
Abstract
The aim of this study was to compare the in vitro osteogenic differentiation potential of within-subject mesenchymal stem cells (MSCs) derived from the dental pulp of permanent teeth (dental pulp stem cells-DPSCs), the dental pulp of deciduous teeth (stem cells from human exfoliated deciduous teeth-SHEDs), and the periodontal ligament of permanent teeth (periodontal ligament stem cells-PDLSCs). A single subject was identified that required concurrent removal of both deciduous and permanent teeth for orthodontic purposes. Primary, mixed population cells from dental pulp, deciduous dental pulp, and periodontal ligament were obtained by the tissue outgrowth method. Subsequently, isolation of STRO-1 +ve cells from their respective primary cell cultures was achieved by immunomagnetic separation. Cells were induced with an osteogenic cocktail of 5 mM β-glycerophosphate, 100 nM dexamethasone, and 50 mg/mL ascorbic acid for up to 21 days. Osteogenic responses were assessed functionally by an alkaline phosphatase (ALP) activity assay and an alizarin red staining assay. Expression of the early osteogenic associated genes, alkaline phosphatase gene (ALPL), runt-related transcription factor 2 (RUNX2), collagen type I alpha 1 (COL1A1), and secreted phosphoprotein 1 (SPP1), was compared by qPCR at days 1, 4, and 7 of differentiation. Functional analysis revealed that there were significant differences in intracellular ALP activity on days 4, 7, 10, and 14 with PDLSCs > SHEDs > DPSCs. Quantification of alizarin red staining showed significantly more mineralization for PDLSCs by day 21. Gene expression analysis showed significant early upregulations of the osteogenic markers ALPL and COL1A1 for PDLSCs over DPSCs and SHEDs. SHEDs showed significantly higher upregulation of ALPL over DPSCs. In conclusion, PDLSCs showed a significantly higher osteogenic differentiation potential than both DPSCs and SHEDs evidenced by functional studies and gene expression. This may be of significance for the use of dentally derived MSCs in bone tissue engineering applications.
Introduction
Mesenchymal stem cells (MSCs) are recognized as offering much promise in the areas of tissue engineering and regenerative medicine due to their ability to differentiate into specific tissue forming cell types. The current practice of tissue engineering involves the combination of cells and/or proteins with biomaterials to generate a new tissue [1]. Factors affecting the success of tissue engineering include the following: the type of stem cell utilized; the strategies used to isolate and expand the specific cells; and the choice of the biomaterial used as a scaffold [2]. MSCs have great potential in bone-tissue engineering applications due to their ability to undergo osteoblastic differentiation under the appropriate conditions. Bone marrow-derived mesenchymal stem cells (BMMSCs) are currently considered the gold standard MSC for bone tissue engineering [3]. Intense research effort has been directed toward examining BMMSCs for applications in bone-tissue engineering [4,5].
MSC populations have also been isolated from several oral tissues such as dental pulp, periodontal ligament (PDL), apical papilla, dental follicle, tooth germ, gingivae, and periosteum [6 –10]. Owing to their relative ease of access and less invasive harvesting techniques in comparison to BMMSCs, dental MSCs represent an attractive alternative MSC source for bone-tissue engineering applications [11,12]. Studies have also found other advantageous properties such as the ability to produce higher colony forming units, a higher cell proliferation rate, and a longer survival time in comparison to BMMSCs [13,14]. A number of studies have demonstrated the osteogenic potential of the various dental MSCs utilizing both in vitro and in vivo models [15]. While an osteogenic potential in general may have been demonstrated, it should be realized that there are significant phenotypic differences between the different types of dental MSCs reflecting their distinct functional properties [16,17]. This heterogeneity may potentially alter the efficacy of the individual lineage's osteogenic potential. To this end, several studies have been performed attempting to compare the osteogenic potential of various selected dental MSCs.
A study investigated the osteogenic potential of periodontal ligament stem cells (PDLSCs) compared to that of stem cells from the pulp of human exfoliated deciduous teeth (SHEDs) [18]. Results showed that both cell types were capable of undergoing osteogenic induction; however, there was a significantly higher cell proliferation of PDLSC over SHEDs, which resulted in greater calcium deposition after 3 weeks of culture. The authors suggested that PDLSCs were a superior stem cell source for osteogenic purposes. Interpretation may be limited due to the use of cells from different donors; results may therefore reflect interindividual variations. The osteogenic differentiation potential of human dental pulp stem cells (DPSCs) has previously been compared with the osteogenic differentiation potential of stem cells from the apical papilla (SCAPs) [19]. Both DPSC and SCAP cell populations were derived from impacted third molars within the same subject. Results demonstrated that although both DPSCs and SCAP were able to differentiate into osteo/odontoblast-like cells, SCAPs had a significantly higher mineralization potential.
