
Abstract
Cardioprotective effects of exosomes derived from human umbilical cord mesenchymal stem cells (hucMSC-exosomes) postmyocardial infarction (post-MI) have been reported in our previous study. It is known that fibroblasts are pro-inflammatory phenotypes, while myofibroblasts are anti-inflammatory phenotypes. This study aimed to investigate whether hucMSC-exosomes promoted cardiac fibroblast-to-myofibroblast differentiation in inflammatory environments and protected cardiomyocytes. Rats were performed by permanent ligation of the left anterior descending coronary artery and underwent intramyocardial injection of hucMSC-exosomes or phosphate-buffered saline (PBS) in surgery. Fibroblasts were stimulated by lipopolysaccharide (LPS) to create inflammatory environments in vitro. Western blot and immunohistochemical and immunofluorescence staining for α-smooth muscle actin were used to demonstrate fibroblast-to-myofibroblast differentiation. Transwell migration assay and CCK-8 assay were used to evaluate migration and proliferation of fibroblasts. Reverse transcription–polymerase chain reaction, western blot, and immunohistochemical staining were used to detect expressions of inflammatory factors. To investigate cardioprotective effects, cardiomyocytes were treated with supernatant derived from fibroblasts pretreated with LPS or LPS plus hucMSC-exosomes in hypoxic environments. Cardiomyocyte apoptosis was determined using TUNEL assay and western blot. Results indicated that hucMSC-exosomes increased the density of myofibroblasts in infarct areas during inflammatory phases post-MI, promoted fibroblast-to-myofibroblast differentiation in inflammatory environments, and attenuated inflammatory responses in vitro and in vivo. Culture medium derived from fibroblasts pretreated with LPS plus hucMSC-exosomes reduced cardiomyocyte apoptosis. In vivo, apoptotic cells in acute myocardial infarction (AMI)+exosomes groups were also less than AMI+PBS groups. In conclusion, hucMSC-exosomes can promote fibroblast-to-myofibroblast differentiation in inflammatory environments, then protecting cardiomyocytes.
Introduction
Acute myocardial infarction (AMI) is the foremost cause of death in human cardiovascular disease. Cardiomyocytes develop irreversible necrosis due to blood supply deficiency after myocardial infarction (MI). Since adult cardiomyocytes are unable to proliferate, the acute loss of myocardium is replaced with a fibrotic scar [1,2]. Fibroblasts are critical in the formation of fibrotic scar [3]. In addition, fibroblasts are involved in different phases of cardiac repair and play different roles in these phases. Cardiac repair response post-MI is mainly divided into the following three stages: the inflammatory phase, the proliferative phase, and the maturation phase. Fibroblasts exhibit different phenotypes at different phases post-MI. During inflammatory phases, fibroblasts are activated and acquire pro-inflammatory phenotypes, which can release inflammatory mediators to promote inflammatory responses. Matrix-degrading activity of fibroblasts is also activated, which degrade extracellular matrix components and maybe lead to cardiac rupture. In addition, cardiac fibroblast-to-myofibroblast differentiation is inhibited during the inflammatory phase. When dead cells are removed, cardiac repair enters proliferative phases. At this phase, the inflammatory response is suppressed and most fibroblasts differentiate into myofibroblasts, which are characterized by anti-inflammatory effect, extracellular matrix production, and contractile ability. Myofibroblasts secrete extracellular matrix proteins, making infarct areas filled with collagen-rich scar to maintain heart structure and functional integrity. Myofibroblasts also secrete some anti-inflammatory factors to attenuate the inflammatory response, which is beneficial for cardiac repair post-MI [4 –8].
It is reported that mesenchymal stem cells (MSCs) or exosomes derived from MSCs have cardioprotective effects [9 –12]. Our research team has previously reported that exosomes derived from human umbilical cord mesenchymal stem cells (hucMSC-exosomes) have cardioprotective effects [13 –15]. Cardioprotective effects are associated with inhibition of excessive inflammation and attenuation of cardiomyocyte apoptosis [16 –18]. Myofibroblasts have the anti-inflammatory characteristic. Therefore, myofibroblasts increased in infarct areas during the inflammatory phase post-MI may be beneficial for cardiac repair post-MI. In this study, we focus on whether hucMSC-exosomes can promote pro-inflammatory fibroblasts to differentiate into anti-inflammatory myofibroblasts in the inflammatory environment to protect cardiomyocytes. Rats underwent permanent ligation of the left anterior descending (LAD) coronary artery, and hucMSC-exosomes were intramyocardially administrated in surgery. In vitro, we used lipopolysaccharide (LPS) to create an inflammatory environment. Results demonstrated that hucMSC-exosomes increased the density of myofibroblasts in infarct areas during the inflammatory phase post-MI and promoted fibroblasts to differentiate into myofibroblasts in the inflammatory environment in vitro. Then, the inflammatory response was attenuated, and cardiomyocyte apoptosis was reduced. Our study suggests that hucMSC-exosomes may exert cardioprotective effect through promoting cardiac fibroblast-to-myofibroblast differentiation in an inflammatory environment.
Materials and Methods
Animals
All animal experiments were performed with the approval of Animal Experimental Center of Jiangsu University, Zhenjiang, China and maintained in accordance with institutional policies. Animals used in the current study were male Sprague-Dawley (SD) rats. SD rats were 4–5 weeks of age with body weight of 180–220 g. Neonatal cardiac fibroblasts and cardiomyocytes were isolated from the heart ventricles of 1- to 3-day-old SD rats.
