
Abstract
Cardiomyocytes (CMs) derived from human embryonic stem cells (hESCs) or induced pluripotent stem cells (iPSCs) are used to study cardiogenesis and mechanisms of heart disease, and are being used in methods for toxiological screening of drugs. The phenotype of stem-cell-derived CMs should ideally resemble native CMs. Here, we compare embryonic/fetal CMs with hESC-derived CMs according to function and morphology. CM clusters were obtained from human embryonic/fetal hearts from elective terminated pregnancies before gestational week 12, and separated into atrial and ventricular tissues. Specific markers for embryonic CMs and primary cilia were visualized using immunofluorescence microscopy analysis. Contracting human embryonic cardiomyocyte (hECM) clusters morphologically and phenotypically resemble CMs in the embryonic/fetal heart. In addition, the contracting hECM clusters expressed primary cilia similar to that of cells in the embryonic/fetal heart. The electrophysiological characteristics of atrial and ventricular CMs were established by recording action potentials (APs) using sharp electrodes. In contrast to ventricular APs, atrial APs displayed a marked early repolarization followed by a plateau phase. hESC-CMs displayed a continuum of AP shapes. In all embryonic/fetal clusters, both atrial and ventricular, AP duration was prolonged by exposure to the KV11.1 channel inhibitor dofetilide (50 nM); however, the prolongation was not significant, possibly due to the relatively small number of experiments. This study provides novel information on APs and functional characteristics of atrial and ventricular CMs in first trimester hearts, and demonstrates that Kv11.1 channels play a functional role already at these early stages. These results provide information needed to validate methods being developed on the basis of in vitro-derived CMs from either hESC or iPSC, and although there was a good correlation between the morphology of the two types of CMs, differences in electrophysiological characteristics exist.
Introduction
Cardiomyocytes (CMs) derived from human embryonic stem cells (hESC-CMs) or induced pluripotent stem cells (iPSC-CMs) have provided new opportunities for studying cardiogenesis [1] and mechanisms underlying heart diseases [2] as well as for safety screening of pharmaceutical compounds [3]. Recently a comprehensive in vitro proarrhythmia assay (CiPA) has been proposed, where assessment of candidate drugs on the electrical activity of human stem-cell-derived ventricular CMs was used to detect ventricular proarrhythmic liability [4]. The KV11.1 voltage-gated potassium channel (also known as hERG) is of particular interest in safety screening programs as many pharmacological compounds bind nonspecifically to KV11.1 channels. These channels mediate the rapid delayed rectifier potassium current, IKr, which is important for the late repolarization phase of the cardiac action potential (AP). A decreased IKr is associated with a prolonged action potential duration (APD), which may cause long QT syndrome (LQTS) and sudden cardiac death in adults due to Torsade de pointes arrhythmias as well as uterine or fetal death [5,6]. Other potassium currents also contribute to the final repolarization phase of the AP in adult CMs resulting in a “repolarization reserve” [7].
hESC- and iPSC-CMs form nodal-, atrial-, and ventricular-like cell types as evidenced by the expression of specific genes and protein markers as well as electrophysiological characteristics, including AP shape [2]. However, both types of stem-cell-derived CMs often display a continuum of AP characteristics rather than chamber-specific phenotypes [8,9], and regulation of the differentiation into specific cardiac cell types is not fully elucidated. Recently, it was shown that retinoic acid signaling is required at the mesoderm stage for atrial specification [10], and nodal-like pacemaker cells can be generated from hESCs by a stage-specific manipulation of developmental signaling pathways [11]. We have demonstrated that hESC-CM clusters in some cases spontaneously organize into nodal-, atrial-, and ventricular-like subtypes in a spatial manner [12]. It is generally accepted that hESC- and iPSC-CMs express an embryonic or fetal phenotype [13,14], and a direct comparison of hESC-derived CMs with primary human fetal CMs corroborated this claim [15]. This presents a major obstacle to using stem-cell-derived CMs as a model system for studying cardiogenesis and heart diseases as well as optimizing strategies for screening drug cardiotoxicity.
Mummery et al. compared CMs derived from hES cells with fetal CMs from fetal hearts (gestational weeks 16–17). They found that most hESCs (28 of 33 total) resembled fetal ventricular cells, and they only found two atrial-like and one pacemaker-like dissociated aggregates [15]. However, the CM clusters were dissociated and replated before electrophysiological measurements, which could influence the hESC electrophysiology. We have previously mapped the intracluster electrophysiological heterogeneity and described areas within complex clusters resembling atrial, ventricular, or nodal APs [12]. A criterion based on the APD20/APD70 ratio and the maximum upstroke velocity was used to divide the APs into three subtypes resulting in 47% nodal-, 32% atrial-, and 21% ventricular-like AP subtypes within the clusters.
In this study, we functionally compared hESC-CM clusters with human embryonic/fetal atrial and ventricular tissue in culture regarding cell morphologies, expression of primary cilia as well as AP characteristics and the functional importance of the delayed rectifying potassium current, IKr.