In a study comparing DPSCs with SHEDs [20], cells were collected and harvested from different donors. SHEDs displayed a higher osteogenic potential over DPSCs evidenced by significantly higher calcium deposition and a significantly higher expression of the osteogenic related genes ALPL (alkaline phosphatase gene), RUNX2 (runt-related transcription factor 2), and COL1A1 (collagen, type I, alpha 1). The in vivo comparisons showed that SHED transplants produced bone-like structures, whereas DPSC transplants formed connective tissue with comparatively smaller amounts of mineralized tissue. A study comparing the osteogenic differentiation potential between DPSCs and PDLSCs has previously been carried out [21]. To eliminate intersubject differences, they harvested and isolated DPSCs and PDLSCs from the same tooth for comparison (one subject). Results overall were inconclusive; PDLSCs had significantly higher COL1A1 but lower OCN (Osteocalcin) gene expression. Mineralization staining assays suggested greater mineralization for DPSC cultures, although no quantification was carried out.
In summary, there is a lack of evidence to recommend a specific dental MSC lineage for bone-tissue engineering purposes. A thorough one-to-one comparison of dental MSCs from a single donor for key osteogenic properties has not been evaluated in detail. It is important to understand whether MSCs derived from close tissues behave differently and have a different osteogenic potential. Possible differences may have implications for dental MSC bone tissue engineering applications. The aim of this study, therefore, was to compare the in vitro osteogenic differentiation potential of MSCs derived from the dental pulp of a permanent tooth (DPSCs), the dental pulp of deciduous teeth (SHEDs), and the periodontal ligament of a permanent tooth (PDLSCs).
Materials and Methods
Study population
Ethical approval was granted by the Office for Research Ethics Northern Ireland, for the collection of teeth from the School of Dentistry, Belfast Health and Social Care Trust (Ethical approval number 08/NIR03/15). Following written consent, subjects were identified that required concurrent removal of both deciduous and permanent teeth for orthodontic purposes from a similar region of the oral cavity. Teeth were caries and periodontal disease free. To rule out potential differences which may be attributable to within-subject variation, experiments were based on explants from a single donor where dental pulp from a deciduous tooth, dental pulp from a permanent tooth, and PDL from a permanent tooth were all successfully explanted and expanded. In this case, a systemically healthy, 16-year-old Caucasian male required orthodontic extraction of an upper left first permanent premolar and the adjacent upper left second deciduous molar teeth due to orthodontic crowding.
Explant, culture, and isolation of dental MSC lineages
Primary, mixed population cells from dental pulp, deciduous dental pulp, and PDL were obtained by the tissue outgrowth method. Briefly, extracted teeth were rinsed with sterile phosphate-buffered saline (PBS, 0.01 M, pH 7.4; Invitrogen, United Kingdom). The PDL tissue from the middle one-third of the permanent premolar root was then scraped off with a sterile scalpel [22]. This PDL tissue was then cut into pieces (∼1 × 1 × 1 mm) and placed into a 25 cm2 culture flask (Falcon). For pulp tissue (permanent and deciduous teeth), the tooth was split open with a bench fitted vice with a cutting edge [23]. The pulp tissue was then carefully removed with a size 40 K Flexofile (Dentsply, Maillefer, Switzerland). This pulp tissue was then similarly minced into tiny pieces (∼1 × 1 × 1 mm) and placed into a T25 culture flask. Cell explants were grown and expanded under identical culture conditions. The culture media consisted of alpha-MEM supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin (all Invitrogen), 1%
Isolation of STRO-1 +ve stem cells from their respective primary cell cultures was achieved by immunomagnetic separation [12]. Briefly, magnetic bead-linked rat anti-mouse IgM (Dynal, Oslo, Norway) beads were incubated with mouse IgM anti-human STRO-1 (Invitrogen). These were then incubated with the different cell types in an isolation buffer for 20 min at 4°C. A magnetic cell sorting rack (DynaMag, Invitrogen) was used to discard cells not magnetically retained. Thus three, STRO-1 +ve, stem cell populations were derived (DPSCs, SHEDs, and PDLSCs).
