
Abstract
What can we learn from embryogenesis to increase our understanding of how regeneration of damaged adult lung tissue could be induced in serious lung diseases such as chronic obstructive pulmonary disease (COPD), idiopathic pulmonary fibrosis (IPF), and asthma? The local tissue niche determines events in both embryogenesis and repair of the adult lung. Important constituents of the niche are extracellular matrix (ECM) molecules, including proteoglycans and glycosaminoglycans (GAGs). GAGs, strategically located in the pericellular and extracellular space, bind developmentally active growth factors (GFs) and morphogens such as fibroblast growth factors (FGFs), transforming growth factor-β (TGF-β), and bone morphogenetic proteins (BMPs) aside from cytokines. These interactions affect activities in many cells, including stem cells, important in development and tissue regeneration. Moreover, it is becoming clear that the “inherent code,” such as sulfation of disaccharides of GAGs, is a strong determinant of cellular outcome. Sulfation patterns, deacetylations, and epimerizations of GAG chains function as tuning forks in gradient formation of morphogens, growth factors, and cytokines. Learning to tune these fine instruments, that is, interactions between GFs, chemokines, and cytokines with the specific disaccharide code of GAGs in the adult lung, could become the key to unlock inherent regenerative forces to override pathological remodeling. This review aims to provide an overview of the role GAGs play during development and similar events in regenerative efforts in the adult lung.
Introduction
During mammalian lung development, the epithelial cells interact closely with underlying mesenchymal cells, including fibroblasts, to form distal airway structures such as bronchioles and alveoli, so called budding [1]. FGF-10, provided by the mesenchyme, is a key regulator during this event, and is closely monitored by the glycosaminoglycans (GAGs) heparan sulfate (HS) as a coreceptor to direct this signaling [2]. It is established that mesenchymal cells, that is, lipofibroblasts, contribute to alveolar type II homeostasis [3], and most of these cells are derived from FGF-10-positive progenitor cells. Moreover, lung-tissue-resident mesenchymal stromal cells (MSCs) are heterogeneous long-lived cell population known for their immunomodulatory properties [4]. Interestingly, the MSC subpopulation that are CD166− differentiate along the lipofibroblast lineage and show epithelial supportive properties correlating with the level of FGF10 in a transforming growth factor-β (TGF-β)-dependent manner [5]. The FGF-10 pathway, dependent on GAG cosignaling in mesenchymal-epithelial crosstalk, is important during lung development, and accumulating evidence indicates that this pathway is reinstated to support adult lung homeostasis and may be disrupted upon epithelial injury in lung disease [6,7]. Moreover, the GAG hyaluronan (HA) plays an important role in lung development and homeostasis by affecting the survival of alveolar macrophages, which are crucial for debris clearance without eliciting an inflammation [8]. These pathways may be disrupted during chronic lung disease, and several GAGs including HA are constituently high during disease, thus perpetuating an inflammatory response [9,10]. Overall, the turnover of extracellular matrix (ECM) is altered in several chronic lung disorders underlining the importance of ECM homeostasis for functional lung physiology. GAGs activate ECM degrading enzymes and function as storage for growth factors (GFs) and cytokines, which may be released upon tissue injury and needed for cell recruitment to heal damage. GAGs are also conformationally altered due to disease [10], and delineating the turnover and changes in GAG structure in disease may elucidate novel therapeutic avenues. To that end, manipulating cell behavior through engineered GAG-coated 3D matrices is a novel thought that in the future may become reality [11]. Novel ECM-like hydrogels, which are biocompatible, porous, 3D structures supporting cell growth in vivo, are promising new vehicles to induce tissue regeneration and engineering [12].