Materials and Methods
hESC-derived cardiomyocytes
The hESC lines LRB010 and LRB024 derived at the Laboratory of Reproductive Biology, University Hospital Copenhagen, Denmark were used [12,16]. Derivation and use of hESCs from surplus donated embryos for disease modeling were approved by the Ethical Committee of the Capital Region in Denmark (H-KF-01-188/03) and conform to the principles outlined in the Declaration of Helsinki. The differentiation protocol used to generate hESC-CMs has previously been reported in detail [12]. In brief, hESCs were aggregated into embryoid bodies and cultured in 96-well plates in 20% fetal bovine serum (FBS) differentiation medium.
Human embryonic/fetal heart clusters
Human embryonic/fetal hearts were obtained from legal elective abortions before gestational week 12. The hearts used in this study were 7–12 gestational weeks old with the vast majority being embryonic (ie, before gestational week 11), and we refer to this material as embryonic. All participants received oral and written information, and gave their informed consent. Maternal exclusion criteria included the following: age of participant below 18 years, chronic diseases, and requirement of an interpreter. Pregnancies with known disorders or morphological malformation were excluded. All embryos and fetuses included in this study were morphologically normal. Participants answered a questionnaire concerning their lifestyle during the pregnancy, including smoking and drinking habits, and the study was approved by The Scientific Ethical Committee for the Capital Region [KF (01) 258206] and [KF (01) 170/99].
After collection, hearts were maintained in culture medium (DMEM/F12 with 10% FBS, 1% GlutaMAX, 1% Pen/Strep, 1% NNEA, 3% sodium pyruvate, and 1% insulin-selenium-transferrin; Thermo Fisher Scientific) at 0°C (on ice) for 6–8 h before they were prepared for culturing (see Supplementary Data for optimization of the procedure). The ventricular tissue was dissected from the atrial tissue and subsequently chopped into small pieces (∼0.1–1 mm3). The pieces were then transferred to gelatin-coated culture dishes, and cultured at 37°C with 20% O2 and 5.0% CO2 in culture medium as described above. The medium was exchanged every 2–3 days, and contracting clusters of 30–50 cells per cluster were counted and passaged every seventh day by dissection from the surrounding noncontracting cells.
Clusters were further divided equally into two parts and transferred to separate dishes. After 7 days of culture, two methods of passaging were performed in parallel: (1) colonies were mechanically split or (2) colonies were enzymatically digested by Tryple Select (Thermo Fisher Scientific, Denmark), and culture was continued for up to seven passages (see Supplementary Data). We evaluated the effect of transport temperature, coating, and mechanical versus enzymatic passaging by counting the number of contracting clusters and measuring the expression of cardiac proteins troponin T2 (TNNT2) and alpha-actinin 2 (ACTN2) (see Supplementary Figure S1).
Cryopreservation and thawing of human embryonic cardiomyocytes
Cardiac tissue that was not immediately put into culture was vitrified according to the open pulled straw method [17]. The tissue was submerged in prewarmed medium consisting of 10% DMSO, 10% ethylene glycol, and 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) (referred to as VS1) for 60 s, and subsequently transferred to prewarmed medium consisting of 20% DMSO, 20% ethylene glycol, 0.5 M sucrose, and 10 mM HEPES (referred to as VS2) for 25 s. Finally, the pieces were transferred to a 5-μL droplet of VS2 medium after which the straw was loaded with the tissue and submerged in liquid nitrogen.
The tissues were warmed by waving the straws in the air for 3 s and expelling the tissue to prewarmed medium consisting of 0.2 M sucrose and 10 mM HEPES. After 60 s, the tissue pieces were transferred to prewarmed medium consisting of 0.1 M sucrose and 10 mM HEPES for 5 min, and finally transferred to gelatin-coated dishes for continued culturing.
Fixation, embedding, and sectioning of embryonic cardiomyocyte clusters for histochemistry
Contracting human embryonic cardiomyocyte (hECM) clusters were isolated from the surrounding tissue using a 23-G needle. The clusters were embedded in 0.5% alginate and 4% agar before fixation in 4% neutral buffered formaldehyde. Five-micrometer histological sections were prepared to use for immunohistochemistry using standard histological methods. One human embryonic heart was fixated in Bouins solution without alginate embedding and processed in parallel with the clusters. Representative sections were stained with hematoxylin and eosin (HE).
Immunofluorescence microscopy analysis
The immunofluorescence microscopy (IFM) analysis was carried out as described previously [12,18]. In brief, samples were deparaffinized following standard protocols, and boiled with citrate or Tris-EGTA buffer (TEG) antigen retrieval buffers for 20 min, cooled and rinsed in tris-buffered saline (TBS). A Dako Pen (Dako, Agilent Santa Clara, CA) was used to encircle the tissue sections, and primary antibodies (Supplementary Table S1) diluted in Dako REAL Antibody Diluent (Dako) were incubated overnight at 4°C in a humidified chamber. After phosphate-buffered saline (PBS) washing steps, cells were incubated in corresponding Alexa Fluor–conjugated secondary antibodies (Supplementary Table S1) for 45 min at room temperature. Cell nuclei were labeled by DAPI staining. Coverslips were mounted in PBS, 90% glycerol, and 2% N-propyl-gallate or with Dako fluorescent mounting medium (Dako) on glass slides and sealed with nail polish.
