
Abstract
Organoids and cells generated in vitro from pluripotent stem cells (PSCs) are considered to be robust models of development and a conceivable source of transplants for putative cell therapy. However, a fundamental question about organoids and cells generated from PSCs is as follows: do they faithfully reproduce the in vivo tissue they are supposed to mimic and replace? This question is particularly relevant to complex tissues such as the cerebral cortex. In this review, we have tackled this issue by comparing cerebral cortices generated in vitro from PSCs to the in vivo cortex, with a particular focus on their respective cellular composition, molecular and epigenetic signatures, and brain connectivity. In short, in vitro cortex generated from PSCs reproduces most of the cardinal features of the in vivo cortex, including temporal corticogenesis and connectivity when PSC-derived cortical cells are grafted in recipient mouse cortex. However, compared to in vivo cortex, in vitro cortex lacks microglia and blood vessels and is less mature. Recent experiments show that the brain of the transplanted host provides these missing cell types together with an environment that promotes the synaptic maturation of the cortical transplant. Taken together, these data suggest that corticogenesis is largely intrinsic and well recapitulated in vitro, while the full maturation of cortical cells requires additional environmental clues. Finally, we propose some lines of work to improve corticogenesis from PSCs as a tool to model corticogenesis and rebuild cortical circuits.
Introduction
Organoids and cells generated in vitro from pluripotent stem cells (PSCs) are considered to be robust models of development and a conceivable source of transplants for cell therapy. However, a fundamental question about organoids and cells generated from PSCs is as follows: do they faithfully reproduce the in vivo tissues? This question is particularly relevant to complex tissues such as the cerebral cortex, which has a six-layered architecture and contains myriad cell types [1]. The vast repertoire of cortical cells is generated during embryogenesis and the perinatal period [1,2]. By contrast, there is probably no neurogenesis in the adult cortex [3 –6]. Because no endogenous neurogenesis could compensate for neuronal loss due to disease or injury, dead cortical neurons must be replaced by artificial means. There are two main strategies for cortical neuron replacement. The first strategy is to produce novel cortical neurons in situ, which can be achieved by activating dormant neural progenitors [7], by stimulating neurogenesis from radial glia-like precursors of neighboring meninges [8], or by transdifferentiating cells into the desired cortical cell type [9]. The second strategy to replace dead neurons is to transplant exogenous cortical cells that have been produced in vitro from PSCs.
A decade ago, the Vanderhaeghen [10] and Sasai [11] laboratories reported the successful generation of cortical-like cells from mouse embryonic stem cells (mESCs). Both laboratories grafted mESC-derived cortical cells into a recipient mouse cortex, paving the way to cortical cell therapy. These pioneer studies were further supported by the generation of cortical-like cells from human ESCs and induced PSCs (hiPSCs) [11 –19]. Corticogenesis from PSCs seems also an interesting system to model corticogenesis in vivo, particularly for modeling human diseases. There is a continuously growing number of protocols to generate cortical-like cells from PSCs, from the initial protocols using mESCs [10,11] to the recently reported direct conversion of human fibroblasts into cortical-like embryoids [12]. Most of these protocols produce cells that recapitulate several features of an in vivo neocortex, including sophisticated features such as cortical connections following their transplantation in mouse cortex. Hereafter, we call “in vitro cortex” PSC-derived cells that reproduce at least one feature of the in vivo cortex.
In this review, we evaluate to what extent in vitro cortex is similar to in vivo cortex in terms of cellular composition, molecular and epigenetic signatures, and connectivity. In short, in vitro cortex generated from PSCs reproduces most of the cardinal features of the in vivo cortex. This includes temporal corticogenesis as well as connectivity when grafted in recipient mouse cortex. However, compared to the in vivo cortex, in vitro cortex lacks microglia and blood vessels and is less mature. Recent experiments show that the host brain provides these missing cell types, together with an environment that promotes the synaptic maturation of the cortical transplant. In summary, this suggests that corticogenesis is intrinsic and well recapitulated in vitro, while the full maturation of cortical cells requires additional environmental clues. Finally, we propose additional lines of work to improve corticogenesis from PSCs as a tool to model corticogenesis and for putative cortical cell therapy.
