
Abstract
Nonobstructive azoospermia (NOA) is a severe form of male infertility, with limited effective treatments. Urine-derived stem cells (USCs) possess multipotent differentiation capacity and paracrine effects, and participate in tissue repair and regeneration. The aim of this study is to investigate whether the transplantation of USCs or USC exosomes (USC-exos) could promote endogenous spermatogenesis restoration in a busulfan-induced NOA mice model. USCs were cultured and characterized by flow cytometry. High-density USCs were cultured in a hollow fiber bioreactor for exosomes collection. USC-exos were isolated from USCs conditional media and identified by transmission electron microscopy, western blotting, and Flow NanoAnalyzer analysis. USC-exos exhibited sphere- or cup-shaped morphology with a mean diameter of 66.5 ± 16.0 nm, and expressed CD63 and CD9. USCs and USC-exos were transplanted into the interstitial space in the testes of NOA mice per the following groups: normal group; groups treated with no injection, phosphate-buffered saline (PBS), USCs or USC-exos on days 3 and 36 after busulfan administration, respectively. Thirty days after USCs and USC-exos transplantation, spermatogenesis was restored by both USCs and USC-exos in NOA mice 36 days after busulfan treatment as confirmed by immunofluorescence staining and hematoxylin and eosin staining. Moreover, spermatogenic genes (Pou5f1, Prm1, SYCP3, and DAZL) and the spermatogenic protein UCHL1 were significantly increased in both the USCs 36 and USC-exos36 groups compared with the PBS group, as demonstrated using quantitative real-time polymerase chain reaction and western blot analysis. However, the transplantation of USCs or USC-exos at day 3 after busulfan treatment did not improve spermatogenesis in NOA mice. Our study demonstrated that USCs could facilitate endogenous spermatogenesis restoration of busulfan-induced NOA mice through paracrine exosomes but could not protect the mouse testicles at the early stage of destruction caused by busulfan. This study provides a novel insight into the treatment of NOA.
Introduction
Azoospermia, which is defined as the absence of spermatozoa in ejaculated semen on at least two separate occasions (World Health Organization), affects ∼1% of men [1] and 15% of infertile men [2]. Azoospermia can be classified as obstructive azoospermia and nonobstructive azoospermia (NOA). Almost 60% of azoospermia men experience NOA [3].
Currently, the treatment options for NOA include hormone therapy, antioxidants, and microdissection testicular sperm extraction [4]. However, at least half of NOA patients have no effective treatment. In recent years, stem cells have brought hope for the treatment of NOA. Embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs), and spermatogonial stem cells (SSCs) have been reported to be able to produce sperms in animal studies [5 –7]. However, ethical concerns and tumor formation risk restrict the usage of ESCs and iPSCs in the clinic. Although SSCs successfully induced haploid production in vitro and in vivo [7,8], long-term culture of human SSCs in vitro remains difficult. Many studies have explored the effects of mesenchymal stem cells (MSCs) in the treatment of NOA. Nayernia claimed that mouse bone marrow stem cells (BMSCs) could be differentiated into male germ cells by transplanting them into NOA mice [9]. However, Lassalle et al. believe that BMSCs cannot be transdifferentiated into spermatogenic cells [10]. Therefore, the effect of MSCs on NOA is controversial, and the mechanisms are unclear.
Recently, we successfully isolated novel MSC-urine-derived stem cells (USCs) by using a noninvasive and low-cost method [11]. Our previous studies have demonstrated that USCs were an optimal cell source for cell therapy of urological diseases [11], and the main mechanism of USCs for cell therapy was paracrine action [12].
Exosomes, which are nanosized particles with a diameter range of 20–150 nm, are secreted by all cell types [13]. Exosomes act as key mediators of cell paracrine effects by carrying miRNAs, mRNAs, proteins, and lipids into target cells [13], and exogenous exosomes may regulate the recipient cell function by regulating target gene or protein expression [14]. Some studies have demonstrated that exosomes could directly stimulate stem cell-like proregenerative effects in injured tissues [15]. Considering that USCs can secrete many growth factors [16] and that exosomes are important mediators of cell activity [17], it would be meaningful to explore whether USCs or USC exosomes (USC-exos) have the ability to restore spermatogenesis.