Fluorescence and differential interference contrast images were captured on a fully motorized Olympus BX63 upright microscope with an Olympus DP72 color, 12.8-megapixel, 4.140 × 3.096-resolution camera and with a fully motorized and automated Olympus IX83 inverted microscope with a Hamamatsu ORCA-Flash 4.0 camera (C11440-22CU). The software used was Olympus CellSens dimension, which was able to perform deconvolution on captured z stacks, and images were processed for publication using Adobe Photoshop CS6.
AP recordings
APs were recorded using an Axon 700B amplifier and pClamp 10.4 software (Axon instruments, Molecular Devices). Data were digitized at 20 kHz and filtered at 10 kHz using a Digidata 1440 digitizer (Axon instruments, Molecular Devices). Sharp electrodes were filled with 3 M KCl, resulting in a 20–50 MΩ resistance when submerged in the bath solution: 140 NaCl mM, 5.4 KCl mM, 1 MgCl2 mM, 1.8 CaCl2 mM, 10 mM HEPES, 10 mM glucose, pH 7.4. All experiments were performed at 37°C. The hESC-CM and hECM clusters were spontaneously contracting, and did not respond to pacing stimuli. APs were recorded in the absence and presence of 50 nM dofetilide (Sigma Aldrich, Denmark) after a 20–40 min wash-in period and a similar period of washout. Data were analyzed using Clampfit 10.4 and Labchart 8 (AD Instruments, United Kingdom). APs from hESC-CM clusters were categorized in the same manner as we have previously published [12]. APs were divided into ventricular like or atrial like based on the ratio between APD at 20% or 70% of repolarization (Fig. 3): nodal like: APD20/APD70 ratio <0.35 and upstroke velocity <25 V/s. Atrial like: APD20/APD70 ratio <0.35 and upstroke velocity >25 V/s. Ventricular like: APD20/APD70 ratio >0.35 and upstroke velocity >25 V/s. APD was measured at 50% upstroke, and APs with amplitudes <30 mV were not included in the analysis.
Cycle length (CL) corrected APD70 was calculated using modified Bazett's formula: cAPD70 = ADP70/CL2. Data are presented as mean ± SEM.
Results
hECM morphology and expression of specific cardiac markers
To characterize clusters of atrial and ventricular CMs from embryonic hearts, we initially performed HE staining and IFM analysis with antibodies against α-smooth muscle actin (α-SMA), β-III tubulin, and myosin light chain 2v (MLC-2V), which mark immature CMs [19], neuronal cells [20], and embryonic ventricular myocardium, respectively [21 –23]. Further, MLC2v has been used to select for ventricular iPSCs [24]. As shown in Fig. 1A, the ventricular cluster with a diameter of ∼4–5 mm exhibited a dense core of CMs with a demarcated periphery, whereas the atrial cluster exhibited a sparser cellular organization (Fig. 1E). Both the ventricular and atrial clusters were obtained from the fourth culturing passage. The α-SMA was expressed in both atrial and ventricular clusters, most predominantly