
Abstract
Human amniotic mesenchymal stem cells (HAMSCs) are promising seed cells with great advantages in promoting angiogenesis. However, the mechanisms underlying angiogenesis facilitated by HAMSCs are still unclear. Long noncoding RNA H19 is involved in many biological processes, such as enhancing angiogenesis and proliferation, invasion, and migration of cancer cells. In this study, we constructed HAMSCs of stable low-expression H19 (HAMSC-shH19) and the scramble control (HAMSC-shNC) using lentiviral vectors, and in a three-dimensional coculture with human umbilical vein endothelial cells (HUVECs) to investigate the effect of H19 knockdown in HAMSCs on angiogenesis. Our results demonstrated that H19 knockdown significantly inhibited the angiogenic function of HAMSCs at an early stage in vitro and in vivo. The results of CCK8 and transwell assays demonstrated that the conditioned medium secreted by HAMSCs reduced proliferation and migration of HUVECs after downregulating H19. The angiogenesis factors expressed and secreted by HAMSC-shH19 were decreased compared with those secreted by the control, while angiogenesis inhibitors were elevated. Furthermore, we conducted chromatin immunoprecipitation and RNA-binding protein immunoprecipitation assays and found that H19 could interact with the histone methyltransferase Enhancer of Zeste homolog 2 (EZH2) and that H19 knockdown inhibited the ability of EZH2 to recruit methyl groups to the promoter region of the angiogenesis inhibitor gene vasohibin-1 (VASH1), thus increasing VASH1 expression and secretion of HAMSCs, suppressing angiogenesis. In summary, our study identified H19 as an important regulator in HAMSCs for promoting angiogenesis, which would help to construct ideal gene-modified seed cells to enhance angiogenesis in regenerative medicine.
Introduction
It is known that angiogenesis plays a crucial role in regenerative medicine [1]. Human amniotic mesenchymal stem cells (HAMSCs) have gained wide interest as promising seed cells due to their low immunogenicity, convenient availability and, especially, their great advantages in promoting angiogenesis [2,3]. HAMSCs express and secrete significantly high levels of representative proangiogenic factors, such as vascular endothelial growth factor (VEGF), hepatocyte growth factor, fibroblast growth factor-2, and angiopoietin-1 [2]. Our previous studies have demonstrated that HAMSCs enhance the sprouting and tube formation of human umbilical vein endothelial cells (HUVECs) in vitro and in vivo [4]. To date, growing evidence supports that the trophic effects of mesenchymal stem cells (MSCs) in repair processes are attributed to the paracrine factors released by MSCs, but not their differentiation into desired cells [5]. Angiogenesis is thought to be regulated by a series of angiogenesis-related cytokines, such as the proangiogenic factor VEGF, platelet-derived growth factor (PDGF), and angiogenin (ANG), and the angiogenesis inhibitors vasohibin-1 (VASH1) and platelet factor 4 (PF4) [6]. HAMSCs promote angiogenesis partly through secreting angiogenic factors [7], but the exact mechanism underlying the proangiogenic process remains unclear.
Long noncoding RNAs (lncRNAs) are a large class of nonprotein-coding RNA with a length of more than 200 nucleotides and account for ∼80% of the transcriptome; lncRNAs are emerging as important regulators in the differentiation and function of stem cells through epigenetic, transcriptional, and posttranscriptional regulation [8]. H19, located on 11p15.5, is an imprinted gene and is highly conserved throughout evolution, indicating that is has an important biological function [9]. The H19 gene does not encode proteins but is transcribed into a 2.3-kb lncRNA H19 [10]. lncRNA H19 is highly expressed in several kinds of cancers, such as bladder cancer, colorectal cancer, and gastric cancer [11]. Cancer cells with H19 high expression possess enhanced proliferation, invasion, migration, and proangiogenic activities, promoting the development of cancer [12,13]. These results prompted us to explore whether H19 is involved in the proangiogenic effect of HAMSCs.
Enhancer of Zeste homolog 2 (EZH2), which is a histone methyltransferase, makes up polycomb-repressive complex 2 (PRC2) [14]. Within this repressive complex, EZH2 catalyzes the trimethylation of histone 3 on lysine 27 (H3K27me3) and modulates the histone methylation of target genes, leading to gene silencing [15]. Several lncRNAs have been reported to bind to PRC2, including PVT1, metastasis-associated lung adenocarcinoma transcript 1, and HOX transcript antisense RNA (HOTAIR), to recruit methyl groups to the promoter region of target genes and silence gene expression [15,16]. For example, HOTAIR is the first lncRNA to be identified to form a repressive complex with PRC2, which suppresses the expression of downstream targets through chromatin remodeling [17]. lncRNA ANCR interacts with EZH2 and inhibits Runx2 expression to regulate osteoblast differentiation [18]. Therefore, we speculated that lncRNA H19 might regulate the proangiogenic effect of HAMSCs by associating with EZH2.