Cells were cultured in a “control media” consisting of 10% fetal bovine serum, 1% penicillin/streptomycin, 1%
Alkaline phosphatase activity
Each of the three cell types (DPSC, SHED, and PDLSCs) was seeded in triplicate in 12-well plates at a density of 4 × 104 cells/mL and grown to ∼70% confluence. Cells were then induced with the osteogenic media (time zero). Osteogenic responses were assessed functionally by an intracellular alkaline phosphatase (ALP) activity assay on days 1, 4, 7, 10, and 14. At the designated time point, cells were rinsed with PBS twice and then lysed by adding 150 μL of lysis buffer (0.2% Triton X-100). This was followed by a 15-min incubation period at 37°C. Wells were then scraped and the lysate transferred to 1.5 mL tubes, before being vortexed and centrifuged at 10,000 rpm at 4°C for 10 min. The supernatant of each sample was collected for ALP quantification using a Diethanolamine Detection Kit (Sigma-Aldrich) utilizing p-nitrophenyl phosphate (PNPP) as the substrate. Values were normalized to total protein content, which was measured using a Bicinchoninic Acid (BCA) Protein Assay Kit (Pierce, Rockford, IL) and expressed as ηmol PNPP/min/mg protein.
Alizarin red staining
Each of the three cell types was seeded in triplicate in six-well plates at a density of 4 × 104 cells/mL and grown to ∼70% confluence. Alizarin red (AR) staining was carried out at days 7, 14, and 21. For each time point triplicate wells were designated as either “Control media” or “Osteogenic media”. At the corresponding time point, cells were rinsed with PBS (Dulbecco's phosphate-buffered saline, without Ca2+/Mg2+; PromoCell, United Kingdom) twice and subsequently fixed in 70% ice-cold ethanol for 30 min. After three washes with distilled water (diH2O), the cells were stained for 30 min with 2% alizarin red, pH 4.2 (Sigma-Aldrich). Cells were then washed five times with diH2O. Full well images and images at × 10 magnification were captured with an inverted microscope with integrated camera (Leica, United Kingdom).
Quantification of the total mineralized tissue produced per well was performed by extracting the AR from the stained sites by adding 2 mL of cetylpyridinium chloride (CPC; Sigma-Aldrich) buffer (10%, w/v) in 10 mM Na2HPO4 (pH 7) for 1 h at 37°C. Subsequently, 100 mL aliquots were transferred to a 96-well plate, and the OD570nm of the solution was measured using a microplate reader (Tecan GENios, MTX Lab Systems). Values were expressed as fold change in optical density (OD570nm) of osteogenic wells over control wells for each cell type at that time point.
Osteogenic gene expression
Each of the three cell types was seeded in triplicate in 12-well plates at a density of 4 × 104/mL and grown to 70% confluence. Cells were then induced with the osteogenic media (time zero). At days 1, 4, and 7 RNA extraction and cDNA synthesis were carried out. Briefly, wells were rinsed twice with cold PBS and RNA extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. RNA yield and purity was assessed on a Take3 plate (BioTek). Following this, cDNA was synthesized using a SuperScript VILO cDNA Synthesis Kit (Invitrogen) and frozen (−80°C). Real-time qPCR for alkaline phosphatase gene (ALPL), collagen type I alpha I (COL1A1) gene, runt-related transcription factor 2 (RUNX2) gene, and secreted phosphoprotein 1 (SPP1) gene was carried out on a Stratagene PCR machine (Agilent Technologies). Normalization of results was achieved using the housekeeping genes Beta-2 microglobulin (B2M) and glucuronidase beta (GUSB). Primer sequences are listed in Table 1. Sequence amplification was performed under the following conditions: denaturation at 55°C for 2 min and 95°C for 10 min. This was then followed by 45 cycles of 95°C for 30 s and 50°C for 1 min. The relative gene expression was calculated by the 2−ΔΔCT method. Results are reported as fold change in gene expression relative to the day 1 DPSC.
Quantitative Polymerase Chain Reaction Primer Information
Statistical analyses
ALP activity data and qPCR data with multiple groups at different time points were analyzed by a two-way Analysis of Variance (ANOVA) test followed by Tukey's post hoc test for multiple comparisons. The AR staining assay was analyzed using one-way ANOVA followed by Tukey's post hoc test for multiple comparisons for each separate time point. Experiments were performed in triplicate and repeated thrice. The level of statistical significance was set at P < 0.05. The statistical analyses were performed with IBM SPSS Statistics for Windows (IBM Corp. Released 2012. Version 21.0. Armonk, NY). Graphs were constructed using Prism version 7.00 for Windows (GraphPad Software, La Jolla, CA,
Results
Three different dental soft tissues (dental pulp from a permanent tooth, dental pulp from a deciduous tooth, and PDL from a permanent tooth) from a single donor were successfully explanted by the tissue outgrowth method. Isolation of STRO-1 +ve cells by immunomagnetic separation was then concurrently undertaken on cultures, to produce three distinct populations: DPSCs, SHEDs, and PDLSCs.