in the peripheral part of the clusters (Fig. 1B, F). In agreement with previous studies [21], MLC-2V was highly expressed in the ventricular cluster (Fig. 1B), with sarcomeres displaying a striated pattern of MLC-2V and α-SMA (Fig. 1C), whereas MLC-2V was absent or expressed at a very low level in atrial clusters as expected (Fig. 1F, G). Finally, βIII-tubulin staining was observed in distinct foci in both ventricular (Fig. 1D) and atrial clusters (Fig. 1H), although most prominently in elongated cellular structures in the former, which may signify neuronal cell types.
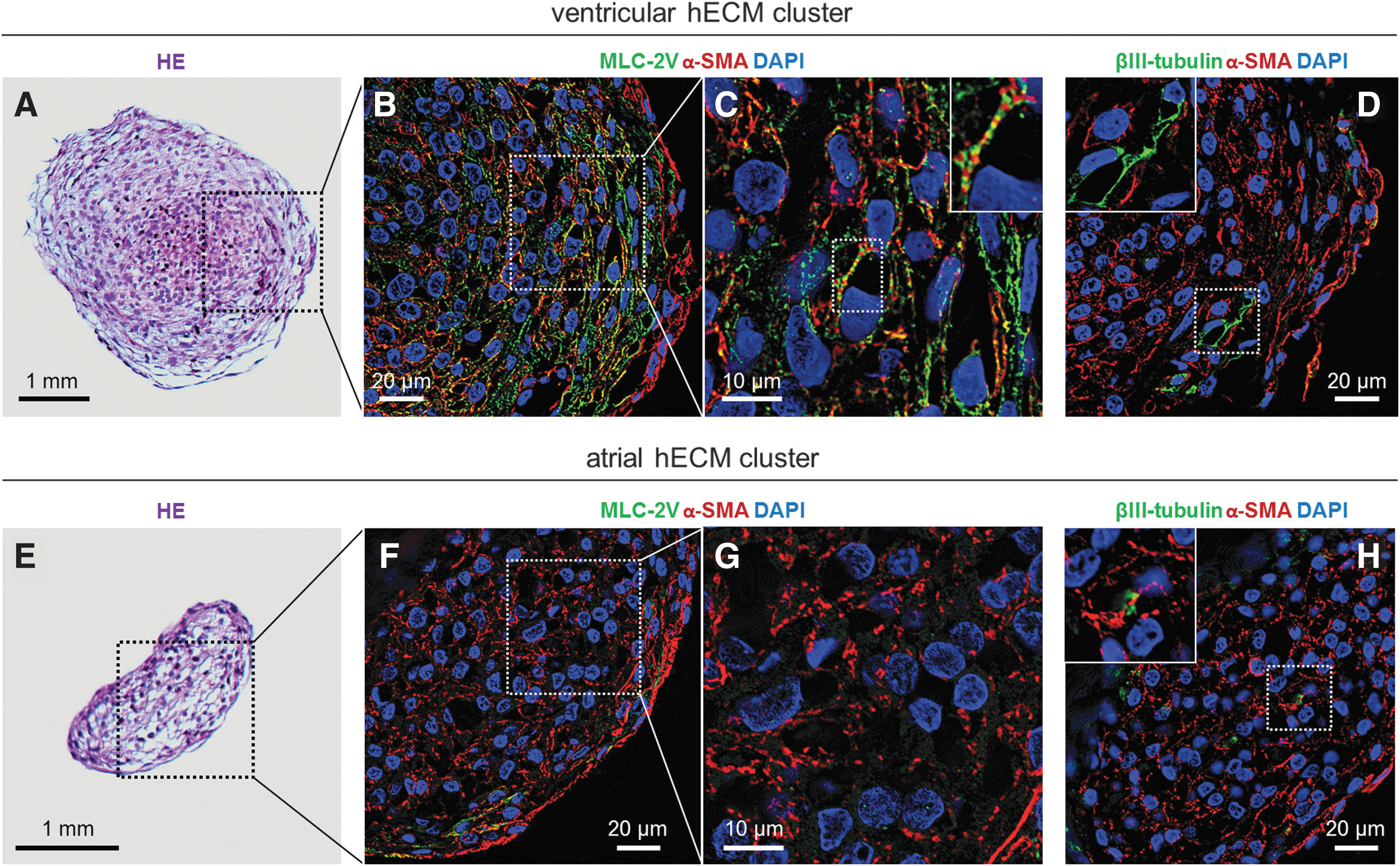
Cell morphology and expression of cardiac markers in hECM clusters.
Expression of primary cilia in hECMs
Primary cilia are solitary organelles that are present on the surface of most quiescent mammalian cell types to coordinate diverse cellular signaling pathways during developmental processes and in tissue homeostasis [25 –27]. We previously demonstrated that hESC-CMs form primary cilia [12,28,29], which are required for in vitro cardiomyogenesis and proper heart development, and when defective can give rise to heart disease [30,31].
To investigate the presence of primary cilia in contracting hECM clusters derived from ventricular tissue we performed IFM with an antibody against ADP-ribosylation factor-like protein 13B (ARL13B), which is a marker for primary cilia [32,33]. Antibody specificity and ciliary formation was further evaluated by IHC analysis of ventricular tissues in isolated human embryonic hearts with a combination of ARL13B and IFT172 antibody. IFT172 is part of the intraflagellar transport machinery and predominantly marks the base and tip of cilia [34]. As shown in Fig. 2A, primary cilia with lengths varying between 3 and 6 μm were present in cells of the myo- and endocardium of the left and right ventricles, in the intraventricular septum as well as at the outer surface of the pericardium (Fig. 2A). Similarly, ARL13B-positive primary cilia with lengths comparable with those observed in the embryonic heart were present in hECMs of the contracting clusters, and to some extent what appears to be regional differences in their orientation. Thus, cells positioned deeply within the cluster often presented a randomly oriented population of cilia, whereas cilia facing the periphery of the clusters, which is the site for predominant α-SMA expression (Fig. 2B), typically oriented parallel to the long axis of the clusters (Fig. 2B, upper left panel). These results show that contracting hECM clusters form primary cilia similar to that of cells in the embryonic heart, although further experiments will be needed to evaluate in detail the spatiotemporal formation, orientation, and sensory function of primary cilia in the context of cell types and cellular organization during the development of contracting hECM clusters.