Reproduction of the Molecular Signature, Cellular Composition, and Cytoarchitecture of In Vivo Cortex by Cortex Generated from PSCs
The cellular composition of the cerebral cortex in vivo was recently revisited using single-cell RNA-seq (Sc-RNA-seq). The developing mouse cortex contains 90% of neural cells (progenitors, excitatory neurons, interneurons, astrocytes, and oligodendrocytes) and 10% of non-neural cells (microglia and endothelial cells) [20]. The adult mouse cortex comprises the same classes of cells, but in different proportions [21,22]. Sc-RNA-seq analysis has revealed that the human developing cortex in vivo also contains excitatory and inhibitory neurons, oligodendrocyte precursors, neural progenitors, astrocytes, and microglia [23]. However, in contrast with the mouse cortex, no endothelial cells were retrieved in the human developing cortex [23].
In vitro cortex generated from human or mouse PSCs largely reproduces the in vivo catalog of cells. As in vivo, glutamatergic neurons are the major cell type in vitro, followed by interneurons [10,15 –17,24], astrocytes [10,13,16], and oligodendrocytes [25]. These data show that corticogenesis from PSCs reproduces the whole neural lineage of in vivo corticogenesis [1,2]. By contrast, microglia and blood vessels were not reported to be present in vitro. The absence of microglia is not surprising since microglia are non-neural cells that originate from erythromyeloid precursors generated in the yolk sac [26]. The absence of blood vessels is more open to discussion since bipotent neurovascular progenitors might exist in vivo [8,27,28]. Conversely, there might be undesired, noncortical cells depending on the protocol of in vitro corticogenesis, on the PSC line [29,30], or on the laboratory: different laboratories using the same PSC lines and differentiation protocol obtain substantial different cell compositions [31]. Sc-RNA-seq is a powerful method to uncover these undesired cells [30] and should also help refining corticogenesis protocols to increase reproducibility.
Special attention was paid to characterize PSC-derived cortical-like neurons at the molecular level. In vitro neurons express the typical molecular markers of the six cortical layers [10,11,13,15 –17,24] (Fig. 1A). Using single-cell transcriptomics, it was found that 68% of hiPSC-derived cortical-like cells express at least one canonical marker of cortical layers [32]. In addition, some neurons co-express deep- and upper-layer markers, as in the human fetal brain [32]. However, upper-layer neurons are underrepresented in mESC-derived cells compared to in vivo [10]. A remarkable feature that is well reproduced by in vitro corticogenesis is temporal corticogenesis (Fig. 1B). In vitro, cortical cells appear sequentially with the same order than during in vivo corticogenesis [2,33], from early multipotent neural progenitors to fate-restricted neurons [10,11,13,15 –17,24] (Fig. 1B). Thus, corticogenesis can take place without influence of the entire brain environment, indicating that it is mainly an intrinsic mechanism [34]. In vitro corticogenesis also reproduces properly the timing of corticogenesis in a species-specific manner (around 21 days to complete corticogenesis for mouse ESCs [10,11], 2 months for nonhuman primates PSCs [35], and 3 months for human PSCs [16,24,35]).

Main characteristics of in vivo cortex that are reproduced by in vitro cortex generated from PSCs. (
In vivo, proper neural activity in the neocortex requires a balanced excitation/inhibition [36]. This suggests that to maintain neural activity, the ESC-derived cortical transplant must contain a defined proportion of excitatory and inhibitory neurons. In vivo, excitatory and inhibitory neurons are known to originate from the dorsal and ventral telencephalons, respectively, and ventral identity is induced by Sonic hedgehog [37]. In vitro, acting on the sonic hedgehog pathway when PSC-derived cells are bipotent dorsal/ventral telencephalic progenitors alters the glutamatergic/interneuron ratio [11,19,38 –40]. Another possibility is to assemble different proportions of dorsal and ventral forebrain spheroids [41]. Finally, in grafting experiments, interneurons of the host invade the cortical transplant generated from hiPSCs [42]. We speculate that the host adjusts the number of interneurons to maintain a balanced neural activity.
