
Abstract
There is compelling evidence that the mature central nervous system (CNS) harbors stem cell populations outside conventional neurogenic regions. We previously demonstrated that brain pericytes (PCs) in both mouse and human exhibit multipotency to differentiate into various neural lineages following cerebral ischemia. PCs are found throughout the CNS, including cerebellum, but it remains unclear whether cerebellar PCs also form ischemia-induced multipotent stem cells (iSCs). In this study, we demonstrate that putative iSCs can be isolated from poststroke human cerebellum (cerebellar iSCs [cl-iSCs]). These cl-iSCs exhibited multipotency and differentiated into electrophysiologically active neurons. Neurogenic potential was also confirmed in single-cell suspensions. DNA microarray analysis revealed highly similar gene expression patterns between PCs and cl-iSCs, suggesting PC origin. Global gene expression comparison with cerebral iSCs revealed general similarity, but cl-iSCs differentially expressed certain cerebellum-specific genes. Thus, putative iSCs are present in poststroke cerebellum and possess region-specific traits, suggesting potential capacity to regenerate functional cerebellar neurons following ischemic stroke.
Introduction
Stroke is the major cause of death throughout the world, and many stroke survivors are left with debilitating sequelae, such as paresis. There are now effective reperfusion therapies, such as recombinant tissue plasminogen activator and/or neuroendovascular therapies [1 –3], but due to the limited time window for success and reperfusion failure, only a small population of stroke patients benefit from these treatments. Thus, it is necessary to establish alternative therapeutic options for patients with ischemic stroke.
Accumulating evidence suggests that many stem cell types, such as embryonic stem (ES) cells, induced pluripotent stem (iPS) cells, and mesenchymal stem cells (MSCs), are potential sources of therapeutic cells for regenerative treatment following stroke [4 –6]. However, ethical issues and possible tumorigenesis are challenges to ES-based and iPS-based therapies, while there is doubt about the capacity of MSCs to differentiate into functional neurons [7,8]. Thus, alternative stem cell sources may be required for successful stem cell-based therapy following ischemic stroke.
Using a mouse model of cerebral infarction, we and others have demonstrated the induction of endogenous stem cells under pathological conditions (called ischemia/injury-induced multipotent stem cells [iSCs]) within the poststroke brain [9 –13]. We also demonstrated that these murine iSCs are self-propagating and differentiate into multiple cell types, including neurons. More recently, we showed that putative iSCs with neuronal differentiation potential are present within the poststroke human cerebrum [14].
Ischemic stroke also occurs outside the cerebrum, such as in the cerebellum. The cerebellum is the second-largest compartment of the brain, and damage and/or dysfunction may impair motor control, gait, gaze stability, and various cognitive functions [15]. In addition, it is reported that cerebellum-derived stem cells with region-specific traits [16] may be superior for the treatment of cerebellar diseases [17]. Thus, it is important to understand the traits of cerebellum-derived stem cells to develop cell-based therapies targeting cerebellar diseases. However, because of the difficulty establishing appropriate animal models of cerebellar diseases, including cerebellar infarction, basic research has progressed slowly, and the characteristics of cerebellum-derived stem cells remain unclear.
In this study, we isolated iSCs from poststroke cerebellum and compared their traits to various stem cells, including cerebral iSCs (cr-iSCs), MSCs, and neural stem/progenitor cells (NSPCs). Furthermore, we examined whether cl-iSCs have multipotency and can differentiate into functional neurons.
Materials and Methods
Patients
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Informed consent was obtained from all individual participants included in the study. The Ethics Committee of Hyogo College of Medicine reviewed and approved the study protocol (approval no.: 1776, 0385). Brain samples were obtained from poststroke areas of two patients who satisfied the following major criteria: [1] requiring both decompressive craniectomy and partial lobectomy for diffuse cerebellar infarction, [2] >20 years of age, and [3] willing to provide written informed consent. Detailed methods and criteria were described previously [14].
Histological examination
Histological analyses were performed as described [14]. In brief, tissue samples obtained from the cerebellar cortex of patients with cerebellar infarction were fixed with periodate-lysine-paraformaldehyde, cryoprotected in 30% sucrose, frozen at −80°C, and cut into 20-μm sections using a cryostat. Sections were then stained with hematoxylin and eosin (H&E) and examined by immunohistochemistry as described below.
Immunohistochemistry
Immunohistochemistry was performed as described [14]. In brief, sections were labeled with antibodies against Tuj1 (1:1,000; STEMCELL Technologies, Vancouver, BC, Canada), microtubule-associated protein 2 (MAP2, 1: 500; Sigma, St. Louis, MO), glial fibrillary acidic protein (GFAP, 1:200; Millipore, Temecula, CA), S-100β (1:100, Abcam; Cambridge, United Kingdom), nestin (1:100, Santa Cruz Biotechnology; Dallas, TX), von Willebrand factor (1:100, vWF; Santa Cruz Biotechnology), neuron-glial antigen 2 (NG2, 1:200; Millipore), and alpha smooth muscle actin (αSMA, 1:100; LifeSpan Biosciences, Seattle, WA). Primary antibodies were visualized using Alexa Fluor 488- or 555-conjugated secondary antibodies (1:500; Molecular Probes, Eugene, OR). Nuclei were counterstained with 4′,6-diamino-2-phenylindole (DAPI, 1:500; Kirkegaard & Perry Laboratories, Gaithersburg, MD). Images of brain sections were captured using a laser scanning microscope (LSM780, Carl Zeiss AG; Oberkochen, Germany) or fluorescence microscopy (Keyence BZ-X700; Keyence, Osaka, Japan). For negative controls in the immunohistochemistry experiments, we omitted the primary antibodies. No staining was observed in the control sections.
Cell culture and immunocytochemistry
The traits of cl-iSCs were investigated as described [14]. In brief, human ischemic tissue samples obtained from the cerebellar cortex were mechanically dissociated by passage through 23- and 27-G needles to yield single-cell suspensions. Suspensions were seeded on 10-cm dishes (Corning, NY) in Dulbecco's modified Eagle's medium/F12 (DMEM/F12; Invitrogen, Carlsbad, CA) containing basic fibroblast growth factor (bFGF, 20 ng/mL; Peprotech, Rocky Hill, NJ), epidermal growth factor (EGF, 20 ng/mL; Peprotech), 10% fetal bovine serum (FBS), and N2 (1%; Invitrogen). When the cells reached confluence, they were treated with trypsin and reseeded under the same conditions.
Passage 5–10 cells (cl-iSCs) were examined by immunocytochemistry, reverse transcriptase-polymerase chain reaction (RT-PCR), and FACS. Immunocytochemistry was performed using antibodies against Ki67 (1:50; BD Pharmingen, San Jose, CA), nestin (1: 100; Santa Cruz Biotechnology), αSMA (1: 200; LifeSpan Biosciences), NG2 (1: 200; Millipore), platelet-derived growth factor receptor beta (PDGFRβ, 1: 200; Santa Cruz Biotechnology), GFAP (1: 1000; DAKO, Glostrup, Denmark), CD31(1: 50; Santa Cruz Biotechnology), and CDH5 (1: 50; Santa Cruz Biotechnology), followed by Alexa Fluor 488- or 555-conjugated secondary antibodies (1: 500; Molecular Probes).
For mesoderm differentiation, cl-iSCs were incubated in medium to promote osteogenic, adipogenic, or chondrocytic differentiation according to the manufacturer's protocol (SC 006; R&D systems, Minneapolis, MN). The medium was changed every three days. Two weeks after incubation, each specimen was subjected to immunocytochemistry using antibodies (SC 006; R&D systems) against osteocalcin, fatty acid binding protein 4 (FABP4), or aggrecan. In addition, iSCs incubated in adipogenic differentiation medium were stained with the lipid-specific dye Oil Red O.
