
Abstract
Mammary gland stem cells (MaSCs), assumed to be the original cells of breast cancer, play essential roles in regulating mammary gland homeostasis and development. Previously, we identified a crucial regulatory role of TAR DNA-binding protein 43 (TDP-43), an RNA-binding protein, in the progression of triple-negative breast cancer. However, the function of TDP-43 in MaSCs is unclear. Based on single-cell data analysis of the mammary gland, TDP-43 showed potential involvement in the regulation of MaSCs. We therefore investigated the effects of TDP-43 on the mammary gland development. Our data both in vitro and in vivo demonstrated that TDP-43 was required for the mammary gland repopulation, which suggested the potential role in the regulation of MaSCs. Knockdown of TDP-43 inhibited proliferation of mammary epithelial cells (MECs) and mammary morphogenesis. RNA-seq data and other experiments identified that loss of TDP-43 induced the upregulation of genes related to the cell cycle, providing a possible mechanism for TDP-43 in regulating mammary gland repopulation. Thus, our findings indicate a previously unknown role of TDP-43 in MECs.
Introduction
Mammary epithelial cells (MECs) are mainly composed of basal and luminal cells. During puberty and subsequent pregnancies in females, the mammary gland undergoes dramatic changes in structure and function, which depend on the function of mammary gland stem cells (MaSCs) and progenitor cells [1,2]. MaSCs possess the ability to self-renew and differentiate into various cell types. Operationally, cells that can generate ductal-lobular epithelial outgrowth when transplanted into cleared fat pads at limiting dilutions are termed mammary repopulating units mammary repopulating units (MRUs) [3]. Both the Notch and Wnt signaling pathways regulate stem cell self-renewal and cell-fate decisions [4,5]. Studies have shown that several transcription factors also play essential roles in regulating the proliferation and differentiation of mammary stem cells and progenitor cells, such as Myc and p63 [6,7]. Since elucidation of the molecular hierarchy of MaSCs is critical to understand the molecular perturbation driving breast cancer [8,9], more molecules are necessary to explore their functions in regulating MaSCs.
TAR DNA-binding protein 43 (TDP-43), encoded by the Tardbp gene, belongs to the heterogeneous nuclear ribonucleoproteins (hnRNPs) family and consists of 414 amino acids with two RNA recognition motifs and a carboxy-terminal glycine-rich domain [10]. Members of the hnRNP family are involved in various cellular pathways such as DNA transcription, premessenger RNA (mRNA) splicing, and mRNA export and import [10,11]. Several studies had identified that aberrant TDP-43 is associated with amyotrophic lateral sclerosis and frontotemporal lobar degeneration [12 –15]. The involvement of TDP-43 is also essential for embryonic development. Research has shown that homozygous deletion of Tardbp results in severe mouse lethality during embryonic stages due to defective outgrowth of the inner cell mass [16,17]. Moreover, TDP-43 is crucial for embryonic stem cell (ESC) survival, with the conditional deletion of Tardbp in ESC known to cause inhibition of proliferation [18]. Our group previously showed that TDP-43 is highly expressed in triple-negative breast cancer progression, and loss of TDP-43 in human breast cancer cell lines can lead to inhibition of cell growth and cell cycle arrest at the G1 phase [19]. These results prompted our exploration of the function of TDP-43 in mammary gland development.
In this study, we identified the involvement of TDP-43 in regulating mammary gland repopulation. Knockdown of TDP-43 inhibited clone formation in vitro. Inducible deletion of Tardbp exhibited diminished mammary gland ductal branching and cell proliferation. Based on RNA-seq analyses, we identified that genes regulating the cell cycle were upregulated following knockdown of TDP-43 in primary MECs. Overall, we demonstrated that TDP-43 plays an essential role in mammary glands.
Materials and Methods
Mice
Transgenic mice harboring the floxed Tardbp allele were generated as reported previously [20]. K14-CreERT mice were obtained from the Nanjing Biomedical Research Institute of Nanjing University, China. All experimental procedures and animal care and handling were performed as per the protocols approved by the Ethics Committee of the Kunming Institute of Zoology, Chinese Academy of Sciences.
