
Abstract
Cells sense and respond to environmental changes induced by gravity. Although reactions to conventional culture have been intensively studied, little is known about the cellular reaction to simulated microgravity conditions. Thus, in this study, we investigated the effects of simulated microgravity on human mesenchymal stem cells using a three-dimensional clinostat (Gravite®), a recently developed device used to generate simulated microgravity condition in vitro. Our time-lapse analysis shows that cells cultured under conventional culture conditions have a stretched morphology and undergo unidirectional migration, whereas cells cultured under simulated microgravity conditions undergo multidirectional migration with directional changes of cell movement. Furthermore, cells cultured under conventional culture conditions maintained their spindle shape through fibronectin fibril formation in their bodies and focal adhesion stabilization with enriched stress fibers. However, cells cultured under simulated microgravity conditions were partially contracted and the fibril structures were degraded in the cell bodies. Additionally, paxillin phosphorylation in the cells cultured under simulated microgravity conditions was more intense at the cell periphery in regions near the leading and trailing edges, but was less expressed in the cell bodies compared with that observed in cells cultured under conventional culture conditions. Furthermore, lamin A/C, a major component of the nuclear lamina, was mainly located on the apical side in cells cultured under conventional culture conditions, indicating basal-to-apical polarization. However, cells cultured under simulated microgravity conditions showed lamin A/C localization on both the apical and basal sides. Taken together, these results demonstrate that simulated microgravity-driven fibronectin assembly affects nuclear lamina organization through the spatial reorganization of the cytoskeleton.
Introduction
Alteration of the physical environment surrounding cells can indirectly lead to changes within the cell [1,2]. In fact, cell behavior is notably influenced by the geometrical space occupied by the cell and is often altered by its external boundaries [3,4]. This is the result of changes in the balance of the mechanical forces exerted on the cell membrane by intracellular components and the outside environment [5,6]. The intrinsic forces exerted on the membrane are mostly the direct result of cytoskeleton reorganization [7] and mechanotransduction through integrin and focal adhesion-mediated connections between the nucleus and extracellular matrix (ECM) [8]. The nucleus reacts directly to changes in both gene regulation and nuclear mechanical properties that are independent of the cytoplasmic biochemical and cytoskeletal responses [9], and the physical connection between the nucleus and the cytoskeleton is important for many cellular processes involving nuclear lamina organization changes [10]. However, the mechanism of this cellular mechanoadaptation in various environments has yet to be fully characterized.
Changes in the physical environment, such as the space and dynamic surroundings, influence cell sensing through indirect mechanisms, including loss of gravity-dependent convection, lack of sedimentation, and buoyancy force disruption [11,12]. In the literature, reaction diffusion systems have been extensively used as fundamental models of spatiotemporal dynamics of various biochemical reactions, including enzyme kinetics and the assembly of complex protein networks [13]. Notably, it has been reported that gravity affects these reaction diffusion systems in both theoretical calculations [14] as well as in practical experiments, proving that gravity influences traveling velocity based on the convection of target patterns relating to their interactions [15,16].
As spaceflight experiments are limited, several ground-based facilities for microgravity environment simulation have been developed, such as rotating wall vessel bioreactors, random positioning machines, and diamagnetic levitation systems [17]. For cultured cells, a three-dimensional (3D) clinostat is a common device used to provide a simulated microgravity environment, and it has been shown to be an effective method for attenuating gravitational effects similar to those observed during spaceflight experiments [18]. When under simulated microgravity conditions established with a 3D clinostat (Gravite®), the cells experience an average gravity vector near zero during a constant rotational speed over 360° in two axes. Notably, this rotation generates functional simulated microgravity effects similar to those observed in a true microgravity environment when the rotation speed is faster than the cellular response time [19]. Although the advent and improvement of these techniques has/have allowed the effects of simulated microgravity to be studied in vitro, the 3D clinostat system has been rarely applied to evaluate a number of cell types, including human mesenchymal stem cells (hMSCs).
In this study, we investigated the effect of simulated microgravity on the cytoskeleton and nucleoskeleton in hMSCs by evaluating the effects on fibronectin assembly. Based on the observed differences in hMSC behavior when under simulated microgravity compared with those under conventional culture conditions, we also evaluated the fundamental gravity-related mechanisms underlying the interactions between the nuclear lamina and the cytoskeleton in hMSCs.
Materials and Methods
Cells and culture conditions
Cryopreserved bone marrow-derived hMSCs (Lot no. 0000374385; Lonza Walkersville, Inc., Walkersville, MD) were thawed according to the manufacturer's instructions. Routine subcultures of hMSCs were cultured using hMSC growth medium (Lonza) at 37°C in a humidified atmosphere containing 5% CO2. At 70% confluency, the cells were detached by enzymatic treatment with a 0.1% trypsin/0.02% ethylenediaminetetraacetic acid solution (Sigma-Aldrich, St. Louis, MO). Cells at their fifth passage were used in these experiments.