To screen for the osteogenic potential of the three cell groups, ALP activity was measured at 1, 4, 7, and 14 days (Fig. 1). At day 1 there was minimal ALP activity in all three cultures with no significant differences. By Day 4, there was a significant increase in ALP activity for PDLSCs, compared to both DPSCs and SHEDs. By day 7, PDLSCs continued to have significantly greater ALP activity than both types of pulp stem cells; however, SHED ALP activity was now significantly greater than DPSC. As time progressed ALP activity for all three cell types increased. A similar pattern of significantly greater ALP activity for PDLSC > SHED > DPSC continued at both day 10 and day 14.

Alkaline phosphatase activity for DPSC, SHED, and PDLSC at days 1, 4, 7, 10, and 14. Values were normalized to total protein content and expressed as mean ± SE ηmol PNPP/min/mg protein, n = 3. *P < 0.05 (Two-way ANOVA–post-test Tukey's). PDLSC, periodontal ligament stem cell; DPSC, dental pulp stem cell; SHED, stem cells from human exfoliated deciduous teeth.
AR staining was used to ascertain the presence of mineralized nodules formed by the three cell osteogenic lineages (Figs. 2 and 3). At day 7 there were subtle differences between the cell lineages cultured in a control media versus those in an osteogenic media. Those in the osteogenic media showed enhanced proliferation with a greater uptake of background AR staining. No nodular formation was observed at this time point. There were no significant differences in quantification of staining between each of the cell types, (Fig. 4A). By day 14 there was noticeable nodule formation in the PDLSC osteogenic culture. Quantification showed significantly greater staining in PDLSC > SHED > DPSCs, (Fig. 4B). By day 21, there was gross mineralization of the whole well for the PDLSC osteogenic culture. Nodule formation was now visible in the SHED osteogenic culture. Evidence of very early nodule formation was observed at magnification for the DPSC osteogenic culture. Also, of note, was the early nodule formation at magnification in the control PDLSC culture. Quantification again showed significantly greater staining in PDLSC > SHED > DPSCs, (Fig. 4C).