Cell culture
The HC11 cell line derived from MECs of Balb/c mice at midgestation were cultured in RPMI-1640 medium with 10% fetal bovine serum (FBS), 5 μg/mL insulin (Sigma, I5500), 10 ng/mL epidermal growth factor (EGF; Gibco, PGH0315), and 5 μg/mL gentamycin sulfate. Primary MECs were dissociated as described previously [21], and the cells were cultured in DMEM/F12 medium with 2% newborn calf serum (NBCS), 10 ng/mL EGF (Gibco, PGH0315), 4 μg/mL heparin solution (Stemcell, 07980), 10 ng/mL basic fibroblast growth factor (bFGF), 5 μM Y-27632 (Selleckchem, S1049), 0.5 μM BIO (Selleckchem, S7198), and 1% penicillin streptomycin. All cells were incubated with 5% carbon dioxide (CO2) at 37°C in a cell culture incubator.
Flow cytometry analysis
For flow cytometry analysis, 1 × 106 cells were resuspended in phosphate-buffered saline (PBS) with 2% FBS and blocked with Mouse BD Fc Block (BD Pharmingen, 553142) for 10 min. The cells were then incubated with primary antibodies against CD31 (biotin-conjugated; BD Biosciences), TER119 (biotin-conjugated; BD Biosciences), CD45 (biotin-conjugated; BD Biosciences), CD24 (PE-conjugated; BD Biosciences), and CD29 (FITC-conjugated; BD Biosciences) on ice in the dark for 30 min. Cells were next washed with PBS buffer containing 2% FBS and stained with PE-Cy7 streptavidin on ice for 30 min. Fluorescence-activated cell sorting (FACS) was performed by flow cytometry, and data were analyzed using FlowJo software.
Bromodeoxyuridine staining
Cells were seeded in 24-well plates. After incubation with bromodeoxyuridine (BrdU) solution at 37°C for 45 min, the cells were fixed with paraformaldehyde for 15 min at room temperature. The cells were then treated with HCl, blocked with goat serum, incubated with BrdU antibody (Cell Signaling Technology, 5292) for 8 h at 4°C, and finally incubated with a second antibody for 1 h at room temperature. The nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI).
Quantitative real-time PCR qPCR
Total RNA was extracted from cells using TRIzol reagent (Life Technologies). Amplification of complementary DNA (cDNA) was performed using a PrimeScript RT reagent kit (TaKaRa) according to the manufacturer's instructions. cDNA was used to perform qPCR with a SYBR Green PCR Master Mix (Applied Biosystems) and QuanStudio 3 Applied Biosystems (Thermo Fisher Scientific). Primer sequences are listed in Supplementary Table S1.
Cell proliferation and cell cycle assay
The viability of cells was assessed by MTS using the CellTiter 96 AQueous One Solution Reagent (Promega, G3581) according to the manufacturer's protocols. For cell cycle analysis, cells were fixed in 70% ethanol for 48 h at 4°C and washed with PBS. Cells were next incubated in PBS with 1 mg/mL RNase A, 100 mg/mL PI, and 0.6% Nonidet P-40 for 30 min at 37°C. Finally, cell cycle distribution was measured by flow cytometry (LSR Fortessa; BD Biosciences), and data were analyzed using FlowJo software.
Mammosphere assay
Cells were seeded into 96-well ultralow attachment dishes at an initial density of 2000 cells per well and cultured in EpiCult-B Basal medium (Stemcell), supplemented with 5% FBS, 5% Matrigel matrix (Corning), 20 ng/mL bFGF (Sigma-Aldrich), 10 ng/mL EGF (Sigma-Aldrich), 4 μg/mL heparin, 10 ng/mL EGF (Gibco, PGH0315), and Y-27632 (Selleckchem, S1049) for 7 d with 5% CO2 at 37°C.
Mammary gland transplantation
Primary MECs were resuspended with 50% Matrigel in PBS, then injected into cleared mammary fat pads. Limiting dilution assay was performed with the indicated cell numbers. After 6–8 weeks posttransplantation, the mammary gland tissues were obtained to examine outgrowth and reconstitution frequency.