GAG Structure and Biosynthesis
The indispensable role for GAGs as interaction partners for GFs, chemokines, and cytokines to convey proper receptor engagement is hidden in the composition of their disaccharide fine structure. GAGs have a net negative charge depending on uronic acid residues and sulfate groups, thus binding cations and water, which affect the affinity of GFs. This results in a hydrated gel-like structure in which matrix proteins, for example, collagens, fibronectin, and fibrillin, are embedded. GAGs are built up in disaccharide units with 50–200 repeats, which in turn have a number of variants. Their structural diversity is a result of complex biosynthetic processes involving numerous enzymes [13], which are carrying out a series of reactions specific for different tissues. There are five groups of GAGs: heparin, HS, HA, chondroitin sulfate/dermatan sulfate (CS/DS), and keratan sulfate (KS) (Fig. 1A). The disaccharide composition differs between HS and CS/DS, where HS is comprised of repeating disaccharide units of N-acetylglucosamine (GlcNAc) and a form of uronic acid; either glucuronate (GlcA) or iduronate (IdoA), compared with CS/DS, which exhibits N-acetylgalactosamine (GalNAc) and GlcA or IdoA (Fig. 1A) [14,15]. Both HS and CS/DS are composed of blocks with similar disaccharides; in HS low- and high-sulfated blocks are found, whereas in CS/DS the blocks are rich in either IdoA-GalNAc-4S or monosulfated GlcA-GalNAc-4/6S. In addition, both glycans contain additional sulfates, which endow specific functions to the polysaccharides. Thus, it has been shown that sulfated IdoA blocks have an important role in mediating the binding to ligands such as fibroblast growth factor 2 (FGF-2), hepatocyte growth factor (HGF), and heparin cofactor II (Fig. 1B) [16 –18]. Variation in sulfation patterns of the GAGs affects not only the binding of GFs but also cellular response resulting from downstream signaling from their cognate receptor [19]. In fact, specific sulfation in position 6 of D-glucosamine of HS GAGs is essential for mitogenic activity [20]. In line, ablating the GlcNAc N-deacetylase/N-sulfotransferase-1 (NDST-1), which is responsible for sulfation of HS, resulted in lethal respiratory distress due to epithelial type II cell immaturity [21]. We have shown that the enzymes dermatan sulfate epimerase 1 and dermatan 4-O-sulfotransferase colocalize to the Golgi, and they physically interact in the process of forming long epimerized 4-O-sulfated blocks in the CS/DS chain [22]. These alterations directly affect differential interactions with their binding partners, thereby immediately stressing their specific activity [23,24]. Interestingly, the content of iduronate in the CS/DS GAG chain affects proliferation of fibroblasts [25].
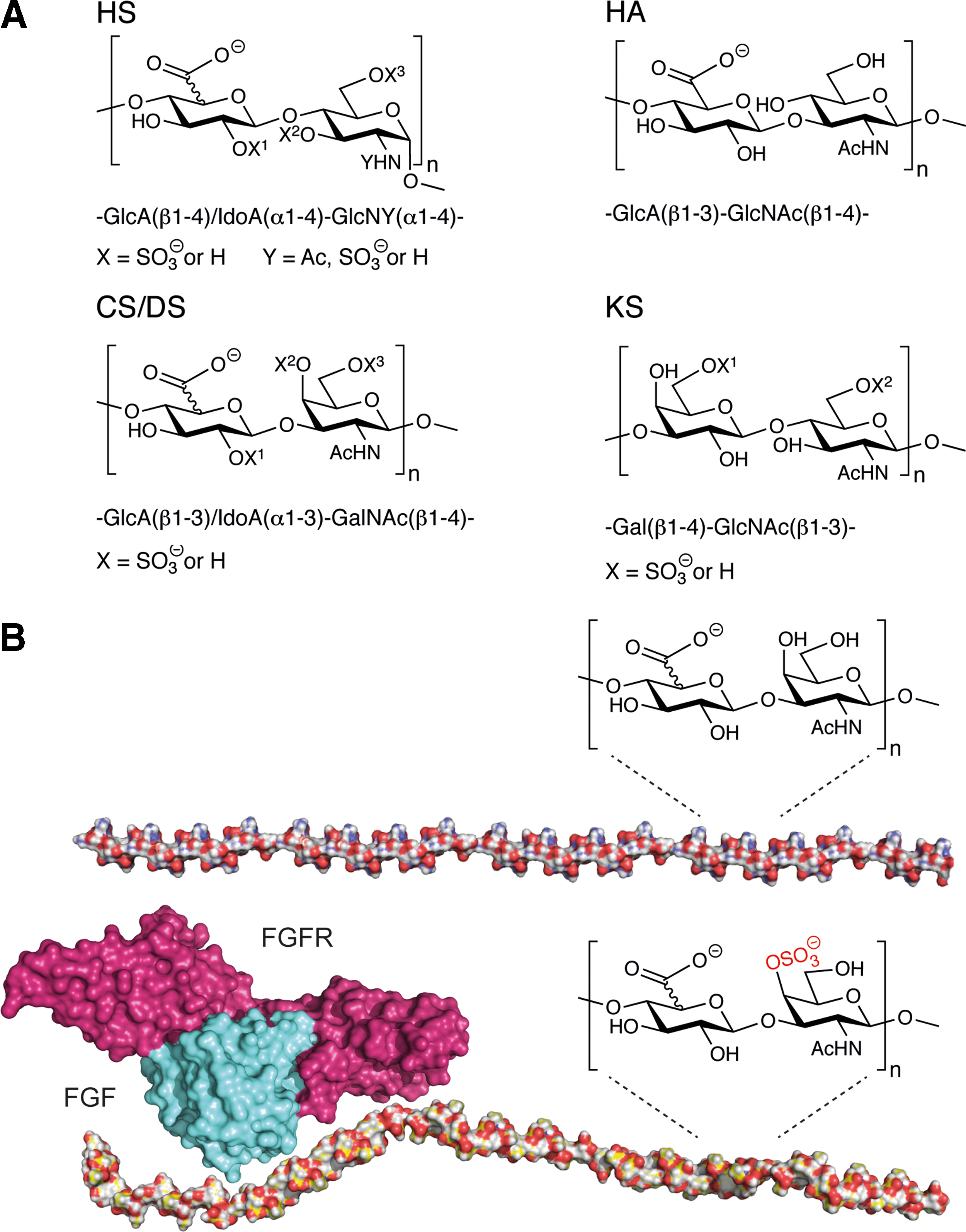
