
Abstract
Tunneling nanotubes (TNTs) are newly discovered tubular structures between two distant cells that facilitate the intercellular exchange of signals and components. Recent reports show that mesenchymal stem cells (MSCs) can rescue injured target cells and promote recovery from a variety of stresses via TNT-mediated mitochondrial transfer. In this study, we explored how TNTs form between bone marrow MSCs and endothelial cells (ECs) by using a human umbilical cord vein endothelial cell (HUVEC) model. TNT formation between MSCs and HUVECs could be induced by treating HUVECs with cytarabine (Ara-C), and human bone marrow mesenchymal stem cells (hBMMSCs) could transfer mitochondria to injured HUVECs through TNTs. Mitochondrial transfer from hBMMSCs to HUVECs via TNTs rescued the injured HUVECs by reducing apoptosis, promoting proliferation and restoring the transmembrane migration ability as well as the capillary angiogenic capacity of HUVECs. This study provides novel insights into the cell–cell communication between MSCs and ECs and supports the results of prior studies indicating that ECs promote hematopoietic regeneration. An improved understanding of MSC-EC cross-talk will promote the development of MSC-directed strategies for improving EC function and hematopoietic system regeneration following myelosuppressive and myeloablative injuries.
Introduction
Many studies have shown that bone marrow resident endothelial cells (ECs) can accelerate hematopoietic reconstitution, and these hematopoietic stimulatory effects have been observed with ECs from a variety of sources [1,2]. Reports have indicated that bone marrow resident ECs support the proliferation and granulocytic differentiation of hematopoietic stem cells (HSCs) [3]. Moreover, ECs can produce hematopoietic growth factors under stimulated and unstimulated conditions to promote hematopoiesis. Studies have associated EC production of IL-6, SCF, G-CSF, and GM-CSF with ex vivo HSC expansion [1]. A number of previous studies have demonstrated that many signaling pathways within the endothelial niche can dictate HSC fate and promote hematopoietic regeneration. In addition, ECs actively contribute to the repair of the bone marrow microenvironment to contribute to hematopoietic reconstitution. In addition to expanding the functions of HSCs ex vivo, the direct injection of exogenous ECs presents hematopoietic stimulatory effects [4]. HSCs predominantly reside either in direct contact or in close proximity to the vascular endothelium throughout their lifespan. From the moment of HSCs embryonic specification from hemogenic endothelium, ECs act as a critical cellular hub that regulates a vast repertoire of biological processes crucial for HSCs maintenance throughout their lifespan. Recent studies have found that endothelial niche-mediated regulation of HSCs function occurs during development, aging, and regenerative conditions and suggested that ECs provide a fertile niche for the maintenance of functional HSCs throughout their lifecycle [5]. Hematopoietic recovery after high-dose chemotherapy (HDC) in the treatment of hematological diseases may be slow and/or incomplete, which is generally attributed to progressive HSC failure, although defective hematopoiesis may also be partially due to poor stromal function [6]. Accordingly, injured ECs in a damaged bone marrow microenvironment under chemotherapy will inevitably hinder hematopoietic reconstitution; thus, it is necessary to repair the injured ECs in bone marrow to help promote hematopoietic reconstitution.
In addition, the administration of human mesenchymal stem cells (MSCs) has been shown to accelerate hematopoietic reconstitution in animal models [7,8]. Accordingly, one of the first clinical uses of MSCs has been to accelerate hematopoietic recovery. The use of MSCs to accelerate hematopoietic reconstitution has been performed in patients with neoplasia who received HDC, and leukocytic and thrombocytic reconstitution appeared to undergo “rapid recovery” [9]. However, the mechanism by which MSCs promote hematopoietic reconstitution is still unknown. Although the in vivo significance of MSCs is still highly debated, one theory is that MSCs in the bone marrow provide a suitable environment for hematopoiesis. MSCs and ECs are two major stromal cells residing in the bone marrow microenvironment, and they are in close proximity to each other; thus, it is very important to clarify the MSC-EC cross-talk in stromal microenvironments to further clarify their contributions to hematopoietic reconstitution. In this study, we hypothesized that MSCs can rescue injured ECs residing in the adjacent bone marrow environment by novel cell–cell communication vehicles called Tunneling nanotubes (TNTs), which contributes to the hematopoietic reconstitution of patients suffering from chemotherapy.
TNTs were first described in 2004 as freely hovering tubular structures between two cells [10]. Reports have suggested that the TNT-mediated transfer of functional mitochondria from healthy MSCs to target cells can rescue cells injured by oxidative stress, ultraviolet radiation, inflammation, and ischemia/reperfusion (I/R) [11]. Our study is the first to investigate the formation of TNTs between MSCs and ECs under chemotherapy stress and to explore the significance of TNT cell–cell communication and the mediation of mitochondrial transfer in the recovery of chemotherapy-stressed ECs. Our results illustrate the new cell–cell communication styles between MSCs and human umbilical cord vein endothelial cells (HUVECs) and provide insights into the mechanism underlying the capacity of MSCs to promote hematopoietic reconstitution.