In vivo, the cerebral cortex has a complex three-dimensional (3D) laminar architecture with six neuronal layers. In vitro, 3D cortex generated using Matrigel, a reconstituted basement membrane matrix, recapitulates this complex 3D organization [11,13,19,35,43,44] (Fig. 1C). In vitro, 3D cortex notably reproduces the stack of cell types of the in vivo cortex with, from bottom to the top of the pile: progenitor (ventricular, subventricular, and intermediate) and neuronal zones (deep-layer and upper-layer neurons, and Cajal-Retzius cells) (Fig. 1C). As in vivo, neuronal layers of in vitro 3D cortex are generated sequentially in an inside-out manner [11,13,19,35] (Fig. 1C). A marked advantage of 3D cortex over two-dimensional (2D) cortex is its ability to model human diseases characterized by disrupted architecture of the neocortex [44]. For instance, Zika virus infection reduces the ventricular thickness of cortical organoids generated from human PSCs [45], which suggests a causative link between Zika infection and the microcephaly observed in Zika-infected human fetuses. On the other hand, 2D corticogenesis is likely more reproducible and scalable for screening purposes than the 3D system. Both 2D and 3D cortices have been successfully transplanted in mouse cortex [10,16,46 –50], but which system is most successful is unknown.
In vivo, glutamatergic cortical neurons orientate and migrate radially on radial glia scaffolds [51,52]. Cortical cells differentiated from mESCs [10] or from hPSCs [17] orientate radially in slice overlay assays (Fig. 1D). In 3D cortex derived from hPSCs, there is a radial glia-like supported migration [53]. Furthermore, interkinetic nuclear migration, the basal/apical oscillations of progenitor nuclei occurring as cell cycle progresses, is preserved in organoids [13]. In addition, the typical migration of interneurons into the dorsal telencephalon is reproduced by coculturing dorsal and ventral forebrain spheroids, with interneurons of the ventral spheroid invading the dorsal spheroid [41]. Taken together, these experiments show that the modes of neural migration occurring during in vivo corticogenesis are well recapitulated in vitro.
Future Prospects to Improve the Molecular Signature, Cellular Composition, and Cytoarchitecture of In Vitro Cortex
In vivo, the extracellular matrix (ECM) acts as a scaffold and regulates cell adhesion, migration, and intercellular communications. ECM orchestrates tissue morphogenesis and influences cell fate [54,55]. ECM is involved in brain development [56,57] and some specific ECM constituents such as REELIN are crucial for human corticogenesis [58]. Recently, a screening approach identified three ECM components that are sufficient to cause the folding of human cortical slices in culture [59]. Taken together, these data suggest that in vitro corticogenesis could be improved by using selected ECM components of the in vivo cortex (in preference to Matrigel that is made from a carcinoma). First, this requires to determine the respective matrisomes (ECM and ECM-associated proteins) of in vitro and in vivo cortices. To our knowledge, there is no study that compares them at the protein level. However, many transcriptomic data are available. The transcriptomes of mouse and human cortices generated in vivo [60], or in vitro [15,29,30,61], are all enriched in Gene Ontology terms for ECM components. In addition, human apical progenitors generated either in vivo or in vitro have similar expression profiles for 16 out of 18 ECM genes [30]. By contrast, Luo and coworkers reported that ECM components are among the most differentially expressed genes between human in vivo and in vitro cortices [61]. Thus, to which extent in vitro cortex reproduces the ECM composition of the in vivo cortex is still elusive and further studies are required to identify which ECM component(s) could improve in vitro corticogenesis.