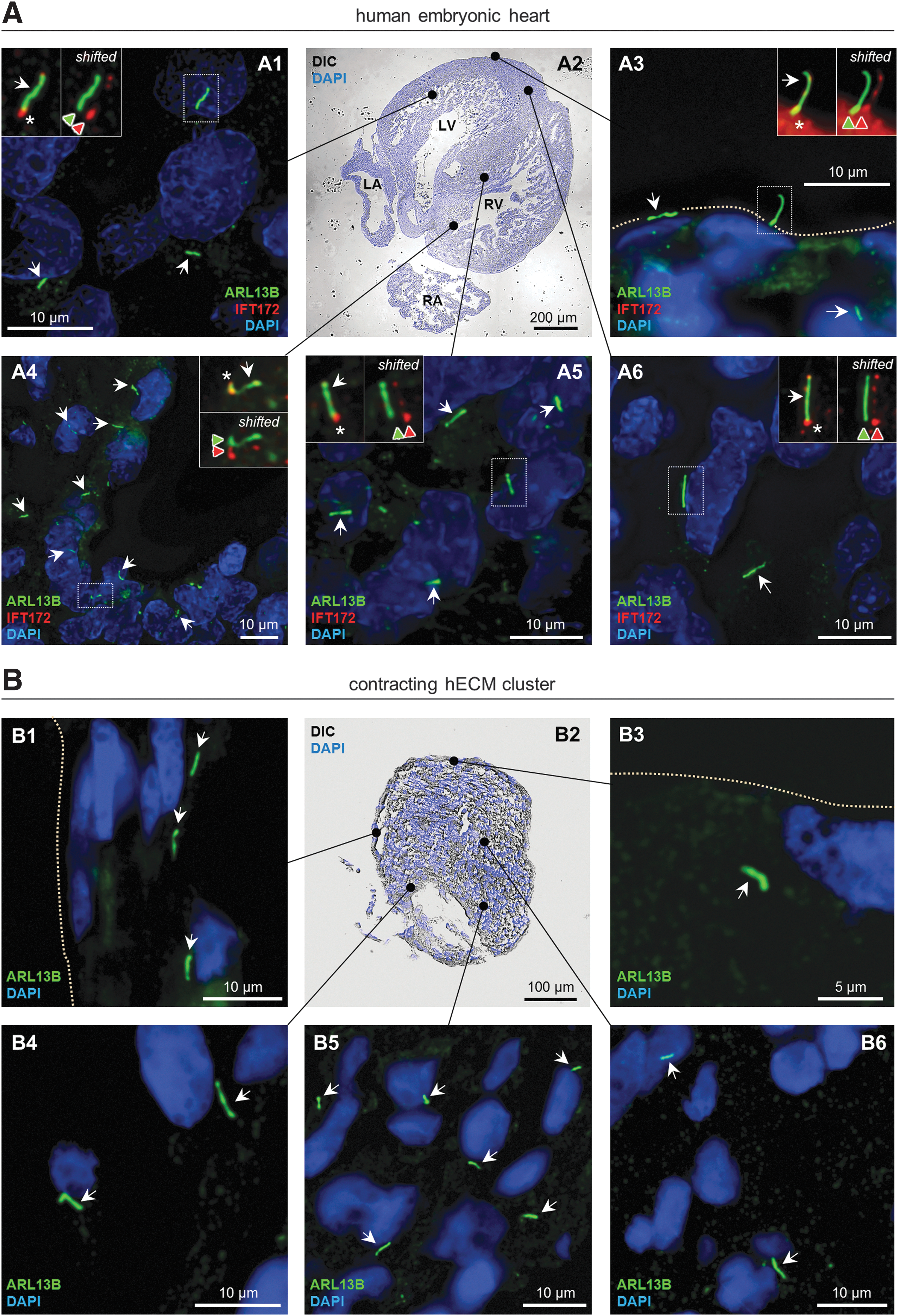
Distribution and organization of primary cilia in human embryonic heart tissue.
AP shape in human embryonic atrial and ventricular clusters compared with hESC-derived cardiomyocyte clusters
AP shape in human embryonic atrial and ventricular clusters was investigated using sharp electrodes (Fig. 3). The mean resting membrane potential (RMP) was comparable between clusters isolated from embryonic atria and ventricles, with RMP = −61 ± 5 and −60 ± 4 mV, respectively (Table 1). Both atrial and ventricular clusters exhibited a slow spontaneous depolarization during phase 4, which triggered a rapid depolarization (>10 V/s). In atrial clusters, the spontaneous phase 4 depolarization was steeper compared with ventricular clusters, compatible with a faster spontaneous contraction rate of atrial clusters (Tables 1 and 2). The AP shape was distinctively different, and consistent with previous findings, we found that APs from human embryonic atrial clusters had a marked early repolarization, whereas ventricular APs did not [15,35 –37] (Fig. 4). This is reflected in significantly shorter APD at 20% repolarization (APD20) for atrial APs compared with ventricular APs (Fig. 4C). APD at 70% repolarization (APD70) was also significantly shorter in embryonic atrial clusters compared with ventricular clusters (Fig. 4D); however, this difference was not present after correction for the differences in CL using Bazett's formula. The ratio between APD20 and APD70 has been used to discriminate atrial- and ventricular-like APs in iPSC and hESC cultures. The APD20/APD70 ratio was significantly different in APs from clusters of embryonic atria compared with APs from embryonic ventricular clusters (Fig. 4E and Table 1). For comparison, APs were recorded from spontaneously contracting hESC-CM clusters. Some clusters or specific areas of clusters exhibited nodal, atrial-, or ventricular-like APs based on APD20/APD70 ratio and the rate of the phase 4 depolarization (Materials and Methods section), but many clusters exhibited intermediate AP characteristics. The characteristics of APs from hESC-CM clusters are summarized in Table 1. For atrial-like APs from hESC-CM clusters, APD20 and APD70 were comparable with APD20 and APD70 of embryonic atrial APs (Fig. 4C, D). However, for ventricular-like APs from hESC-CM clusters, both APD20 and APD70 were significantly shorter compared with those of embryonic ventricular APs. This suggests a difference in repolarizing currents in hESC-CM and hECM clusters.

Action potentials recorded from human embryonic atrial or ventricular clusters. Action potentials were recorded using sharp electrodes from spontaneously contracting human embryonic atrial or ventricular clusters.

Action potentials recorded from hESC-derived cardiomyocyte clusters. Action potentials recorded from hESC-derived cardiomyocyte clusters exhibit a high degree of variation in contrast to action potentials from human embryonic atrial or ventricular clusters. Action potentials were recorded using sharp electrodes from spontaneously contracting clusters.