For all experiments, hMSCs were expanded for the specified number of days in Dulbecco's modified Eagle's medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum and antibiotics (Life Technologies, Carlsbad, CA) at a depth of 4 mm in the culture vessels. The seeding density was 5.0 × 103 cells/cm2, and the number of viable cells was estimated by a Trypan Blue exclusion test through the direct counting of suspended cells with a hemocytometer.
Cell culture under simulated microgravity conditions
The Gravite, a simulated microgravity generator, was produced by Space Bio-Laboratories Co., Ltd. (Hiroshima, Japan). Using a simultaneous rotation on two axes with constant angular velocity, the Gravite cancels the cumulative gravity vector at the center around the device, producing a simulated microgravity environment of 10−3 G in 8 min after the operation estimated by an acceleration sensor in the Gravite. This is accomplished by rotating a chamber at the center of the device to disperse the gravity vector uniformly within a spherical volume.
On day 0, hMSCs were seeded into a 12.5-cm2 T-flask with vented filter cap (Corning, Tewksbury, MA). Then, on day 1, the medium was removed from that flask. Subsequently, the flask was completely filled with fresh medium without any bubbles. After that, the cells were cultured in a simulated microgravity environment at the center of the Gravite until day 3 without any medium change. For the time-lapse observations, a phase-contrast microscope with a 10 × objective lens has been installed into the Gravite. After locking position of the culture flask above the lens, the machine started rotation and the images were automatically obtained every 10 min during this machine's operation continuously. For control condition, on day 1 after fully filling the T-flask with fresh medium without any bubbles, the cells were cultured under conventional culture with static condition.
Immunofluorescent staining
Immunostaining was carried out as described previously [20]. Briefly, cells were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS; Wako Pure Chemical Industries, Tokyo, Japan) for 10 min at room temperature. Then, the cells were incubated in PBS with 0.5% polyoxyethylene octylphenyl ether for 10 min. After masking the nonspecific proteins with Block Ace (Dainippon Sumitomo Pharma Co., Ltd., Osaka, Japan) at 4°C overnight, the cells were incubated with primary antibodies at 4°C overnight. In this study, we used primary antibodies against paxillin (Millipore, Billerica, MA), phosphorylated paxillin (Cell Signaling Technology, Inc., Danvers, MA), fibronectin (Santa Cruz Biotechnology, Dallas, TX), lamin A/C (Santa Cruz), and lamin B (Abcam, Cambridge, United Kingdom). All of the antibodies were adequately diluted in PBS containing 10% Block Ace. After staining, the cells were washed with Tris-buffered saline (Dako, Glostrup, Denmark) and immunolabeled with the appropriate secondary antibodies [Alexa Flour 488-conjugated anti-mouse, anti-rabbit, or anti-goat immunoglobulin G (IgG) or Alexa Flour 594-conjugated anti-mouse IgG; both from Life Technologies] for 1 h at room temperature. The nuclei and F-actin were stained with 4′,6-diamidino-2-phenylindole and fluorescent rhodamine phalloidin (Life Technologies), respectively. Images were obtained using a confocal laser scanning microscope (FV-1000; Olympus, Tokyo, Japan) through a 60 × or 100 × objective lens.
Quantitative real-time polymerase chain reaction
RNA isolation and quantitative real-time polymerase chain reaction (RT-PCR) assays were carried out as described previously [21]. Total RNA was isolated from the cells using an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to a commercially available protocol. The RNA was then reverse transcribed using a PrimeScript® RT Reagent Kit (TaKaRa Bio, Inc., Shiga, Japan). The quantitative RT-PCR assays were conducted using SYBR® Premix Ex Taq (TaKaRa Bio, Inc.) and carried out on a real-time PCR system (Applied Biosystems 7300; Thermo Fisher Scientific, Waltham, MA). We utilized the following specific primers according to previous reports as shown in Table 1 [22,23]. Relative mRNA expression of the target gene was evaluated by the cycle threshold value (Ct) and normalized to that of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) using the 2−ΔΔCt method.
Primer Sequences Used in This Study
MT1-MMP, membrane type-1 matrix metalloproteinase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Protein extraction and western blot analysis
Protein extraction and western blot analysis were performed as described previously [21]. Briefly, total protein was extracted from cells using RIPA lysis buffer (Sigma-Aldrich) and a Halt™ protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific) on ice for 30 min. Protein concentration was estimated using a BCA Protein Assay Kit (Thermo Fisher Scientific). Equal amounts of total lysates were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes using standard methods. The membrane was blocked with ECL™ Blocking Agent (GE Healthcare, Chicago, IL) for an hour at room temperature and then incubated with primary antibodies against paxillin (Millipore), phosphorylated paxillin (Cell Signaling Technology, Inc.), and GAPDH (Sigma-Aldrich) at 4°C overnight. Later, the membrane was incubated with secondary antibodies [DyLight® 800 anti-Rabbit IgG (Thermo Fisher Scientific), Starbright™ blue 700 anti-mouse IgG (Bio-Rad)] for an hour at room temperature. Fluorescent signals were displayed with a ChemiDoc™ MP Imaging system (Bio-Rad). Western blots were performed for three individual experiments. The optical intensity of protein signals was quantified using an image processing and analysis software (Image Lab™; Bio-Rad).