To further improve in vitro corticogenesis, a research area complementary to the ECM is scaffolding. Scaffolding consists in assembling polymers to mimic the architecture of the selected organ. A floating scaffold made of microfilaments of poly(Lactide-co-Glycolide) copolymers (PLGA) improves corticogenesis and cytoarchitecture of 3D cortex from hPSCs [62] (Fig. 1E).
In vivo, the neocortex contains numerous areas that are associated with specific functions (such as object recognition and motion perception for the visual cortex [63]). The area identity of in vitro cortices can be inferred from their axonal connections observed after their transplantation in vivo. There are many reports on PSC-derived cortical transplants with connections that are typical of visual/limbic cortex [10,16,47,48,64], few reports on PSC-derived cortex displaying motor identity [49,65], and, to our knowledge, no report on other cortical areas. It is likely that future cortical therapies and developmental studies will have needs of additional cortical areas. Similarly, we could imagine that a stroke selectively destroys a given cortical layer. To date, there is no protocol to specifically generate a given cortical layer from PSCs, but there is some evidence that the identity of cortical neurons can be switched in vivo. Indeed, overexpressing Fezf2 in vivo, either in layer IV neurons [66] or in layer II/III neurons [67] switches their identity to a layer V identity.
Finally, in vitro cortex is less mature than in vivo cortex. The current consensus is that neural cells produced from hPSCs resemble a human brain at mid-gestation [16,30,43,49]. Human iPSC-derived cortex acquires over time a transcriptomic signature gradually resembling that of the brain of the autopsy donor [68]. However, transcriptomic signatures of in vitro and in vivo cortices never fully match [68]. Corticogenesis is intrinsic [10,16,34] and we speculate that maturation of in vitro cortex depends on environmental clues. It is well known that the environment influences brain development [69]. The environmental clues missing in vitro could be other cell types (such as microglia and blood vessels), other neighboring brain areas that connect and develop the concert with cortex (such as the thalamus [70]), and/or circulating factors. The synapses of in vitro cortex generated from hPSCs continue to mature following their transplantation in mouse cortex [42]. To which degree the maturity of the transplanted cortex compares to that of the cortex maintained in vitro is unknown. This can be tested by transplanting human PSC-derived cells in a mouse cortex and comparing its transcriptome after its development/maturation in vivo to the transcriptome of parallel cultures developed in vitro.
Reproduction of the Epigenetic Signature of In Vivo Cortex by Cortex Generated from PSCs
In vivo, epigenetics modifications, including DNA methylation, post-translational modifications of histones, and remodeling of chromatin, are crucial to cortex development and function [71 –74]. Mutations in the epigenetic machinery lead to neurodevelopmental diseases [75]. The aberrant gain or loss of histone marks is a hallmark of neurodegenerative diseases [76]. As culturing cells in vitro can alter their epigenetic profiles [77 –79], it is important to compare the epigenetic signatures of cortex generated in vitro from PSCs to that of in vivo cortex.
The DNA methylomes of human cerebral organoids and human fetal brain are largely identical for CpG methylation at regulatory sequences and for non-CpG methylation (an epigenetic mark enriched in the brain [80]) [61] (Fig. 1F). However, organoids contain aberrant hypomethylation at pericentromeric regions [61]. Genomic imprinting is an epigenetic mechanism where different epigenetic marks are present on parental alleles at a few loci, leading to the parent-of-origin depend expression of imprinted genes. In vitro cortex from mESCs largely reproduces parental genomic imprinting of the developing mouse cortex [29] (Fig. 1F). Fourteen out of 18 differentially methylated regions (DMRs) are identically methylated between in vivo and in vitro cortices [29]. Zdbf2-DMR is the most differentially methylated DMR, with a gain of DNA methylation on the maternal allele in vitro [29]. Importantly, ZDBF2-DMR hypermethylation is also observed in certain patients with Beckwith-Wiedemann syndrome [81], an imprinting disorder characterized by overgrowth and predisposition to develop tumors. Thus, grafting an in vitro cortex with this aberrant methylation might be detrimental to the host.