Short hairpin RNA-mediated knockdown
The short hairpin RNAs (shRNAs) targeting TDP-43 were inserted into the cloning vector (pLKO.1) and sh-GFP (Addgene No. 30323) and sh-Control (Addgene No. 10879) vectors were obtained from Addgene. The shRNAs were transfected with psPAX2 and pMD2.G (4:3:1) into 293T cells to produce lentiviral particles. The produced shRNA lentiviral particles were concentrated by ultrafiltration (25,000g) within 20% sucrose in the bottom of virus suspension and resuspend in 0.1% bovine serum albumin. After testing the viral titers, the concentrated lentiviral particles were used to infect target cells in appropriate virus multiplicity of infection (MOI) for 12 h, following changes with normal cell culture medium. After 72 h, cells were collected, and knockdown efficiency was verified by real-time qPCR and western blot analysis. The shRNA sequences are listed in Supplementary Table S1.
Western blot analysis
Cells were treated in lysis buffer with protease inhibitor (1:1000) for 30 min on ice. Protein lysates were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. The membranes were blocked by 5% nonfat milk diluted in phosphate buffered saline with Tween 20, then probed with specific primary antibodies at 4°C overnight, and detected with chemiluminescent horseradish peroxidase (HRP) substrate (Millipore). The antibodies used for immunoblotting were α-tubulin (Sigma, T5168), TDP-43 (Abcam, ab109535), and P21 (Abcam, ab109199).
Whole-mount staining
Whole mammary glands were fixed in 75% ethanol and 25% acetic acid for 1 h and stained with carmine alum solution at 4°C overnight. The following day, tissues were treated in a series of graded alcohol for dehydration, then cleared in xylene, and mounted with Permount.
Histology and immunofluorescence
Mammary glands were fixed in 10% formalin, then embedded in paraffin, sectioned, and stained with hematoxylin and eosin (H&E). For immunofluorescence, sections were blocked with 10% goat serum in PBS for 2 h, incubated with primary antibody at room temperature for 2 h, then incubated with secondary antibody for 1 h. DAPI was used to stain nuclei. The antibodies used for immunoblotting were TDP-43 (Abcam, ab109535) and Ki67 (Abcam, ab15580).
RNA-seq analysis
The RNA library was constructed following the Illumina library preparation protocols. High throughput sequencing was performed on a HiSeq2500. Sequenced reads were mapped to the mouse genome (mm10) using hisat2 (v2.1.0) with the following parameters: “–no-softclip-k 20 –dta –dta-cufflinks – score-min L, 0, -0.4 –no- discorda- nt –known-splicesite-infile.” The gene annotation file in gtf format was downloaded from the Ensembl website (release 91). FeatureCounts was used to generate the count matrix with the following parameters: “-g gene name -p.” Differential expression analysis was performed using edgeR (v3.24.0). Genes with a P value <0.05 and fold-change >2 were regarded as differentially expressed. Gene ontology (GO) analysis was performed using DAVID bioinformatics tools.
Data deposition
The data reported in this article were deposited in the Sequence Read Archive database (
Results
TDP-43 expression is positively correlated with MaSCs
To identify potential members in MaSCs regulation, we analyzed single-cell sequencing data using published data [22,23]. Results suggested that TDP-43 was highly expressed in mammary gland epithelial cells on embryonic day 18 (E18) and in some adult protein c receptor positive (Procr+) cells (Fig. 1A, B). Previous studies have shown that the frequency of MaSCs at embryonic day 18.5 is higher than that in the adult gland, that is, fetal MaSCs peak during the late phase of embryogenesis [24], and Procr+ cells are a mark of multipotent mouse MaSCs [25]. Our single cell RNA-seq analyses indicated that TDP-43 is possibly involved in the regulation of MaSCs. To further characterize the expression of TDP-43, mammary gland tissues were obtained from virgin female C57BL/6 mice for immunofluorescence staining with an antibody against TDP-43. Results showed that TDP-43 was ubiquitously distributed in the mammary glands of virgin mice (Fig. 1C). We next sorted basal (Lin-CD24+CD29high), luminal (Lin-CD24+CD29low), and stromal-enriched cell subpopulations (Lin−CD24lowCD29low) from adult mice by FACS (Fig. 1D). Results indicated that TDP-43 was highly expressed in basal cells enriched with MaSCs and in luminal cells (Fig. 1E). Overall, our expression data indicated that TDP-43 may play a role in mammary gland development.