Statistical analysis
All quantitative data were obtained from triplicate experiments, which are expressed as the mean ± standard deviation. Comparisons between the groups were evaluated by one-way analysis of variance and the Tukey's honest significance test. P-values < 0.01 were considered statistically significant.
Results
Changes in hMSC behavior under simulated microgravity conditions
To investigate the effects of simulated microgravity on hMSCs, cells were first grown under conventional culture conditions for 1 day. Then, a Gravite was used to generate simulated microgravity conditions in the culture environment, and the changes in cell behavior were monitored. Notably, the time-lapse images show that the cells present extensions and dynamic protrusions under both culture conditions (Supplementary Movies S1 and S2). Under conventional culture conditions, the spindle-shaped cells underwent unidirectional migration through extension of the leading edge followed by contraction of the trailing edge. In contrast, cells under simulated microgravity conditions showed multidirectional migration with active extension of leading edge together with partial contraction of cell rear (Supplementary Fig. S1A). Moreover, cells under simulated microgravity conditions exhibited higher frequency of directional changes of migration than those under conventional culture conditions (Fig. S1B).
Cytoskeletal organization and focal adhesion formation under simulated microgravity conditions
To clarify cytoskeletal organization, hMSCs grown under conventional and simulated microgravity conditions were stained with F-actin on days 1 and 3. The cells under conventional culture conditions on day 1 (Fig. 1A and Supplementary Movie S3) and day 3 (Fig. 1B and Supplementary Movie S4) had longitudinal actin stress fibers along both the apical and basal sides. Alternatively, the cells grown under simulated microgravity conditions presented larger cortical actin fibers in the filopodia and lamellipodia regions and had less stress fibers located mainly along the basal side (Fig. 1C and Supplementary Movie S5). Moreover, paxillin, a focal adhesion protein, was also analyzed at these time points in cells under both conditions. Our results show that paxillin is present in the peripheral and central regions of cells in both culture conditions, with intensive staining being observed in the longitudinal direction of the stress fibers in a scattered manner. Furthermore, cells under simulated microgravity conditions presented greater paxillin aggregation, particularly at the leading edges of the cells (Fig. 1C). In contrast, cells under conventional culture conditions exhibited more dispersed paxillin at both the cell edges and bodies (Fig. 1A, B).