Materials and Methods
Cell cultures and cell lines
This study was approved by the Peking University People Hospital Medical Ethics Committee. Bone marrow samples were collected from healthy donors who provided informed consent. Mononuclear cells (MNCs) were isolated from bone marrow samples via Ficoll-Hypaque density gradient centrifugation. MNCs were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) with 15% FBS at 37°C and under 5% circulating CO2. Explants were kept in medium for 5 days; the medium was changed every 2 days. Once cell outgrowth was subconfluent, cells were harvested with 0.25% (w/v) trypsin-EDTA (Gibco) and subcultured with a split ratio of 1:3. Subsequently, bone marrow mesenchymal stem cells (BMMSCs) were maintained in DMEM supplemented with 10% FBS. Cells at passage 3 were used in the following experiments.
HUVECs were purchased and cultured in Endothelial Cell Growth Medium-2 (EGM-2; Lonza, Walkersville, MD) containing a growth factor cocktail with 5% FBS at 37°C and under 5% circulating CO2. Then, the third passage MSCs and HUVECs were confirmed by the FCM and immunofluorescence experiments (data not shown).
hBMMSCs cocultured with HUVECs and TNT formation and breakage
Induction of TNT formation between hBMMSCs and HUVECs
hBMMSCs were cocultured with HUVECs in 1:1 ratio in glass-bottomed dishes (Costar Corning, NY) and incubated at 37°C under 5% CO2 for 24 h. HUVECs were pretreated with cytarabine (Ara-C; 4 μM) for 24 h to induce stress (HUVECs Ara-C), while untreated HUVECs were used as a control group (HUVECs CON). Both groups were then cocultured with hBMMSCs.
TNT breakage
To selectively inhibit TNT formation, the medium of the direct coculture system was treated with microfilament depolymerization reagents cytochalasin D (CyD, 7.5 μM, 5 h; Sigma, St. Louis, MO) or latrunculin A (LAT, 100 nM, 19 h; Sigma) and microtubule depolymerization reagent nocodazole (NOC, 30 μM, 5 h; Sigma).
HUVEC stress and apoptosis detection
hBMMSCs and HUVECs were individually cultured overnight in 24-well plates (1 × 105 cells/mL) (Corning) at a 1:1 ratio. Gradually increasing concentrations of cytarabine (Ara-C; 0.0, 0.4, 4, 40 μM) were added to the medium, and the cells were cultured for 24 h to induce stress.
Apoptosis of HUVECs and hBMMSCs was measured using Annexin V (PE) and 7AA-D (PerCP) with an apoptosis detection kit (BD Pharmingen, SanDiego, CA) according to the manufacturer's protocol to test the tolerance of these two cells to cytarabine. The samples were analyzed via LSRFortessa flow cytometry (FCM; BD Biosciences, Franklin Lakes, NJ). Fluorescence-activated cell sorting (FACS), scanning, and data analysis were performed using a FACS analyzer and CellQuest software (BD Biosciences).
Immunofluorescence detection and confocal imaging of TNTs and mitochondrial transfer
Membrane staining
Cocultures of hBMMSCs and HUVECs in glass-bottom dishes (Corning) were fixed with 4% paraformaldehyde (PFA). Immunofluorescence staining was performed according to standard procedures using mouse monoclonal anti-CD31 primary antibody (Abcam, Cambridge, MA) with goat anti-mouse- Alexa Fluor 488 or goat anti-mouse- Alexa Fluor647 secondary antibodies (Invitrogen, Carlsbad, CA), as appropriate, to stain the HUVEC membranes.
Cytoskeletal staining
To determine the composition of TNTs, hBMMSCs and HUVECs were seeded onto glass-bottom dishes (2 × 104 cells/mL) at a 1:1 ratio and cocultured for 12 h, and the coculture system was treated with 4% PFA. Immunofluorescence staining was performed according to standard procedures: rabbit monoclonal anti-α-tubulin (Abcam) was used as the primary antibody and goat anti-rabbit-Alexa Fluor 633 (Invitrogen) was used as the secondary antibody to stain microtubules, while F-actin was labeled with rhodamine phalloidin (Invitrogen).
TNT formation process between BMMSCs and HUVECs
To determine how TNTs form between BMMSCs and HUVECs, HUVECs were labeled with Alexa Fluor® 647-conjugated wheat germ agglutinin (WGA) (Invitrogen), and hBMMSCs were labeled with Alexa Fluor 555-conjugated WGA (Invitrogen). Then, hBMMSCs and HUVECs were cocultured, and the glass-bottomed dishes were imaged using live-cell workstations at 37°C under 5% CO2 after 3 h of coculture.
TNT counts
Cocultures of hBMMSCs and HUVECs (pretreated with 4 μM Ara-C or not) were incubated with anti-human CD31 (1:300) to specifically mark HUVECs. TNTs were counted in 20 randomly selected fields of view under a confocal microscope and averaged in the two groups. Experiments were performed in duplicate.
Mitochondrial transfer between BMMSCs and HUVECs via TNTs
To observe the TNT-mediated mitochondrial transfer between HUVECs and BMMSCs, viral transfection of BMMSCs or HUVECs and CellTracker tracing of HUVECs or BMMSCs were alternately performed followed by the detection of mitochondrial transfer from BMMSCs to HUVECs or from HUVECs to BMMSCs. The details were as follows. At 70% confluence of BMMSCs or HUVECs, the mitochondria in living cells were transfected for 24 h with Ad-Mito-EGFP, which contained the mitochondria-specific targeting sequence 5′-ATGTCCGTCCTGACGCCGCTGCTGCTGCGGGGCTTGA-3′ (Jikai Gene, Shanghai, China). HUVECs were stained with 10 mM CellTracker™ Orange CMRA Dye (Invitrogen) at 37°C for 30 min and washed twice. After 3 h of coculture of the CellTracker-labeled HUVECs with BMMSCs transfected with Ad-Mito-enhanced green fluorescent protein (EGFP) (BMMSC-Mito-eGFP), the glass-bottom dish was imaged in live-cell workstations at 37°C under 5% CO2. Two-hour recordings of mitochondrial transfer from BMMSCs to HUVECs were acquired after 3 h of coculture. A similar method was used to detect mitochondrial transfer from HUVECs to BMMSCs.