To test this hypothesis, we established H19 stable low-expression HAMSCs (HAMSC-shH19) that encoded the lentivirus gene vector and then explored the influence of H19 on the proangiogenic capacity of HAMSCs in vitro and in vivo. We also investigated the mechanism underlying the effect of H19 on angiogenesis at the molecular level.
Materials and Methods
Isolation and culture of primary cells
Primary HAMSCs were isolated from amnions with 2.4 U/mL dispase II (Roche, Germany) and 1 mg/mL collagenase D (Roche) digestion and were then cultured in α-minimum essential medium (α-MEM; Gibco, USA) supplemented with 5% human platelet lysis (Helios, German), 2 U/mL heparin (Sigma, USA), and 1% antibiotics. Primary HUVECs were isolated from human umbilical cords through digestion with 0.1% (w/v) collagenase type I (Roche) and then cultured in endothelial cell medium (ScienCell, USA). Both cell lines were identified by flow cytometry as previously described and cultured in a humidified incubator at 37°C with 5% carbon dioxide. The details of the isolation and identification were also described in our previous study [4]. The experiments have been approved by the Ethics Committee of Nanjing Medical University (No. PJ2013-037-001). Informed consent was obtained from all the participants enrolled in the study.
Lentivirus and small interfering RNA transfection
Lentiviral vectors harboring RNAi sequences that target the H19 gene (shH19-1 and shH19-2) and scramble control (shNC) were purchased from GenePharma (Shanghai, China), and the knockdown efficiency in HAMSCs was verified. Primary HAMSCs at passages 2 to 3 and 50%–60% confluence were treated with polybrene (5 μg/mL) and lentiviral vectors containing green fluorescent protein (GFP) for 10 h for lentivirus transfection. The infected cells were screened for GFP expression by inverted fluorescence microscopy (Leica, Germany). The cells with an infection rate >95% were used as the stably expressing model. The sequence of small interfering RNAs (siRNAs) targeting EZH2 was AUCAGCUCGUCUGAACCUCUU [19], and the siRNA universal negative control (Invitrogen, USA) was used as a negative control. HAMSCs at 60%–70% confluence were transfected with control siRNA or with siRNA targeting EZH2 using Lipofectamine RNAiMAX (Invitrogen).
In vitro and in vivo three-dimensional fibrin vasculogenic assays
Primary HUVECs were labeled with red fluorescent protein (RFP) and cocultured with HAMSCs (shNC and shH19 groups, respectively) under three-dimensional (3D) fibrin matrices. The procedure of 3D coculture was in accordance with the previously described method [4]. RFP-HUVECs and HAMSCs (HAMSC-shNC or HAMSC-shH19) were mixed in a 2.5 mg/mL human fibrinogen (Sigma) solution at a ratio of 1:1. Five microliters of thrombin (25 U/mL; Sigma) was added to every 100 μL fibrinogen–cell suspension to catalyze solidification. Fluorescent images were captured using inverted fluorescence microscopy at 0, 24, 48, and 72 h, and the sprouting lengths of HUVECs were measured by Image-Pro Plus Software (version 6.0). Three random sights were selected for each group.
In vivo, we used male 8-week-old BALB/c-nu mice to study angiogenesis with subcutaneous injection in all experiments according to previous studies [4,20]. HAMSC-shNC and HAMSC-shH19 were respectively mixed with HUVECs at a ratio of 1:1 in 300 μL of a fibrinogen solution at a final concentration of 10 million cells per mL. Then, cell mixtures were immediately subcutaneously injected into mice after adding 15 μL thrombin to catalyze solidification. After 3D coculturing for 7 days, the implants were removed and fixed in 10% buffered formalin for histological and immunohistochemical analyses. All animal protocols were approved by the Institutional Animal Care and Use Committee of Nanjing Medical University (No. IACUC-1701014).
Histology and immunohistochemistry
Samples were embedded in paraffin and sliced into 5 μm-thick sections for histopathological analyses. Histological and immunohistochemical staining were performed as previously described [21]. A rabbit monoclonal anti-CD31 primary antibody (ab134168; Abcam, UK) was used to mark the vessel-like structures (VLS) newly formed by HUVECs.