Cell culture
hucMSCs were isolated and cultured according to the established method [19]. hucMSCs were obtained from The Affiliated Hospital of Jiangsu University (Jiangsu, China), and all people were provided informed consent for the use of the cord in this study. hucMSCs were cultured in low-glucose Dulbecco's modified Eagle's medium (L-DMEM; Gibco, Grand Island) with 10% fetal bovine serum (FBS; Biological Industries, Israel) at 37°C in 5% CO2. The morphology of primary and the second passage hucMSCs was photographed. Neonatal cardiac fibroblasts and cardiomyocytes were isolated from 1- to 3-day-old rats. The ventricles were minced and digested using trypsin and collagenase II (Worthington, Lakewood). According to different attachment time that most fibroblasts are attached in 90 min, the digested cells were plated on dishes and incubated for 90 min at 37°C in 5% CO2. Adherent cells were harvested as cardiac fibroblasts, while nonadherent cells were harvested as cardiomyocytes. Primary fibroblasts and cardiomyocytes were used in experiments.
Extraction and characterization of hucMSC-exosomes
When the density of cultured hucMSCs was up to 80%, hucMSCs were cultured with 10% exosome-free FBS L-DMEM. Forty-eight hours later, we collected the supernatant of hucMSCs. The supernatant was centrifuged at 300 g (10 min at 4°C), 2,000 g (20 min at 4°C), and 10,000 g (30 min at 4°C) to remove cell debris. Then, the supernatant was concentrated using a 100-kDa molecular weight cutoff (MWCO) hollow fiber membrane (Millipore, MA) at 1,000 g for 30 min. The final concentrate was mixed with ExoQuick-TC exosome isolation reagent (System Biosciences, CA) overnight at 4°C. Next day, mixtures were centrifuged at 1,500 g (30 min at 4°C). Finally, the extracted exosomes were suspended in 1 mL of phosphate-buffered saline (PBS) and stored at −80°C. The protein concentration of exosomes was determined using a BCA Kit (CWBIO, Beijing, China).
To characterize hucMSC-exosomes, a transmission electron microscope was used to observe the morphology of exosomes. Nanoparticle tracking analysis (NTA) was performed to analyze the particle size and video image of exosomes. Western blot was used to detect surface markers of exosomes, such as CD9 and CD63.
AMI model
Male SD rats were randomly divided into three groups as follows: sham groups, AMI+PBS groups, and AMI+exosomes groups. All animals underwent permanent LAD coronary artery ligation according to the reported method, except for sham groups [15]. In surgery, rats underwent intramyocardial administration and received either hucMSC-exosomes (400 μg) or PBS control. Following AMI, rats were continuously housed. These rats were sacrificed at day 2, 4, and 7 after surgery.
Adipogenic and osteogenic differentiation
HucMSCs were plated in 24-well culture plates. HucMSCs were cultured in adipogenic or osteogenic differentiation medium (Biological Industries) and then replaced adipogenic or osteogenic differentiation medium every 3 days. Ten days later, hucMSCs were stained by corresponding adipo-staining kit or osteo-staining kit (VivaCell Biosciences, Shanghai, China) to detect the adipogenic and osteogenic potential of hucMSCs.
Flow cytometry
HucMSCs were grown in six-well culture plates. When the density of cultured hucMSCs was up to 80%–90%, cells were digested. Cells were incubated with monoclonal antibodies against CD19, CD34, CD45 (FITC-conjugated), CD29, CD90, and CD105 (PE-conjugated; eBioscience) for 30 min at 4°C, then washed twice. Finally, cells were resuspended in PBS and immediately analyzed with a flow cytometer (Becton Dickinson).
Exosome labeling and internalization
HucMSC-exosomes were labeled with CellTracker™ CM-Dil (Invitrogen, CA), according to the manufacturer's protocol. The labeled exosomes were washed by PBS and filtered through a 100-kDa MWCO hollow fiber membrane (Millipore) to remove unbound dye. CM-Dil also did the same treatment and was used as a negative control. Fibroblasts were grown on coverslips (NEST Biotechnology, Wuxi, China). Next day, cells were incubated with labeled exosomes at 37°C for 12 h and then washed with PBS and fixed in 4% paraformaldehyde. The nuclei were stained with Hoechst (Sigma). Images were acquired with a fluorescence microscope (Olympus DP73, Japan).
Immunohistochemistry
Two days, 4 days, and 1 week post-MI, hearts of rats were excised and then fixed in 4% paraformaldehyde. Next, hearts were embedded in paraffin and chopped in serial 3-μm thick slices. For immunohistochemical staining, slices were incubated with primary antibodies: anti–α-Smooth muscle actin (α-SMA; Maxim, Fuzhou, China), anti-periostin (Proteintech, Wuhan, China), anti-collagen I (Boster, Wuhan, China), interleukin (IL)-1β (Bioworld Technology), tumor necrosis factor (TNF)-α (Bioworld Technology), and transforming growth factor (TGF)-β (Abcam, Britain) overnight at 4°C and an HRP-conjugated secondary antibody (Maxim) in the next processing. Immunohistochemical images were captured using Pannoramic MIDI (3DHISTECH, Hungary).
Reverse transcription–polymerase chain reaction
Primary fibroblasts treated with LPS (100 ng/mL) (ABCONE, Shanghai, China) for 6, 12, 24, and 48 h and treated with LPS (100 ng/mL) (ABCONE) or LPS plus hucMSC-exosomes (200 μg/mL) for 12 and 24 h were collected. Infarct areas of heart tissues were collected. Total RNA was extracted using TRIzol reagent (Invitrogen). Complementary DNA was obtained using Reverse Transcription–Polymerase Chain Reaction (RT-PCR) Kits (CWBIO). Real-time PCR was accomplished using UltraSYBR Mixture (CWBIO). The sequences for sense and antisense primers are as follows: IL-1β (forward: 5′-ATCTCACAGCAGCATCTCGACAAG-3′, reverse: 5′-CACACTAGCAGGTCG TCATCATCC-3′); TNF-α (forward: 5′-GCATGATCCGAGATGTGG AACTGG-3′, reverse: 5′-CGCCACGAGCAGGAATGAGAAG-3′); and TGF-β (forward: 5′-GGCACCATCCATGACATGAACCG-3′, reverse: 5′-GCCGTAC ACAGCAGTTCTTCTCTG-3′). Levels of gene expression were normalized to β-actin (forward: 5′-TGTCACCAACTGGGACGATA-3′, reverse: 5′-GGGGTGTTGAAGGTCTCA AA-3′).