For neuronal differentiation, cl-iSCs were incubated for 1 week in DMEM/F12 containing bFGF, leukemia inhibitory factor (LIF; Millipore), 10% FBS, and N2 to promote the formation of neurosphere-like cell clusters, as previously described [14]. The medium was changed every three days. Then, cell clusters were picked up and subjected to RT-PCR and immunocytochemistry. The differentiated cells were immunostained with antibodies against Tuj1 (1:500; STEMCELL Technologies), neurofilament (NF, 1:200; Millipore), and MAP2 (1:500; Cell Signaling Technology, Danvers, MA) followed by Alexa Fluor 488- or 555-conjugated secondary antibodies (1: 500; Molecular Probes).
Human pediatric brain-derived endothelial cells (ECs, ACBRI-376; Cell Systems, Kirkland, WA), human adult bone marrow (BM)-derived MSCs (PT-2501; Lonza, Basel, Switzerland), human NSPCs derived from H9 embryonic stem cells (NSPCs, Part. No. SCC003; Millipore), human fetal brain-derived astrocytes (ACs, #1800; ScienCell Research Laboratories, Carlsbad, CA), and human fetal brain-derived pericytes (PCs, #1200; ScienCell Research Laboratories) were maintained in media specified by the supplier.
Briefly, ECs were planted in 10-cm dishes (Corning, NY) and maintained in EGM-2 medium containing vascular endothelial growth factor, bFGF, EGF, insulin-like growth factor, hydrocortisone (CC3156; Lonza), and 10% FBS. MSCs were planted in T25 flasks (Corning, NY) and maintained in mesenchymal stem basal medium (Lonza) containing MSC growth supplement (Lonza),
The ECs were immunostained with antibodies against CD31 and CDH5 as a positive control. Extracted RNA samples from ECs, MSCs, NSPCs, ACs, and PCs were used as controls for microarray analyses of cl-iSC gene expression patterns.
Cell proliferation
To evaluate the proliferative capacity of cl-iSCs, 1 × 104 cells were plated onto 12-well plates in DMEM/F12 containing bFGF, EGF, 10% FBS, and N2. Then, the numbers of cl-iSCs were counted manually over 7 days (N = 4). Doubling time was calculated as described [14,18].
Reverse transcriptase-polymerase chain reaction
Total RNA was isolated from cl-iSCs and ACs using an RNeasy Micro Kit (Qiagen, Hilden, Germany). cDNA was amplified according to the manufacturer's protocol as described [9 –11,14,19]. The primer sequences used in this study are listed in Supplementary Table S1.
Microarray analysis
Total RNA was isolated from cl-iSCs from the first patient (cl-iSCs-1) and second patient (cl-iSCs-2), as well as from ECs, MSCs, NSPCs, ACs, and PCs using an RNeasy Micro Kit (Qiagen, Hilden, Germany). In addition, total RNA was obtained from two different cerebral stroke patients (cr-iSCs-1, cr-iSCs-2) [14]. For microarray analysis, three samples were used for each cell type. Microarray analysis, including RNA quality assessment, labeling, hybridization, scanning, and data analysis, was performed by a contract service (Takara Bio, Inc., Shiga, Japan). In brief, biotinylated cRNA was synthesized using the GeneChip 3′IVT PLUS Reagent Kit (Affymetrix; Thermo Fisher Scientific, Inc., Waltham, MA) from 250 ng total RNA. Biotinylated cRNA production was checked using a NanoDrop ND-2000 Spectrophotometer. Following fragmentation, 15 μg of cRNA was hybridized for 16 h at 45°C on the GeneChip Human Genome U133 Plus 2.0 Array (Thermo Fisher Scientific, Inc.). Arrays were washed and stained in the GeneChip Fluidics Station 450 (Affymetrix) and then scanned using GeneChip Scanner 3000 7G. Single Array Analysis was calculated by Microarray Suite version 5.0 (MAS5.0) with Affymetrix default settings and global scaling as the normalization method. The trimmed mean target intensity of each array was arbitrarily set to 500. Principal component analysis (PCA) and unsupervised hierarchical clustering were performed on samples based on normalized expression of all genes (total of 54,613 ID genes).
Flow cytometric analysis
Suspended single cl-iSCs, hMSCs, and hNSPCs were labeled with antibodies against CD19, CD44, CD45, CD90, CD105, CD166 (SC017; R&D Systems, Minneapolis, MN), and CD133 (Proteintech, Rosemont, IL) according to the manufacturer's instructions. Cell subpopulations were quantified using a fluorescence-activated cell sorter (FACS, BD LSRFortessaÔX-20; BD Pharmingen) as described [14].
Electrophysiology by multielectrode arrays
Electrophysiological properties were analyzed using multielectrode arrays (MEA2100 Lite, Multi Channel Systems MCS GmbH, Reutlingen, Germany) as described [13,20,21]. In brief, cl-iSCs (80 μL of a 1 × 105 cells/mL suspension) were plated onto 60 electrode dishes (60MEA200/30) precoated with Matrigel™ (Corning). During experimental recordings, the medium, including growth factors, was changed every 3 days. The spontaneous activity was recorded by the MEA2100-Lite system (Multi Channel Systems). The electrophysiological signals were detected using MC_Rack software (Multi Channel Systems), and the data were digitized with a 60-channel A/D converter at rate of 20 kHz with 16-bit resolution. Data obtained from multielectrode were converted to Axon Binary File format and analyzed using LabChart 8.0 Japanese software (AD Instruments Japan, Inc., Nagoya, Japan).
Analysis of neurogenic potential by single cell from cl-iSCs
Individual clone cells were isolated by the limited dilution method as described [22]. In brief, single-cell suspensions of cl-iSCs (passage 3) were plated on 96-well plates (1 cell per well) in DMEM/F12 containing bFGF, EGF, 10% FBS, and N2. The medium was changed every 2 days. On day 26 after incubation, expanded cl-iSCs were treated with trypsin and reseeded under the same conditions. For neurogenic differentiation, iSCs expanded from single cells were further incubated in a new medium containing DMEM/F12 plus bFGF, LIF, 10% FBS, and N2 and then examined by immunohistochemistry or multielectrode arrays as described above.
Statistical analysis
Data are presented as mean ± standard deviation. Differences were analyzed using Student's t-test. A gene was considered differentially expressed when its adjusted P value was lower than 0.05 (P < 0.05).
Results
Clinical history of two patients with cerebellar infarction undergoing decompressive lobectomy
The first case (Case 1, Supplementary Table S2) was an 84-year-old female hospitalized at the cardiovascular division of our hospital for postoperative management of percutaneous coronary intervention (PCI). On day 6 after PCI, she suddenly complained of nausea and consciousness disturbance. She had paraparesis, and her NIH stroke scale score was 39 (severe). Diffusion-weighted magnetic resonance imaging (MRI) showed a large early hyperintensity in the basilar artery territory (Fig. 1A), and magnetic resonance angiography (MRA) showed basilar artery occlusion (Fig. 1B). Given the risk of mortality, she received mechanical thrombectomy. Although reperfusion was achieved, there was no improvement of consciousness disturbance. On the day following thrombectomy, computed tomography (CT) showed upward tentorial herniation (Fig. 1C). She received internal and external decompression 36 h after ischemic onset (day 6). Postoperative CT showed successful evacuation of the infarct areas (Fig. 1D).