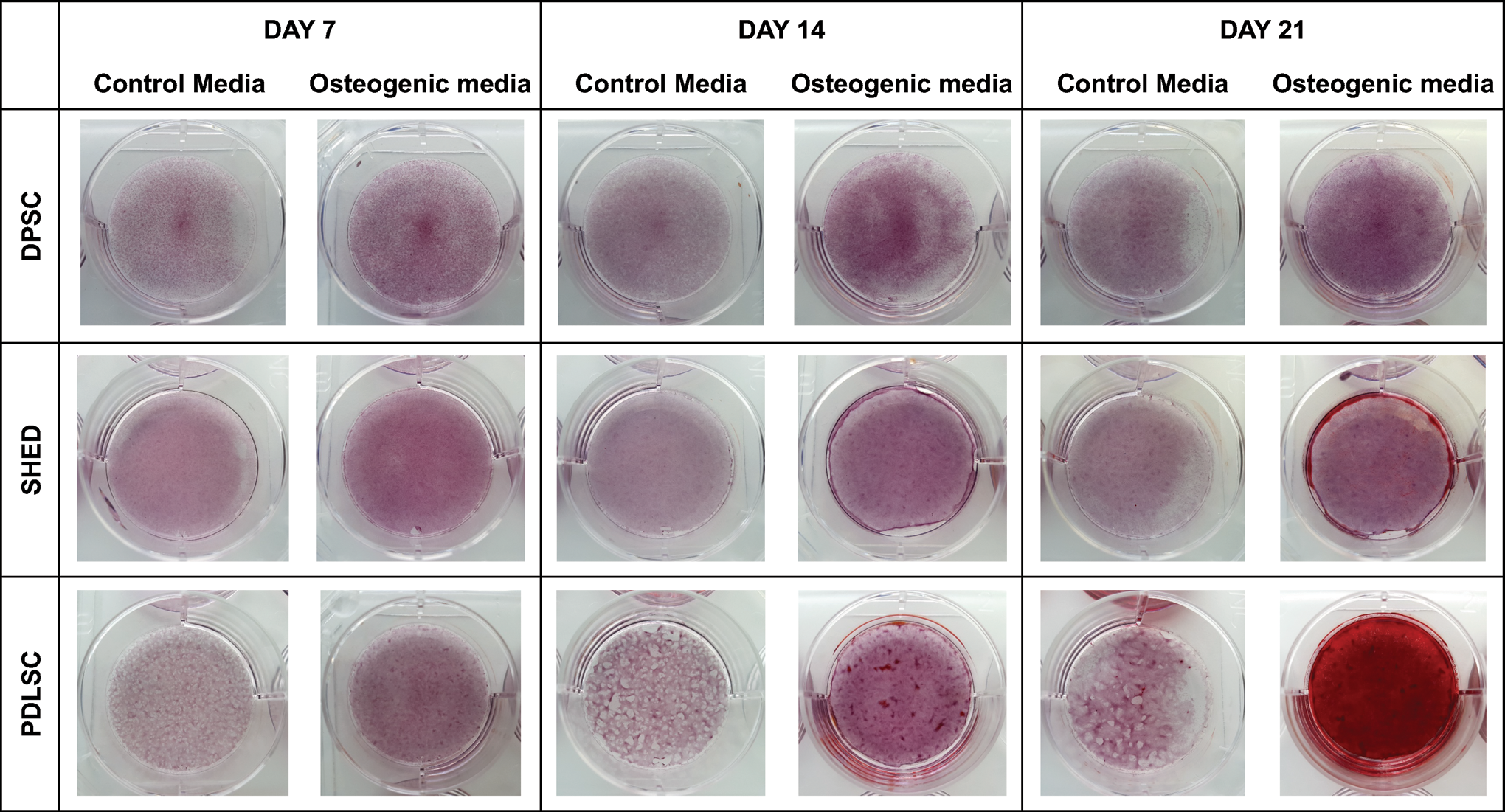
Alizarin red staining images for DPSCs, SHEDs, and PDLSCs cultured in both a control media and osteogenic media on days 7, 14, and 21. Representative well view. Color images are available online.

High magnification ( × 10) alizarin red staining images for DPSCs, SHEDs, and PDLSCs cultured in both a control media and osteogenic media on days 7, 14, and 21. Color images are available online.
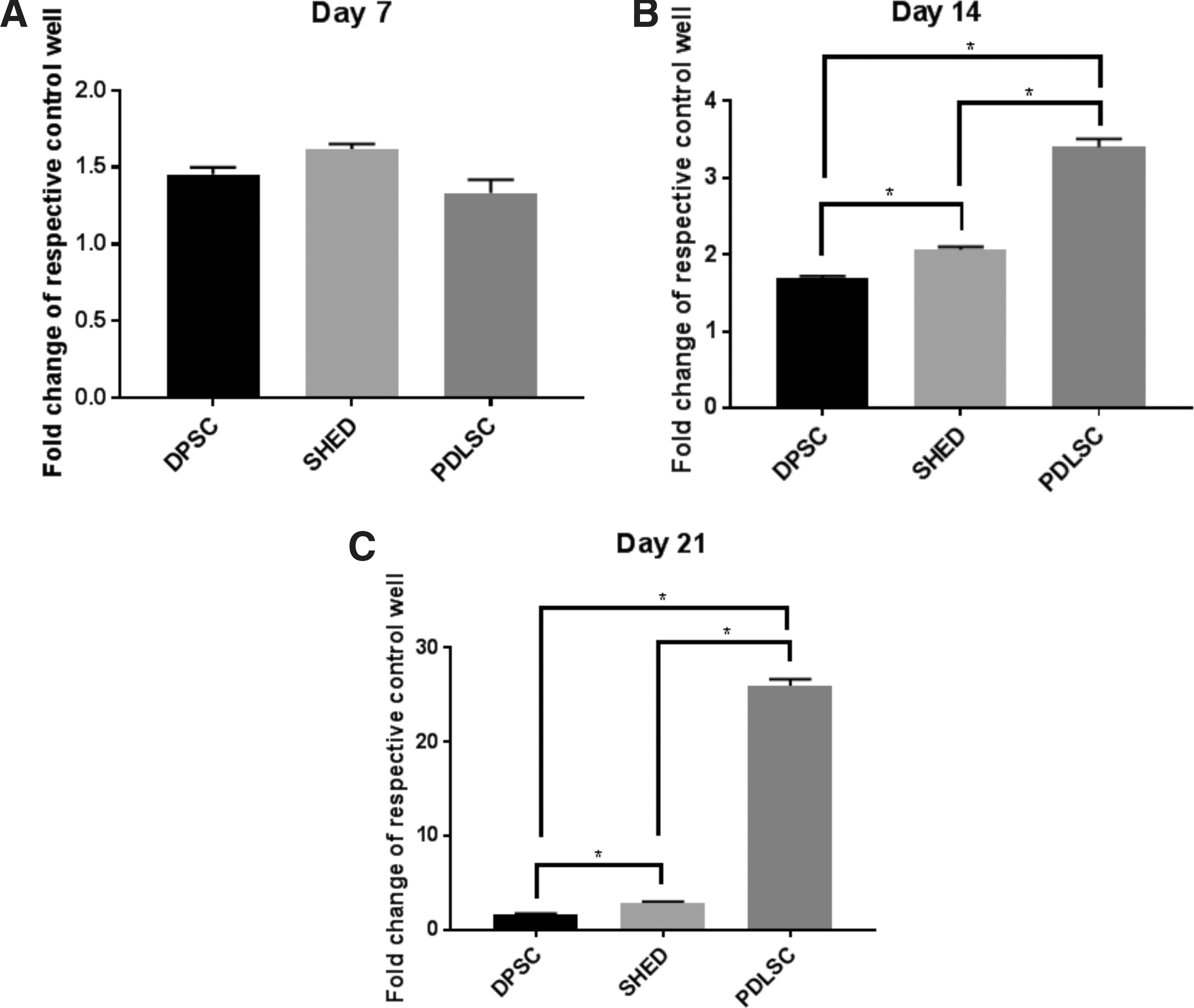
Alizarin Red staining quantification assay for DPSC, SHED, and PDLSC at days 7
The osteogenic phenotype of DPSCs, SHEDs, and PDLSCs following culture in an osteogenic media was confirmed at the transcriptional level by genes encoding ALP, COL1A1, RUNX2, and SPP1, (Fig. 5). Across the four genes studied the ALPL expression provided the most interpretable information to what was being observed functionally: There were no significant differences in gene expression at Day 1. At day 4 there was a stepped pattern with PDLSCs having a ×8.6-fold increase in ALPL expression, SHEDs a ×4.1 increase, and DPSC × 1.3. By day 21 there was a ×25.2-fold increase for PDLSCs, ×8.5-fold increase for SHEDs, and a ×3.0-fold increase for DPSCs. Also, of note, was the gene expression for COL1A1. PDLSC showed a modest early upregulation of COL1A1 with a ×2.7-fold increase versus ∼ × 1.0 for both types of pulp stem cell. A similar pattern was observed at Day 4 with significantly greater upregulation of ×3.7 for PDLSCs, although expression in DPSCs and SHEDs had now increased to × 1.8 for both. By day 7 upregulation for PDLSCs had fallen to ×2.7, with no significant difference to DPSCs (×2.5). Both DPSCs and PDLSCs had significantly greater expression than SHEDs (×1.8). Although statistically significant differences were observed for both RUNX2 and SPP1, fold change values were small (<2.0) across groups.

Bone-specific gene expression for ALPL
Discussion