Action Potential Characteristics
Parameters of action potentials recorded from human embryonic atrial and ventricular clusters (7–9 weeks post conception) and hESC-CM clusters. Action potentials from hESC-CM clusters were categorized into being either nodal like, ventricular like, or atrial like based on the APD20/APD70 ratio and upstroke velocity (see the Materials and Methods section). The three rows with data from hESC-CMs have previously been published but are included for the sake of comparison12. Numbers in parentheses indicate the number of hearts the atrial or ventricular clusters originated from.
n = 5, for three ventricular clusters we did not find consecutive spontaneous contractions, only random contractions with large intervals.
RMP, resting membrane potential; APA, action potential amplitude; APD, action potential duration at 20% or 70% repolarization; cAPD70, corrected APD70 using Bazett's formula; hESC, human embryonic stem cell.
Effects of Dofetilide on Action Potentials Characteristics
Parameters of action potentials recorded from human embryonic atrial and ventricular clusters (7–9 weeks post conception) and hESC-CM clusters divided into nodal-, atria-, and ventricle like (see the Materials and Methods section) before and after 20–40 min of superfusion with 50 nM dofetilide. Numbers in parentheses indicate the number of hearts the atrial or ventricular clusters originated from. Mean ± SEM are shown, P values calculated by a paired t-test.
= P < 0.05.
= P < 0.01.
n = 2; for one of the clusters there was no spontaneous contraction but only random contractions with large intervals, hence correction for cycle length was not possible.
To further investigate the difference in repolarization, the IKr blocker dofetilide was applied. Dofetilide blocks Kv11.1 with an IC50 of 15.3 nM [38] and application of 50 nM prolonged APD70 in embryonic ventricular clusters from 308 ± 40 ms to 512 ± 5 ms and in atrial clusters from 101 ± 21 to 212 ± 67 ms (Fig. 5 and Table 2). All hECM and hESC-CM clusters responded with an increased APD70 (Fig. 5D, Table 2 and Supplementary Fig. S2). The prolongation was not significant, possibly due to the relatively small number of experiments. In some clusters, the spontaneous beating rate changed during the experiment; after correction for CL, the change in APD70 in the presence of dofetilide was still not significant (Table 2). The effect of dofetilide was reversible, and APD shortened after washout (data not shown). In several embryonic ventricular clusters, one or more consecutive early after-depolarizations (EAD) were observed, suggesting that IKr is of major functional importance in embryonic hearts. Despite a prolongation of embryonic atrial APs, EADs were not observed in embryonic atrial clusters. Dofetilide did not alter the RMP, AP amplitude, or upstroke velocity significantly in embryonic clusters. In hESC-CM clusters, dofetilide also prolonged APD; however, not to the same extent as in hECM clusters (Table 2 and Supplementary Fig. 3) and no EADs were observed. In several hESC-CM clusters, dofetilide had a depolarizing effect on the membrane potential, resulting in a loss of AP activity, consistent with previous reports [8,39]. These clusters were not included in the statistics. Interestingly, this phenomenon was not observed in hECM clusters, suggesting that IKr is crucial for setting the RMP in hESC-CMs, as also demonstrated previously for hiPSC [8,40]. IK1 is important for setting the RMP in adult ventricular CMs, but IK1 is very low or negligible in hiPSCs [8,40]. We speculate that IK1 may also be minor or negligible in hESC-CM, leaving IKr to set the RMP.
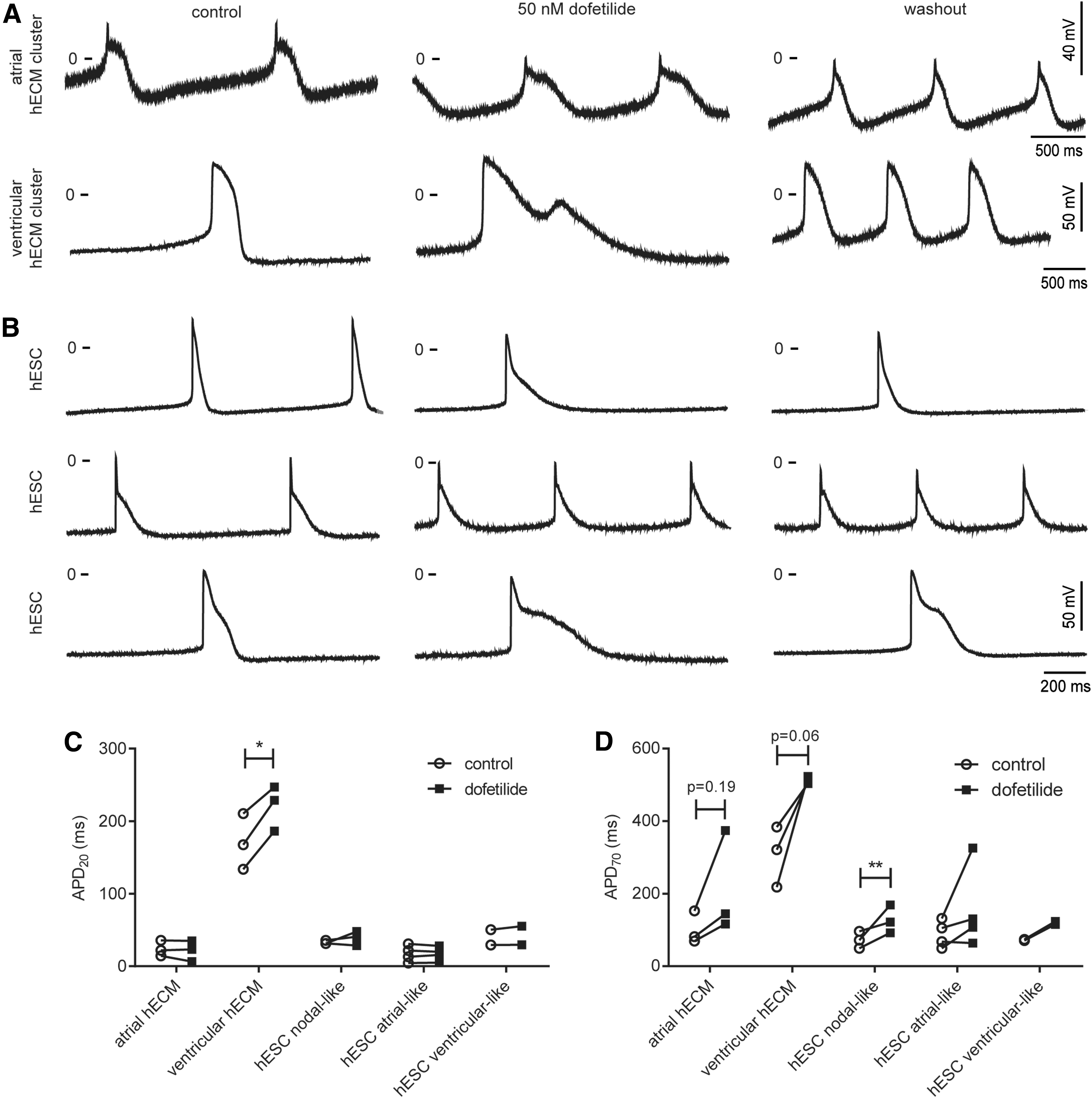
Effect of dofetilide on action potentials recorded from human embryonic atrial or ventricular clusters and hESC-derived cardiomyocyte clusters. Action potentials were recorded using sharp electrodes from spontaneously contracting clusters before and after 20–40 min exposure to 50 nM dofetilide followed by washout.