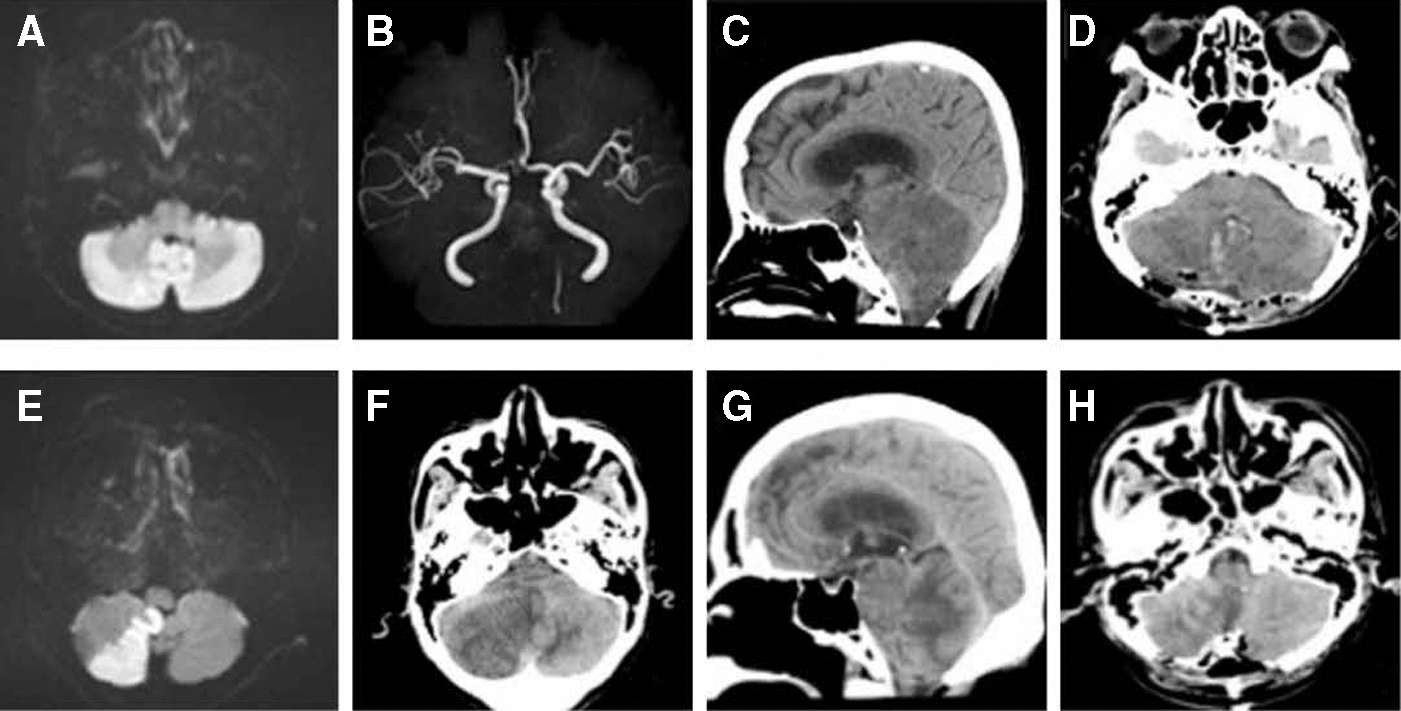
Neuroimaging of two patients with cerebellar infarction. Diffusion-weighted MRI
The second case (Case 2, Supplementary Table S2) was an 80-year-old male who visited his doctor due to gait ataxia. Diffusion-weighted MRI showed right cerebellar infarction (Fig. 1E). Thus, he was immediately transferred to our hospital. We started medical treatment, but on day 5 after admission, his consciousness got worse. Head CT revealed a large low-density area in the right cerebellar hemisphere with brainstem compression and hydrocephalus (Fig. 1F, G). Emergent decompressive craniectomy and partial lobectomy were performed 5 days after stroke onset. Postoperative CT confirmed successful decompressive lobectomy (Fig. 1H).
Histological examination of poststroke cerebellar tissue sections
We performed histological examination of poststroke brain tissue sections obtained from the first patient (Fig. 2A). H&E staining revealed numerous pyknotic cell nuclei indicative of apoptosis/necrosis in the cerebellar cortex, including the molecular, Purkinje cell, and granular layers (Fig. 2B). No mature neural cells, including neurons [Tuj1+ cells (Fig. 2C) and MAP2+ cells (Fig. 2D)] and ACs [GFAP+ cells (Fig. 2E) and S100β+ cells (Fig. 2F)], were found in the molecular layer of the cerebellar cortex. These findings suggest that most of neurons and ACs had already disappeared due to apoptosis/necrosis. This is also consistent with previous findings showing that under pathological conditions, such as after ischemic stroke, mature neural cells within ischemic areas undergo cell death within a couple of days [14,23].

Histological findings of poststroke human cerebellum obtained from the first patient. Hematoxylin and eosin staining of surgically excised tissue
Using a mouse model of cerebral infarction, we and others previously identified putative iSCs within ischemic areas [9 –13]. Very recently, we also demonstrated putative iSCs in the human ischemic cerebrum [14]. These findings led us to hypothesize that putative iSCs might also be present in the human ischemic cerebellum. Cerebral iSCs expressed the NSPC marker nestin [9 –13], so we investigated the presence of nestin+ cells in the ischemic cerebellum of the first patient. Immunohistochemistry revealed nestin+ cells in the molecular layer of the cerebellar cortex (Fig. 2G).
Our previous studies showed that nestin+ iSCs are likely derived from brain PCs following ischemia (ischemic pericytes; iPCs) , so we further investigated the localization of nestin+ cells. Immunohistochemistry revealed the presence of nestin+ cells near vWF+ ECs (Fig. 2H) in the molecular layer of the cerebellar cortex. In addition, nestin+ cells in the molecular and granular layers of the cerebellar cortex coexpressed the pericytic markers NG2 (Fig. 2I–K) and αSMA (Fig. 2L–N), respectively. Similar histological characteristics were observed in the ischemic cerebellum tissue from the second patient (Supplementary Fig. S1A–D).
Characterization of cerebellar stem cell origin and marker phenotype
We further investigated the phenotypic characteristics of these nestin+ cells by polymerase chain reaction (PCR) and immunocytochemistry. Ischemic brain tissue obtained from the first patient was mechanically dissociated into single cells by passage through needles, and these dissociated cells were incubated as adherent monolayer cultures under conditions promoting iSC proliferation. Adherent cells (Fig. 3A) were highly proliferative, and immunohistochemistry revealed expression of the dividing cell marker Ki67 (Fig. 3B) and nestin (Fig. 3C). These cells also expressed pericytic markers, including αSMA (Fig. 3D–F), NG2 (Fig. 3G–I), and PDGFRβ (Fig. 3J–L), results confirmed by PCR (Fig. 3P).