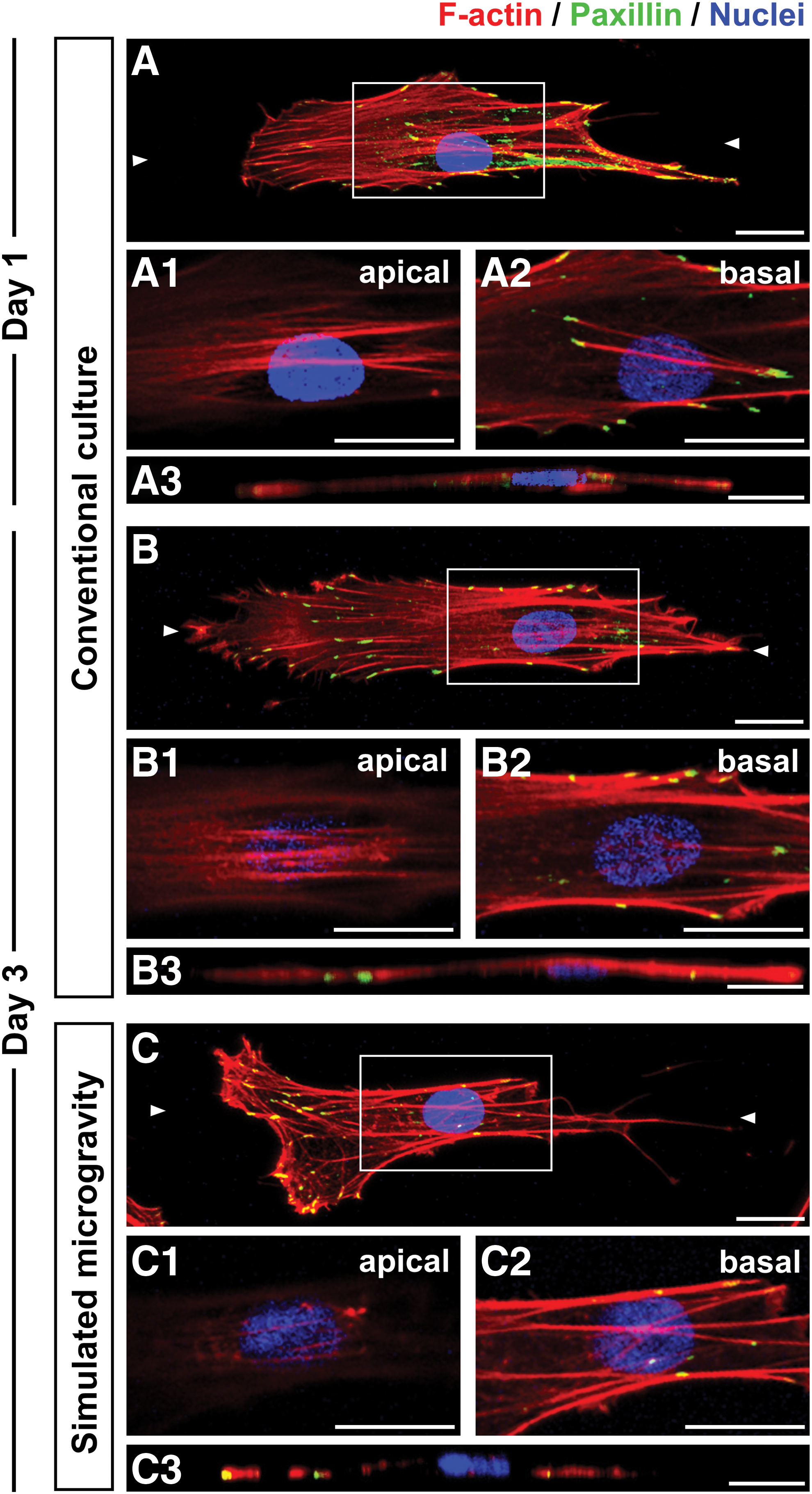
Actin cytoskeleton structures and focal adhesion construction in hMSCs. Representative images showing F-actin (red) and paxillin (green) expression, as well as the nuclei (blue) in cells on day 1
To examine focal adhesion protein dynamics, hMSCs on day 3 were investigated by immunostaining and western blot analysis with both paxillin and phosphorylated paxillin. In cells grown under conventional culture conditions, phosphorylated paxillin was largely colocalized with paxillin in the central and peripheral areas (Fig. 2A). However, cells grown under simulated microgravity conditions presented phosphorylated paxillin, more intensive formation at the cell edges (Fig. 2B1), compared with those grown under conventional culture conditions (Fig. 2A1). Although total paxillin against GAPDH was the same level between two culture conditions, the average intensity ratio of phosphorylated paxillin per total paxillin in cells under simulated microgravity conditions was significantly higher than those under conventional culture conditions (Fig. 2C, D), indicating enhancement of paxillin phosphorylation in cells under simulated microgravity conditions.

Focal adhesion protein phosphorylation in hMSCs. Representative images showing paxillin (red) and phosphorylated-paxillin (green) expression as well as the nuclei (blue) on day 3 in cells cultured under conventional culture conditions
Fibronectin assembly and matrix-degrading enzyme activity under simulated microgravity conditions
Cytoskeletal formation and fibronectin assembly were observed by immunostaining the cells grown under both conditions for F-actin and fibronectin on days 1 and 3. In both types of gravity-treated cells, concentrated fibronectin was detected at the interface between the cells and culture plate surface. On day 1, fibronectin with fibril structures were not found at the cell periphery in regions near the leading and trailing edges (Fig. 3A1), but were detected in the longitudinal direction under cell bodies together with stress fibers in the cells (Fig. 3A2). On day 3, the cells grown under conventional culture conditions had fibronectin fibrillogenesis, showing as an increase of fibronectin fibrils under the cell bodies (Fig. 3B2) than the cells on day 1 (Fig. 3A2). However, cells cultured under simulated microgravity conditions on day 3, we observed fibronectin under the cell bodies, but fewer fibril structures (Fig. 3C2), in addition to there being more aggregated structures than we noted for the cells grown under conventional culture conditions (Fig. 3B2). Furthermore, cells under simulated microgravity conditions had significantly lower mRNA expression of fibronectin than the cells under conventional culture conditions (Supplementary Fig. S2).