In this study, we investigated the effects and mechanisms of USCs and USC-exos on spermatogenesis in an NOA mouse model induced by busulfan.
Materials and Methods
Experimental animals and ethics statement
Human urine samples were obtained from six healthy male volunteer donors. All donors provided informed consent. The procedure was approved by the Sun Yat-sen University Health Sciences Institutional Review Board. Forty-eight male C57BL/6 mice (8–10 weeks old) were purchased from the Animal Centre of Sun Yat-sen University (Guangzhou, Guangdong Province, P.R. China). The animals were kept in a standard, pathogen-free environment on a 12-h light/12-h dark cycle with free access to laboratory chow and water. The animal procedures were approved by the Institutional Animal Care and Use Committee of Sun Yat-sen University.
Isolation, culture, and identification of human USCs
USCs were isolated from healthy males as in our previous methods [16]. In brief, 20 voided urine samples (100–400 mL) were collected from six healthy male donors (20–35 years old). Urine specimens were centrifuged at 500g for 5 min, and the supernatant was removed. The cell pellets were resuspended in mixed medium composed of embryo fibroblast medium and keratinocyte serum-free medium (1:1 ratio) per our previous methods, and plated in 24-well plates (p0) [18]. The medium was changed every 2 days. Primary USCs were observed with an inverted phase-contrast microscope (Nikon TS100; Tokyo, Japan). Independent clones appeared 3–5 days after plating. It took 7–10 days for a single USC clone to reach confluence in the initial culture (p0), and then the cells were serially passaged every 3–4 days. Each single-cell clone was trypsinized and transferred into six-well dishes when the cells reached a confluence of 70%–80% (p1). Finally, cell cultures were transferred to a 150-mm culture dish (p2) for expansion; USCs (<p5) were used for all experiments. Human USCs were identified as previously described [16].
High-density USCs culture in a hollow fiber bioreactor for collecting exosomes
To collect a sufficient amount of exosomes of USCs for in vivo transplantation [19], a hollow fiber bioreactor (FiberCell Systems, Inc., New Market, MD) equipped with a cartridge was used. The cartridge was treated sequentially with phosphate-buffered saline (PBS), serum-free Dulbecco's modified Eagle medium (DMEM), and DMEM (high glucose) with 10% exosome-free fetal bovine serum (FBS) and 1% penicillin/streptomycin, for each 24 h. Bovine exosomes of FBS were depleted by ultracentrifugation at 100,000g for 18 h [20]. Then, a total of 1.8 × 108 USCs were seeded into the cartridge and incubated with complete medium containing exosome-free FBS. The glucose level of the medium was measured by a blood glucose meter (Yuyue 580; Jiangsu, China) every day to evaluate the cell growth rate, and culture medium was changed once the glucose level decreased to half the original level. When the glucose level dropped to 50% within a day, the medium was changed to mixed medium composed of keratinocyte-serum-free medium and progenitor cell medium in a 1:1 ratio. Afterward, exosome-containing medium was collected from the cartridge every day for ∼1 month.
Isolation and identification of USCs-exos
Detailed procedures to isolate exosomes from USCs medium are shown in Fig. 1C [21]. In brief, when the glucose level dropped by >50% within a day, conditioned USCs medium was collected and centrifuged at 300g for 10 min and 2,000g to remove cells. Then, the supernatant was centrifuged at 12,000g for 30 min to remove cell debris. The supernatant was ultracentrifuged (BECKMAN COULTER Optima XE-100 Ultracentrifuge 100 Ti; CA) at 100,000g for 70 min to pellet small vesicles that contain exosomes. After the first ultracentrifugation, the pellet was washed in a large volume of PBS to eliminate contaminating protein, and the suspension was filtered with a 0.22-μm filter sterilized SteritopTM (Millipore). Finally, the solution was centrifuged at 100,000g for 70 min to obtain exosomes.