Immunofluorescence assay
Fibroblasts were grown on coverslips (NEST Biotechnology, Wuxi, China). Next day, fibroblasts were treated with LPS or LPS plus hucMSC-exosomes for 12, 24, 48, and 72 h. Control groups did not add any stimulus. After that, cells were washed with PBS and fixed for 20 min with 4% paraformaldehyde at room temperature and then washed and permeabilized with 0.1% Triton X-100 for 10 min. Next, they were blocked with 5% bovine serum albumin for 20 min and incubated with primary antibodies for α-SMA (1:100; Boster, Wuhan, China) overnight at 4°C. Next day, cells were incubated with fluorescence-conjugated secondary antibodies (1:1,000; Cell Signaling Technology, Boston), followed by Hoechst (Sigma). Images were obtained by a fluorescence microscope (Olympus DP73).
Transwell migration assay
Cardiac fibroblast migration was determined using Transwell chambers (Corning). Primary fibroblasts were treated with LPS or LPS plus hucMSC-exosomes for 24 h and then harvested and resuspended in serum-free L-DMEM, respectively. Two hundred microliters of cell suspension (3 × 104 cells) per group was added to the upper chamber of the insert. Six hundred microliters of 10% FBS L-DMEM were added to lower chambers. After incubation for 8 h at 37°C in 5% CO2, cells that had migrated through membranes were stained using 0.1% crystal violet. Migrated cells were visualized and photographed using a stereo microscope (Olympus DP73). Three randomly selected fields were photographed, and cells were counted.
Cell proliferation assay
Proliferation of cardiac fibroblasts was assessed using CCK-8 assay (Beyotime Biotechnology, China). Briefly, cells were harvested and resuspended in 10% FBS L-DMEM. One hundred microliters of cell suspension (3,000 cells) per group was plated in 96-well culture plates. Next day, fibroblasts were treated with LPS or LPS plus hucMSC-exosomes. Control groups were cultured with 10% FBS L-DMEM without additional stimulus. Twelve and 24 h later, medium was replaced with 100 μL serum-free L-DMEM, and 10 μL of CCK-8 reagent was added. Two hours later, we measured the absorbance at a wavelength of 450 nm with a microplate spectrophotometer (BioTek, Vermont).
Collagen gel contraction assay
Fibroblasts were harvested and resuspended in serum-free L-DMEM to achieve the density of 2.5 × 105 cells/mL. Collagen matrix was prepared by mixing 8 mL collagen I (3 mg/mL; Gibco, Grand Island) with 1 mL 10 × DMEM (Gibco) and 1 mL 0.1 M NaOH to neutralize the pH on ice. Cell suspension and collagen matrix were mixed in a ratio of 1:1. Subsequently, 500 μL of this mixture was plated in 24-well culture plates and incubated at 37°C for 1 h. Gels were detached and cultured in 10% FBS L-DMEM containing LPS or LPS plus hucMSC-exosomes. Control groups were only cultured in 10% FBS L-DMEM. After 24 h, pictures of gels were taken, and diameters of floating gels were measured.
Hypoxic experiments in vitro
Cardiomyocytes were grown on coverslips or six-well culture plates. Next day, cardiomyocytes were treated with supernatant that fibroblasts were continuously cultured with 10% FBS L-DMEM for 12 and 24 h after withdrawal of LPS or LPS plus hucMSC-exosomes and then incubated in humidified air with 5% CO2, 2% O2, and 93% N2 at 37°C. At the same time, hypoxic control group was cultured with 0.2% FBS L-DMEM in same hypoxic condition, and normal control group was cultured with 10% FBS H-DMEM in humidified air with 5% CO2.
TUNEL staining
The apoptosis of cardiomyocytes was measured by the TUNEL assay (Boster) according to the manufacturer's instructions.
Western blot analysis
Protein samples were extracted from hucMSC-exosomes, cells, or infarct areas of heart tissues. Briefly, treated cells or infarct areas of heart tissues were lysed in RIPA lysis buffer containing phenylmethanesulfonyl fluoride (PMSF), protease inhibitors, and phosphatase inhibitors. Protein samples were fractionated by 12% SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) and transferred to polyvinylidene fluoride membranes (Millipore). The membranes were blocked and then incubated with the primary antibodies at 4°C overnight. Primary antibodies were CD9 (1:1,000; Cell Signaling Technology), CD63 (1:1,000; Cell Signaling Technology), α-SMA (1:1,000; Cell Signaling Technology), Bax (1:1,000; Cell Signaling Technology), Bcl-2 (1:1,000; Cell Signaling Technology), IL-1β (Bioworld Technology), TNF-α (Bioworld Technology), and TGF-β (Abcam, Britain). Next day, the membranes were incubated with goat anti-rabbit or anti-mouse antibodies (1:3,000; ABM, Canada) at 37°C for 1 h. Protein expressions were normalized to GAPDH (1: 2,000; CWBIO).
Statistical analyses
We used GraphPad Prism 5.0 software to perform all statistical analyses. Data are expressed as mean ± standard error of mean. Student's t-test or one-way ANOVA was used to compare experimental groups and relative control groups. A value of P < 0.05 was considered significant.