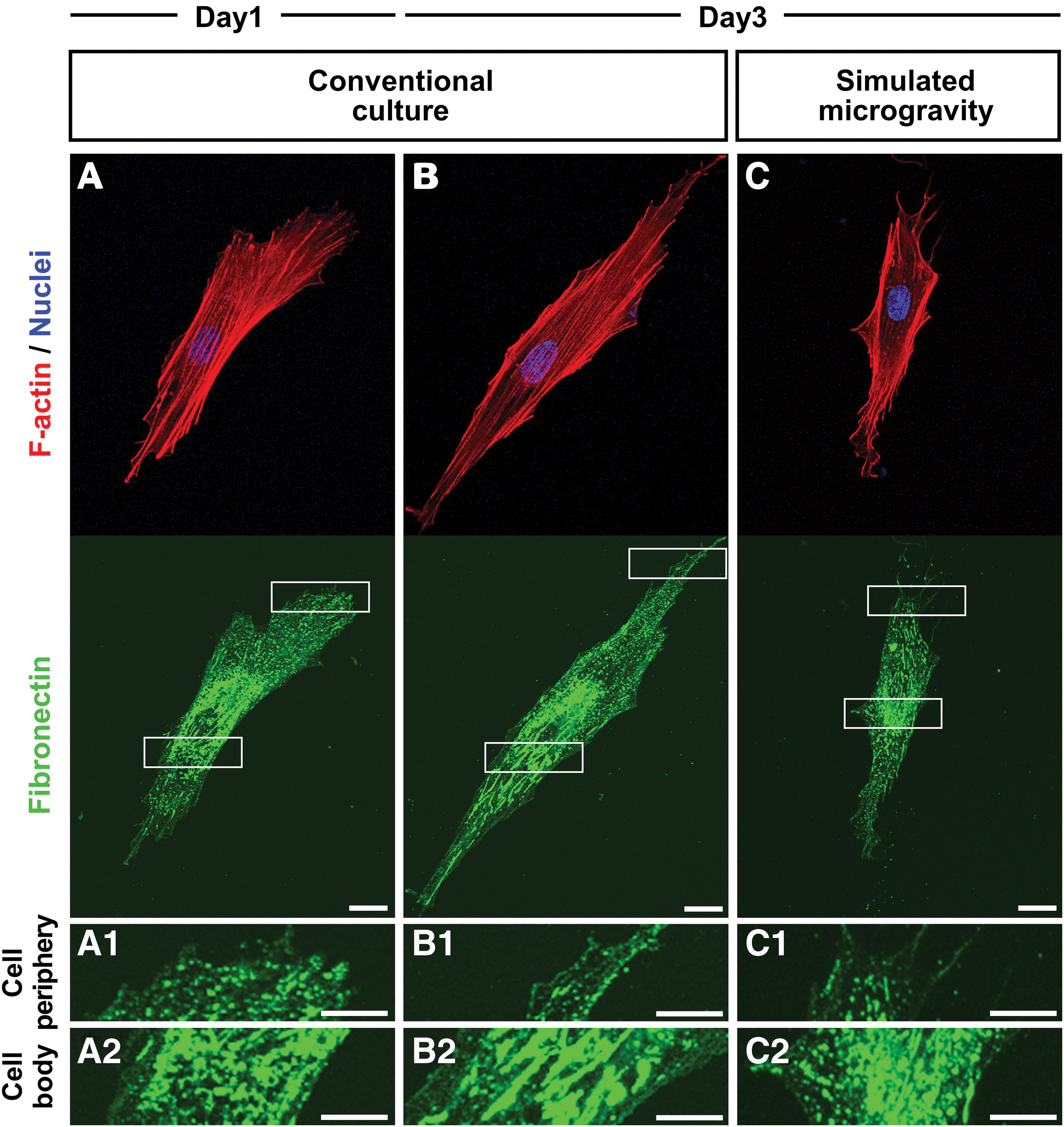
Fibronectin assembly and the actin cytoskeleton in hMSCs. Representative images showing F-actin (red) and fibronectin (green) expression as well as the nuclei (blue) on day 1
Subsequently, the expression of membrane type-1 matrix metalloproteinase (MT1-MMP or MMP14), a surface enzyme responsible for fibronectin degradation, was evaluated by quantitative RT-PCR in cells grown under both gravity conditions on days 1 and 3. As shown in Fig. 4, the relative mRNA expression of MT1-MMP in cells grown under conventional culture conditions was significantly lower on day 3 than it was on day 1. However, cells grown under simulated microgravity conditions exhibited a significantly greater MT1-MMP expression on day 3 than that observed on day 1. Furthermore, MT1-MMP expression in cells grown under simulated microgravity conditions was approximately three-fold higher than that of cell grown under conventional culture conditions on day 3.
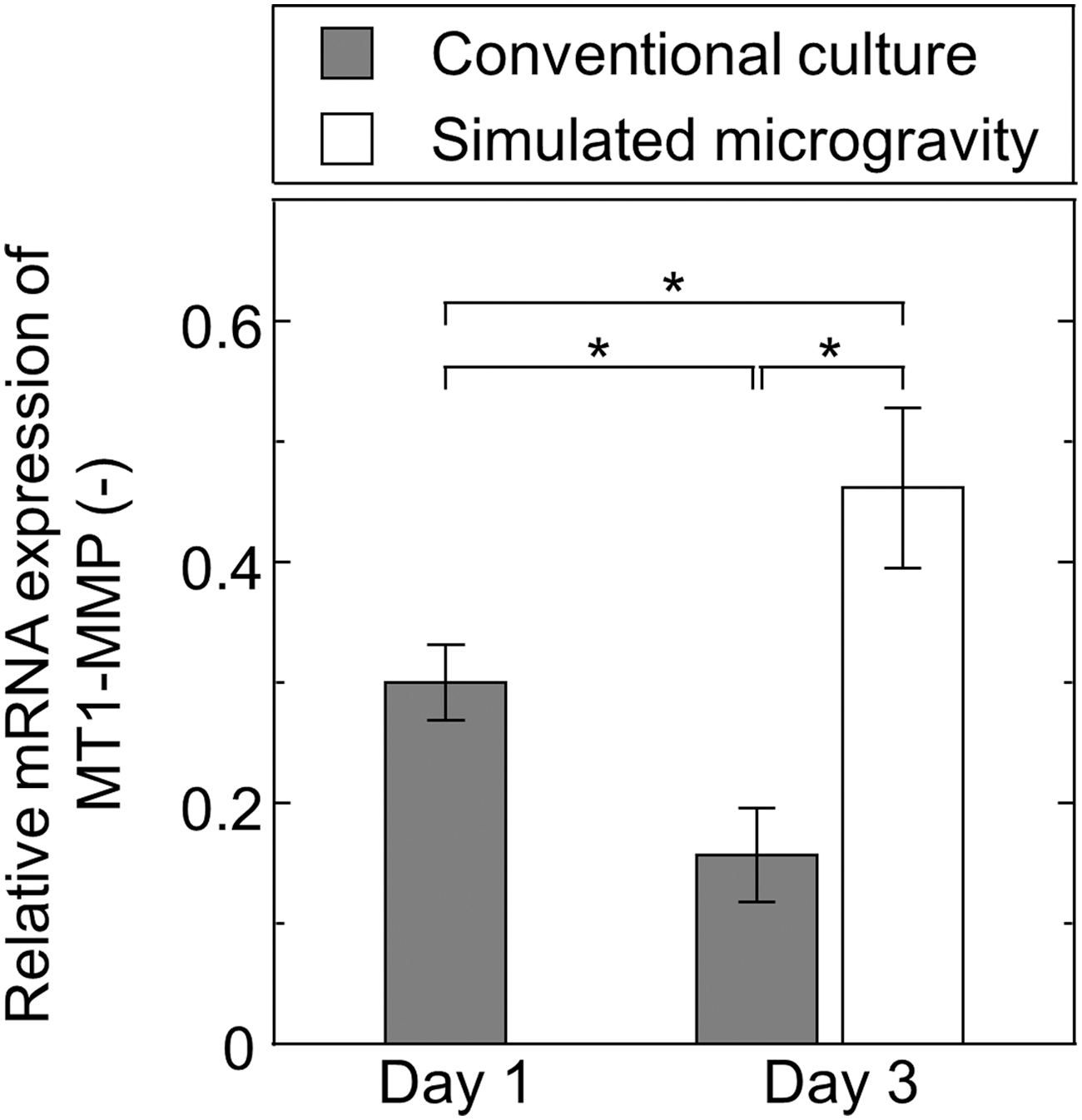
Quantitative real-time polymerase chain reaction analysis of MT1-MMP expression in hMSCs cultured under conventional culture conditions (shaded bars) on day 1 and 3, as well as simulated microgravity conditions (open bar) on day 3. Data represent the mean ± standard deviation (n = 3). *P < 0.01. MT1-MMP, membrane type-1 matrix metalloproteinase.
Spatial organization of nuclear lamina under simulated microgravity conditions
We next investigated the formation of nuclear lamins (types A/C and B), which are major structural components of the peripheral nuclear lamina, in the nuclei of cells cultured for 3 days under both gravitational conditions using immunostaining. Our results show that lamin A/C and lamin B were detected mainly at the periphery of the nucleus in all culture conditions (Fig. 5A1a–B3a and Supplementary Fig. S3). However, cells cultured under simulated microgravity showed more concentrate of lamin B at the periphery than those in conventional