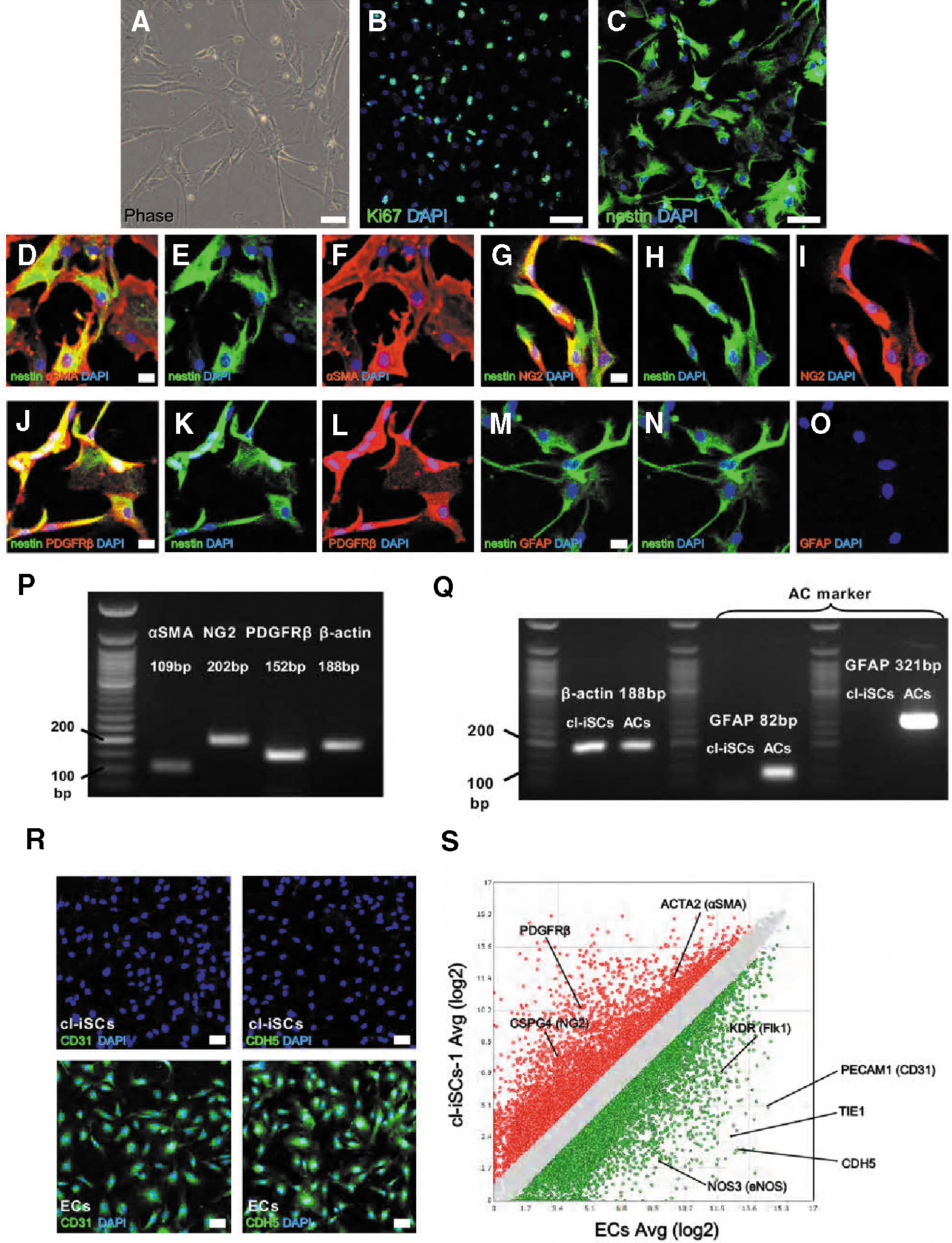
Isolation and characterization of human cl-iSCs from the first patient. Immunohistochemistry showed that putative cl-iSCs
To exclude the possibility that these nestin+ cells are reactive ACs [24], we further investigated whether the nestin+ cells expressed the astrocytic marker GFAP but found no such expression (Fig. 3M–O). Furthermore, no GFAP mRNA expression was detected by PCR using two different GFAP primers, although GFAP mRNA expression was detectable in commercially obtained ACs using the same PCR methods (Fig. 3Q). In addition, heat mapping and scatter plot analysis showed strong expression of astrocytic markers GFAP, S100β, aquaporin4 (AQP4), and FABP7 in ACs compared to cl-iSCs (Supplementary Fig. S2A–D). These results strongly indicate that these nestin+ cells are not of the astrocytic lineage.
We then examined whether putative cl-iSCs contain EC populations, because it has been reported that nestin+ cells can also originate from vascular lineage cells, including ECs [25,26]. Immunohistochemistry revealed no detectable expression of EC markers, such as CD31 and CDH5, by cl-iSCs, although expression of these markers was observed in human ECs (ECs) used as a positive control (Fig. 3R). Scatter plot analysis comparing global gene expression profiles between cl-iSCs-1 and ECs revealed stronger expression of pericytic-related genes [eg, ACTA2 (αSMA), CSPG4 (NG2), PDGFRβ] in cl-iSCs (>2-fold higher than in ECs, red) and stronger expression of EC-related genes [eg, PECAM1 (CD31), CDH5, KDR (Flk1), TIE1, and NOS3 (eNOS)] in ECs (>2-fold higher than in cl-iSCs, green) (Fig. 3S). Similar lineage traits were confirmed in putative cl-iSCs obtained from the second patient (cl-iSCs-2) (Supplementary Fig. S3A–T).
Multipotency of cerebellar iSCs and differentiation potential to electrophysiologically active neurons
We next investigated whether cl-iSCs have multipotency. They displayed a high proliferation rate as demonstrated by a doubling time of 65.8 ± 6.3 h (Fig. 4A). In addition, cl-iSCs maintained expression of various stem cell and undifferentiated cell markers even after several passages, including nestin, c-myc, Klf4, and Sox2 (Fig. 4B). This finding suggests that cellular phenotypes regarding stemness are not variable during in vitro treatment (eg, repeated passages, chemical stimulation, and so on), at least under the tested conditions. To confirm this multipotency, cl-iSCs were incubated separately under osteoblastic, adipogenic, and chondrocytic differentiation conditions. Under these respective conditions, cl-iSCs differentiated into mesodermal lineages, including osteocalcin+ osteoblasts (Fig. 4C), FABP4+ and Oil red O+ adipocytes (Figs. 4D, E), and aggrecan+ chondrocytes (Fig. 4F).
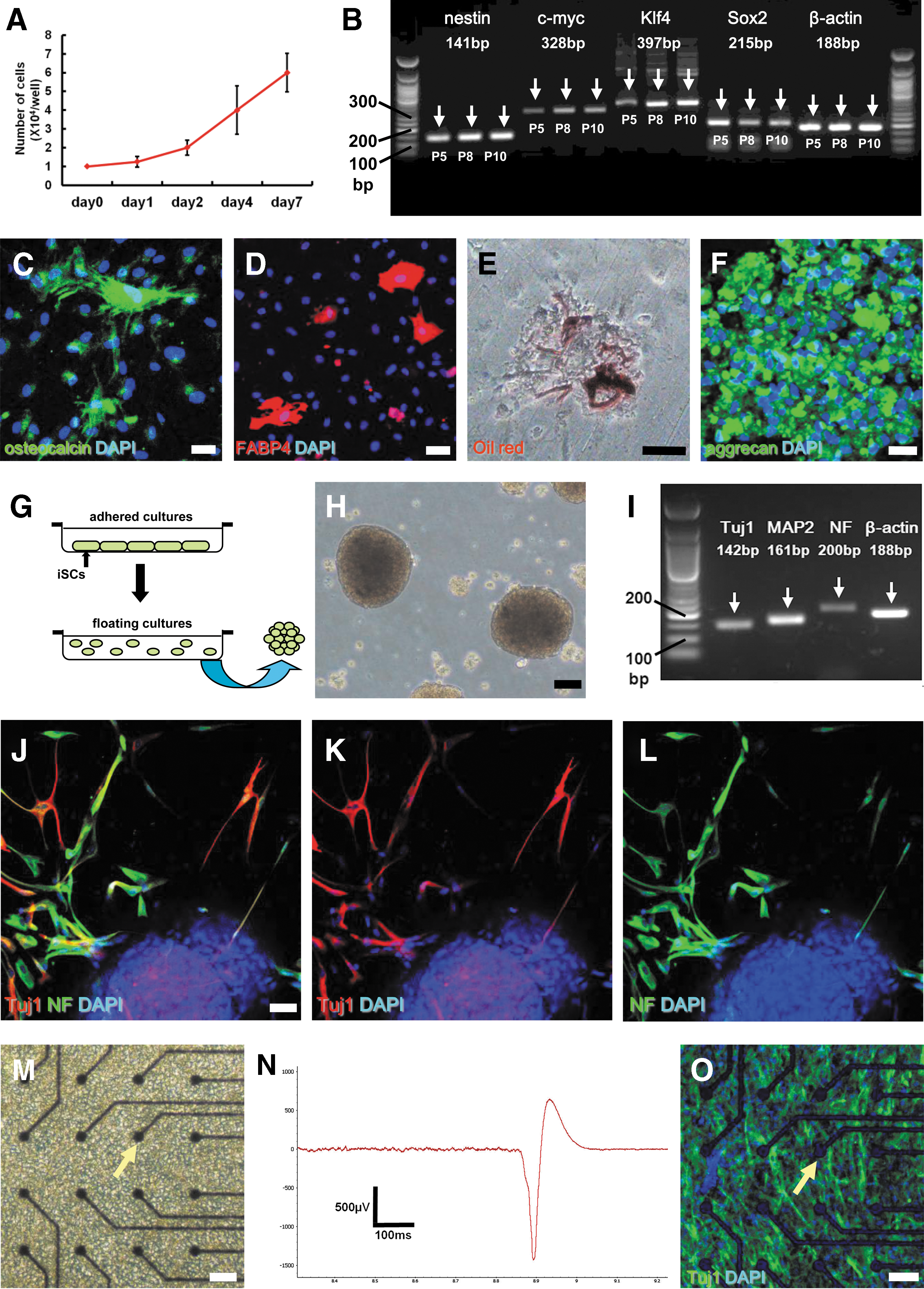
Multipotent stem cell activity of human cl-iSCs obtained from the first patient. Putative cl-iSCs demonstrated high proliferative potential