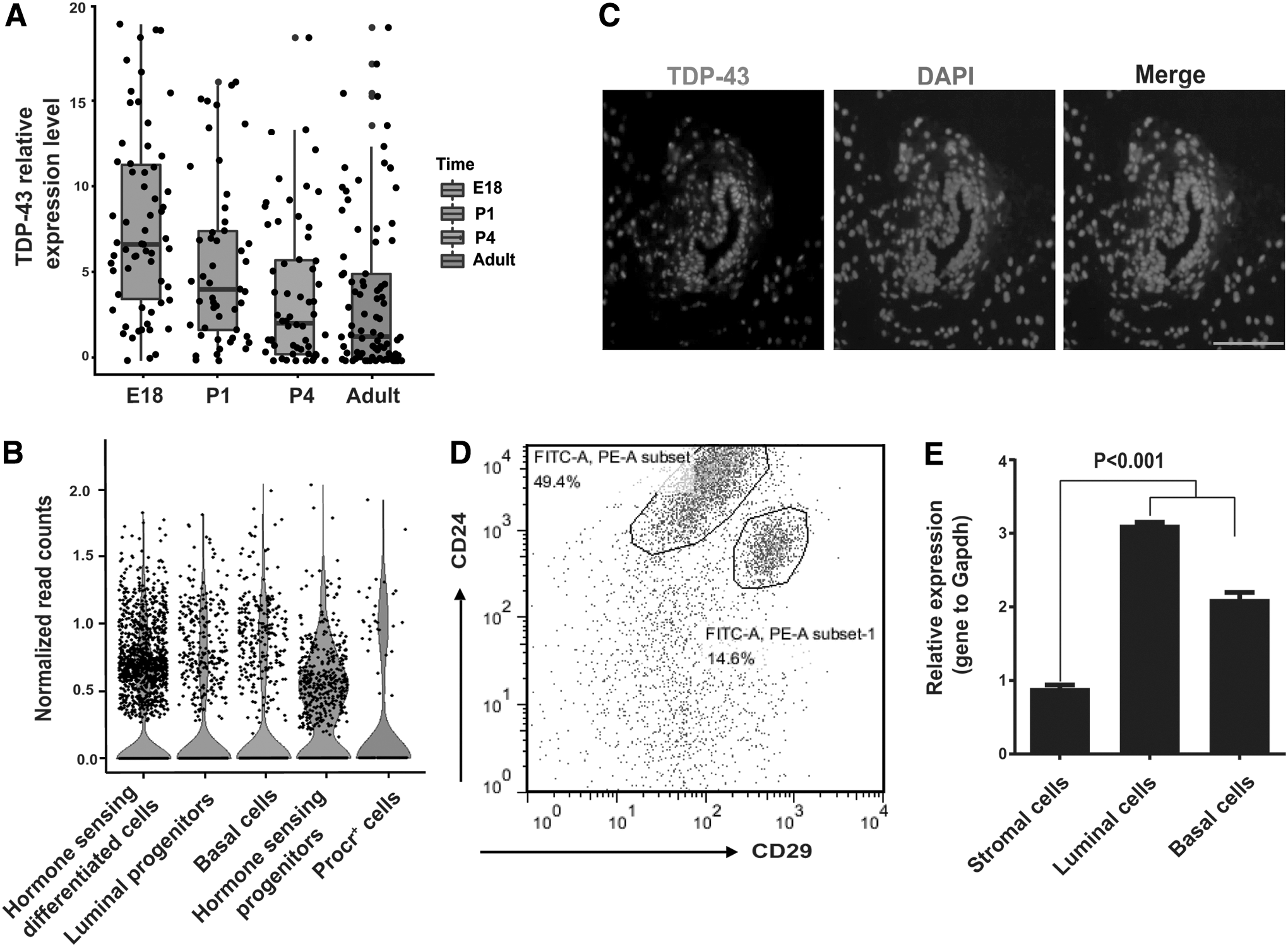
TDP-43 expression is positively correlated with MaSCs.