All imagings were performed using a Zeiss LSM780 confocal laser scanning microscope (Carl Zeiss, Oberkochen, Germany). Post-acquisition image analysis was performed with ZEN 2012 x32 blue software (Carl Zeiss). Images were taken with an oil-immersed 63X/1.4NA objective (Carl Zeiss).
Rescue of stressed HUVECs by TNTs
Grouping: HUVEC treatments
(i) Healthy untreated HUVECs were used as a control group (HUVECs CON). (ii) HUVECs were treated with Ara-C (4 μM) for 24 h to induce apoptosis and were then treated with Ara-C (0.1 μM) for 48 h to induce proliferation inhibition (HUVECs Ara-C). (iii) Stressed HUVECs were directly cocultured with healthy MSCs for 24 h at a 1:1 ratio. (iv–vi) During the direct coculture process, the cells were treated with CyD, LAT, or NOC to block TNT formation.
HUVEC apoptosis detection
Apoptosis was assessed in HUVECs subjected to different treatments as described above and was measured using annexin V (PE)/7AA-D (PerCP) in CD31+ cells with an apoptosis detection kit according to the manufacturer's protocol.
Detection of carboxyfluorescein diacetate succinimidyl ester in HUVECs
HUVECs were stained with a carboxyfluorescein diacetate succinimidyl ester (CFSE) fluorescent probe (Invitrogen) and PE-CD31 antibody in each group. Fluorescence dilution of CFSE and the MFI of CFSE in CD31+ cells were detected by FCM (BD Biosciences).
Coculture and TNT effects on capillary angiogenesis and transmembrane migration
Cells of different groups were digested and incubated for 24 h in a 24-well plate precoated with a thawed Matrigel solution (BD Biosciences, Baltimore, MD) according the manufacturer's protocol to observe the capacity of capillaries to undergo angiogenesis, and the cells were transferred to the upper chamber of the corresponding 12-well Transwell plate (Corning) and incubated for 24 h to perform the transmembrane migration experiment. Finally, the tubes and knots of capillaries and the migrated cells were observed and counted under an inverse phase-contrast microscope (Olympus, Japan).
Statistical analysis
Data are presented as the mean ± standard deviation from at least three independent experiments. The statistical analysis was performed using SPSS 17.0 (SPSS, Inc.). Student's t-test or analysis of variance (ANOVA) combined with a post hoc LSD test was used to analyze the differences among groups (P < 0.05 was considered statistically significant).
Results
TNT formation between hBMMSCs and HUVECs mediated bidirectional mitochondrial transfer
Immunofluorescence staining was performed, and confocal microscopy showed that elongated membrane-like structures gradually formed between hBMMSCs (WGA-555) and HUVECs (WGA-647) (Fig. 1A). Cocultured HUVECs were stained positively with anti-CD31 antibody, while the membrane-like structures could be stained with phalloidin and anti-α-tubulin antibody, which showed that the structural components of these TNTs contained F-actin and microtubules (Fig. 1B). This result indicates that TNTs allow for the direct transfer of mitochondria between hBMMSCs and HUVECs (Fig. 1C). The TNT connections in cocultures that were pretreated with CyD, LAT, and NOC were destroyed, whereas such destruction was not observed in the cells without pretreatment showed under an inverted phase-contrast microscope (data not shown).