Results
Characterization of hucMSC and hucMSC-exosomes
Flow analysis displayed that hucMSCs expressed CD29, CD90, and CD105, but did not express CD19, CD34, and CD45 (Fig. 1A). hucMSCs isolated from human umbilical cord tissues were spindle shaped (Fig. 1B). Adipogenic differentiation and osteogenic differentiation assay in vitro indicated that hucMSCs differentiated into adipocytes and osteocytes successfully (Fig. 1C). The morphology of exosomes observed by a transmission electron microscope showed a typical cup shape (Fig. 1D). The size distribution displayed a homogeneous population of about 100 nm (Fig. 1E). The video image of hucMSC-exosomes was also recorded by NTA (Fig. 1F). The isolated hucMSC-exosomes also expressed CD9 and CD63 (Fig. 1G). To detect the internalization of exosomes by fibroblasts, exosomes were labeled with CM-Dil and then incubated with fibroblasts for 12 h. Results showed that hucMSC-exosomes were internalized by fibroblasts (Fig. 1H).
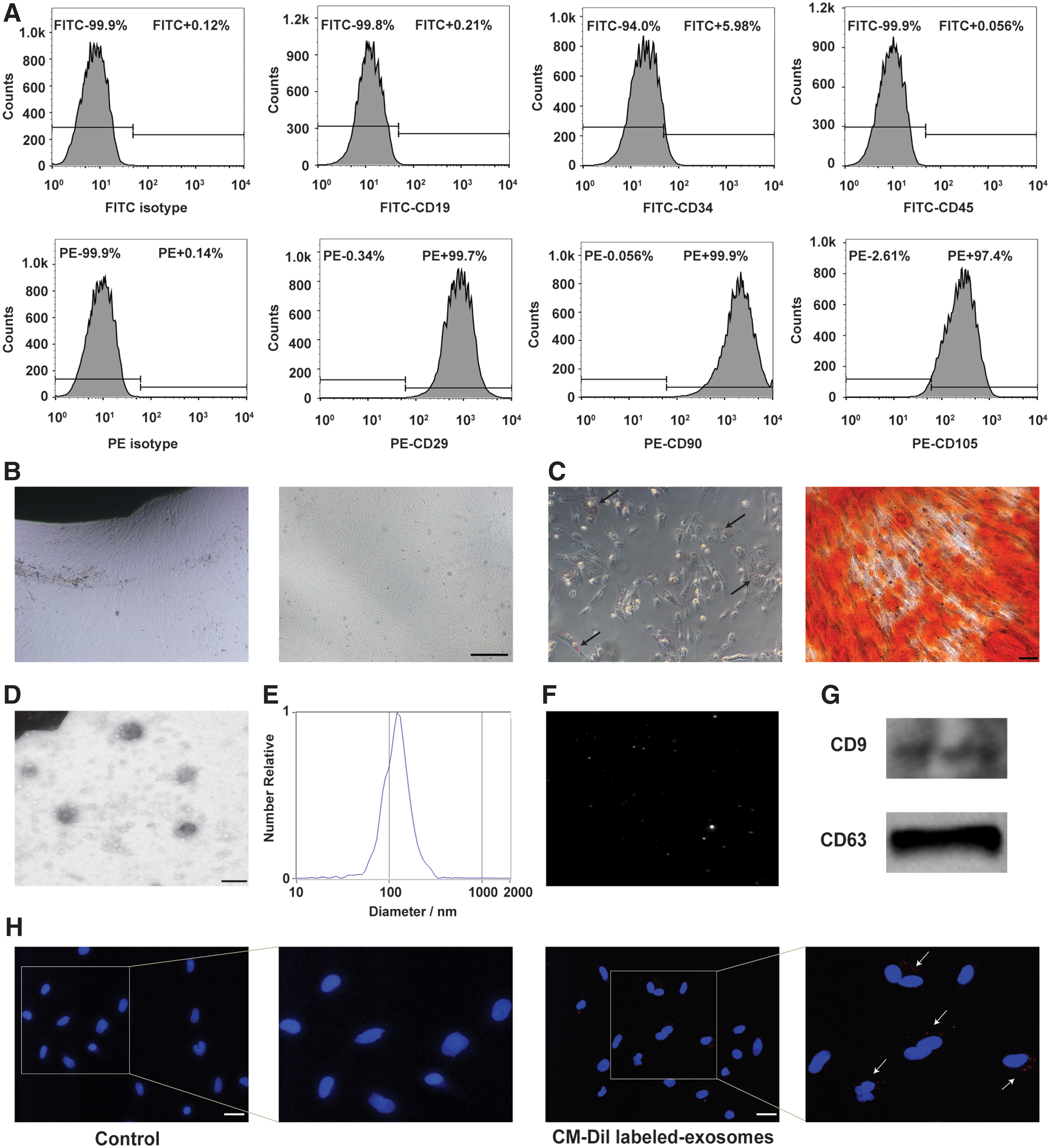
Characterization of hucMSCs and hucMSC-exosomes.
hucMSC-exosomes increased the density of myofibroblasts in infarct areas during the inflammatory phase post-MI
During the inflammatory phase post-MI, cardiac fibroblast-to-myofibroblast differentiation is inhibited. Therefore, the density of myofibroblasts in infarct areas at this phase is small, which are mainly abundant in the proliferative phase. The inflammatory phase usually lasts from first day to third day post-MI. The proliferative phase lasts from third day to fifth day post-MI, then entering the maturation phase [4]. We sacrificed rats at second day, fourth day, and seventh day post-MI and collected their hearts to perform immunohistochemical staining for α-SMA, which is the most common marker to identify myofibroblasts. In addition, it has been reported that periostin and collagen I were used to identify myofibroblasts. Therefore, we also performed immunohistochemical staining for periostin and collagen I. Results showed that hucMSC-exosomes increased the density of myofibroblasts in infarct areas during the inflammatory phase (Fig. 2A and Supplementary Figs. S1A and S2A), but there is no significant difference about the density of myofibroblasts between AMI+PBS groups and AMI+exosomes groups during the proliferative phase and the maturation phase (Fig. 2B, C and Supplementary Figs. S1B, C and S2B, C).