The results of this study suggest that the in vitro osteogenic potential of PDLSCs is superior to SHEDs, which is in turn superior to DPSCs, (PDLSCs > SHEDs > DPSCs). This was demonstrated at a functional level by intracellular ALP enzyme activity and additionally by mineralized nodule deposition with AR staining. At a gene level, results reflecting the functional findings were observed for the osteogenic associated genes ALPL and COL1A1. This study provides evidence that despite the anatomical proximity of their tissues of origin, dental MSCs represent distinct MSC populations with respect to their in vitro osteogenic potential. This may have implications for in vivo dental MSC bone tissue engineering applications.
Although this is a novel study, comparability to results from previous studies can be inferred: Chadipiralla and coworkers showed that PDLSCs had a greater osteogenic potential than SHEDs [18]; and Wang and colleagues showed that SHEDs had a greater osteogenic potential than DPSCs [20]. The novelty of the present study relates to the fact that all three cell populations were examined concurrently, and a robust experimental design was used. An important aspect of this design was the use of a single donor to yield the three cell populations. Although it could be argued that the generalizability of the findings from a single study may be limited, this is somewhat outweighed by the numerous advantages offered by a single donor approach. In studies comparing efficacies of different cell types where cells have been harvested from different donors, it is difficult to determine if observed functional differences are indeed due to different cell phenotypes or alternatively due to genetic variability relating to the use of different donors. Connected with this, if using donors of different ages, studies have shown that there is a donor age-related decrease in osteogenic differentiation capacity [24,25]. Other factors which may potentially affect the osteogenic differentiation capacity when using multiple donors relate to: state of confluence; method of isolation; storage time; and conditions; properties of culture media used; and incubation environments [19]. It has also been found that epigenetic and genetic alterations occur during culture expansion (including passaging), which can unequivocally affect osteoblastic lineage commitment/differentiation [26].