During in vivo development, chromatin is reorganized so that selected genomic regions get into contact, while others get apart (Fig. 1G). This 3D reorganization of chromatin is thought to be important for the regulation of gene expression and thus, cell fate [82]. Using ultra-deep Hi-C (to quantify the level of interaction between two genomic regions at a global level), RNA-seq (to quantify gene expression), and ChIP-seq (to quantify histone marks), it was found that some trends of spatial genome architecture of mouse cortex are well reproduced during in vitro corticogenesis from mESCs [83].
Future Prospects to Improve the Epigenetic Signature of In Vitro Cortex
Taken together, these studies suggest that some epigenetic features of in vivo cortex are well recapitulated in vitro. Nevertheless, the failure to mimic certain epigenetic features cannot be neglected, notably in the prospect of cortical cell therapy. Overall, it is important to note that we have only a partial knowledge on the epigenetic signature of the in vitro cortex. The Roadmap Epigenomics Consortium has generated a comprehensive repertoire of histone modification patterns, DNA accessibility, DNA methylation, and RNA expression on 111 human tissues, including dorsolateral prefrontal cortex and cortical neurospheres derived from hESCs [84]. These data could be mined to compare the epigenetic signatures of in vivo and in vitro human cortices.
How to improve the epigenetic signature of the in vitro cortex? Improving the cell culture media of PSCs is an interesting avenue of research as media formulation has a considerable impact on the epigenetic signature of PSCs [85]. Among media components, Vitamin C profoundly remodels the epigenome of PSCs by Tet-dependent DNA demethylation [86]. Treating mESC-derived cortical cells with valproate, a histone deacetylase inhibitor, increases the number of upper-layer cortical neurons during mESC corticogenesis [87]. However, valproate is not specific of a given DNA loci and it deacetylates histones in a genome-wide manner. To increase target selectivity, a given aberrant epigenetic mark could be edited using CRISPR with a dead Cas9 (dCas9) fused to an epigenetic modifier brought to the genomic locus of interest by a matching sgRNA guide [88,89]. This strategy has been successfully applied to Fragile X syndrome (FXS). In FXS, the FRM1 gene is silenced by hypermethylation. Tet1 fused to dCas9 efficiently demethylates the promoter of FRM1, rescuing FRM1 expression in neurons derived from FXS hiPSCs [90].
Reproduction of Connectivity of the In Vivo Cortex: Insights from Transplantation Experiments of Cortex Generated from PSCs
In vivo, the connections of cortical neurons with different brain areas, including the cortex itself, are highly specific [33,91,92]. In vitro cortex connects with typical cortical targets when it is transplanted in the cortex of living rodents at several stages: embryonic [10], newborn [10,16,46], and adult [42,47 –50,64,65,93] (Fig. 2). The axons of grafted mouse and human PSC-derived neurons connect with cortical targets, including at long distance [10,16,46 –50] (Fig. 2A). Transplanted neurons establish synapses with the host (Fig. 2B). Interestingly, successful connectivity requires an identity match between the cortical areas of the host and the in vitro transplant [47,48,65] (Fig. 2C). Their synapses mature over time and are selectively conserved, or eliminated [42]. Grafted neurons are electrophysiologically active [16,47,48,93]. There is experimental evidence for functional connectivity between the transplant and the host [48,93,94] (Fig. 2D). Interestingly, when in vitro cortex is transplanted in mouse brain, it is invaded by the host microglia [42,93,95] (Fig. 2E). In addition, the transplant also gets vascularized by the host [42,93] (Fig. 2E). Thus, for putative cortical therapy, it is likely not crucial to bioengineer an in vitro cortex with blood vessels and microglia as these two cell types may be provided by the host following transplantation. Taken together, these data suggest that PSC-derived cortical cells could be used to reconstruct cortical tracts in the future.