The effect of hucMSC-exosomes on the density of myofibroblasts in infarct hearts post-MI. Immunohistochemical staining for α-SMA was used to examine the density of myofibroblasts in infarct hearts at day 2
hucMSC-exosomes promoted cardiac fibroblast-to-myofibroblast differentiation in an inflammatory environment in vitro
Resident fibroblasts in unstressed hearts are inactivated. Following MI, fibroblasts are activated, but their differentiation into myofibroblasts is inhibited during the inflammatory phase. The phenomenon that the density of myofibroblasts in infarct areas during the inflammatory phase was increased in AMI+exosomes groups may be because hucMSC-exosomes promoted cardiac fibroblast-to-myofibroblast differentiation in the inflammatory environment. To verify this hypothesis, we used LPS to stimulate primary fibroblasts to create an inflammatory environment. Primary fibroblasts were stimulated with LPS for 6, 12, 24, and 48 h. IL-1β, TNF-α, and TGF-β mRNA levels were measured by RT-PCR. Results showed that the inflammatory response was up to its peak at 12 h and lasted for about 48 h (Fig. 3A–C). Then, we detected the α-SMA protein expression of fibroblasts treated with LPS or LPS plus hucMSC-exosomes for 12, 24, 48, and 72 h by western blot and immunofluorescence staining. Results demonstrated that hucMSC-exosomes could promote cardiac fibroblasts to differentiate into myofibroblasts at 12, 24, and 48 h, but this promotion disappeared at 72 h (Fig. 3D, E). It may be that this promotion effect of hucMSC-exosomes only function in inflammatory environments. Meanwhile, fibroblast-to-myofibroblast differentiation promoted by hucMSC-exosomes was triggered but not intense at 12 h.

hucMSC-exosomes promoted cardiac fibroblast-to-myofibroblast differentiation in the inflammatory environment in vitro. Cardiac fibroblasts were stimulated with LPS (100 ng/mL) for 6, 12, 24, and 48 h, then IL-1β
The effect of hucMSC-exosomes on fibroblast migration, proliferation, and contraction
Next, we examined the effect of hucMSC-exosomes on fibroblast migration, proliferation, and contraction. When fibroblasts differentiate into myofibroblasts, their contractile abilities are enhanced. We assessed changes in contractile ability of fibroblasts treated with LPS or LPS plus hucMSC-exosomes. Results revealed that inflammation which was created by LPS inhibited collagen gel contraction, but the addition of hucMSC-exosomes improved collagen gel contraction (Fig. 4A, B). Moreover, the migration ability of fibroblasts was decreased when they differentiated into myofibroblasts (Fig. 4C, D). Finally, we explored the effect of hucMSC-exosomes on fibroblast proliferation. Results showed that hucMSC-exosomes had no effect on fibroblast proliferation (Fig. 4E, F).
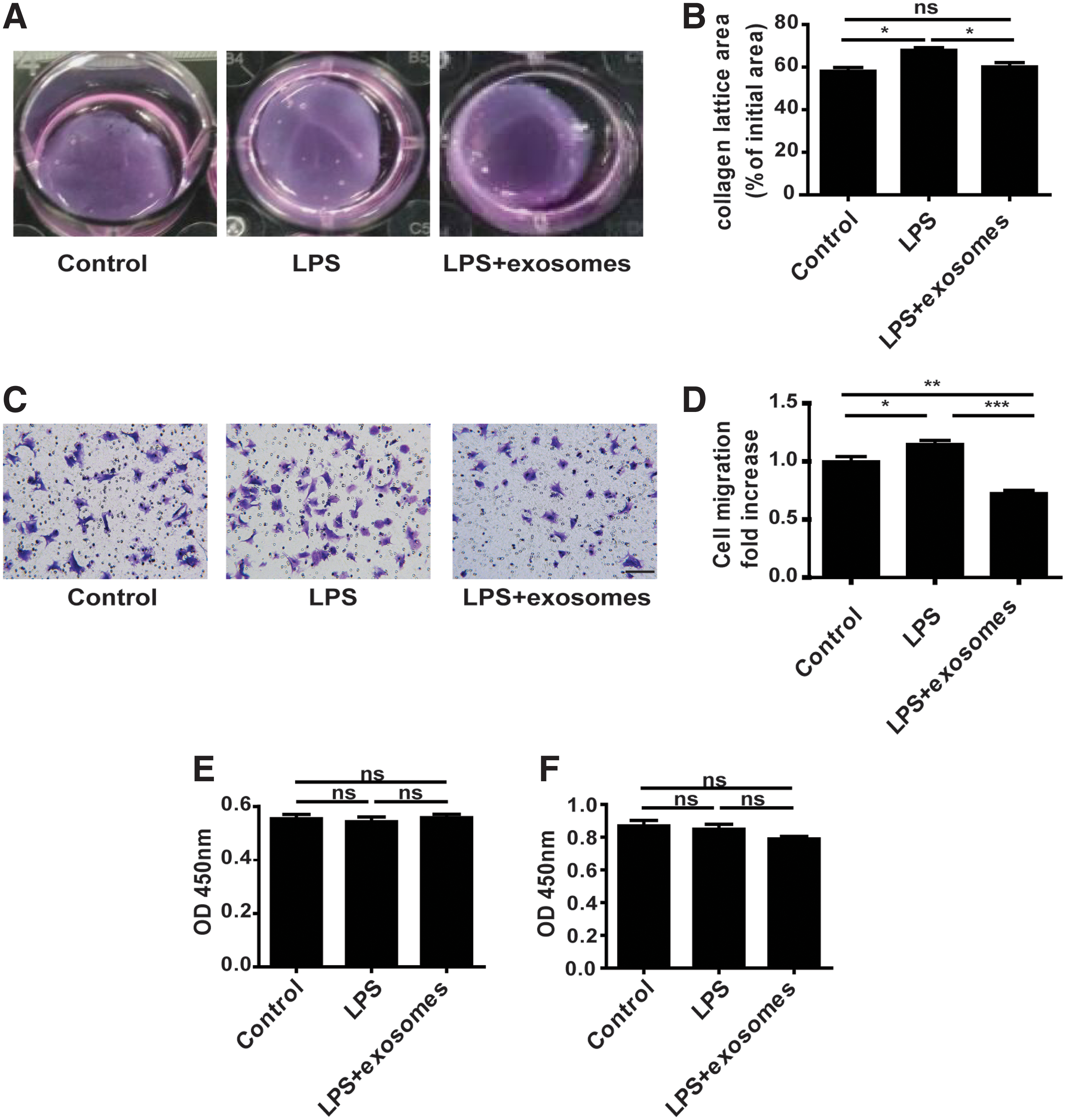
The role of hucMSC-exosomes in fibroblast migration, proliferation, and contraction.