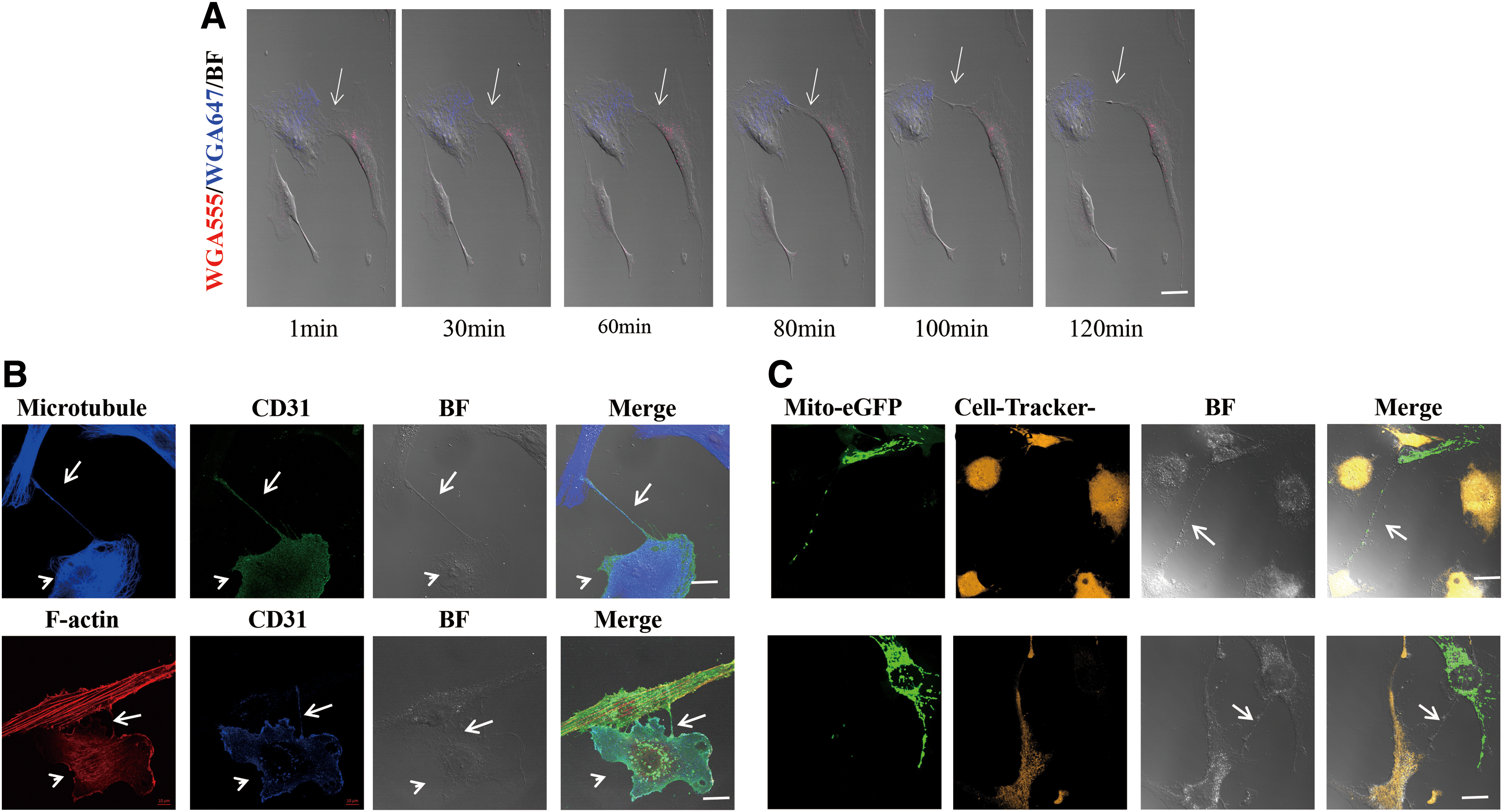
TNT formation and mediated bidirectional transport of mitochondria between hBMMSCs and HUVECs.
TNTs mediate unidirectional mitochondrial transfer from hBMMSCs to stressed HUVECs with cytarabine pretreatment
To investigate the rescue effect between hBMMSCs and HUVECs, increasing concentrations of Ara-C were used to induce chemotherapy stress in hBMMSCs and HUVECs. The results showed that the apoptosis rate of HUVECs and MSCs increased with increases in the cytarabine concentration. The apoptosis rate of HUVECs treated with 4 μM Ara-C was significantly higher than that of hBMMSCs treated with 4 μM Ara-C (20.00% ± 4.10% vs. 6.06% ± 0.90%, P < 0.05) (Fig. 2A). Since the results showed that HUVECs were more susceptible to 4 μM Ara-C-induced damage than hBMMSCs, HUVECs with or without pretreatment with 4 μM Ara-C cocultured with healthy hBMMSCs were evaluated, which was followed by the detection of TNTs between heterotypic cells. The results showed that significantly more TNTs formed between hBMMSCs and stressed HUVECs with 4 μM Ara-C pretreatment than between hBMMSCs and HUVECs without pretreatment, with 8.2 versus 1.8 TNTs observed across an average of 20 visual fields (P < 0.05, n = 4) (Fig. 2B).