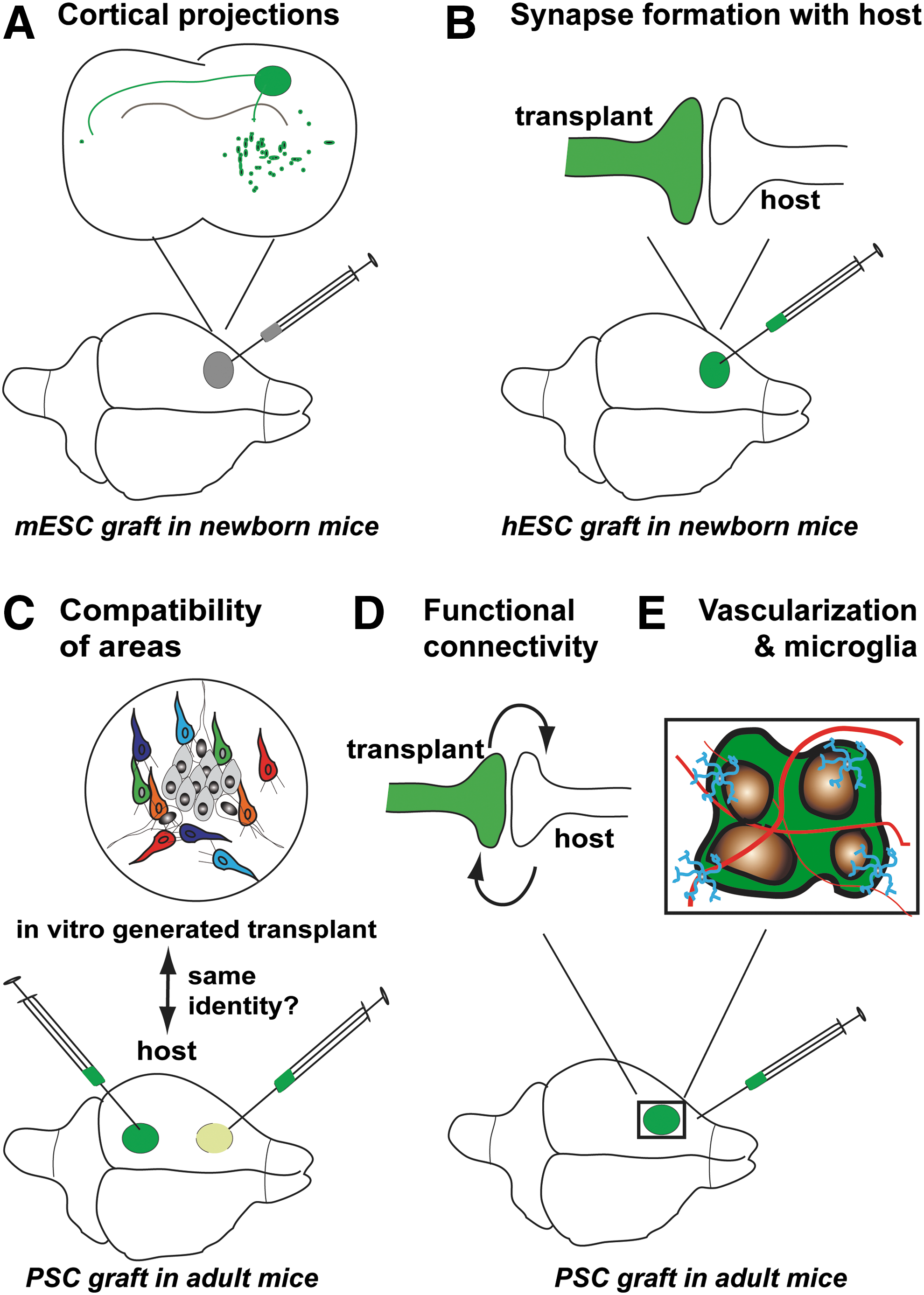
Reproduction of connectivity of the in vivo cortex by in vitro cortex following its transplantation in rodent cortex.