hucMSC-exosomes attenuated inflammatory responses in vitro and in vivo
It is reported that myofibroblasts have anti-inflammatory properties. Therefore, we explored whether the inflammatory response was reduced when hucMSC-exosomes promoted fibroblasts to differentiate into myofibroblasts. Primary fibroblasts were stimulated with LPS or LPS plus hucMSC-exosomes for 12 or 24 h. Next, RT-PCR, western blot, and immunohistochemical staining were used to detect the expression of IL-1β and TNF-α, which are pro-inflammatory factors highly expressed during the inflammatory phase, and TGF-β, which is the major anti-inflammatory factor expressed in the proliferative phase. Results indicated that hucMSC-exosomes attenuated inflammation in cardiac fibroblasts at 24 h (Fig. 5C, D). However, inflammation was not relieved at 12 h (Fig. 5A, B). In vivo, the inflammatory response was also attenuated in AMI+exosomes groups at day 2 post-MI (Fig. 5E, F).
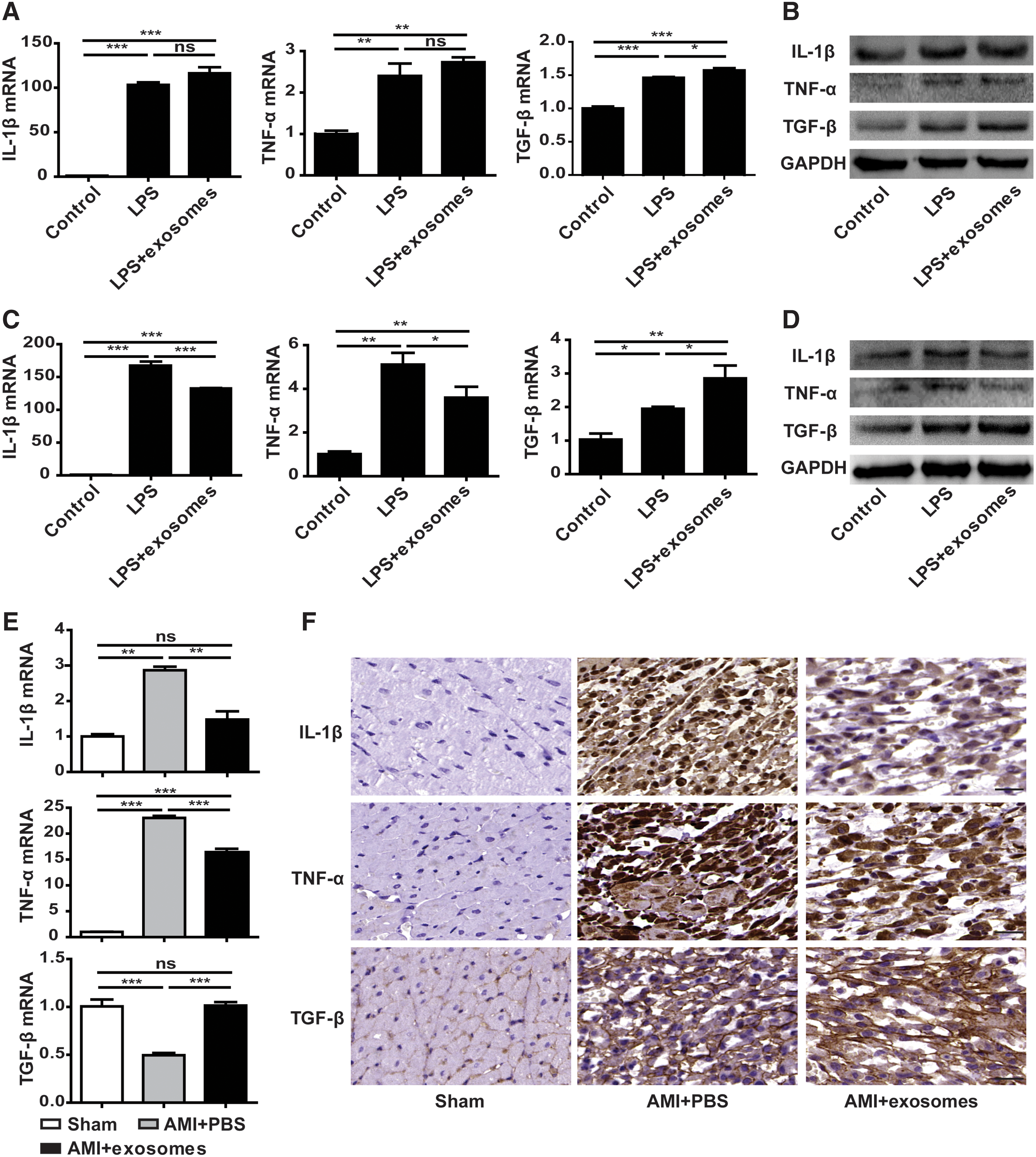
hucMSC-exosomes attenuated inflammatory responses in vitro and in vivo. Cardiac fibroblasts were stimulated with LPS (100 ng/mL) or LPS plus hucMSC-exosomes (200 μg/mL) for 12 h
hucMSC-exosomes promoting fibroblast-to-myofibroblast differentiation reduced hypoxia-induced cardiomyocyte apoptosis in vitro
Inhibition of excessive inflammation can attenuate cardiomyocyte apoptosis. Therefore, we next aimed to examine the effect of myofibroblasts into which hucMSC-exosomes promoted fibroblast differentiation on cardiomyocyte apoptosis in response to hypoxia. Cardiomyocytes were treated with supernatant secreted by fibroblasts pretreated with LPS or LPS plus hucMSC-exosomes. To eliminate