To investigate whether cl-iSCs can also differentiate into neuronal lineages, adherent cells were incubated in floating cultures under conditions that promote the formation of neurosphere-like cell clusters (Fig. 4G) [9,10,27,28]. On day 4 of incubation, cell clusters were observed (Fig. 4H), and PCR analysis revealed expression of the neuronal markers Tuj1, NF, and MAP2 (Fig. 4I). Immunohistochemistry also showed that these clusters produced Tuj1+ and NF+ neuronal cells (Fig. 4J–L). Furthermore, electrophysiological recordings using multielectrode arrays (Fig. 4M) showed that cl-iSCs exhibited the potential to differentiate into electrophysiologically active neurons (Fig. 4N). In addition, the presence of Tuj1+ neuronal cells on microelectrode dishes was confirmed by immunohistochemistry (Fig. 4O).
To exclude possible contamination by mature neurons in cl-iSC populations, single-cell suspensions made from cl-iSCs were diluted in medium and plated in 96-well plates as described [22]. One day after plating, wells successfully plated (1 cell per well) were marked and incubated for further analysis (Fig. 5A). Single cells gradually expanded under conditions that promote the proliferation of cl-iSCs (Fig. 5B). Then, the capacity of expanded cl-iSCs to produce functional neurons was investigated by further incubation under conditions that promote differentiation to neuronal lineages. Indeed, these cells expressed neuronal markers, including Tuj1, NF, and MAP2 (Fig. 5C–H). In addition, these cells were electrically functional (Fig. 5I–K). Collectively, these results confirm that cl-iSCs can give rise to functional neurons. Similar multipotency was confirmed in cl-iSCs obtained from the second patient (cl-iSCs-2) (Supplementary Fig. S4A–O), and multielectrode arrays revealed stimulus-evoked responses (Supplementary Fig. S5).
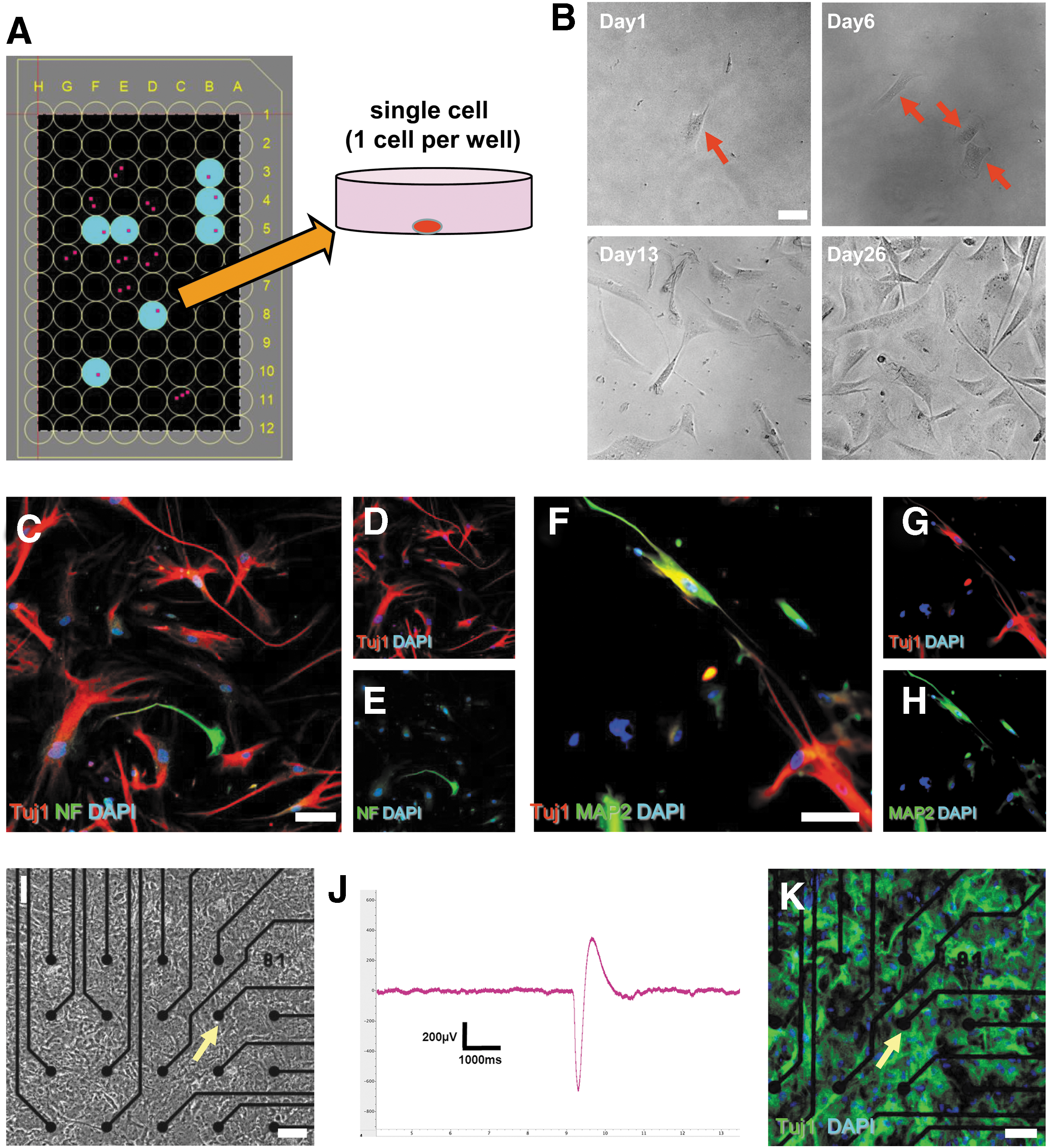
Neurogenic potential of single-cell suspensions from cl-iSCs. Using the limited dilution assay, single-cell suspensions obtained from cl-iSCs were plated in 96-well plates
Comparison of cerebellar iSCs to other types of cells, including stem cells
Our data thus far demonstrate that cl-iSCs can differentiate into both mesodermal and neural lineage cells. Thus, we next compared the traits of cl-iSCs to MSCs and NSPCs that mainly commit to the mesoderm and neural lineage, respectively. FACS analysis revealed that cl-iSCs-1 (Fig. 6A) and cl-iSCs-2 (Fig. 6B) rarely expressed hematopoietic lineage markers (CD19 and CD45) but did express various MSC markers (CD44, CD90, CD105, and CD166), similar to established MSCs (Fig. 6C). These findings are consistent with our previous results showing that cr-iSCs share several markers with MSCs [14]. NSPCs also express MSC markers [29]. However, the NSPC marker CD133 [30] was specifically expressed in established NSPCs but not cl-iSCs and MSCs (Fig. 6D).