We have shown that the disaccharide composition of the GAGs is a determinant of its biological effect, and alterations in the chain by specific enzymes affect cellular response such as apoptosis [26]. Interestingly, we have shown that interacting GFs or cytokines such as TGF-β can affect the fine structure of CS/DS and HS both concerning sulfation and epimerization [27,28]. Taken that TGF-β is an important player during fibrotic events in lung disorders, this finding may have implications in the negative feedback loop leading to fibrosis in idiopathic pulmonary fibrosis (IPF), chronic obstructive pulmonary disease (COPD), and asthma. Indeed, we have shown that highly sulfated HS was located in the border zone between fibrotic areas and more normal looking alveolar parenchyma in IPF [10]. Taken together, the fine structure of GAGs highly affects the binding to cytokines, chemokines, and GFs.
Proteoglycans and GAGs Are Ubiquitous Fine Tuners of Cell Response
GAGs are found freely or covalently attached to the core protein of proteoglycans and depending on the degree of sulfation providing specific binding motifs for GF interaction, different downstream signaling occurs [29]. GFs such as TGF-β interact with both the core protein and the GAG side chains of proteoglycans [30]. Proteoglycans are structurally diverse glycosylated proteins found throughout the body. They exist in cell-bound forms or secreted into the ECM. For instance, glypicans and syndecans are present at the cell surface, being attached to the plasma membrane through a glycosyl-phosphatidyl inositol anchor or a transmembrane domain, respectively [31,32]. In contrast, perlecans are secreted and distributed mainly in the ECM, specifically in the basement membrane of alveolar and blood vessel structures [33]. GAGs are crucial binding partners for some chemokines, and CXCL8 (interleukin 8) is involved in neutrophil recruitment to inflammatory sites. Interestingly, the GAG–IL-8 interaction, apart from establishing a chemokine gradient for neutrophil attraction, has an effect on downstream signaling of endothelial cells such as cytoskeletal organization, cell adhesion, and cell mobility [34]. The formation of chemokine gradients enables a GAG-mediated mechanism for spatially restricting the engagement between the chemokine or GF and its receptor near the source of mediator production [35]. In this way, GAGs are thought to control a spatial and temporal release of factors upon direct receptor interaction. The spatial and temporal control GAGs have on GF release and engagement with their receptors is of major importance during embryonic development and also in adult lung homeostasis and regeneration (Fig. 2). FGF2R is expressed throughout the airway tree during lung development as well as its ligand FGF-10. Importantly, FGF-10 is only chemotactic for distal airways, not for proximal epithelium. This is due to O-sulfated HS as critical binding partner to FGF-10 to restrict interaction to the distal epithelium (Fig. 1B) [2].