A clear pattern unfolded with regards to mineralization and nodule formation by AR staining. PDLSCs readily formed nodules at day 14, and by day 21 there was complete mineralization of the well. SHEDs, behind PDLSCs, had clear nodule formation by day 21, whereas DPSCs had only very early evidence of nodule formation when observed under × 10 microscope. Of interest, PDLSCs kept in a control media also started to show signs of nodule formation at day 21. This is potentially initiated by the high adhesion forces occurring between the cells after lengthy confluency, making the monolayer contractile, and hence behaving more like a tissue and conferring PDLSC's role to tissue homeostasis and repair. Quantification of the AR staining showed a clear pattern with significantly greater staining uptake in PDLSC>SHED>DPSC osteogenic cultures at both day 14 and day 21.
Undifferentiated MSCs show weak ALP activity, whereas fully differentiated osteoblasts have higher activity [27]. The results indicate that PDLSCs had significantly higher ALP activity than both types of pulp stem cells at days 4, 7, 10, and 14. SHEDs had significantly greater ALP activity than DPSCs at days 7, 10, and 14. The overall pattern observed suggests that all three phenotypes were committing to an osteoblastic type lineage although at different rates: PDLSCs at a greater rate than SHEDs and DPSCs, and SHEDs at a greater rate than DPSCs. At the gene level, the ALPL codes for the production of alkaline phosphatase. The upregulation of alkaline phosphatase is generally regarded as an early marker of osteogenic differentiation. The results mirrored what was being observed at a functional level specifically for ALPL. At day 4 there was a stepped pattern with PDLSCs having a ×9-fold increase in ALPL expression, SHEDs a ×4 increase, and DPSC remaining at ×1. By day 21 there was a ×25-fold increase for PDLSCs, ×8-fold increase for SHEDs, and a ×3-fold increase for DPSCs. Also, of note, was the gene expression for COL1A1, which codes for production of type I collagen and plays an important role in cell adhesion, proliferation, and differentiation of the osteoblast phenotype. PDLSC showed a modest early upregulation of COL1A1 with a ×3-fold increase versus ×1 for both types of pulp stem cells. The interpretation of the gene expression for RUNX2 and SPP1 genes was inconclusive. Results of the osteogenic gene expression analysis add further evidence to what was being observed functionally: PDLSCs were further along the osteogenic differentiation pathway than SHEDs, which were in turn further along than DPSCs.
A potential explanatory factor for the observed results relates to the likely heterogeneity of cells within a STRO-1 +ve sorted cell population. Heterogeneity within a sorted population may give rise to differences in proliferative capacity, as well as multipotency [28]. No ideal technique of cell sorting has been identified that will result in perfectly homogenous populations of MSCs [29]. This is a recognized limitation across efficacy type studies comparing populations of MSCs derived from different tissues. STRO-1 +ve cells derived from the PDL, in particular, may contain a subpopulation of STRO-1+ve preosteoblast type cells [30], which could potentially contribute to increased ALP activity. However, given that starting values for ALP activity were comparatively low across all three cell types (Fig. 1), this was unlikely to have been a major factor.