Assessment of cellular proliferation and migration
HAMSC-shNC and HAMSC-shH19 were cultured with α-MEM medium for 48 h, and supernatants were collected as the conditioned medium (shNC and shH19 groups). CCK8 and transwell assays were conducted to assess the proliferation and migration of HUVECs cultured in conditioned medium. Briefly, HUVECs were seeded into 96-well culture plates at a density of 2,000 cells per well. After 24, 48, and 72 h of culturing in conditioned medium, HUVEC-targeted wells were treated with a 10% CCK8 (Sigma) solution and incubated for 1 h. Then, the absorbance values were measured at 450 nm by a SpectraMax M2/M2e (Molecular Devices, USA). All experiments were performed with six replicates (n = 6). Migration assays were performed using a transwell chamber (Corning, USA). HUVECs were seeded into a transwell chamber at a density of 5,000 cells per chamber. We placed the chambers into 24-well plates and added 200 μL conditioned medium per well. The number of cells migrating across chambers was counted after culturing for 24 and 48 h.
Enzyme-linked immunosorbent assays
The concentrations of ANG and VASH1 in HAMSC-shH19- and HAMSC-shNC-conditioned media were detected using Enzyme-Linked Immunosorbent Assay (ELISA) Kits (BD Biosciences, USA) according to the manufacturer's instructions. All experiments were performed on six samples per group (n = 6).
RNA extraction and quantitative reverse transcription–PCR
Total cellular RNA was extracted by TRIzol (Invitrogen) and then reverse transcribed into complementary DNA (cDNA). Quantitative reverse transcription–PCR (qRT-PCR) was performed to detect the relative gene expression levels with an ABI 7300 real-time PCR System (Applied Biosystems, USA) with the following settings: 95°C for 10 min followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal control for messenger RNAs (mRNAs) and lncRNAs. The related primer sequences for H19, ANG, PDGF, VASH1, PF4, and GAPDH are listed in Supplementary Table S1. The data were analyzed using the 2−ΔΔCt relative expression method as described previously [22].
Western blot analysis
Cells were collected and lysed with RIPA (Beyotime, China). The converted proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, electrotransferred to polyvinylidene fluoride (PVDF) membranes, and incubated with primary antibodies against ANG (ab10600; Abcam), PDGF (ab23914; Abcam), VASH1 (ab199732; Abcam), PF4 (ab129183; Abcam), EZH2 (ab186006; Abcam), and GAPDH (ab181602; Abcam) overnight at 4°C. Protein bands were detected by chemiluminescence using the ECL system (Thermo Scientific, USA).
Chromatin immunoprecipitation assays
Chromatin immunoprecipitation (ChIP) assays were performed using the EZ-Magna CHIP Kit (Millipore, USA) according to the manufacturer's protocols. Briefly, HAMSC-shNC and HAMSC-shH19 were treated with formaldehyde to crosslink protein and DNA, respectively, and were then added to the lysis buffer. Then, lysates were sonicated to shear the chromatin into fragments of 200–300 bp. We removed the first 1% of the supernatant as the “Input,” and every 10% of the supernatant was immunoprecipitated with antibodies as follows: EZH2 (Millipore), H3K27me3 (Millipore), and the rabbit immunoglobulin G (IgG) (supplied in the CHIP kit) as the negative control. Finally, free DNA was eluted from protein/DNA complexes by proteinase K and then purified. Immunoprecipitated DNA was quantified by qPCR, and the data were calculated by 2[Input Ct – Target Ct] × 0.1 relative to the input. The ChIP primer sequences are listed in Supplementary Table S2.
RNA-binding protein immunoprecipitation assays
RNA-binding protein immunoprecipitation (RIP) assays were performed using the EZ-Magna RIP Kit (Millipore) according to the manufacturer's protocols. Briefly, HAMSC-shNC and HAMSC-shH19 were scraped off and treated with RIP lysis buffer. We removed 10 μL of the supernatant from the RIP lysate as the “Input,” and every 100 μL was incubated with the magnetic beads–antibody complex buffer at 4°C overnight. The antibodies used for the RIP assays were EZH2 (Millipore) and rabbit IgG (supplied in the RIP kit), which served as the negative control. Then, each immunoprecipitate was exposed to proteinase K buffer and RIP wash buffer for RNA purification. Purified RNA was reverse transcribed to cDNA and analyzed by qRT-PCR with specific primers for H19.