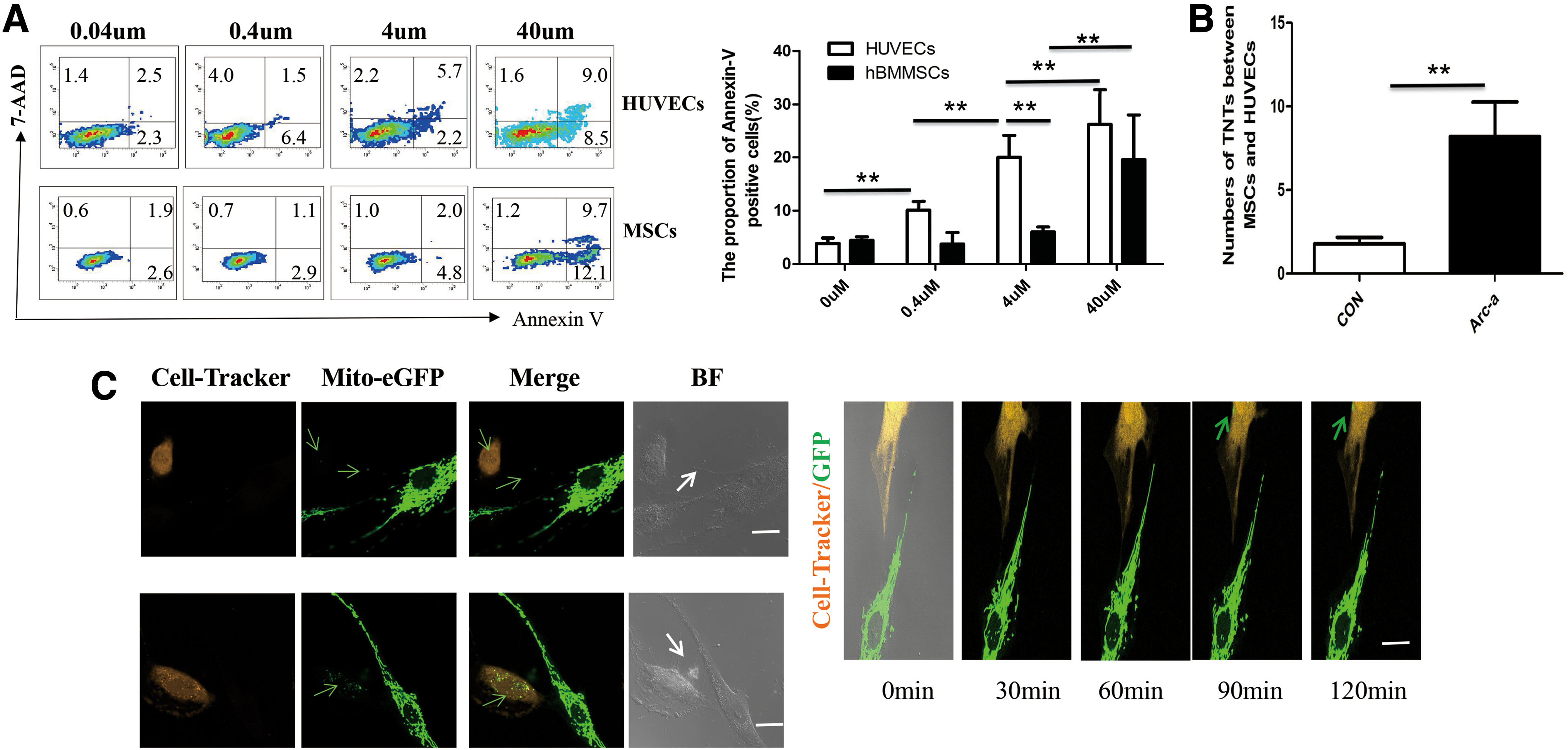
TNTs formed between hBMMSCs and HUVECs with treatment by cytarabine can unidirectionally transport mitochondria from hBMMSCs to stressed HUVECs.
To further study mitochondrial transfer mediated by TNTs between hBMMSCs and HUVECs stressed by Ara-C, HUVECs with pretreatment with cytarabine were cocultured with healthy hBMMSCs for 24 h. Notably, after cocultivation, we were not able to detect green-fluorescing mitochondria in the hBMMSCs coming from stressed HUVECs (data can be acquired). The HUVECs with cytarabine pretreatment labeled by CellTracker-Orange were also cocultured with hBMMSCs transfected with Ad-Mito-eGFP for 24 h. Mito-eGFP signal can be observed transferring from hBMMSCs to HUVECs through TNTs, and a considerable amount of Ad-Mito-eGFP can be seen in the plasma of HUVECs tracked by CellTracker-Orange (Fig. 2C left panel). A series of confocal images from the time-lapse movies (movies can be acquired) show the movement process of Ad-Mito-EGFP-labeled mitochondria in TNTs between hBMMSCs and HUVECs treated with Ara-C. These results indicated that hBMMSCs can unidirectionally transfer mitochondria to HUVECs stressed with Ara-C. When cocultures were treated with CyD, LAT, and NOC to destroy TNTs, the mitochondrial transfer rates correspondingly decreased compared with the control coculture group without treatment (P < 0.05, n = 3)(Supplementary Fig. S1).
Transfer of mitochondria from BMMSCs to HUVECs can alleviate HUVECsAra-C apoptosis and promote HUVECAra-C proliferation
The results showed that the apoptosis rate of HUVECs Ara-C was significantly higher than that of HUVECs CON (20.02 ± 6.96 vs. 5.52 ± 3.18, P < 0.01, n = 5). Direct coculture with hBMMSCs significantly alleviated the apoptosis rate of HUVECs Ara-C (8.78 ± 2.85 vs. 20.02 ± 6.96, P < 0.01, n = 5). The rescue effect of the direct coculture system on HUVECs Ara-C could be alleviated by pretreatment of the coculture after system with CyD, LAT, and NOC, which disrupt TNTs. TNT apoptosis significantly increased in the coculture after pretreatment with CyD, LAT, or NOC compared with the direct coculture group without pretreatment (18.44 ± 5.46 due to CyD, 21.60 ± 8.66 due to LAT, and 18.56 ± 7.25 due to NOC vs. 8.78 ± 2.85 in coculture; P < 0.05, n = 5) (Fig. 3A).

Transport of mitochondria through TNTs from hBMMSCs to HUVECs attenuates the apoptosis rate and promotes the proliferation of HUVECs treated with 4 μM Ara-C.