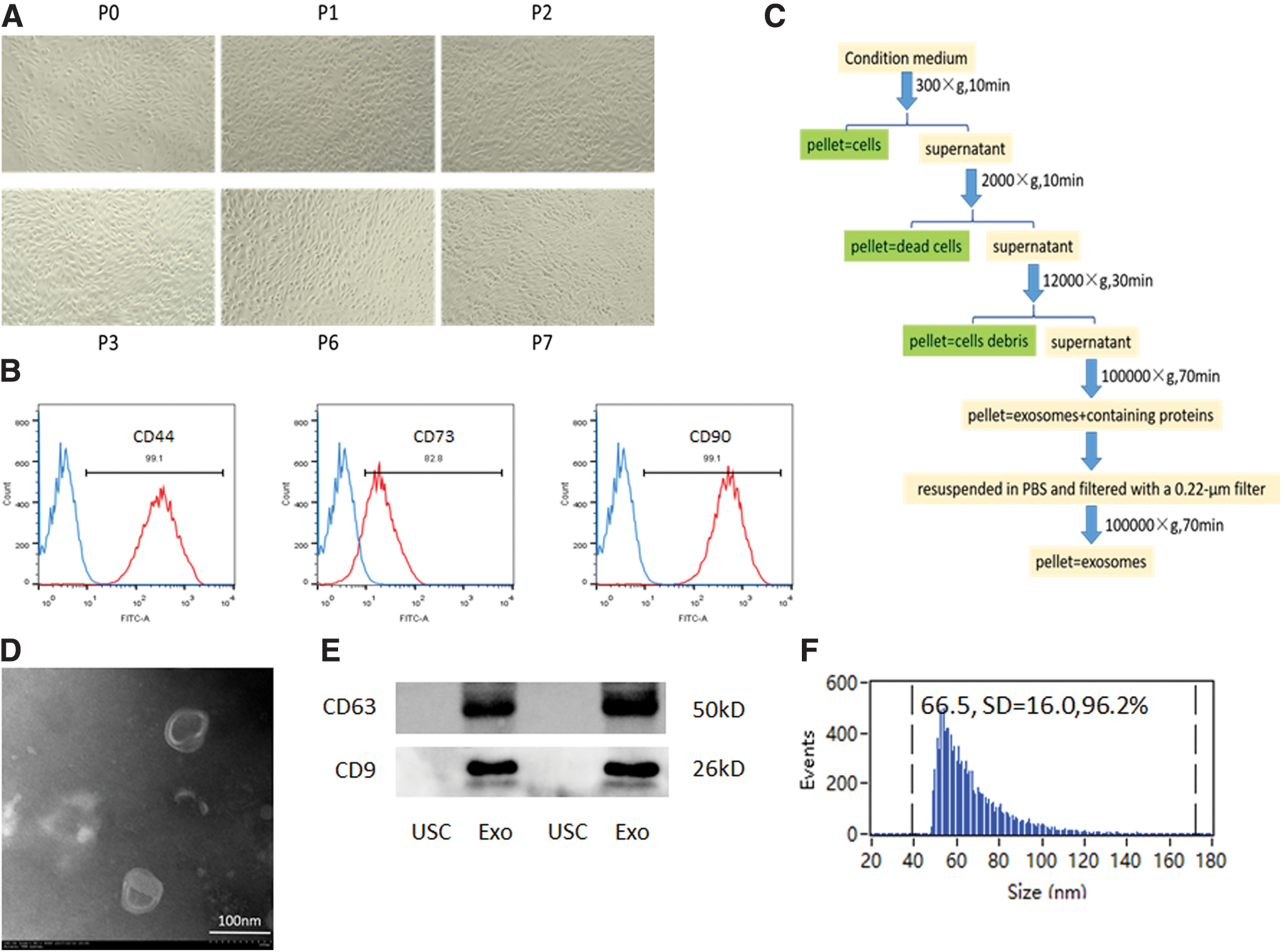
Acquisition and identification of USCs and USCs-exos.
To identify the extracellular vesicles, USC-exos morphologies were observed by transmission electron microscopy as previously described [21]. Exosomes that were obtained from USCs conditioned medium were added to the copper network. The exosomes were adsorbed on the copper mesh, stained with 2% tungstophosphoric acid, and observed under transmission electron microscopy (Japan Electron Optics Laboratory Co., Ltd.; JEM-1200EX). Exosome surface markers, including CD9 (ab92726, dilution 1:1,000; Abcam) and CD63 (ab59479, dilution 1:1,000; Abcam), were analyzed by western blot analysis. To analyze the size and number of exosomes, a Flow NanoAnalyzer N30 (NanoFCM, Inc., Xiamen, China) was used according to the manufacturer's instructions [22].