Results
H19 knockdown inhibited the proangiogenic capacity of HAMSCs in vitro and in vivo
Both the shH19-1 and shH19-2 lentiviral vectors decreased the expression level of H19 in HAMSCs. Given the higher interference efficiency of lenti-shH19-1 (Supplementary Fig. S1), HAMSCs transfected with the shH19-1 lentiviral vector were used as the stably expressing model in subsequent experiments. To investigate whether H19 was involved in the process of promoting angiogenesis in HAMSCs, HAMSC-shH19 and HAMSC-shNC were 3D cocultured with HUVECs in vitro and in vivo, respectively.
In vitro, we marked HUVECs with RFP and cocultured them with HAMSC-shH19 and HAMSC-shNC in 3D fibrinogen gels. We focused on HUVECs to evaluate the function of lncRNA H19 in angiogenesis. The average sprouting lengths of HUVECs were measured after coculturing for 24, 48, and 72 h. As shown in Fig. 1, HUVEC sprouting of the HAMSC-shH19 cocultured group was significantly inhibited compared with that of the control group at every time point (P < 0.01).

H19 knockdown inhibited the proangiogenic capacity of HAMSCs in vitro.
In vivo, the proangiogenic capacity of HAMSC-shH19 was evaluated using a subcutaneous model in nude mice (Fig. 2). HAMSC-shH19 and HAMSC-shNC were mixed with HUVECs in fibrinogen gels and immediately subcutaneously injected into nude mice. After in vivo culturing for 7 days, the implants were removed for histological analysis. Histomorphometric analysis of Hematoxylin and Eosin (H&E)-stained sections showed that fewer newly formed VLS were observed in the HAMSC-shH19-cocultured implants than in the control groups, especially in the inner layer. To verify the results of the H&E staining, immunohistochemical analysis of human CD31 was used to mark the newly formed VLS originating from the implanted HUVECs. The results further demonstrated that the density of VLS was decreased in the HAMSC-shH19-cocultured group compared with the control group. The percentage of the CD31-positive area showed the same trend between the two cocultured groups, suggesting that the angiogenic capacity of HUVECs in the HAMSC-shH19-cocultured group was reduced. Taken together, these data suggested that downregulation of lncRNA H19 contributed to the decreased proangiogenic capacity of HAMSCs in the early stage.