Dental MSCs are known to be of ectomesenchymal origin and are considered to share a common lineage of being derived from neural crest cells [31]. Most dental MSCs identified thus far have generic MSC-like properties, including expression of marker genes and differentiation into mesenchymal cell lineages (osteoblasts, adipocytes, and chondrocytes) in vitro and, to some extent, in vivo [32]. Despite similarities across dental MSCs, phenotypic differences do exist. The difference in osteogenic potential found in the present study most likely relates to the phenotypic differences across PDLSCs, SHEDs, and DPSCs [17]. If we consider the tissue of origin of each of these types of cells, it would seem logical that PDLSCs should have a greater osteogenic potential due to the PDL's intimate relationship with alveolar bone. This would also agree with the premise that the function of PDLSCs is thought to be repair of damaged tissue, including regeneration of lost alveolar bone [33]. Based on their anatomic location, it should follow that DPSCs would be more likely to follow an odontoblastic route (for dentine repair), rather than osteogenic route. Although several publications indicate the osteogenic potential of DPSCs [15], comparative gene expression studies with DPSC-derived osteoblast-like cells and normal osteoblasts derived from BMMSCs suggest significant molecular differences [34]. A sensitivity analysis (Supplementary Data) was performed to investigate gene expression of dentin sialophosphoprotein (DSPP) across the three cell types during osteogenic stimulation (Supplementary Fig. S1). The DSPP gene provides instructions for synthesis of the protein dentin sialophosphoprotein, which is an important component of dentine. Relative to PDLSCs, the fold change expression of DSPP by both SHEDs and DPSCs was higher at Day 1 (×1.0 for PDLSCs and ×3.0 for both SHEDs and DPSCs) and at Day 7 (×2.2 for PDLSC, ×5.8 for SHED, ×4.4 for DPSC). This gives some indication that both DPSCs and SHEDs may have been committing to more of an odontoblastic or odonto/osteoblastic lineage compared to that of a predominantly osteoblastic lineage in PDLSCs.
Mineralization and osteogenesis from DPSCs also differ markedly from what has been reported with BMMSCs, with osteo-induced DPSCs resembling osteo-dentin rather than an osteoblast [17]. Tatullo and colleagues recently carried out a study comparing the osteogenic potential of DPSCs versus Periapical Cyst Mesenchymal Stem Cells (PCy-MSCs) [35]. Following osteogenic induction DPSCs seem to be directed toward dentinogenesis, while PCy-MSCs were directed toward osteogenesis.
SHEDs, considered immature DPSCs, do however have strong osteogenic capacity to generate bone following in vivo transplantation, as opposed to DPSCs which generate dentin [9]. This may relate to a greater degree of “stemness” and pluripotency in comparison to DPSCs [20]. Further to this, an important comparison between SHEDs and DPSCs is the significantly higher proliferation rate of SHEDs [36]. There is evidence to suggest that the expression of osteoblast related genes may occur faster in highly proliferative MSC populations [37]. At a molecular level, differences in populations of MSCs derived from the pulp of both deciduous and permanent teeth have also been investigated [38]. Using tailored microarray panels, RT-qPCR, western blot, and immunohistochemistry, it was found that high mobility group AT-hook 2 (HMGA2) was strongly expressed in deciduous pulp but not in permanent tooth pulp cells. Furthermore, genes associated with mitosis were strongly expressed in deciduous pulp cells, but in comparison matrix gene and signaling molecules were more highly expressed in the adult pulp cells. The authors concluded that based on molecular level findings MSCs derived from deciduous teeth may be the more appropriate choice for tissue engineering than cells from permanent teeth [38].
A question raised, based on this evidence, is whether PDLSCs from deciduous teeth possess a greater osteogenic potential than those from permanent teeth. Khoshhal and colleagues carried out a study comparing the osteogenic properties of PDLSCs from deciduous teeth versus PDLSCs from permanent teeth [39]. Teeth were sourced from different subjects, with osteogenic potential assessed by AR staining and ALP activity. Results showed that following osteogenic induction, both isolated cell populations were able to form colonies and differentiate into osteoblasts. When comparing the two cell populations, there was a higher ALP activity in PDLSC cultures from permanent teeth. However, AR quantitative analysis revealed no statistically significant differences in mineralization potential between the two cell types. The results of this study contrast with a study carried out by Silvério and colleagues [40]. Isolates of permanent PDLSCs and deciduous PDLSCs collected from different donors were again compared for their osteogenic potential. When examining osteogenic gene expression by qPCR, they found higher mRNA levels for ALPL and bone sialoprotein (BSP) expression in deciduous PDLSCs compared to permanent. This suggested a greater osteogenic potential of deciduous PDLSCs. The limited number of studies and ambiguity in results mean that no conclusion can be made on the osteogenic potential of PDLSCs from permanent teeth versus deciduous.
Conclusion
PDLSCs showed a significantly higher in vitro osteogenic differentiation potential than both SHEDs and DPSCs evidenced by functional studies and gene expression. This may be of significance for the use of dental MSCs in bone tissue engineering applications. Further studies are warranted to validate whether these in vitro results are replicated in an in vivo environment.
Footnotes
Author Disclosure Statement
The authors declare that there are no conflicts of interest. The study was supported by a research grant from the British Society of Periodontology.
Supplementary Material
Supplementary Data
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.