the influence of LPS and hucMSC-exosomes in supernatant, fibroblasts were stimulated with LPS or LPS plus hucMSC-exosomes for 12 h. Twelve hours later, medium was replaced with 10% FBS L-DMEM to continue culturing fibroblasts for 12, 24, and 48 h, and supernatant was collected (Fig. 6A). Next, we detected α-SMA protein levels of fibroblasts treated according to above methods by western blot. The difference of α-SMA expression between LPS groups and LPS+exosomes groups was found at 12 h + 12 h. However, the difference disappeared at 12 h + 24 h and 12 h + 48 h (Fig. 6B). Cardiomyocytes were treated with supernatant that fibroblasts were continuously cultured with 10% FBS L-DMEM for 12 h and 24 h after withdrawal of treatment in hypoxic conditions. The apoptosis of cardiomyocytes was determined using TUNEL assay. Results indicated that cardiomyocyte apoptosis was increased in hypoxic control group compared with normal control group. After culturing cardiomyocytes with supernatant that fibroblasts were continuously cultured with 10% FBS L-DMEM for 12 h after withdrawal of LPS, cardiomyocyte apoptosis was further increased. However, the rate of apoptosis was reduced when cardiomyocytes were treated with supernatant that fibroblasts were continuously cultured with 10% FBS L-DMEM for 12 h after withdrawal of LPS plus hucMSC-exosomes (Fig. 6D, E). Western blot was undertaken to detect the expression of apoptosis-related proteins (Fig. 6C). Results were consistent with results of TUNEL assay.
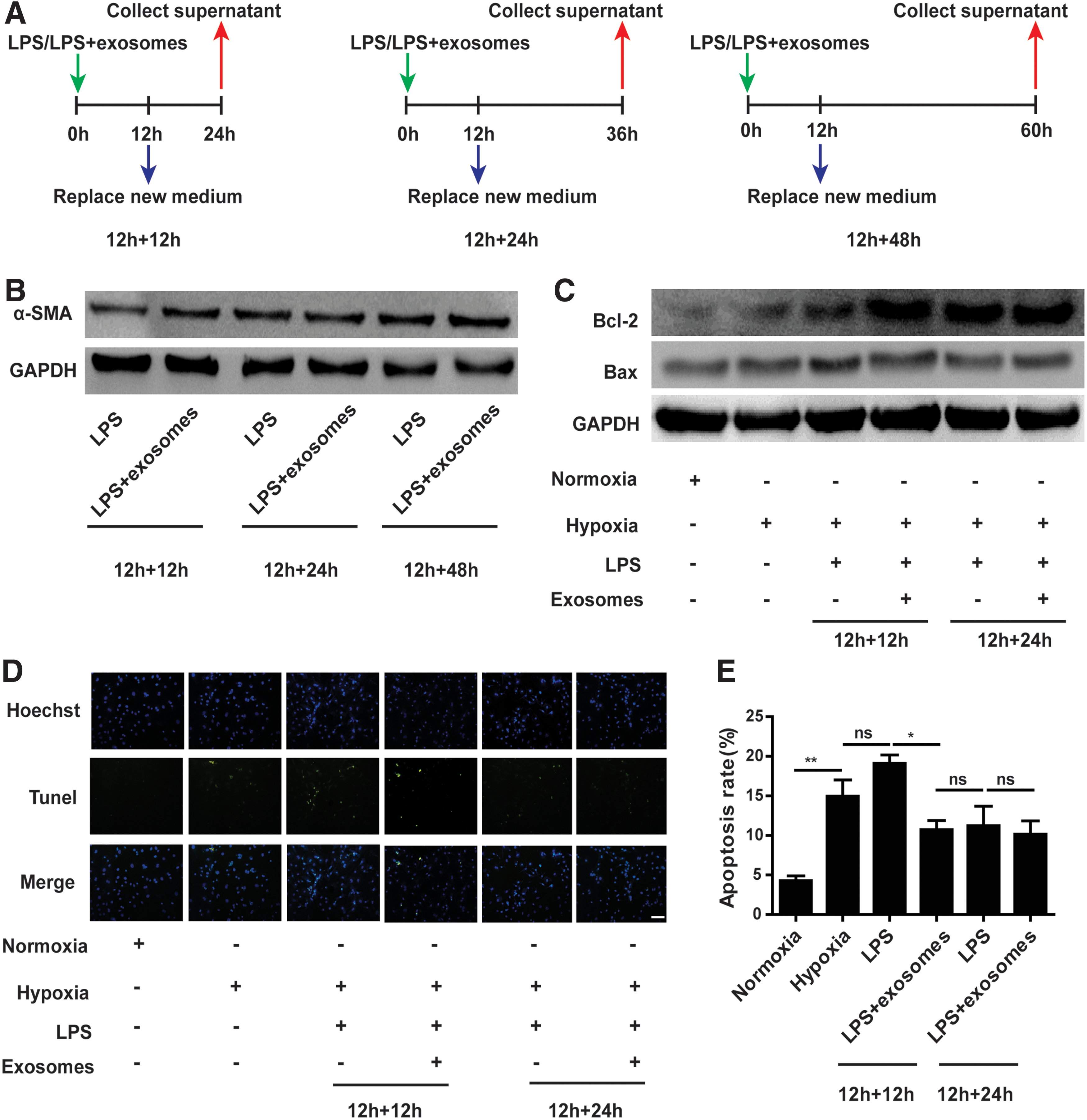
hucMSC-exosomes promoting cardiac fibroblast-to-myofibroblast differentiation influenced hypoxia-induced cardiomyocyte apoptosis.