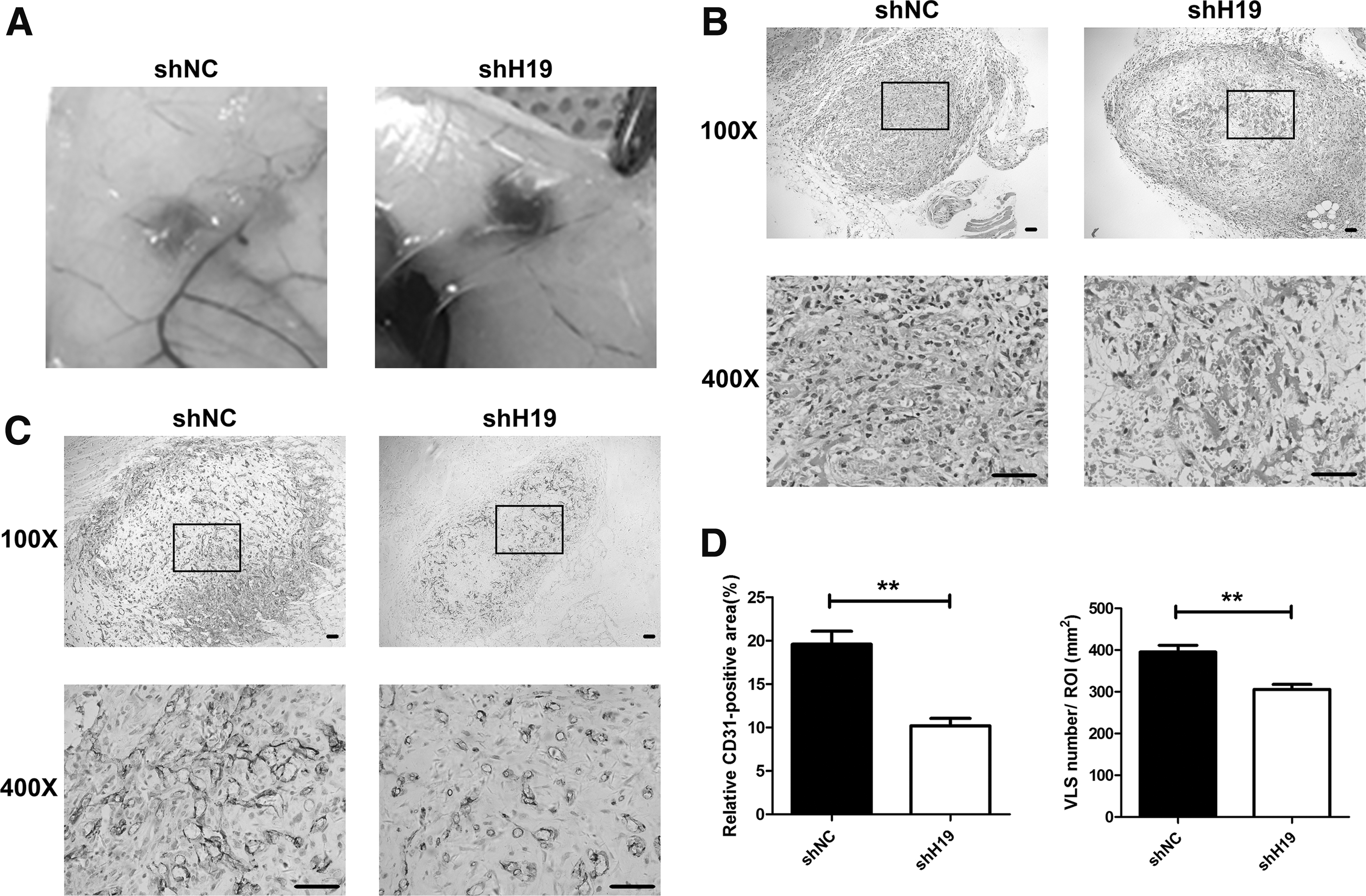
H19 knockdown inhibited the proangiogenic capacity of HAMSCs in vivo.
Conditioned medium secreted by HAMSCs reduced proliferation and migration of HUVECs after downregulating H19
Our previous study found that the conditioned medium secreted by HAMSCs contained growth factor-facilitated proliferation and angiogenesis of HUVECs. Thus, we hypothesized that H19 knockdown could affect the paracrine action of HAMSCs and then influence the proliferation and migration of HUVECs. To verify this hypothesis, we collected the conditioned media from HAMSC-shH19 and HAMSC-shNC to use for culturing HUVECs. CCK8 assays were conducted after culturing HUVECs in conditioned medium for 24, 48, and 72 h. The results showed that the conditioned medium secreted by HAMSC-shH19 significantly repressed the proliferation of HUVECs compared with that of the control group at every time point (P < 0.01; Fig. 3A). We also conducted transwell assays to assess the migration of HUVECs after culturing in conditioned medium for 24 and 48 h. The results indicated that the migration of HUVECs cultured in HAMSC-shH19-conditioned medium was reduced in comparison with that of the control group (P < 0.01; Fig. 3B).
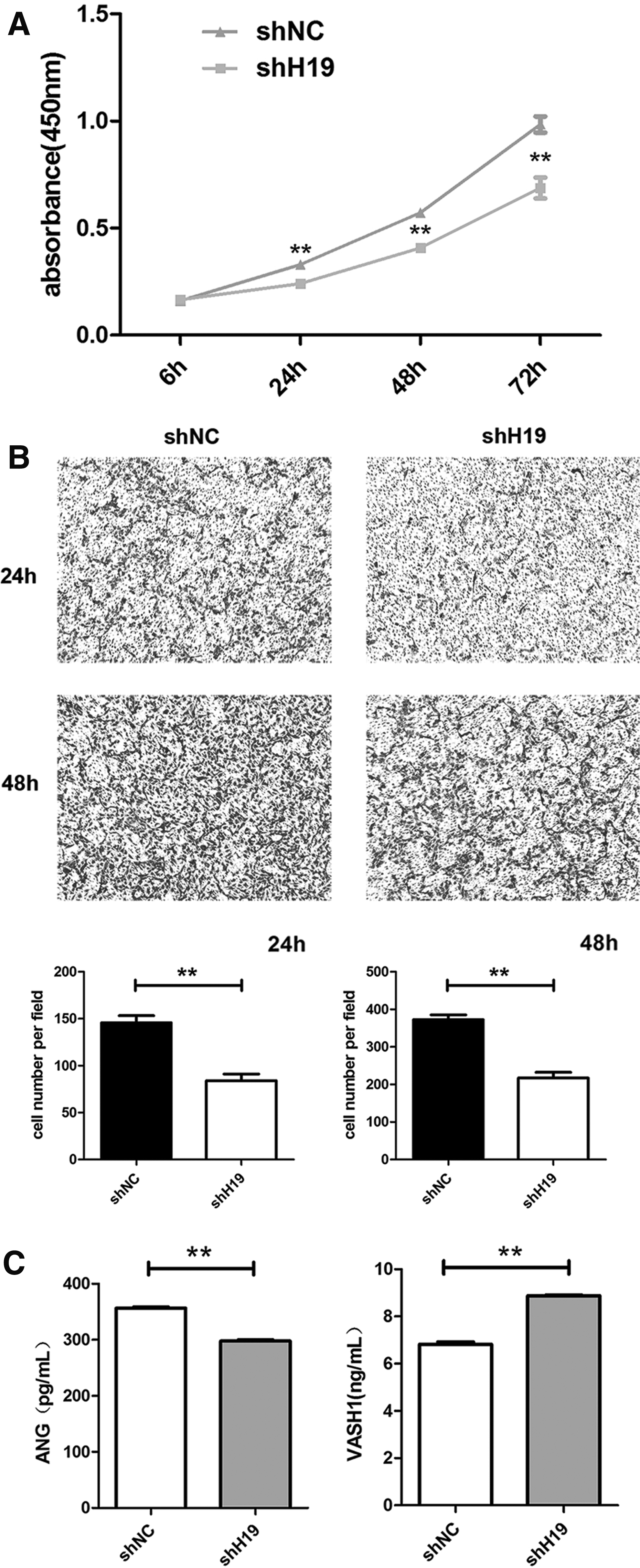
Conditioned media secreted by HAMSC-shNC and HAMSC-shH19 influenced the proliferation and migration of HUVECs.
Angiogenesis factors expressed and secreted by HAMSC-shH19 were decreased compared with the control, while angiogenesis inhibitors were elevated