Knockdown of TDP-43 reduces mammary gland repopulation
To investigate the potential role of TDP-43 in mammary gland repopulation units (MRUs), we used concentrated lentiviral particles with MOI = 50 to disrupt the expression of TDP-43in both primary MECs and HC11 cells. The knockdown efficiencies were validated in primary MECs and HC11 cells (Fig. 2A; Supplementary Fig. S1A). The colony numbers of mammospheres were remarkably reduced in both primary MECs and HC11 cells (Fig. 2B, C). We also applied MOI = 20 or MOI = 100 to infect primary MECs, and found the similar phenotype (Supplementary Fig. S1). To further characterize the importance of TDP-43 for maintaining repopulation capacity, we performed cleared fat pad repopulation assay. Primary MECs treated with shRNA were transplanted with limiting dilution into the cleared fat pad of 3-week-old NOID/SCID mice. Whole mount analysis revealed that knockdown of TDP-43 in MECs generated less mammary reconstitution and ductal outgrowth than their control groups (Fig. 2D). Collectively, these results suggest that the down-regulation of TDP-43 inhibits mammosphere formation and mammary reconstitution.

TDP-43 downregulation decreases repopulation of MECs
Deletion of Tardbp inhibits mammary morphogenesis
We next examined whether TDP-43 plays a role in promoting mammary morphogenesis. To bypass embryonic lethality of Tardbp-null mice [20], we disrupted TDP-43 in an inducible Tardbp knockout mouse line (K14-CreERT; Tardbp fl/fl) by crossing the floxed Tardbp allele [20] with transgenic mice expressing a Cre recombinase fused with the tamoxifen (TAM) responsive domain of estrogen receptor (ERT) under the control of the K14 promoter (K14-CreERT). The postnatal transgenic mice did not yield active Cre recombinase until TAM was administered [26] (Fig. 3A). To disrupt the expression of TDP-43 in virgin mammary glands, we administrated 6 mg/25 g body weight of TAM (three injections every other day) to 4-week-old mice [27] (Fig. 3B). The deletion efficiency of TDP-43 in MECs was confirmed by immunofluorescence, and results showed that the Tardbp gene was partially deleted in MECs (Fig. 3C). We also costained with a marker of mammary epithelium (EpCAM) with TDP-43 to determine ontologically equivalent mammary gland ductal structures (Supplementary Fig. S2A). Whole mount analysis showed that mammary ductal branching was reduced in Tardbp− /− (K14-CreERT; Tardbp fl/fl) mice compared with wild-type (WT) (K14-CreERT; Tardbp wt/wt) mice (Fig. 3D, E). To further investigate any morphological changes, we performed histological analyses by H&E staining. Results showed that the mammary gland ducts from Tardbp− /− mice exhibited fewer cellular layers than that of WT mice (Fig. 4F), which was possibly due to MaSCs loss, as reported in previous research [1]. Immunofluorescence staining with Ki67 antibody demonstrated that proliferation was inhibited in Tardbp− /− mice compared with that in WT mice (Fig. 3G). Thus, taken together, our data suggest that TDP-43 is important for maintaining mammary morphogenesis.

Deletion of Tardbp results in reduction of mammary gland ductal branching.

Downregulation of TDP-43 inhibits mammary epithelial cell proliferation.
Loss of TDP-43 inhibits proliferation of MECs
To explore the underlying mechanism of the effects of TDP-43 on the proliferation of MECs in vivo (Fig. 3G), we confirmed the reduction effects by TDP-43 loss in vitro. MTS assay demonstrated that knockdown of TDP-43 inhibited HC11 cell proliferation (Fig. 4A). Consistent with the MTS assay, BrdU staining showed suppression of cell cycle progression, as evidenced by the decreased BrdU-positive signals in the HC11 cells (Fig. 4B). Further observation by flow cytometry indicated that the reduced TDP-43 expression led to cell cycle arrest at the G1 phase (Fig. 4C).