Establishment of an NOA mouse model
A total of 48 male C57BL/6 mice aged 8–12 weeks (22–25 g) were used in this study. NOA mice were developed by a modified method based on a previous report [23]. In brief, busulfan (HY-B0245/CS-2224; MedChemExpress) was dissolved in dimethyl sulfoxide (DMSO, 196055; MP Biomedicals, France) to a concentration of 20 mg/mL. To avoid DMSO toxicity and reduce mortality, we modified the modeling method by diluting the DMSO concentration to 2.5%. In detail, 39 mL of isotonic saline water was added to ∼1 mL busulfan–DMSO solution before injection. Each mouse was intraperitoneally (ip) injected with diluted busulfan–DMSO solution (40 mg/kg body weight, total dose needed was equally divided into four injections within 2 days). At 30 days after injection, mouse testes were collected for hematoxylin and eosin (H&E) staining to confirm NOA [24].
USCs and USC-exos transplantation
To prepare cell injections, cultured USCs (p3) were trypsinized. The final cell suspension (1.8 × 107 cells/mL) in PBS was prepared for transplantation.
USC-exos were enriched in a Fibercell system and isolated by ultracentrifuge as described above. USC-exos were resuspended in PBS, and concentrations of USC-exos were determined using the Pierce BCA Protein Assay Kit (Thermo 23225).
To investigate both effects of USCs and USC-exos on NOA, we treated NOA mice with USCs and USC-exos at 3 days and 36 days after busulfan treatment separately. Eight groups (n = 8) were assigned. The mice in Groups 1–4 were at day 3 after busulfan administration. Group 1 (M3 group) animals received no injection as a blank control; Group 2 (P3 group) received an injection of 10 μL PBS as a negative control; and Group 3 (U3 group) received an injection of 1 × 105 USCs in 10 μL PBS after busulfan treatment on day 3; Group 4 (Exo3 group) received an injection of 40–50 μg USC-exos in 10 μL PBS per testis after busulfan treatment on day 3. The mice in Groups 5–8 were at day 36 after busulfan administration. Group 5 (M36 group) received no injection as a blank control; Group 6 (P36 group) received an injection of 10 μL PBS as a negative control; and Group 7 (U36 group) received an injection of 1 × 105 USCs in 10 μL PBS; and Group 8 (Exo36 group) received an injection of 40–50 μg USC-exos in 10 μL PBS per testis. The above eight groups consisted of NOA mice; supplementary NOA mice were added whenever the animals died during the experiment. The ninth group (Normal group) consisted of normal mice as a positive control.
All the recipient mice were anesthetized with pentobarbital sodium (40 mg/kg, ip) for transplantation. The mice were positioned supine decubitus on the operation table and were fixed with a rubber band. A small incision was made, and 10 μL of the cell suspension (∼1 × 105 cells), 10 μL exosomes (∼40 μg), or 10 μL PBS per testis was directly injected under the tunica albuginea [25]. At 30 days after the injection, testes and epididymis of the recipient mice were collected for further analysis.
Testicular weight ratio and epididymis weight ratio
Testicular weight ratio (TWR) and epididymis weight ratio (EWR) were measured at 30 days after the injection. All mice were weighted before being anesthetized. After mice were sacrificed using pentobarbital (150 mg/kg, ip), the testes and epididymides were immediately removed and weighted. TWR was defined as testis weight/body weight (g/g). EWR was defined as epididymis weight/body weight (g/g).
Histological and immunohistological analysis
To evaluate the spermatogenesis in all groups, the spermatogenesis-specific marker protein DAZL was detected by immunofluorescence staining. Testes of all groups were fixed in 4% paraformaldehyde and embedded in paraffin for histological analysis. After embedding in paraffin, 5-μm-thick sections were cut. For immunofluorescence staining, deparaffinized testis slides were placed in a container with antigen retrieval buffer (AR0024; Boster Biological Technology Co. Ltd.). The containers were heated in a microwave at high power for 7 min and middle power for 14 min. The slides were cooled to room temperature for 30 min and washed three times in PBS for 5 min. Then, the slides were blocked for 30 min with 0.3% Triton X-100/5% bovine serum albumin in PBS and incubated with primary antibodies against DAZL (A1114, dilution in 1:100; ABclonal). After overnight incubation at 4°C, slides were washed three times for 5 min in PBS, then incubated with secondary antibodies conjugated with Cy3 (Goat Antimouse immunoglobulin G Cy3-Conjugated, BST13E18B31; Boster Biological Technology Co. Ltd.) at a 1:100 dilution for 1 h at 37°C. Finally, 10 μg/mL 4′6-diamidino-2-phenylindole (Invitrogen) staining was performed for 5 min to visualize the nuclei, followed by three washes with PBS. Images were captured with a Leica TCS SP8 confocal microscope.