culture. Nevertheless, cells grown under conventional culture conditions presented lamin A/C mainly on the apical side, indicating basal-to-apical polarization of this lamin (Fig. 5A1a). Notably, lamin A/C was localized on both the apical and basal sides of cells grown under simulated microgravity conditions (Fig. 5B1a), suggesting that this cellular polarization has less occurrence under these conditions.

Structural organization of nuclear lamins in hMSCs. Representative images showing lamin A/C
Discussion
Changes in the external environment are widely known to affect cell behavior; however, the mechanism underlying this cellular mechanoadaptation has been largely unstudied in environments with altered gravitational conditions. Cells under conventional culture conditions interact with adsorbed surface proteins through integrins that stabilize adhesion of the cell body [24]. These integrin–surface protein interactions may be governed by the conformation and adsorption of ECM proteins, such as fibronectin [25]. Thus, variations in ECM structure or mechanics will therefore alter the ECM's ability to resist cell traction forces exerted on integrins [26]. The interactions between the cells and the surface result in cell migration that occurs through ordered changes in the cytoskeleton and the regulated formation, turnover, and distribution of focal adhesions [27]. Paxillin phosphorylation leads to the formation of active lamellipodial protrusions [28] and is often used to indicate the active turnover of focal adhesions [29]. Notably, focal adhesion turnover and distribution is regulated by various factors, including MT1-MMP [30]. It has also been reported that ECM degradation is mediated by MMPs [31]. Indeed, MT1-MMP has been shown to play a role in fibrillar fibronectin degradation [32]. Furthermore, the cellular mechanical response through ECM–integrin–cytoskeleton–nucleus axis is one of the most important mechanotransduction systems in these cells [33].