The results showed that HUVEC proliferation was inhibited by 0.1 μm Ara-C treatment for 48 h. The CFSE signal from HUVECsAra-C was significantly increased compared with that from HUVECsCON (5234.50 ± 1439.54 vs. 1761.25 ± 641.10, P < 0.01, n = 4). The MFI of the CFSE signal from the direct coculture group was significantly lower than that of the HUVECAra-C group (2675 ± 1271.96 vs. 5234.50 ± 1439.54, P < 0.05, n = 4). Compared with the direct coculture group, the CFSE signal was significantly increased in HUVECs pretreated with CyD and NOC (5178 ± 1961.27 due to CyD, 5240.50 ± 1879.67 due to NOC vs. 2675 ± 1271.96 in the direct coculture group; P < 0.05, n = 4), while the CFSE signal in the LAT pretreatment group showed a nonsignificant increase (4538.75 ± 2048.57 vs. 2675 ± 1271.96 in the direct coculture group; P = 0.232, n = 3) (Fig. 3B).
Mitochondrial transfer from hBMMSCs to HUVECs can improve HUVEC transmembrane migration capacity and capillary angiogenesis
The number of migrated cells in the HUVECs Ara-C group was significantly decreased compared with that in the HUVECsCON group (57.25 ± 9.85 vs. 15.00 ± 7.48), while direct coculture with hBMMSCs significantly increased the migrated cell count of the HUVECsAra-C group (29.50 ± 8.06 vs.15.00 ± 7.48, P < 0.01, n = 4). In the groups pretreated with CyD and LAT, the number of transmembrane-migrated HUVECs was significantly reduced compared with that in the direct coculture group without pretreatment (14.50 ± 6.40 for the CyD-group, 16.00 ± 2.45 for the LAT- group vs. 29.50 ± 8.06 for the direct coculture group; P < 0.05, n = 4). The number of migrated HUVECs in the NOC group was less than that in the direct coculture group (11.75 ± 2.22 vs. 29.50 ± 8.06, P < 0.01, n = 4). The above results showed that the Ara-C treatment significantly weakened the transmembrane migration of HUVECs and that direct coculture with hBMMSCs improved the capacity of HUVECsAra-C via TNT cell–cell communication between hBMMSCs and HUVECsAra-C (Fig. 4A).
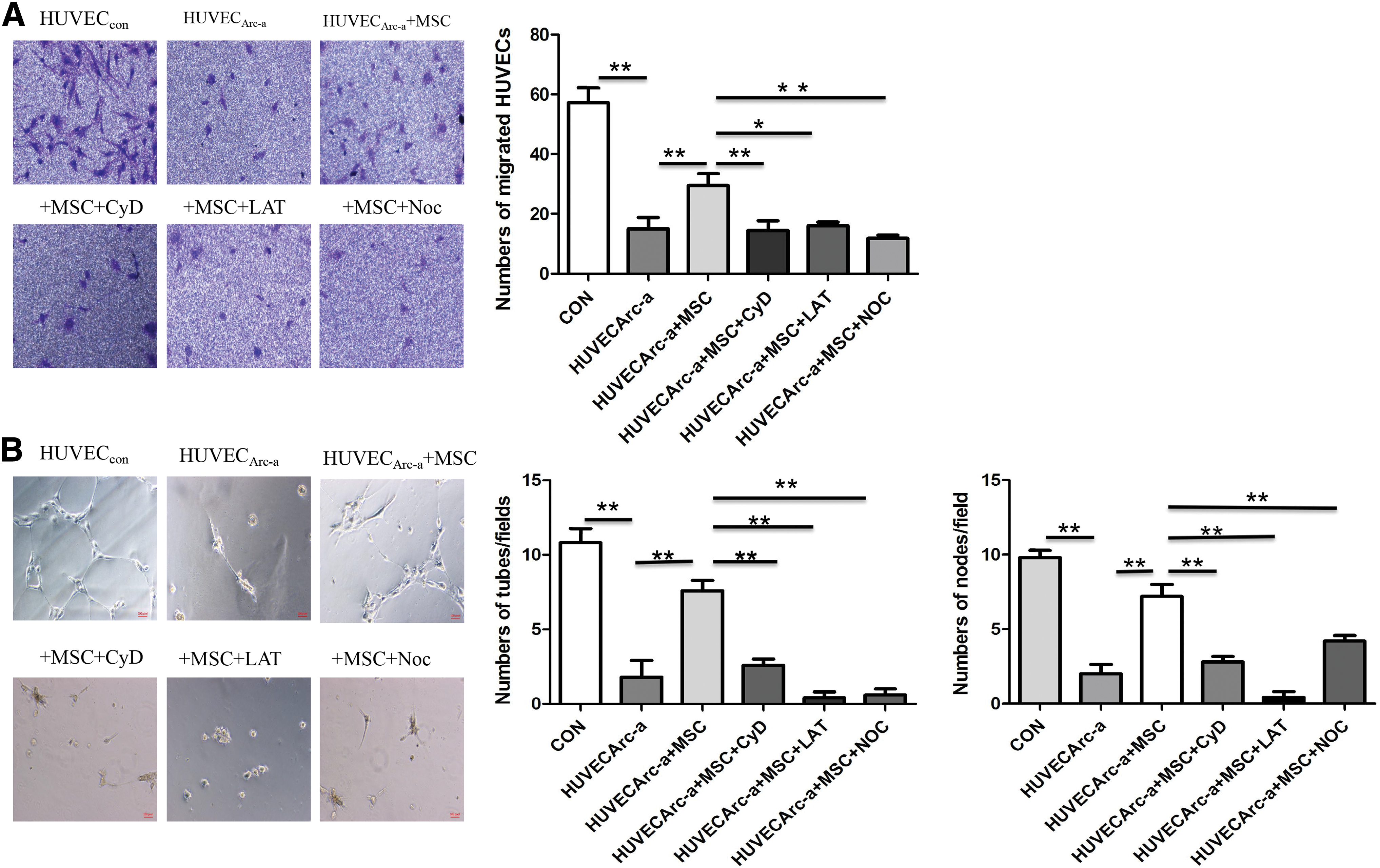
Direct coculture of hBMMSCs and Ara-C-treated HUVECs improves the capillary development on Matrigel and Transwell transmembrane migration capacities of stressed HUVECs.