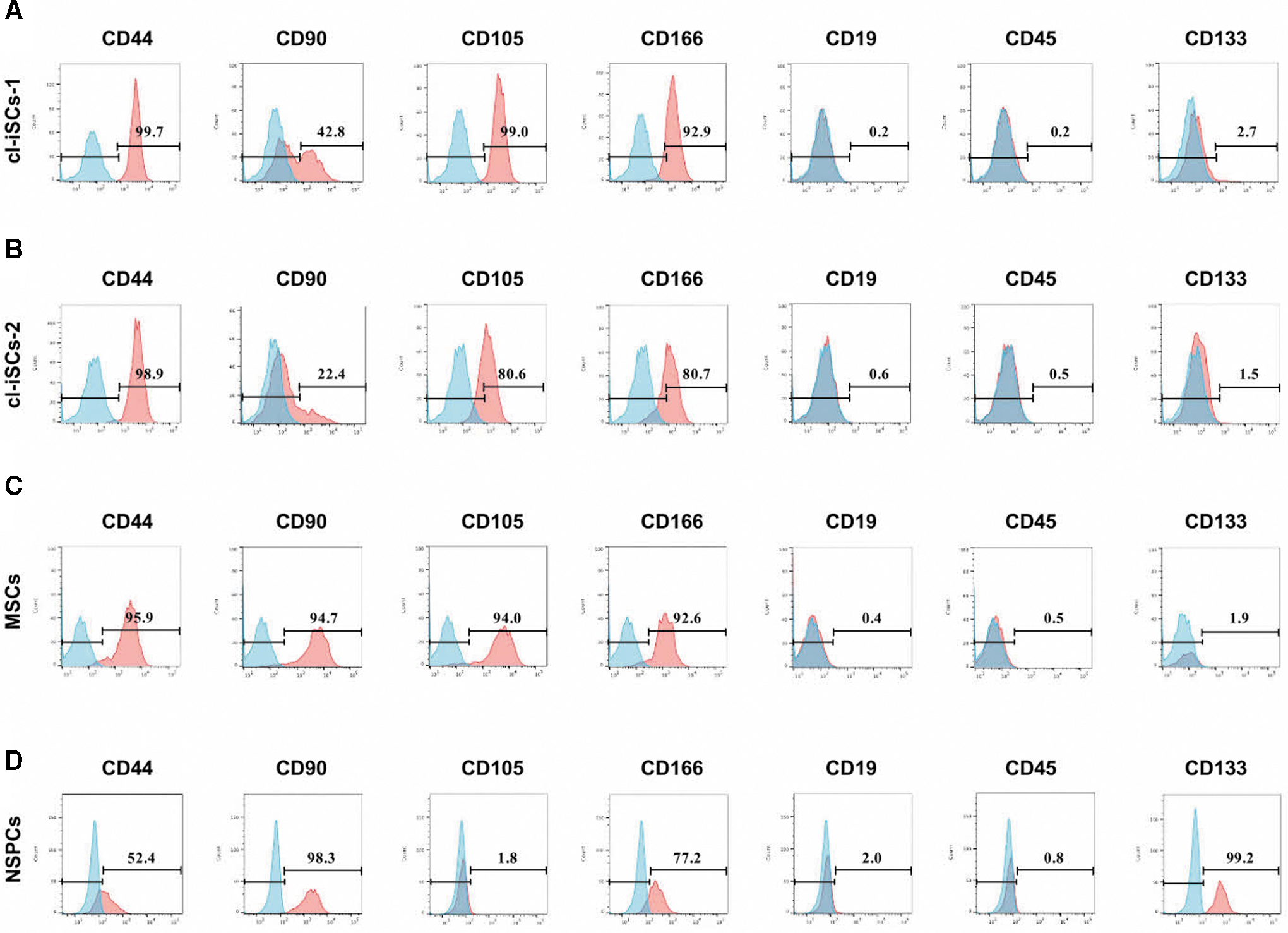
Flow cytometric analysis of human cl-iSCs
To further investigate the traits of cl-iSCs, global gene expression profiles were analyzed in cl-iSCs obtained from both patients. In scatter plot analysis comparing global gene expression profiles between cl-iSCs-1 and MSCs [log2 (cl-iSCs-1/MSCs ratio)], ∼74% of all gene sequences were with the difference range ≥−2-fold to ≤2-fold change) [gray areas in Fig. 7A (left panel) and Supplementary Fig. S6A]. In the scatter plot analysis comparing global gene expression profiles between cl-iSCs-1 and NSPCs [log2 (cl-iSCs-1/MSCs ratio)], ∼57% of total gene sequences were present in this range [gray areas in Fig. 7B (left panel) and Supplementary Fig. S7A]. Similarly, in global gene expression profile comparison between cl-iSCs-2 and MSCs [log2 (cl-iSCs-2/MSCs ratio)], ∼74% of sequences were within this range [gray areas in Fig. 7A (middle panel) and Supplementary Fig. S6B]. Moreover, in the comparison between combined cl-iSCs from both patients (cl-iSCs-1,2) and MSCs [log2 (cl-iSCs-1,2/MSCs ratio)], 77% of total gene sequences were present within this range [gray areas in Fig. 7A (right panel) and Supplementary Fig. S6C]. In the comparison of global gene expression profiles between cl-iSCs-2 and NSPCs [log2 (cl-iSCs-2/NSPCs ratio)] and between cl-iSCs-1,2 and NSPCs [log2 (cl-iSCs-1,2/NSPCs ratio)], ∼57% [gray areas in Fig. 7B (middle panel) and Supplementary Fig. S7B] and 59% of total gene sequences, respectively, were within the ≥−2-fold and ≤2-fold change range [gray areas in Fig. 7B (right panel) and Supplementary Fig. S7C]. These results indicate that the global gene expression pattern of cl-iSCs is closer to that of MSCs than NSPCs.