We compared the concentration of angiogenesis-related factors in HAMSC-shH19 and HAMSC-shNC-conditioned media by protein microarray analysis. Compared with the HAMSC-shNC-conditioned medium, the concentrations of angiogenesis factors, such as ANG, PDGF, and matrix metallopeptidase 9 (MMP9), in HAMSC-shH19 were decreased and the concentrations of angiogenesis inhibitors, such as PF4 and VASH1, were increased. As validated by ELISA, similar trends were apparent for the concentrations of VASH1 and ANG (Fig. 3C). Furthermore, we explored the protein and mRNA expression changes in angiogenesis-related genes between HAMSC-shH19 and HAMSC-shNC using RT-PCR and western blotting. Notably, the expression of the angiogenesis factors, ANG and PDGF, was markedly downregulated and that of the angiogenesis inhibitors, VASH1 and PF4, was elevated after H19 knockdown in HAMSCs (Fig. 4).

Expression of angiogenesis-related genes between HAMSC-shNC and HAMSC-shH19.
lncRNA H19 could bind to methyltransferase EZH2 and enhance the recruitment of methyl groups to the promoter region of the angiogenesis inhibitor gene VASH1, thus decreasing VASH1 expression in HAMSCs
To provide insight into the molecular mechanisms by which H19 contributes to the proangiogenic effect in HAMSCs, we continued to explore how H19 regulates angiogenesis-related genes. Several studies have reported that lncRNAs, including H19, could interact with the histone methyltransferase EZH2 to enhance the epigenetic regulation of EZH2 and silence downstream genes [23]. EZH2 is a histone methyltransferase that catalyzes the trimethylation of histone 3 on lysine 27 (H3K27) and modulates the histone methylation of target genes. To further explore the potential target genes, we investigated the epigenetic regulation modification factors of these angiogenesis-related genes by using UCSC (

lncRNA H19 binds to EZH2 and enhances H3K27me3 in the promoter region of the angiogenesis inhibitor gene VASH1 in HAMSCs.
To confirm the interaction between H19 and EZH2 in HAMSCs, we performed RIP assays to confirm their direct binding using an EZH2-specific antibody to immunoprecipitate RNA followed by a primer to detect H19 RNA. A significant enrichment of H19 RNA was observed with the EZH2 antibody, but no enrichment with the negative control IgG antibody was found (Fig. 5E).