For all experiments, there was no difference in clusters derived from embryos/fetuses obtained from women who smoked compared with those who did not.
Discussion
The hESC-CMs resemble hECMs to a large extent, although some important differences were found, including differences in APD and response to the Kv11.1 blocker dofetilide. The study provides new information for validation of the methods being developed on the basis of in vitro-derived CMs from either hESC or iPSC. Further, this study demonstrates that functionally active hECMs can be cultured for several passages, and that cells maintain their characteristics after freezing and thawing.
Cell morphology and expression of cardiac markers
Morphologically, hESC-CMs resemble immature CMs from various stages of embryonic development with regard to structural organization and AP properties [41 –43]. hESC-CMs do not exhibit the homogeneous rod shape seen in adult CMs. The hESC-CM and hECM cell clusters both exhibited a variety of forms, including spindle, multiangular, or rod shape with bundled, rather than highly ordered, sarcomeres found in the homogeneous rod-shaped adult CMs, as we have previously reported [12]. Consistent with this, transcriptional analysis of hESC-CMs typically displays a wide array of genes characteristic of fetal CMs [40]. Further, the hESC-CMs exhibited differential spatial expression of βIII-tubulin, MLC2v, and α-SMA [12], and a similar organization was found for hECM clusters. The α-SMA was most pronounced in cells lining the peripheral parts of the clusters, which is an indication of CM progenitor cells or myofibroblast formation [12,44]. We previously demonstrated that the βIII-tubulin-positive cell populations reflect nodal CMs, and βIII-tubulin expression was confined to minor cell populations [12].
The gene expression profile of embryonic and fetal CMs varies a lot during different stages of development and between heart chambers. The changes in gene expression have been investigated by Berg and colleagues [45]. They demonstrated that based on gene expression profile, hPSC-CMs have an expression profile most similar to first-trimester fetal hearts. Our results suggest that hESC-CMs display an even younger phenotype than week 7–12, including differences in AP repolarization and a lack of specialization into different cell types. It would be interesting to investigate expression profile of genes encoding proteins that modify AP waveform, but due to the very limited supply of samples it was not possible to carry out gene expression assays.
Formation and orientation of primary cilia in hECM clusters
Primary cilia coordinate multiple cellular processes during heart development and play a central role in the pathogenesis of congenital heart disease, which is the most prevalent birth defect [30,31]. Specification and organization of CMs as well as cardiac tissues during in vivo heart development are associated with variations in ciliary orientation [33,44,46], and primary cilia play a critical role in regulating the balanced output of diverse signaling pathways in cardiovascular development, such as in the regulation of TGFβ/BMP signaling [29,47,48]. In this study, primary cilia were demonstrated in hECM clusters derived from ventricular tissue, and we show that cilia are formed to an extent similar to that of the human embryonic heart as well as of CMs derived from hESCs [12]. We further noticed that hECM clusters displayed regional difference in orientation of primary cilia, such that cilia in α-SMA-positive cells at the periphery of the clusters predominantly oriented parallel to the long axis of the clusters, whereas cells within the clusters displayed a more randomly oriented population of cilia. We suggest that hECM clusters in the context of primary cilia may serve as a novel human cardiac organoid model to study cardiogenesis and mechanisms underlying heart disease as well as for drug screening, in which the identity of ciliary signaling pathways that contribute to cluster formation, organization, and function can be addressed in a spatial-temporal manner.