Downregulation of TDP-43 induces cell cycle arrest via upregulation of lincRNA-p21 and p21
To reveal how TDP-43 affects the proliferation of MECs, RNA-sequencing was performed to identify differentially expressed genes after knockdown of TDP-43 in primary MECs. In total, 1539 genes exhibited changes following loss of TDP-43 expression (Supplementary Fig. S3A). GO identified that these genes were enriched in terms related to cell growth, including mitotic nuclear division, cell cycle, and cell division (Fig. 5A). To further demonstrate the changes of cell cycle progression, we examined whether the expression of cell cyclins, key components of the core cell cycle machinery, was affected by downregulation of TDP-43. qPCR analysis demonstrated that mRNA expression of cyclin F, cyclin A2, and cyclin B2 were upregulated following downregulation of TDP-43 (Fig. 5B). It has been shown that these cyclins involved in cell cycle regulation [28,29]. Western blot showed that expression levels of p21 were upregulated (Fig. 5C). This was consistent with our earlier observation that proliferation was inhibited in Tardbp− /− mice and reduced TDP-43 expression resulted in cell cycle arrest at the G1 phase (Figs. 3G and 4C). Also, we tested the status of p53, but there was no change compared with that in control group (Supplementary Fig. S3B). Interestingly, heatmap analysis of lincRNA demonstrated that the expression level of lincRNA-p21 (also named Trp53cor1), a long noncoding RNA (lncRNA) involved in cell cycle regulation [30], was markedly upregulated (Fig. 5D) and partial lncRNA genes that previously reported with functions associated with development showed nonsignificant changes (Supplementary Fig. S3C). LincRNA-p21 upregulation was further demonstrated by qPCR analysis (Fig. 5E). These results indicate that the regulation of TDP-43 on the proliferation of MECs is likely derived from changes in the cell cycle.
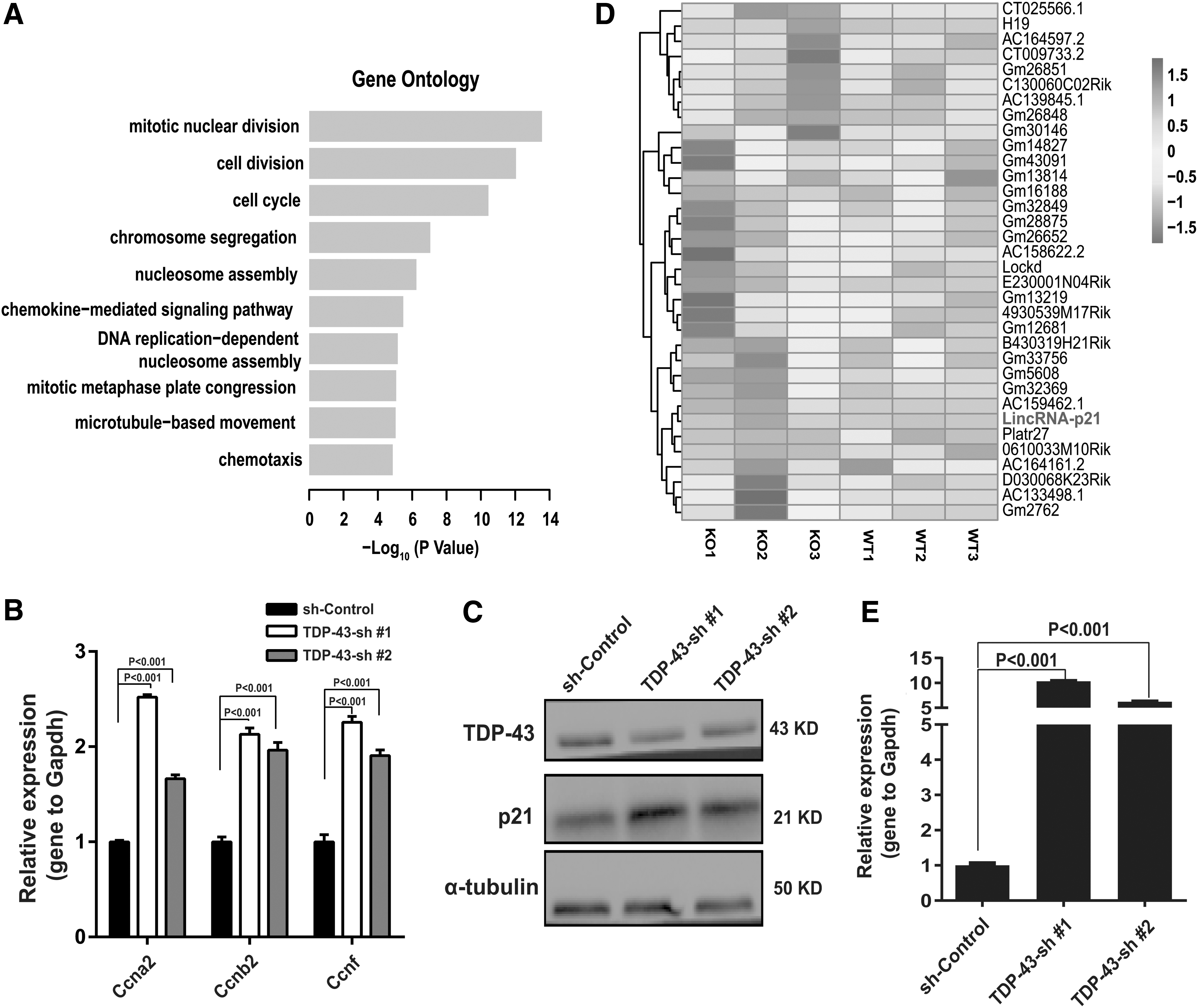
Downregulation of TDP-43 induces genes related to the cell cycle.
Discussion
Our study demonstrated that TDP-43 plays essential roles in mammary morphogenesis. TDP-43 is abundant in many tissues and evolutionarily conserved among mammals and invertebrates [31]. Based on single-cell sequencing, we identified that TDP-43 was highly expressed in mammary gland epithelial cells at the E18 stage and in some Procr+ cells, which are highly enriched in MaSCs. Moreover, we found that TDP-43 was highly expressed in basal cells enriched in MaSCs. These data