Taken together, these results demonstrated that lncRNA H19 could bind to the methyltransferase EZH2 and that H19 knockdown inhibited EZH2 to recruit methyl groups to the promoter region of the angiogenesis inhibitor gene VASH1, thus increasing VASH1 expression and the secretion of HAMSCs, suppressing angiogenesis (Fig. 6).
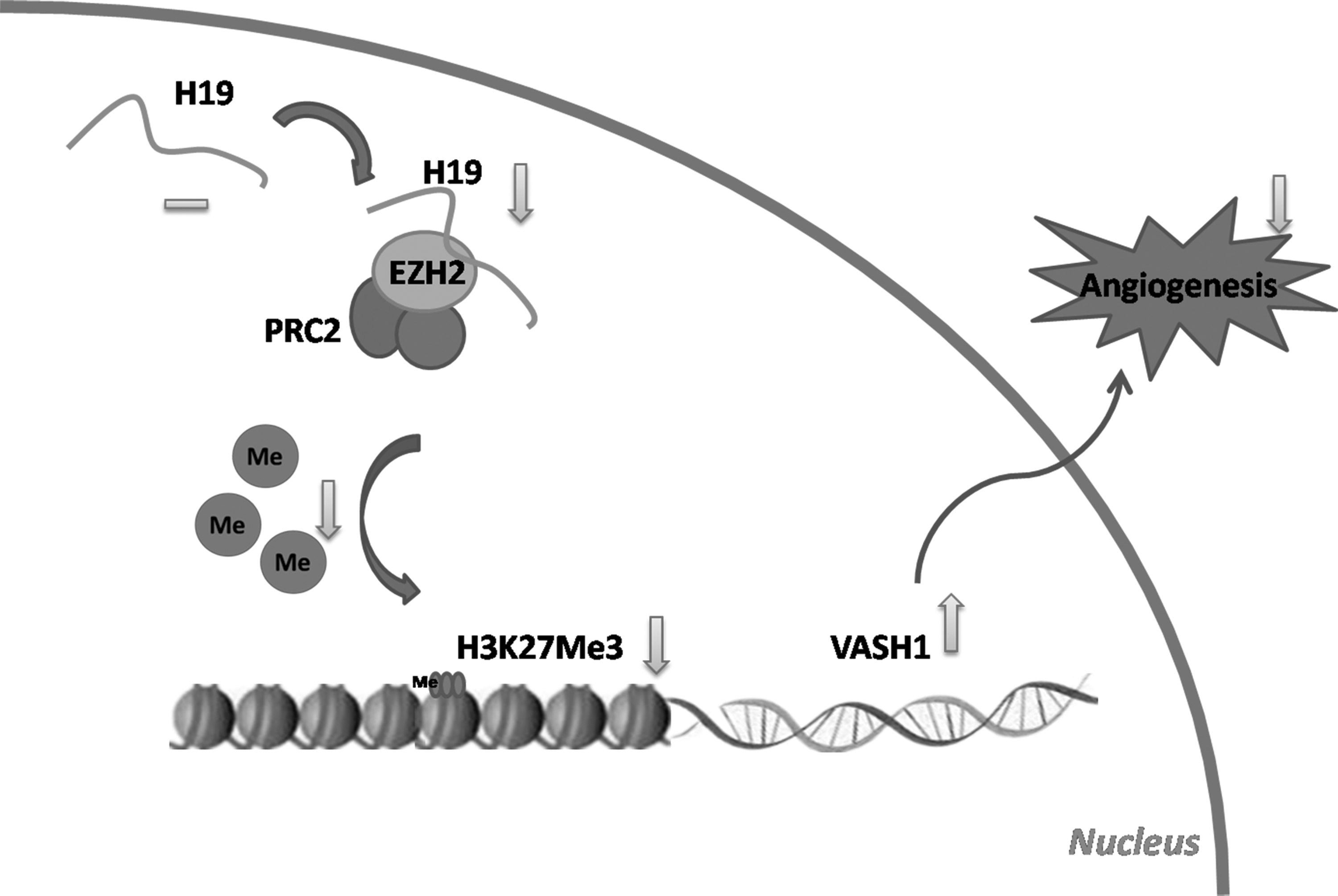
Proposed model by which H19 knockdown in HAMSC suppresses angiogenesis. H19 interacts with histone methyltransferase EZH2. H19 knockdown inhibits EZH2 recruitment of methyl groups to the promoter region of the angiogenesis inhibitor gene VASH1, thus increasing VASH1 expression and the secretion of HAMSCs, suppressing angiogenesis.