Illustration of parallel involvements of GAGs and growth factors in lung development
GAGs Are Important During Development
Disruption of HS biosynthesis during airway development results in malformed lungs in mice and obstruction of normal tracheal branching in Drosophila [36]. As mentioned above, HS modulates FGF signaling during embryonic lung development. For instance, digestion of HS in embryonic week 11.5 lung epithelial explants results in cell death, similar to when they are cultured without FGF1. This suggests that lack of HS results in failure in FGF signaling, which leads to impaired lung morphogenesis [37]. HS, as coreceptor for FGF-10 signaling, is also required for airway branching [2]. Importantly, binding of FGF-10 to HS is highly specific and dependent on precise modification patterns of HS [38]. Similarly, in Drosophila, mutations in genes encoding HS sulfation modification enzymes hamper FGF signaling, resulting in disruption of tracheal morphogenesis [39,40]. Moreover, epithelial HS has been shown to be a key regulator for dictating Sonic Hedgehog (SHH) signaling linking to the importance of FGF-10 during lung morphogenesis. When HS was depleted in a biosynthetic step that involves attachment of altering GlcNAc and GlcA units to the growing HS chain, the biologically active form of SHH was lost and thereby also proper expression of FGF-10. This led to tip expansion and reduced branching [41]. The importance of HS in lung development and repair is fortified by a large amount of studies [42], but the role of CS/DS is less explored. Studies on rat lung development point out the importance of CS proteoglycans and sulfation of these in growth and branching morphogenesis [43]. The action of sulfatases determining the degree of sulfation of GAGs is crucial for correct GF signaling during alveolarization [44]. To go further in detail regarding GAG and GF structure interaction, it has been shown that epimerizations are important in this interplay. Interestingly, it has been shown that the interaction of defined structures of CS/DS with FGF-10 is important for proliferation and migration of epithelial cells, and subsequently re-epithelialization for rapid repair of burn wounds [45]. The direct function of CS/DS binding to FGF-10 for cell signaling is not clear but as a strong anion, CS/DS and also HS may stabilize FGF-10 and improve its functionality [46]. Similar to FGF-10, the lung developmental factor FGF-2 interacts with HS and DS with repeats of IdoA-GalNAc4S in the same manner [18]. In contrast to the FGFs, the developmental factor HGF just generally binds to GAGs with a wider variety of sulfation patterns. HGF requires a minimum binding sequence of disulfated trisaccharide comprised of an internal iduronate of variable sulfation pattern. However, additional sulfation further enhances the affinity of HGF [47]. In addition, it has been shown that CS/DS binds HGF, and that esophagus squamous carcinoma cells deficient in IdoA-containing CS/DS exhibit decreased migration and invasion capabilities in vitro [48]. Important factors for the branching program that interact with GAGs are the WNTs and TGF-β/BMP/activin pathways controlling cell proliferation, differentiation, polarity, and linage specification. More specifically, not only 2-O-sulfation but also 6-O-sulfation has been pointed out as important structures in HS for WNT interactions, especially for WNT5a [49,50]. These structures are found on membrane-bound proteoglycans such as betaglycan and syndecans, which also bind all three TGF-β isoforms, bone morphogenetic protein (BMP), and activin. Thus, several developmental factors closely interact with each other, and a common link is the GAG structure. This is illustrated in one of many studies where overexpression of WNT5a affects both FGF-10 and SHH signaling, resulting in disruption of epithelial–mesenchymal interactions and reduced epithelial branching and dilated distal airways [51,52]. WNT3a has been described to bind not only to HS but also to CS/DS, and the precise interaction is represented by the 4,6-O-sulfated E disaccharide unit of CS [53]. Interestingly, both WNT5a and WNT3a have been found in the border zone where an active regeneration and repair take place in lung tissue from IPF patients [54] together with regions of HS containing 2-O- and 6-O-sulfated disaccharides [10].
Another important factor that is critical for embryonic and lung development is CXCL12 (SDF-1) that has been described to bind to HS/heparin [55]; however, the precise structure for this interaction has at present not been described. Taken together, several of the pathways involving GAG–GF interactions during embryonic lung development have been shown to be important during adult lung homeostasis, remodeling, and regeneration (Fig. 2). This may give cues on how to approach lung regeneration in future therapeutic development.