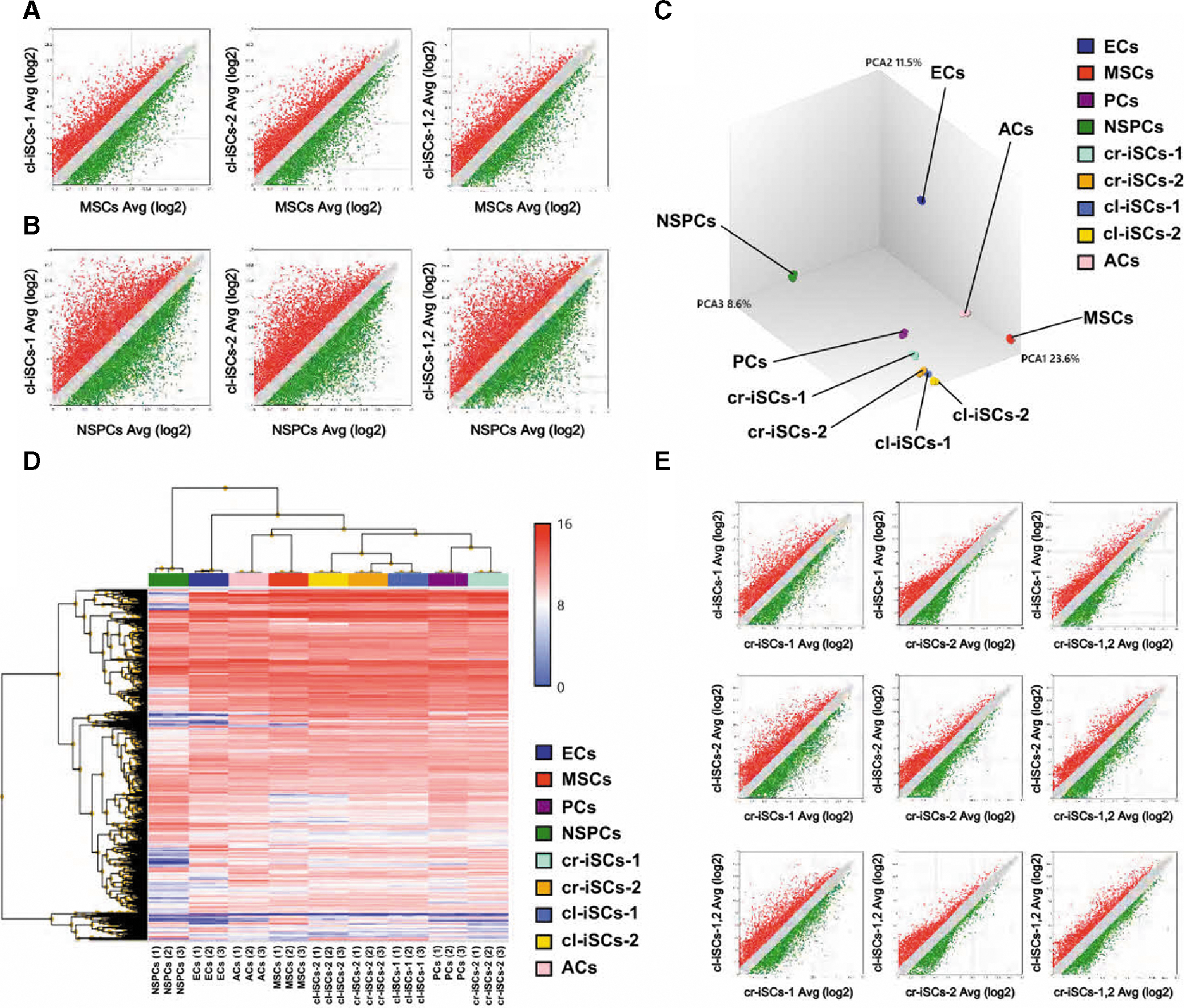
Global gene expression profiles of human cl-iSCs. Scatter plot analysis comparing global gene expression profiles between cl-iSCs and MSCs
While NSPCs are neural plate derivatives, certain brain PCs are neural crest derivatives [31]. Thus, we further compared the expression levels of genes related to neural plate or neural crest origin between NSPCs and cl-iSCs-1,2 [log2 (cl-iSCs-1,2/NSPCs ratio)]. Expression levels of numerous neural plate-related genes (early neural genes) were lower in cl-iSCs-1,2 than NSPCs (<−2-fold change) (Supplementary Table S3), whereas many neural crest-related genes were higher in cl-iSCs-1,2 than NSPCs (>2-fold change) (Supplementary Table S4).
We further compared global gene expression patterns between cl-iSCs and cr-iSCs obtained from two different cerebral stroke patients (cr-iSCs-1, cr-iSCs-2) [14]. PCA mapping (Fig. 7C) and heat mapping (Fig. 7D) showed that the gene profiles of cl-iSCs and cr-iSCs were closer to PCs than to ECs, MSCs, NSPCs, and ACs. These data indicate that iSCs, which are likely brain PC derivatives, are present in poststroke cerebellum, as well as cerebrum.
In addition, scatter plot analysis of each pairwise comparison (cl-iSCs-1 vs. cr-iSCs-1; cl-iSCs-1 vs. cr-iSCs-2; cl-iSCs-1,2 vs. cr-iSCs-1; cl-iSCs-2 vs. cr-iSCs-1; cl-iSCs-2 vs. cr-iSCs-2; cl-iSCs-2 vs. cr-iSCs-1,2; cl-iSCs-1,2 vs. cr-iSCs-1; cl-iSCs-1,2 vs. cr-iSCs-2, cl-iSCs-1,2 vs. cr-iSCs-1,2) showed that most genes (∼74%–86%) were restricted within the −2-fold ≥ and ≤2-fold change range (Fig. 7E, Supplementary Fig. S8A–I), indicating a strong lineage resemblance between cl-iSCs and cr-iSCs.
However, PCs in forebrain regions are reported to be neural crest derivatives, while PCs in caudal regions are considered to be mesoderm derivatives [32,33]. Thus, we compared the neural crest-related gene expression levels between cl-iSCs-1,2 and cr-iSCs-1,2 [log2 (cl-iSCs-1,2/cr-iSCs-1,2 ratio)] (Supplementary Table S5). Expression levels of ERBB3, TFAP2A, ETS1, and BGN were significantly lower in cl-iSCs than cr-iSCs, while Sox9, ID1, ID2, MGP, and FOXD3 expression levels were significantly higher in cl-iSCs than cr-iSCs. However, expression levels of most neural crest-related genes were within the −2-fold ≥ and ≤2-fold change range. Thus, we further examined the expression levels of cerebellar specific genes (Supplementary Table S6) and found that of the genes exhibiting a >1.5-fold change [log2 (cl-iSCs-1,2/cr-iSCs-1,2 ratio)], the cerebellar granule cell marker PDEIC was significantly higher in cl-iSCs-1,2 than cr-iSCs-1,2 (P < 0.05). These results indicate that cl-iSCs resemble cr-iSCs but possess cerebellar specific traits.
Discussion
This study demonstrates for the first time that putative iSCs are present in poststroke human cerebellum. There is strong evidence that a stem cell population exists in the mouse cerebellum [16,17], although the precise characteristics remain unclear. In addition, nestin+ stem/progenitor cells have been identified in the developing mouse cerebellum [34,35] and found to give rise to neurons and glia following injury [35]. Consistent with these reports, our current study revealed that nestin+ iSCs can be isolated from poststroke adult human cerebellum, will follow neural lineages under appropriate conditions, and give rise to electrophysiologically active neuron-like cells.
It is well known that NSPCs reside in specific regions of adult mammalian central nervous system (CNS) (so-called neurogenic regions), such as the subventricular zone (SVZ) and the subgranular zone (SGZ) of the hippocampus [36 –38]. Our current study showed that iSCs can also be isolated from the mature human cerebellum. Cerebellum is distant from the SVZ and SGZ, so these cl-iSCs are likely a completely independent stem cell population. In support of this idea, cl-iSCs and NSPCs exhibited distinct gene expression profiles, and cl-iSCs expressed fewer early neural plate markers than NSPCs.
Although the precise origin of cl-iSCs remains unclear, we and other researchers previously demonstrated that stem/progenitor cells originate, in part, from leptomeninges covering the surface of the whole CNS, including the brain and spinal cord [10,39 –48]. Although leptomeninges are considered to be histologically continuous with PCs in the parenchyma of the brain along the blood vessels [10,49], the present study revealed that gene expression patterns were very similar among cl-iSCs, cr-iSCs, and brain PCs. This suggests that cl-iSCs are brain PC derivatives. These results also support previous findings showing that iSCs developing within poststroke mouse [9 –12] and human cerebrum [14] are likely PC derivatives. Although the precise relationship between leptomeninges and PCs remains unclear, there is increasing evidence that nestin+ cells, which contain stem cell populations with neurogenic potential, are present within leptomeninges of the cerebrum [10,39,42 –46,48], as well as the cerebellum [50]. Thus, cerebellar leptomeninges are also a potential origin of cl-iSCs. Nevertheless, the precise origin of cl-iSCs warrants further investigation.