HUVECsCON incubated on Matrigel for 24 h formed a complete capillary-like structure, in which cells were connected in a mesh-like arrangement, whereas HUVECsAra-C were clustered and formed markedly reduced lumen structures and a few incomplete capillary-like structures. After pretreatment with Ara-C, the capillary formation capacity of stressed HUVECs was markedly attenuated. Significantly, fewer integrated capillary structures were observed in the HUVECsAra-C group than in the control group (tubes: 1.8 ± 2.5 vs. 10.8 ± 2.2; knots: 2 ± 1.41 vs. 9.8 ± 1.1; P < 0.01, n = 5). More tubes and knots formed in the direct coculture group than in the HUVECsAra-C group (tubes: 7.6 ± 1.5 vs. 1.8 ± 2.5; knots: 7.2 ± 1.8 vs. 2.0 ± 1.4; P < 0.01, n = 5). The use of CyD, LAT, and NOC to disrupt TNTs in the direct coculture groups led to less complete tube formation on Matrigel than in the untreated direct coculture group, and significantly, fewer tubes and knots formed in the CyD, LAT, and NOC pretreatment groups than in the direct coculture group without pretreatment (tubes: 2.6 ± 0.89 due to CyD, 0.4 ± 0.89 due to LAT, and 0.6 ± 0.89 due to NOC vs. 7.6 ± 1.5 in direct coculture; knots: 2.8 ± 0.8 due to CyD, 0.4 ± 0.9 due to LAT, and 4.2 ± 0.8 due to NOC vs. 7.2 ± 1.8 in direct coculture; P < 0.01, n = 5) (Fig. 4B).
MSC proliferation assays were performed using the Cell Counting Kit-8 (CCK-8), and several common cytokine (HGF, EGF, VEGF, and TGF-β) concentrations in supernatants of different MSC populations were assessed using ELISA to demonstrate that pretreatment with CyD, LAT, and NOC does not affect other functional properties of MSCs, including viability and capacity, to secrete soluble factors. Our results indicated that there was no significant difference between the drug treatment groups and the control group in terms of both MSC viability and their capacity to secrete soluble factors (P > 0.05, n = 4) (Supplementary Fig. S2).
Discussion
hBMMSCs have been extensively studied for the treatment of bone marrow failure and have been clinically used to promote hematopoietic recovery following HSCT. However, the mechanism of hBMMSC-facilitated hematopoietic recovery is not yet fully understood. Therefore, it is possible that bone marrow MSCs and ECs interact by cell–cell communication to coordinate the hematopoietic microenvironment and to promote hematopoietic reconstruction after chemotherapy. TNTs are a newly discovered cell-to-cell communication modality that allows for the transmission of cytoplasmic contents, signals, and organelles between cells and thus may participate in altering the biological functions of involved cells. Reports have shown that hBMMSCs can communicate with a variety of injured target cells via TNTs to repair and rescue target cells. Reports have shown that hBMMSCs can transfer mitochondria to cells following I/R injury via TNTs to rescue the stressed target cells [12 –16]. This study is the first to explore TNTs between two types of stromal cells resident in bone marrow, that is, MSCs and ECs, and the role of TNTs in transferring mitochondria to repair chemotherapy-stressed ECs and thereby promote hematopoietic reconstruction.
In our study, TNTs that formed between hBMMSCs and HUVECs could be observed under confocal microscopy. The entire length of the TNTs is composed of F-actin and microtubules, and TNT formation can be blocked by pretreatment with an actin and a microtubule depolymerization agent. Both microtubules and F-actin were found to be necessary components of the TNTs that formed between hBMMSCs and HUVECs, which represent a key piece of evidence that complements previous studies [16]. The results showed that the formation and fracture of TNTs are dynamic processes and that TNTs are formed between hBMMSCs and HUVECs by retaining a thin thread of membrane upon dislodgement. This process has been reported to be the most common mode of TNT formation [17,18], and it differs from the process described in PC12 cells by Rustom et al., in which cells extend protrusions or pseudopodia to adjacent cells to form TNT bridges [10].
In the coculture system, TNTs appeared as membrane structures that originated from HUVECs and were extended to hBMMSCs, and these TNTs mediated mitochondrial transfer from hBMMSCs to HUVECs. Then, we established a direct coculture system of hBMMSCs and HUVECs to further study the role of TNTs through using Ara-C to stress HUVECs. We found that significantly more TNTs were formed between hBMMSCs and injured HUVECs than between hBMMSCs and healthy HUVECs. This result suggests that in addition to I/R [13] and inflammatory [18] stresses, chemotherapy stress may stimulate TNT formation, which has not been reported in previous studies. TNTs may be easily formed by injured cells as a rescue signal from surrounding cells.