Nuclear lamins are also known to interact with membrane-associated proteins to form nuclear lamina at the nuclear envelope [34]. The cytoskeleton, which is linked to the nuclear lamina and thereby to chromatin, has recently been proposed to impact chromatin remodeling and transcriptional activity [35]. This physical connection provides a structural basis for mechanical signal transduction between integrins and the nucleus through the actin cytoskeleton [36]. Recent studies have also provided critical information highlighting the significant contribution of lamins to nuclear mechanics and the spatial organization [37 –39]. Lamin A/C, for example, contributes to nuclear stiffness [40,41] and is involved in actin cap function [37], which is an essential aspect of biophysical signaling between the ECM and nucleus [42]. Notably, Ihalainen et al. [43] have also reported that basal-to-apical polarization of lamin A/C is mediated by nuclear compression. Their data indicate that the differential basal-to-apical accessibility of lamin A/C epitopes in the nuclear lamina are regulated by changes in cytoskeletal tension in response to environmental changes.
In the present study, we investigated the nuclear lamina and cytoskeleton in hMSCs cultured under simulated microgravity conditions. Figure 6 shows a diagram outlining the effects of simulated microgravity on fibronectin assembly and the related formation of focal adhesions and actin cytoskeleton network in cell–surface interfaces. The recent development of the Gravite system, which allows cells to be cultured under simulated microgravity, has enabled the study of this particular change in gravity on cell behavior compared with those cultured under conventional culture conditions. Cells grown under conventional culture conditions had a stretched morphology and unidirectional migration (Supplementary Movie S1) and maintained their spindle shape through fibronectin fibril formation in their bodies and focal adhesion stabilization with enriched stress fibers (Figs. 1 and 3). However, cells grown under simulated microgravity conditions migrated multidirectionally through cell movement with active extension of leading edge (Supplementary Movie S2) and were partially contracted and the fibril structures were degraded in the trailing edge (Figs. 1 and 3). These results were supported by immunostaining of paxillin and phosphorylated paxillin, which indicates a higher level of the phosphorylated form at the edges of the cells cultured under simulated microgravity conditions. This suggests that cells grown under simulated microgravity have an active extension of their leading edges through the inhibition of fibronectin fibrillogenesis by increasing of MT1-MMP, which in turn causes the release of adhesions and immature stress fiber formation that affect directional changes in cell migration, similar to previous findings of hMSCs grown on dendrimer-immobilized surfaces under conventional culture [23]. In addition, another similar phenomenon was also reported by Guignandon et al. [44] who have reported in a study evaluating the effects of a microgravity environment on osteoblastic cells. Moreover, the promotion of phosphorylated paxillin-mediated focal adhesion turnover by active cell migration has also been documented [45]. Thus, based on our results and those of others in similar or related studies, simulated microgravity-driven fibronectin assembly by higher expression of MT1-MMP, show that ECM appears to be responsible for the formation of focal adhesions as well as the actin cytoskeleton during cell–surface interactions under this particular gravitational condition. The focal adhesions subsequently initiate intracellular signaling cascades that determine cell behavior [46]. To our knowledge, this is the first time the effects of simulated microgravity have been evaluated with regard to the behavior of hMSCs. Thus, studying the mechanical forces influencing the assembly, stabilization, and dissociation of focal adhesions and ultimately cell behavior in response to the mechanical environment provides significant insight into how cells function.