hucMSC-exosomes reduced cardiomyocyte apoptosis following MI in vivo
Finally, we explored the effect of hucMSC-exosomes on cardiomyocyte apoptosis in vivo. Hearts of rats sacrificed at day 2 and 7 post-MI were used for following experiments. TUNEL assay was used to explore cardiomyocyte apoptosis. Results revealed that hucMSC-exosomes reduced cardiomyocyte apoptosis at day 2 (Fig. 7A, B) and day 7 (Fig. 7C, D) post-MI. Western blot was also used to detect the expression of apoptosis-related proteins in infarct areas, and same results were shown (Fig. 7E, F).

hucMSC-exosomes influenced cardiomyocyte apoptosis post-MI in vivo. TUNEL assay was used to detect cardiomyocyte apoptosis at day 2
Discussion
It is reported that MSCs restore cardiac function post-MI by paracrine signaling or differentiating into cardiomyocytes and vasculature cells [20 –22]. Extracellular vesicles secreted by MSCs, including exosomes, can improve cardiac function post-MI. We have previously reported that hucMSC-exosomes had cardioprotective effects post-MI. In this study, we mainly explored the effect of hucMSC-exosomes on fibroblast phenotypic differentiation and functional change during the inflammatory phase and the effect on cardiomyocytes. When MI occurs, cardiac repair response enters the inflammatory phase first. Some pro-inflammatory cytokines and chemokines are released, such as IL-1β. It is reported that IL-1β inhibits the migration of fibroblasts to infarct areas and also inhibits cardiac fibroblast-to-myofibroblast differentiation. As dead cells and degraded matrix are removed by macrophages, the inflammatory response is suppressed. IL-1β synthesis is suppressed, and TGF-β synthesis is facilitated. It means that cardiac repair enters the proliferative phase, and infarct areas are filled with a large number of fibroblasts, which subsequently differentiate into myofibroblasts [6,7,23]. In vivo, we found that the density of myofibroblasts in infarct areas was increased in AMI+exosomes groups during the inflammation phase post-MI. Subsequently, results indicated that hucMSC-exosomes could promote fibroblasts to differentiate into myofibroblasts in the inflammatory environment in vitro.
However, increased myofibroblasts will promote cardiac fibrosis. Our findings demonstrated that the density of myofibroblasts in infarct areas was increased in AMI+exosomes groups during the inflammation phase while there were no differences in AMI+PBS groups and AMI+exosomes groups during the proliferative phase and the maturation phase. In vitro, we also demonstrated that the promotion of hucMSC-exosomes was only maintained in the inflammatory environment, which was created by LPS and lasted about 48 h. In addition, our previous studies also demonstrated that hucMSC-exosomes did not aggravate cardiac fibrosis post-MI [15]. Therefore, there is no need to worry that this promotion during the inflammation phase will aggravate cardiac fibrosis in later stage of MI. It may be that this promotion of hucMSC-exosomes only occurs when inflammation is present, and it vanishes after inflammation is suppressed.
In the inflammation environment, more myofibroblasts may be beneficial for cardiac repair, due to the characteristics of myofibroblasts. Myofibroblasts have anti-inflammatory effect. We found that hucMSC-exosomes attenuated inflammatory responses in cardiac fibroblasts in vitro, which may attribute that hucMSC-exosomes promoted pro-inflammatory phenotype fibroblasts to differentiate into anti-inflammatory phenotype myofibroblasts. At the same time, inflammation in AMI+exosomes groups was also attenuated in vivo. However, this phenomenon attributed that hucMSC-exosomes promoted cardiac fibroblast-to-myofibroblast differentiation. hucMSC-exosomes may regulate other inflammatory cells, such as neutrophils and macrophages. In infarct areas, improved scar contraction may attenuate adverse remodeling. Furthermore, scar contraction is associated with accumulations of α-SMA-expressing myofibroblasts. In vitro, we found that fibroblasts treated with hucMSC-exosomes in the inflammation environment enhanced collagen gel contraction. However, it is reported that enhanced contraction is not due to increased myofibroblast differentiation [24,25]; perhaps, other mechanisms induced by hucMSC-exosomes are involved in enhanced collagen contraction.
It is reported that inflammation is too strong to be disadvantageous for cardiac repair. Therefore, attenuated inflammatory response may be advantageous [26,27]. Finally, we explored whether fibroblasts treated with hucMSC-exosomes could reduce cardiomyocyte apoptosis. Results showed that cardiomyocyte apoptosis was reduced, when treated with supernatant which was secreted by fibroblasts pretreated with LPS plus hucMSC-exosomes. Meanwhile, cardiomyocyte apoptosis was also reduced in AMI+exosomes groups in vivo.
Our study demonstrates that hucMSC-exosomes can promote cardiac fibroblast-to-myofibroblast differentiation during the inflammatory environment and then protect cardiomyocytes from apoptosis. Most of the researches focused on the role of exosomes in directly protecting cardiomyocytes and regulating inflammatory cells post-MI. Our study may provide new ideas for further studies on the mechanism of cardioprotective effects of exosomes.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant nos. 81800270, 81472334) and the Municipal Social Development Guide Project (grant nos. FZ2017065).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.