Discussion
Angiogenesis plays a crucial role in regenerative medicine. It is known that HAMSCs promote angiogenesis. Thus, exploration of the mechanisms underlying angiogenesis facilitated by HAMSCs is necessary and would help us to construct ideal gene-modified seed cells to enhance angiogenesis in regenerative medicine. The functions of stem cells could be regulated by epigenetic and transcriptional control [24]. Angiogenesis is the process by which vascular endothelial cells sprout, proliferate, migrate, and form tubules under the stimulation of a series of angiogenesis and antiapoptosis-related factors [25]. In this study, we used HUVECs as the experimental cells to study angiogenesis and identified lncRNA H19 as an important regulator of HAMSCs for promoting angiogenesis in vitro and in vivo.
Transplanted MSCs practically disappear 1 week after implantation [26], and their angiogenic function mainly relies on the secretion of angiogenic growth factors [27]. A previous study has demonstrated that HAMSCs express and secrete significantly high levels of representative angiogenic factors [2], whereas H19 affects the secretion of these factors, promoting the proliferation and migration of HUVECs to form prevascular structures at the early stage, which may further facilitate angiogenesis. Among them, ANG can activate vascular endothelial cells and promote angiogenesis by activating a series of signaling pathways that are involved in cell migration, invasion, proliferation, and tube formation [28]. PDGF can stimulate the proliferation and migration of specific cells and promote capillary formation. PDGF has been approved for clinical use and has led to great progress in the treatment of chronic ulcers and periodontal tissue regeneration [29]. PF4, also known as CXCL4, is a chemokine that inhibits angiogenesis [30]. VASH1 is an angiogenic inhibitor that specifically inhibits the proliferation, migration, and tube formation of endothelial cells, but not other cells [31]. In in vitro experiments, VASH1 did not affect the proliferation of tumor cells, but suppressed tumor growth and angiogenesis [32]. We showed that knockdown of H19 increased the expression of angiogenic inhibitors, including VASH1 in HAMSCs, prompting us to further explore the biological mechanisms by which H19 regulates the angiogenesis of HAMSCs.
lncRNAs have emerged as important regulators in gene regulation and cellular development [33]. Accumulating studies report that lncRNAs could serve as molecular scaffolds and epigenetic modifiers to regulate target gene expression [34] and are widely involved in various cellular biological processes, including regulation of cellular apoptosis and invasion and the remodeling of stem cell pluripotency [35]. lncRNA H19 is highly expressed in the early stage of stem cell differentiation. H19 can be found in the nucleus and cytoplasm [23]. In the cytoplasm, H19 participates in cellular biological processes by functioning as competing endogenous RNAs or regulating mRNA stability and splicing [36,37]. For instance, Huang et al. [38] demonstrated that H19 could promote osteoblast differentiation by deriving miR-675 and activating the TGF-b1/Smad3/HDAC pathway. In the nucleus, H19 can guide and recruit DNA, histone protein modification enzymes, or transcription factors to specific DNA sequences in the promoters of target genes [23]. In this study, we mainly focused on the function of intranuclear H19 and demonstrated that H19 could interact with the histone methyltransferase EZH2 and guide H19–PRC2 complexes to bind to the promoter of VASH, enhancing the epigenetic regulation of the downstream gene VASH1 through catalyzing the trimethylation of H3K27 in the promoter region, thus decreasing the expression and secretion of the angiogenesis inhibitor gene VASH1 in HAMSCs and promoting angiogenesis. Lu et al. [39] have found that EZH2 can bind to the VASH1 promoter and mediate VASH1 methylation and silencing, regulating tumor angiogenesis, in agreement with our results and the bioinformatics predictions. Our research not only verified EZH2-VASH1 promoter binding but also demonstrated that H19 could affect the binding and further regulate angiogenesis.
There were several limitations to this study. First, we failed to construct HAMSCs of stably overexpressed H19, so the effect of H19 in HAMSCs on angiogenesis was not fully investigated. Thus, we further explored the biological mechanisms by which H19 regulates the angiogenesis of HAMSCs to clarify the function of H19 at the molecular level. Second, in this study, we mainly focused on the function of intranuclear H19, while the effect of H19 in the cytoplasm was not assessed.
Our study supports the hypothesis that lncRNA H19 plays a critical role in HAMSCs-mediated angiogenesis both in vitro and in vivo and provides important insights into the molecular mechanisms, which will be helpful in developing gene-modified MSC-based therapy for angiogenesis in regenerative medicine.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (grant no. 81670966).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.