It has been reported that PCs in various organs can function as multipotent stem cells [51 –59]. However, our previous study of mice at different developmental stages, including embryonic, postnatal, and adult, showed that nestin+ brain PCs gradually decreased in number during the postnatal period and were eventually lost in adulthood [60]. This suggests that adult brain PCs have somatic cell rather than stem cell traits, although it appears that brain PCs maintain stem cell-like features up to the early developmental period. However, we have shown that brain PCs within poststroke areas reexpress nestin and reacquire stemness while brain PCs within nonischemic areas do not [9 –11].
In the present study, the gene expression profile of patient-derived iSCs resembled that of commercially available human brain PCs, including high expression of nestin (data not shown), indicating that brain PCs maintain stem cell features. Considering that the commercial human brain PCs used in this study originated from fetal brains and that brain PCs retain stemness during early development [60], their stem-like features are not surprising. Taken together, these results suggest that adult brain PCs reacquire the stemness traits of fetal brain PCs under ischemia. Although the precise mechanisms by which they acquired such features remain unclear, we previously showed that adult mouse and human brain PCs reacquired stemness presumably through cellular reprogramming [9,14]. Similar to our findings, other groups have demonstrated that adult human brain PCs can be reprogrammed into neuronal cells through unfolding of a neural stem cell-like program [61,62], indicating that adult brain PCs have phenotypic plasticity.
Several types of cells, including ECs, MSCs, NSPCs, ACs, and PCs, were used in the present study. Similar to iSCs, it was methodologically ideal to use all cell types obtained from adulthood. Unfortunately some cell types were not commercially available from adulthood. Similar to the traits of PCs, the traits of brain ECs are known to differ across developmental stages [63]. Thus, further comparative studies using cell samples obtained from same developmental state are needed.
Unfortunately, there are few appropriate mouse models of cerebellar infarction, so we could not confirm that cl-iSCs are also present in adult mouse cerebellar. Due to ethical limitations, it is also unclear whether cl-iSCs are present within healthy adult human cerebellum. However, our finding that nestin+ cl-iSCs develop within postischemic areas even in aged human brain indicates that adult human brain PCs can function as stem cells under pathological conditions, such as after ischemic stroke, as is the case with adult mouse brain PCs following cerebral infarction [9 –12].
In the present study, we found that cl-iSCs express various markers in common with MSCs, including CD44, CD90, CD105, and CD106, and that they can differentiate along various mesoderm lineages, including osteoblast, adipocyte, and chondrocyte. Akin to PCs, MSCs are multipotent stem cells that reside in the perivascular regions of multiple organs. The precise lineage and functional relationships between PCs and MSCs remain unclear. However, because PCs and MSCs have similar markers and both demonstrate multipotency [64 –69], certain MSCs are thought to be derived from PCs [70 –72]. Thus, it is reasonable that cl-iSCs and MSCs have similar markers and traits.
In the present study, we compared the traits of BM-derived MSCs with cl-iSCs. However, recent studies showed that brain also harbors perivascular MSCs that possess traits similar to PCs [64,67]. In addition, it is reported that MSC traits vary among organs [68]. Thus, compared to BM-derived MSCs, brain MSCs may show a greater phenotypic similarity with cl-iSCs, although the precise relationships among brain MSCs, brain PCs, and iSCs should be clarified in further studies.
Brain PCs can differentiate into various cells, including neurons [9,11,51,73]. In the present study, we also showed that cl-iSCs, which likely originate from brain PCs, have the potential to differentiate into electrophysiologically active neurons. Although the mechanisms by which brain PCs differentiate into neurons remain unclear, we and others have demonstrated that adult brain PCs can acquire NSPC-like features through cellular reprogramming by upregulated expression of transcriptions factors associated with the neuronal phenotype, such as Sox2 and ASCL1 [9,14,61,62].
During early brain development, neuroepithelial cells generate NSPCs that differentiate into various types of neural cells, including neurons. Then, some neuroepithelial cells form the neural crest, where cells attain complex features of neural and mesenchymal lineages through epithelial–mesenchymal transition [74]. Certain brain PCs, particularly in forebrain regions, are reported to originate from the neural crest, while PCs in other brain regions are mesoderm derivatives [32,33]. In the present study, we found that many neural crest-related genes were higher in iSCs than in commercial NSPCs. Unexpectedly, however, expression levels of these genes did not differ markedly between cr-iSCs and cl-iSCs. Although the detailed phenotypic differences between cr-iSCs and cl-iSCs should be clarified in future studies using larger sample sizes, these results suggest that like forebrain PCs, cerebellar PCs originate, at least in part, from the neural crest. Considering that the neural crest can function as a source of multipotent neural crest stem cells that can differentiate into various neural cells, including neurons [75 –81], cl-iSCs may also give rise to neurons. However, the precise origin of cerebellar PCs should be clarified in future studies.
Cerebellum is composed of several phenotypically distinct types of neurons (eg, Purkinje cells, granule cells, Golgi cells, and basket cells). A previous study reported that cerebellar stem cells possess intrinsic regional characteristics and produce cerebellar-specific neurons, such as calbindin-expressing neurons with Purkinje cell morphology [16]. Although we did not obtain evidence that cl-iSCs differentiate into calbindin+ neurons (data not shown), cl-iSCs strongly expressed cerebellar granule cell markers compared to cr-iSCs. It is reported that granule cells are derived from precursors in the cerebrum rhombic lip [82], while Purkinje cells originate from cerebellar ventricular zone progenitors [83]. In accord with these findings, we have previously shown that cr-iSCs are derived from stem cell populations at the brain surface, including leptomeninges that contain abundant PCs rather than the SVZ [10,40]. In addition to PC, several other CNS cell types reside in the perivascular region, including radial glia-like cells [84], oligodendrocyte precursor cells [85,86], and perivascular fibroblasts [87 –89]. Although the precise traits of these cells in cerebellum remain unclear, they possess similar markers and traits as cr-iSCs [31]. Thus, the precise origin, traits, and subtypes of cerebellar stem cells should be determined in future experiments.
In conclusion, we show that iSCs are present in the human cerebellum following stroke. These cl-iSCs differentiated into electrophysiologically functional neurons, suggesting that the factors/molecules modulating iSCs fate (eg, survival, proliferation, differentiation, and migration) may be novel therapies to promote neurogenesis following ischemic stroke. Alternatively, cl-iSCs could be used as autologous stem cells for transplantation in patients following severe ischemic stroke because cl-iSCs can be rapidly expanded in vitro while maintaining stemness. A more complete understanding of cerebellar iSCs is required for realization of poststroke restoration of cerebellar function.
Footnotes
Acknowledgments
This work was partially supported by Japan Society for the Promotion of Science (JSPS) KAKENHI (15K06723), a Strategic Program Grant for Research Institute Development in Private Institute from Ministry of Education, Culture, Sports, Science and Technology (MEXT) in Japan, Japan Agency for Medical Research and Development (AMED), Grant-in-Aid for Graduate Student, Hyogo College of Medicine (2018), and Grant-in-Aid from the Japanese Society of Cerebral Blood Flow and Metabolism. The authors thank S. Kamachi, M. Shirakawa, K. Uchida, H. Kageyama, and Y. Minato for helpful assistance.
Author Disclosure Statement
Department of therapeutic progress in brain diseases is financially supported by Daiichi Sankyo Co., Ltd., Nippon Zoki Pharmaceutical Co., Ltd., and CLEA Japan, Inc. The sponsors had no roles in this study, including those of study design, data collection, data analysis, data interpretation, and article writing.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.