AP characteristics in hECM and hESC-CM clusters
AP morphology differs in isolated cells and in multicellular preparations [49] and with regard to electrophysiology, this study differs from the studies by Mummery et al. [15] and Danielsson [35] and colleagues, as we did not dissociate the hESC-CM clusters before electrophysiological measurements. In embryonic clusters, atrial and ventricular APs could clearly be discerned. In contrast, the APs in hESC-CM clusters displayed a continuum of characteristics. APD70 was markedly longer in embryonic ventricular clusters compared with all hESC-CM clusters. In isolated embryonic CMs, the APD50 has previously been reported as 150–200 ms [35] and the APD90 reported as 370 ms [15], which is well in agreement with the APD70 of 228 ± 31 ms for embryonic ventricular clusters.
Mutations in the KCNH2 gene encoding the Kv11.1 channel have been implicated in fetal and intrauterine deaths [6], and IKr has previously been demonstrated in embryonic isolated CMs. While the Mummery et al. study [15] did look at the chronotropic effect of the calcium antagonist verapamil and several other compounds on the spontaneous beating of hESC-CMs and fetal CMs, they did not investigate the effect of blocking IKr. We found that in embryonic atria and ventricular clusters, dofetilide induced a marked increase in APD, confirming that IKr play a functional role.
In a study by Danielsson and coworkers, cells from gestational week 5 did not respond with an increase in APD to the Kv11.1 blocker E-4031, whereas cells from weeks 7 to 9.5 did. All cells showed subthreshold oscillations of the membrane potential and disturbances in rhythm generation [35]. We did not observe these oscillations, but this is likely explained by the different experimental conditions as cell coupling in multicellular preparations tends to even out oscillations. However, we did observe several ventricular EADs during dofetilide treatment. This is an important finding, and suggests that IKr blocking compounds or substances that affect Kv11.1 channels may be a contributing factor to the relatively high frequency of pregnancy failures during the first trimester of pregnancies. Indeed, epidemiological studies have linked the use of specific drugs associated with LQTS and torsade de pointes, such as clarithromycin [50], paroxetine, and venlafaxine, to an increased risk of spontaneous abortions [51], and other Kv11.1 inhibitors such as the antidepressant sertraline and citalopram [52,53] have been associated with septal heart defects in humans. Teratology studies in animal models suggest that malformations and embryonic deaths are due to cardiac arrhythmias and hypoxia-reoxygenation damage [35,54,55].
Dofetilide has also been indicated as a highly proarrhythmic drug in a multisite hiPSC-CM electrophysiological model [56]. In agreement with this, dofetilide increased APD in most of the hESC-CM clusters, but in several clusters application of dofetilide resulted in depolarization of the membrane potential leading to loss of excitability, in agreement with previous observations in iPSC-CMs after application of the IKr blocker E-4031 [8,39]. This phenomenon was not observed in hECM clusters, suggesting that IKr is crucial for setting the RMP in hESC-CMs, whereas a more complex repolarization reserve may be present in human embryonic hearts even at very early stages of cardiac development. The immature phenotype of hESC-CMs with regard to AP characteristics as well as differences in response to Kv11.1 inhibitors compared with embryonic and adult CMs poses challenges for using hESC-CMs as models for CMs. Ideally, more pathways and factors that stimulate development of CMs into more specific and more mature phenotypes should be identified [13]. We acknowledge that the results of dofetilide are preliminary and should be tested in more detail in future studies.
The fact that hESC-CMs display an apparent immature phenotype compared with hECMs could indicate differences in the developmental paths of the two cell types. This information should be taken into account when validating new methods being developed on the basis of stem-cell-derived CMs. This requires more in-depth analysis such as single-cell RNA-seq as well as IFM analysis. In this context, it would be interesting to analyze in detail the function of primary cilia in hECMs and hESC-derived CMs, since these organelles are major coordinators of developmental signaling pathways.
In conclusion, before gestational week 12 the electrophysiological characteristics of atrial and ventricular CMs were established. Even though the morphology of the CMs is immature with rounded and spindle-shaped cells, clear electrophysiological distinctions between atria and ventricle CMs were observed. The study provides new information for validation of the methods being developed on the basis of stem-cell-derived CMs, and although there was a good correlation between the two types of CMs slight differences appeared to exist. Further, we suggest that hECM clusters in the context of primary cilia may serve as a novel system to study cardiac development and disease as well as for drug screening.
Footnotes
Acknowledgments
We thank technician Marjo Westerdahl for help with cultures of hESC.
This work was supported by the Faculty of Health Sciences, University of Copenhagen (M.L.V., K.K.), The University of Copenhagen Excellence Programme for Interdisciplinary Research (2016 Funds) (S.T.C.) and the Danish Council for Independent Research (1331-00254) (S.T.C.) and Rigshospitalets Forskningspuljer (L.S.M.).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.