We observed that hBMMSCs unidirectionally transferred mitochondria to stressed HUVECs, whereas hBMMSCs could bidirectionally exchange mitochondria with unstressed HUVECs. The blockage of TNT formation significantly reduced mitochondrial transfer from hBMMSCs to stressed HUVECs. In addition, HUVEC apoptosis and proliferation in different treatment groups were detected by flow cytometry. The results showed that hBMMSCs could rescue Ara-C-stressed HUVECs via TNT-mediated mitochondrial transfer from MSCs to HUVECs through reducing HUVEC apoptosis and promoting HUVEC proliferation. Mitochondria are the energy factories of cells and control cell survival, stress, and death [19]. Mitochondria also play key roles in the maintenance of oxidative phosphorylation, glucose and fat aerobic metabolism, calcium signaling, and apoptosis [19]. During apoptosis, a reduction in mitochondrial membrane potential is an early event that activates the respiratory chain by releasing cytochrome C and activating the caspase cascade [20]. Using fluorescent imaging and flow cytometry, we observed that extensive mitochondrial transfer occurred from MSCs to stressed HUVECs and occurred at least partially through TNT-like structures. Moreover, coculture with hBMMSCs reduced the apoptosis rate of chemotherapy-stressed HUVECs, and the blockage of TNT formation reduced mitochondrial transfer and abrogated the rescue effect of MSCs, which indicated that TNT-mediated mitochondrial transfer from hBMMSCs to HUVECs takes part in the rescue effect of MSCs. Many previous reports have indicated that TNT-mediated mitochondria transfer can exert an effect on the apoptosis of targeted cells [13,16]. It has been reported that the apoptosis of UV-stressed PC12 cells can be rescued by transferring mitochondria from healthy cells [21]. Therefore, we concluded that TNT-mediated mitochondrial transfer of MSCs plays a key role in the rescue effect on chemotherapy-stressed HUVECs; whether other cytoplasmic contents are also transferred via TNTs and participate in the rescue effect on stressed HUVECs still requires further research. These research results provide a verification of and supplement to previous studies that showed that hBMMSCs can improve cerebral ischemia via TNTs, such as in structure-mediated mitochondrial transfer to decrease the apoptosis of HUVECs [16]. We also found that hBMMSCs promoted the proliferation of stressed HUVECs. The finding that injured target cell proliferation was promoted by TNT-mediated mitochondrial transfer is consistent with that of a previous study reported by Dumler, who showed that vascular smooth muscle cells (VSMCs) promoted MSC proliferation through TNT-mediated mitochondrial transfer from VSMCs to injured MSCs [22]. In addition, hBMMSCs improved capillary formation and the migration of stressed HUVECs, indicating that TNT-mediated mitochondrial transfer from hBMMSCs to HUVECs could also restore the characteristics of HUVECs, which is similar to the capacity of TNT-mediated mitochondrial transfer from hBMMSCs to macrophages to enhance macrophage phagocytosis in models of ARDS. TNT-mediated mitochondrial transfer may essentially repair the characteristics of damaged cells [23].
In summary, this study showed that hBMMSCs could form TNTs with HUVECs and could rescue Ara-C-stressed ECs in vitro by reducing apoptosis, promoting cell proliferation, and restoring the capillary formation and migration capacities of injured HUVECs via TNT-mediated mitochondrial transfer. According to previous reports, TNTs can also transfer cytoplasmic material and organelles between two distant cells, and our study indicated that the rescue effect of MSCs on HUVECs was at least partially related to mitochondrial transfer via TNT-like structures, which are dependent on direct cell contact. Interestingly, our study indicated that TNTs may transport other additional cytoplasmic material and may be involved in other organelle transfer mechanisms during the repair of HUVECs, possibilities that require further study. In the experiment presented here, we observed the release of exosome-encapsulated mitochondria into the medium under confocal microscopy, which indicated that hBMMSCs may have exerted a rescue effect on cytarabine-stressed HUVECs through some other cell–cell communication styles.
As shown above, bone marrow resident ECs support the proliferation and differentiation of HSCs and the production of hematopoietic growth factors to promote hematopoiesis and actively contribute to the repair of the bone marrow microenvironment to promote hematopoietic reconstitution. In summary, ECs resident in bone marrow provide a fertile niche for the maintenance of functional HSCs. Thus, according to our research results and previous reports, hBMMSCs might contribute to hematopoietic recovery via the transfer of mitochondria to stressed bone marrow ECs. Nonetheless, these findings provide a theoretical basis and a new direction for the study of cell–cell talk between stromal cells residing in the bone marrow microenvironment as well as insights into their contributions to hematopoietic recovery. In a future study, we plan to further elucidate the effect of ECs via mitochondrial transfer on hematopoietic recovery.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (no. 81270572 and no. 81760026). We express our grateful acknowledgement to professor YY Zhang for her constant support throughout the work. The authors also acknowledge the assistance of American Journal Experts (AJE) for their English language editing. The authors are grateful for the experimental platform afforded by the Institute of Vascular Medicine of Peking University Third Hospital.
Our abstract had been selected by the Scientific Program Committee for an oral presentation during the 22nd Congress of the European Hematology Association held in Madrid, Spain on Friday, June 23, 2017 (Submission ID: EHA-3566, Final Abstract Code: S138).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.