The Role of GAGs in Regeneration of Human Lungs
In line, we and others have identified factors in lung tissue from patients with COPD that are active during both development and adult lung regeneration, such as HGF, TGF-β, and vascular endothelial growth factor [56 –60]. We found that these factors are increased in early but not late stages of the disease [57], which may point toward a regenerative response to insults (eg, smoke) that the body is trying to heal. Moreover, SDF-1, shown to interact with HS, is an important homing factor for recruiting fibrocytes from blood into tissue, and may be involved in repair during pathological changes in IPF, bronchiolitis obliterans syndrome (BOS), and asthma [61 –63]. The downstream signaling from these factors on cell activity is dependent on interaction with the specific code or sulfation pattern in the GAGs (Fig. 1B) [28]. Therefore, the specific disaccharide structure of the GAG determines the cell output in development and regeneration. For example, specific CS sulfation motifs interact with proteoglycans on activated stem/progenitor cells in areas of tissue development and also in tissue repair [64]. In fact, one study showed that silencing sulfotransferase 15, which generates C6-sulfation of GalNAc on CS GAGs, inhibited the development of murine pulmonary fibrosis [65]. In line, the developmentally and regeneratively active FGFs interact with specifically sulfated disaccharide motifs in HS and CS/DS to convey mitogenic activity (Fig. 1B) [66]. Studies suggest that FGF-10 plays a protective role in diverse acute lung injuries [67]. A precise interaction between FGF-10 and specific patterns in the GAG has been determined to enclose IdoA and 4-O-sulfated GalNAc within oligosaccharide structures of ∼5–10 disaccharide units of DS to convey maximal capacity to enable FGF-10 receptor stimulation, where DS has a greater potency than HS in the interaction with FGF-10 [68].
The enormous flexibility in modifications of GAGs concerning structure, sulfation, and epimerization plays an important role in lung diseases. It has been shown that specific patterns in GAGs, including sulfation, are altered in the disease progression of asthma and pulmonary fibrosis [69,70]. In line, we have found that HS, CS/DS, and HA were increased in IPF lungs along with a relative increase in sulfation of HS and CS/DS. Interestingly, the highly sulfated HS was found in the fringes of fibrotic areas bordering normal tissue. The changes colocalized with perlecan in basement membranes as well as in the alveolar interstitium [10]. Concurrently, we have shown that the CS/DS-associated proteoglycan biglycan stimulated migration of lung-derived fibroblasts [71] and was highly expressed in the alveolar compartment in lung-transplanted patients who later developed BOS [72]. Furthermore, biglycan, another proteoglycan associated with CS/DS, was increased in uncontrolled asthma along with collagen, versican, and decorin [73]. Further, we have shown that IdoA structures in CS/DS have an impact on migration of smooth muscle cells. However, more studies are strongly warranted to determine the exact role of the fine structure within GAGs in lung pathology. In Fig. 2, we depict a potential scenario for lung regeneration in the adult lung.
A recent study illustrates that intratracheal delivery of FGF-10 to rats results in mobilization of lung-resident MSCs. Upon transplantation of FGF-10-pretreated MSCs to LPS-injured rat lungs, lung injury and inflammation were significantly reduced [74]. Recently, it was shown that FGF-10 also plays a pivotal role in epithelial stem cell regulation in the lung. Basal stem cells were shown to establish their own niche through the expression of FGF-10 by airway smooth muscle cells. Upon epithelial injury, the Hippo pathway upregulates intracellular Yap giving rise to the release of Wnt7b, which induces the generation of FGF-10 in the niche airway smooth muscle cells. FGF-10 interacts with its receptor FGFR2b on basal stem cells leading to their expansion and ability to regenerate the damaged epithelium [75]. These findings indicate that GAGs are important for stem cell behavior during lung disease.