indicate that TDP-43 is involved in the regulation of mammary gland morphogenesis. Previous studies have shown that ESCs with loss of TDP-43 exhibit reduced colony size [18]. Moreover, the number of surviving ESCs are also significantly reduced [18]. In accordance with the results demonstrating that TDP-43 was essential for ESC survival and proliferation, we demonstrated that knockdown of TDP-43 inhibited the repopulation of MECs based on mammosphere-formation and transplantation assays, which indicated that loss of TDP-43 in MECs reduced the number of mammospheres and generated less mammary reconstitution and ductal outgrowth. Furthermore, partial deletion of Tardbp in MECs resulted in the reduction of mammary gland ductal branching and inhibition of MEC proliferation in virgin mammary glands. These data suggest that TDP-43 plays an essential role in maintaining the repopulation of MECs and mammary morphogenesis.
Previous research on mouse ESCs has shown that the pluripotency of stem cells is linked with specific cell cycle regulation and shortened G1 phase [32]. Moreover, pluripotent stem cells show an extended G1 phase during differentiation [33,34]. Through RNA-seq results in primary MECs, we determined that numerous cell growth genes exhibited significant changes. We observed that p21 and lincRNA-p21 were upregulated after knockdown of TDP-43. It is well understood that lincRNA-p21 is involved in cell cycle regulation [35] and that p21 [36], acting as cyclin-dependent kinase inhibitors, inhibit G1 phase progression. In the current study, deletion of TDP-43 inhibited proliferation in vivo and induced cell cycle arrest at the G1 phase in HC11 cells. These results indicate that the effects of TDP-43 on MECs were possibly derived from changes in the cell cycle.
In conclusion, we found that TDP-43 participated in the functional regulation of MECs, and it is possible that this regulatory inhibition of repopulation was derived from an extended G1 phase. The detailed association between TDP-43 and the cell cycle of MECs needs further elaboration.
Footnotes
Acknowledgments
This work was equally supported by the National Key Research and Development Program of China (2016YFA0100900), Strategic Priority Research Program of the Chinese Academy of Sciences (XDB13030400), National Science Foundation of China (31371502), and Yunnan Applied Basic Research Key Projects (2018FA002, 2015HA026). We thank Dr. Christine Watts for English editing.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.