzed the capacity of CeCa-MSCs to modify CD73 expression in CeCa tumor cells and therefore their ability to generate Ado. By coculturing CeCa tumor cells with CeCa-MSCs, we found that the tumor cells increased CD73 expression and the ability to generate Ado in a manner that was dependent on the proportion of CeCa-MSCs added to the cocultures. This phenomenon was not only exclusive of the presence of CeCa-MSCs in the cocultures but also occurred when the CeCa cells were cocultured with NCx-MSCs. Taking into consideration that TGF-β1 is a cytokine that induces CD39 and CD73 expression in immunoregulatory cells, such as Tregs [56,57], dendritic cells [58], and myeloid suppressor cells in mouse tumors [46] and is even able to induce the expression of these ectonucleotidases in cells with the Th17 phenotype [58], we proceeded to analyze whether the ability of CeCa-MSCs to induce increased CD73 expression in CeCa tumor cells functioned through TGF-β1 production. We initially confirmed TGF-β1 functions to induce CD73 expression in CeCa tumor cells through CeCa cell culture in the presence of rh-TGF-β1. Furthermore, the increase in CD73 in the CeCa cell membrane was significantly related to the TGF-β1 content in the cell coculture supernatants; a higher TGF-β1 concentration was detected in cocultures with a higher proportion of MSCs, and increased TGF-β1 mRNA expression also detected in tumor cells. The addition of anti-hTGF-β neutralizing antibodies strongly reversed the CD73 level in the tumor cells (by more than 50%). However, the fact that a higher TGF-β1 concentration was detected in CeCa/CeCa-MSCs than in CeCa/Ncx-MSCs cocultures suggests that at least in NCx-MSCs other factors produced under these culture conditions could participate in the induction of CD73. It will be interesting to evaluate this possibility in future studies.
Recent studies have shown that MSCs can induce functional changes, such as an increase in CD73 activity in tumor cells and NK cells, either through the exchange of biological material through extracellular vesicles, including exosomes, or through paracrine activity [43,59]. In the present study, we provide evidence that the TGF-β1 contained in the supernatant of cocultures of CeCa tumor cells with MSCs induces increased CD73 expression in the tumor cells. This property has been observed in splenic T cells, BM-derived dendritic cells, and peritoneal macrophages, upon in vitro stimulation with TGF-β1 [45]. In fact, Fausther et al. already showed that SMAD proteins may control CD73 transcription [60]. Then, the MSCs that reside in the TME and constitute 0.1%–1% of the total number of cells in solid tumor tissues [8] can be determining factors in the induction of functional changes in the tumor cells or exacerbate their immunosuppressive capacity through the adenosinergic pathway. In the particular case of CeCa, several studies have shown that the presence of TGF-β1 in the sera and tumor tissues of patients correlates directly with the degree of disease progression [32,33,61,62]. Therefore, because our results show that communication between CeCa cells and MSCs generates high amounts of TGF-β1 and that induction of CD73 in CeCa cells depends heavily on the presence of TGF-β1, suggest that the presence of MSCs into the TME promote CeCa cells to express high levels of CD73 through the secretion of TGF-β1 and contribute to suppression of the immune response through the production of large amounts of Ado, implying that this route may have clinical importance as a therapeutic target.
Conclusions
In this study, we found that CeCa tumor cells cocultured with CeCa-MSCs or NCx-MSCs were induced to significantly increase CD73 expression by the production of TGF-β1, since addition of anti-hTGF-β neutralizing antibodies strongly reversed CD73 expression in the tumor cells. These results suggest that the interaction of MSCs with CeCa tumor cells in the TME can trigger the production of TGF-β1 to maintain an immunosuppressive status not only through the activity of this cytokine per se, but also through its ability to induce CD73 expression in the tumor cells and generate an immunosuppressive microenvironment rich in Ado.
Footnotes
Acknowledgments
The authors are indebted for CONACYT support to A.M.-G. (grant no. 240635) and DGAPA-PAPIIT support to M.L.M.G. (grant no. IN226516); and Mexican Institute of Social Security, IMSS, support to A.M.-G. (grant nos. 1161, 1314, 1383, and 1613) are gratefully acknowledged. They also thank the support provided to LRAI and DBTP by the Postdoctoral Scholarship Program of the UNAM. Finally, the authors thank the “Laboratorio Nacional para Servicios Especializados de Investigación, Desarrollo e Innovación (I+D+i) para Farmoquímicos y Biotecnológicos (LANSEIDI-FarBiotec-CONACyT)” for facilitating the use of the UPLC system.
Author Disclosure Statement
No competing financial interests exist.