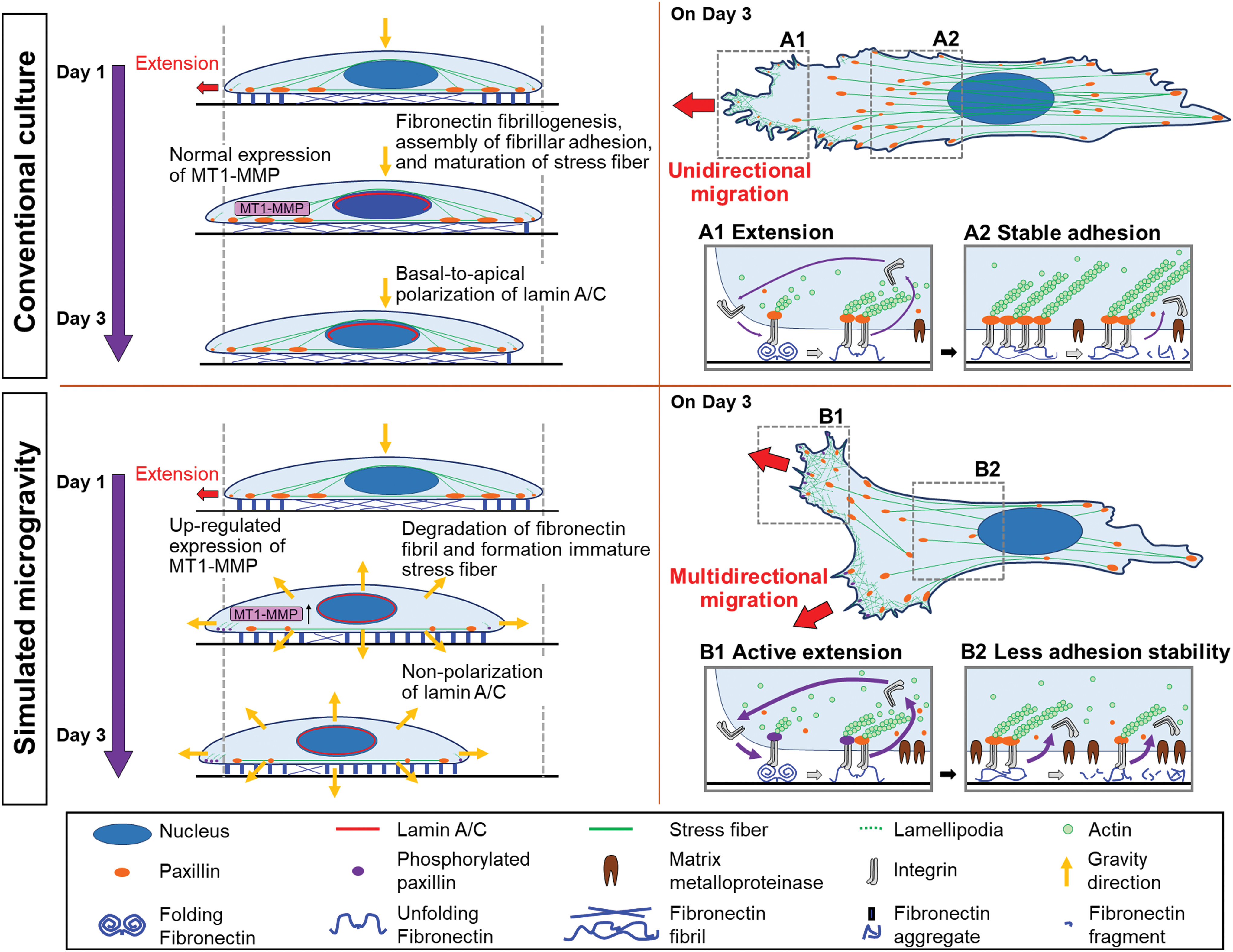
Schematic illustration showing the processes of fibronectin assembly, focal adhesion formation, and actin cytoskeleton maintenance and the influence of organization changes in the nuclear lamina in hMSCs suggested by our data. When cultured under conventional culture conditions, the cells undergo unidirectional migration with extension at the cell edge
In the present study, we also describe changes in the nuclear lamina–cytoskeleton interaction in hMSCs that appears to be mediated by the simulated microgravity-driven fibronectin assembly. We observed basal-to-apical polarization of lamin A/C in the hMSCs grown under conventional culture conditions, but this polarization was much lower when the cells were grown under simulated microgravity conditions (Fig. 5). Based on these previous studies [42,43], it is possible that environmental changes, including gravitational changes, might induce alternations in the nuclear lamina–cytoskeleton interactions through disruption of ECM–actin cytoskeleton signaling. Because the cell's actin cytoskeleton network dynamics induced by actomyosin interaction can exert active, contractile forces on the ECM are transduced through adhesion complexes through simulated microgravity-driven fibronectin assembly. During the response and adaptation of hMSCs cultured under simulated microgravity conditions, it is likely that multiple mechanosensitive systems, molecules, and signaling pathways are involved. These results indicate that simulated microgravity induces mechanotransduction changes in hMSCs through the loss of lamin A/C polarization and disruption of the apical stress fibers. These cytoskeletal and nucleoskeletal changes in hMSCs observed under simulated microgravity condition seemed to be similar to the concept of “force isotropy” relevant to cytoskeletal forces and relevant to extracellular loads acting on cells, reported by Nava et al. [47]. According to their work, isotropic cytoskeletal tension is characterized by adhesion-mediated traction forces of similar magnitude at varying orientation, resulting in a roundish nuclear morphology. It is most likely that the simulated microgravity induces isotropic properties which exhibits an isotropic distribution both in position and momentum. In future work, further examination will be conducted to evidence the cytoskeletal tension and lamin A/C nucleoskeletal changes by isotropic-inducing factors under simulated microgravity condition.
Conclusion
In the present study, we evaluated the cytoskeletal and nucleoskeletal changes in hMSCs when cultured under simulated microgravity conditions. Our results indicate that the observed changes in cellular behavior are largely due to disruption of the fibronectin assembly. Fibronectin structural changes induced by MMPs are widely known to play important roles in the reorganization of cell–substrate interactions and the subsequent modification of nuclear lamina–cytoskeleton interactions. Taken together, this study provides insight into the effects of gravity on cell structure and behavior in addition to being a practical guide for device design, operation, and application in the study of gravity on cells in vitro.
Footnotes
Acknowledgments
The authors are grateful to Mr. Shinzou Sato, Mr. Ken-Ichi Nishisako, Mr. Toshinori Tawa, and Mr. Yoshitaka Kanetada (Miyuki Ironworks Co., Ltd., Hiroshima, Japan) for preparation of the Gravite® used in this study to generate the simulated microgravity conditions. This work was supported partially by the Program to accomplish research activities of female researchers and by scholar project program from Toyota Physical and Chemical Research Institute.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Movie S1
Supplementary Movie S2
Supplementary Movie S3
Supplementary Movie S4
Supplementary Movie S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.