The Importance of GAGs for Stem Cell Fate
The involvement of GAGs in stem cell fate has been touched upon above, and more evidence shows that GAGs are altered in proteoglycans such as lumican during embryonic stem cell (ESC) differentiation. These alterations correlate with changed transcript levels of HS biosynthetic enzymes [76]. HS has proven to be critical for lineage commitment of ESCs, and lack of HS results in failure of ESC differentiation. When heparin was administered to HS-deficient (Ext1−/−) ESC, differentiation capacity in ESC was restored by conferring the correct responsiveness of signaling molecules such as WNT and BMPs [77]. As previously discussed, WNT has a preponderance for 2-O-sulfation but also 6-O-sulfation [49,50] and for ESC differentiation also N-sulfation was essential, suggesting that the structure of the chain domains and organization of sulfate groups delicately control important cues for proper ESC differentiation [78]. Not only does sulfation pattern, which may alter the net charge of GAGs, affect the affinity for GFs but also the length of the GAGs. Shorter sequences (12-mer) provided a greater effect on differentiation of stem cells than longer chains [79]. The mechanism behind this is multifaceted, as HS binds multiple signaling cues involved in the balance between self-renewal and differentiation. However, evidence suggests that HS is required for proper cell surface binding of FGF and downstream intracellular signaling, without which ESCs fail to exit from their self-renewal state [80]. HA, on the contrary, has proven to be important for preserving MSC quiescence and maintenance of stemness. Interestingly, when MSCs were treated with HA a delayed cellular aging was observed [81]. Lung MSCs, other mesenchymal cells, and epithelial stem cells and their niche give promise to utilize their inherent milieu to induce differentiation into cell types needed to restore lung function. The ECM niche has in many studies been shown to be crucial for stem cell function, and the development of artificial ECM niches would enable a new era of stem cell management to treat lung disease. In line, altering the sulfation pattern in GAGs incorporated into a hydrogel used as 3D cell scaffold, the cell fate could be intricately affected. This was evidenced by a study showing that morphogen gradients of the sulfation pattern controlled the affinity of GFs, and therefore also the release and bioavailability for cells. In this way, it was possible to direct cells toward a specific fate [82].
Tissue Engineering for Treatment of Lung Disease
The ability of adult human lung growth and alveolarization after pneumonectomy has been demonstrated in a case report study in which exercise and the induced stretch were relevant contributions to the success [83]. Pneumonectomy has in animal models been associated with dilation of alveolar structure but not the growth of more alveoli [84]. This indicates that the human lung has an inherent capacity to self-new with the right type of stimuli such as the right combination of signaling factors, progenitor cells, and biomechanical loading as in exercising. An exciting field of novel therapeutic strategies is tissue engineering using modified acellular ECM scaffolds. FGF-2 was in one study shown to hamper cardiac myofibroblast contraction of such scaffold, and this effect was augmented by combining heparin with FGF-2. Heparin was shown to enhance the effect of FGF-2 as an antifibrotic factor [85]. Consequently, the regulatory properties of GAGs may influence regenerative processes in the adult lung, as GF pathways such as the FGF circuitry become reactivated after injury. Moreover, the need for developing novel approaches to serious lung diseases, which are currently treated with lung transplantation, is urgent. For this reason, the lung bioengineering field is promising. Moreover, the age of the cells producing the ECM plays an important role in stem cell decision. Old cells grown on ECM produced by young cells exhibited a clearly enhanced proliferative capacity and intriguingly their capacity to form tissues [86]. In line, aged mice exhibit decreased basal levels of both FGF-7 and FGF-10, which are linked to a blunted wound healing [87]. This pathway could be one of many links to delayed healing in elderly persons. It is furthermore known that there is a shift in 4-O-sulfation to 6-O-sulfation in elderly [88].
Improving treatment of serious lung diseases is an essential task in medicine. It has, however, been slow due to discrepancies between in vivo/in vitro models and the human diseases. Recently, progress in the bioengineering field enables complex 3D environments to mimic the lung [89]. In these settings, cell behavior can be evaluated in a milieu as close to lung as possible. We have shown that the ECM influences cell fate, and may be a trigger for a negative feedback loop in disease and impaired regenerative ability. Normal human bronchial epithelial (NHBE) cells cultured on lung ECM derived from COPD patients induced differential gene expression associated with COPD pathophysiology, which was not seen for NHBE cultured on normal ECM. Likewise, we have demonstrated that lung ECM derived from IPF patients induced normal human primary fibroblasts to produce more of already upregulated proteins and less or none of under-represented proteins, glycoproteins being one group, compared with fibroblasts cultured on normal lung ECM (unpublished data).