In addition, the transplantation of cortex generated from human PSCs gives insight into human diseases characterized by impaired neural connectivity and function. When hESC-derived neurons are grafted in Alzheimer disease mice, human neurons accumulate synaptic markers around Aβ plaques and they degenerate with no observable tangle pathology [95]. Grafted neurons derived from Down syndrome hiPSCs have increased synaptic stability and their neural networks are less active than control human neurons [42].
Future Prospects to Improve Transplantation of PSC-Derived Cortical Cells
The functional benefit of cortical transplants generated from PSCs is largely unknown. One study has shown that transplanted hiPSC-derived cortical progenitors improve the motor function of rats after focal ischemia in the motor cortex [50]. In this study, the readout of the motor function (forelimb asymmetry test) was quite easily quantifiable. However, for other cortical areas, it might be more difficult to quantity an improvement in the associated cortical functions.
An additional issue is xenotransplantation. To our knowledge, all grafting experiments of in vitro cortex from human PSCs have been performed in rodents, whose cortices are far less complex than primate cortices. The validity of the rodent cortex as a model for transplantation may depend on the cortical area to be repaired. For instance, the mouse visual cortex seems an appropriate model to extrapolate to the primate cortex because the flow of visual information is processed essentially by similar mechanisms in mouse and primates [96,97]. However, it is possible that the full maturation of cortical cells differentiated from primate PSCs will require the specific environment of the primate brain.
The benefit of transplanting PSC-derived tissue might also be limited by the presence of undifferentiated or poorly differentiated PSCs that could form teratomas in the host brain. In this context, it was reported that a high concentration of L-Alanine selectively eliminates undifferentiated hiPSCs [98]. Alternatively, hiPSCs expressing an inducible Caspase-9 might be an interesting tool to induce the selective death of the transplant in case of adverse effects [99]. Transplantation experiments are also limited by histocompatibility. Immunosuppression of mice is required to prevent rejection of hPSC cortical grafts beyond 2 months of transplantation [16]. Accordingly, transplantations of hPSC-derived cortex are performed in immunocompromised mice [16,42,93]. In the future, allografts will therefore require treating the patient with immunosuppressors. Transplantation of cortical-like cells generated from iPSCs of the receiving patient might be a solution to this histocompatibility concern.
Conclusions
To conclude, considerable efforts have been made to characterize in vitro cortex from PSCs, notably on its cellular composition, molecular and epigenetic signatures, and brain connectivity. In vitro cortex reproduces most of the characteristics of the in vivo cerebral cortex. Thus, in vitro cortex is likely a robust model to study corticogenesis and to test drugs. In vitro cortex is also a conceivable source of neural transplants for future cortical repair. However, compared to in vivo cortex, in vitro cortex lacks microglia and blood vessels and is less mature. Interestingly, recent experiments suggest that the brain of the transplanted host provides these missing cell types together with an environment that promotes the synaptic maturation of the cortical transplant. To date, there is no evidence that putative cortical cell therapy will require grafting the whole repertoire of cortical cells. By contrast, modeling in vivo corticogenesis likely requires reproducing precisely the cell type composition of the in vivo developing cortex, as the outcome and interpretation of in vitro corticogenesis experiments might be biased by the absence or disproportion of cell types normally present in the developing cortex. For instance, corticogenesis is known to be influenced by microglia [26,100] and blood vessels [28], which are both absent in vitro.
Taken together, this suggests that corticogenesis is intrinsic and is quite well recapitulated in vitro. By contrast, the full maturation of cortical cells requires additional environmental clues. These environmental clues remain to be identified and will certainly improve the generation of in vitro cortex from pluripotent stem cells.
Footnotes
Acknowledgment
We thank Joël Bockaert for helpful discussions and critical reading of our review.
Author Disclosure Statement
The authors declare no conflicts of interest.