With accumulating evidence that GAGs play a vital role in regeneration and repair, there is an increasing interest in developing biomaterials, both biological and synthetic, with incorporated GAGs for controlled delivery of GF cytokines, and progenitor cells. In Fig. 3, we speculate on the potential regenerative properties of such biomaterial incorporated into emphysematous lung.

A hypothesis of how a biomaterial distributed to the emphysematous lung may act upon the border zone between healthy
There are already numerous materials being produced with GAGs for bioengineering, and their structural, physiochemical, and biological characteristics have been meticulously reviewed elsewhere [90,91]. The fact that GAGs sequestrate positively charged factors in vivo enables a plethora of opportunities to design biomaterials with different compositions of GAGs to introduce gradients, modulate signaling, and to mimic the natural environment [92]. In fact, efforts are now being made to produce GAGs with specific motifs that bind GFs and cytokines. This library of recombinantly expressed proteoglycans with specified GAG structures and well-defined GAG chains is referred to as GAGome [13]. This will be an invaluable addition in the design of to-the-point biomaterials that exert specific instructions to cells. Forming microhydrogels from GAGs is a novel way of creating a system for controlled release of cytokines [93]. Engineered biomaterials may serve as an in vitro model that recapitulates the human lung structure and biomechanics. It is a promising future therapeutic approach for lung diseases, which provides an artificial “healthy” scaffold. Loading these scaffolds with cell-instructive factors (GFs, cyto/chemokines, and progenitor cells) may enable lung-resident cells toward regeneration of damaged tissue.
HA-based biomaterials have long been used to induce chondrogenesis [94]. In a study using CS, it was shown that human MSCs encapsulated in a 3D hydrogel made up of synthetic polyethylene glycol and CS were able to differentiate in a more controlled manner upon biomechanical loading, and avoiding terminal differentiation into hypertrophic cartilage [95]. Altering the sulfation pattern is another way to control the sequestration of GF and thereby the cellular response. By increasing the degree of sulfation in a collagen biomaterial, using HA, CS, and heparin, and incubating with the same amount of insulin-like growth factor 1 (IGF-1), it was shown that the metabolic activity of equine tenocytes for tendon regeneration was enhanced in a similar dose-dependent manner as increasing soluble levels of IGF-1 [96]. A hydrogel using CS of different sulfation patterns, using monosulfated CS-4 and CS-6, as well as disulfated CS-4,6 was developed to create an endogenous niche for neural stem cells (NSCs). The sulfated CS of different grades bound trophic factors FGF-2, BDNF, and IL-10 in the hydrogel, and the material as a carrier of NSC was shown to improve NSC self-renewal and prolong survival rate of the cells [97]. In a heparin-based hydrogel, researchers have been able to sequester chemokines in an opposite manner for wound closure. Their hydrogel bound and neutralized chemokines released from injured tissue and immune cells, including tumor necrosis factor (TNF), IL-1β, IL-6, MCP-1, and IL-8. By lowering the concentration of the proinflammatory mediators, the inflammatory response was attenuated and the wound was healed. Again, specific sulfation pattern was of importance for optimal effects [98]. As lung has a remarkable capacity to regenerate in normal conditions, this opens up the possibility to construct a material with the right combinations of factors to trigger regeneration in the diseased state.
Conclusion
In summary, GAGs are intimately involved in the regulation of morphogen gradients and GF signaling during development. However, given the diversity in GAGs, both temporally and spatially during development, the precise structure–function relationship between these macromolecules and cell fate decisions remains elusive. Transfer of information from developmental pathways to local repair and regeneration of the tissue niche will open up new clues for understanding disease, diagnosing and developing new therapies. During the last decade, the bioengineering field has flourished and now produce biomaterial with incorporated signaling molecules to be used in diverse fields, such as diabetes and cancer. By combining specific structures of GAGs and their potential to bind cytokines and GFs to modulate biological activity, it will be possible to fine tune effects on the local tissue niche; thus, opening up the therapeutic window. Conclusively, we are entering a new and exciting era with new techniques rapidly developing, to study advanced and detailed structures of carbohydrates and GAGs combined with increased options with single-cell techniques to isolate tissue-specific stem cells. Together, this will really raise interesting and novel chances for the future in regenerative medicine and repair of the adult lung.
Footnotes
Author Disclosure Statement
The authors have no actual or potential commercial associations that might create a conflict of interest in association with the publication of this article.