To track the injected human USCs in testis 30 days after transplantation, testis slides were incubated with a primary antibody against an antinucleus antibody (MAB1281, dilution in 1:100; Sigma-Aldrich), which specifically binds to human nuclei.
Quantitative real-time polymerase chain reaction
Furthermore, the expression levels of spermatogenesis-specific genes (Table 1) were analyzed by quantitative real-time polymerase chain reaction (qRT-PCR). One month after injection, mice were sacrificed, and total RNA was extracted from testes using RNA iso Plus (Takara, Kusatsu, Japan) according to the manufacturer's instructions. Total RNA from each sample was reverse transcribed into cDNA with PrimeScript™ RT Master Mix (Perfect Real Time) (Takara). qRT-PCR was performed by using TB Green™ Premix Ex Taq™ II (Tli RNaseH Plus) (Takara). Reactions were processed and analyzed on a LightCycler® 480 System (Roche, Basel, Switzerland). Relative gene expression was calculated using the 2−ΔΔCT method, and β-actin was used as a housekeeping gene for normalization. All the primer sequences used for qRT-PCR are shown in Table 1.
Primer Sequences for Real-Time Polymerase Chain Reaction
Western blot analysis
Thirty days after USCs and USC-exos transplantation, spermatogenesis-specific markers in mice testes were analyzed by western blotting. In brief, whole protein samples were extracted, and the concentrations were measured by using the Pierce BCA Protein Assay Kit (Thermo 23225). Then, equal amounts of protein (20 μg per lane) were loaded and separated on a 10% sodium dodecyl sulfate polyacrylamide gel. Primary antibodies were used after the proteins were transferred to 0.45-μm Immobilon-P polyvinylidene fluoride Membrane; GAPDH was used as the loading control. Signals were obtained in the linear range of detection and quantified with the Bio-Rad ChemiDoc Imaging System. The primary antibodies used in this study were as follows: UCHL1 (#13179, dilution in 1:1,000; Cell Signalling) and DAZL (A1114, dilution in 1:1,000; ABclonal).
Statistical analysis
Continuous variables from the experiment were presented as the means ± standard deviation. Statistical analyses were performed using SPSS (version 20.0; SPSS IBM Corp., Armonk, NY) and GraphPad Prism version 5.0 (GraphPad Software, La Jolla, CA). Multiple groups were compared by using one-way analysis of variance. Differences between two groups were evaluated by using Student's t-test. Values of P < 0.05 were considered statistically significant.
Results
Characterization of cultured USCs
USC colonies appeared 3–5 days after plating. Primary USC colonies exhibited a typical rice-shaped appearance (Fig. 1A), and they could be passaged >12 times. The cells that were used in this study were strongly positive for the MSC markers CD44, CD73, and CD90, as confirmed by flow cytometry analysis (Fig. 1B).
Massive USC-exos produced by hollow fiber bioreactor and their characterization
USC-exos exhibited a typical cup or sphere shape under transmission electron microscopy (Fig. 1D). Western blot analysis showed that USC-exos were enriched for the typical exosomal markers CD63 and CD9 (Fig. 1E). Importantly, USC-exos exhibited an ideal mean diameter of 66.5 ± 16.0 nm (Fig. 1F) and a concentration of 3.82 × 1010/mL, as detected by Flow NanoAnalyzer. By using the Pierce BCA Protein Assay Kit, the final concentration of USC-exos was 4.2 μg/μL.
NOA mice model establishment
The survival rate of mice after the administration of busulfan was 100%. No mice died during the experiment.
NOA mice were confirmed by H&E staining. As shown by H&E staining, most of the seminiferous epithelium became thinner in testes of NOA mice (Fig. 2C[b, g]) compared with normal mice (Fig. 2C[a, f]). Spermatogenic cells, including spermatocytes, spermatids, and spermatozoa, were completely cleared.
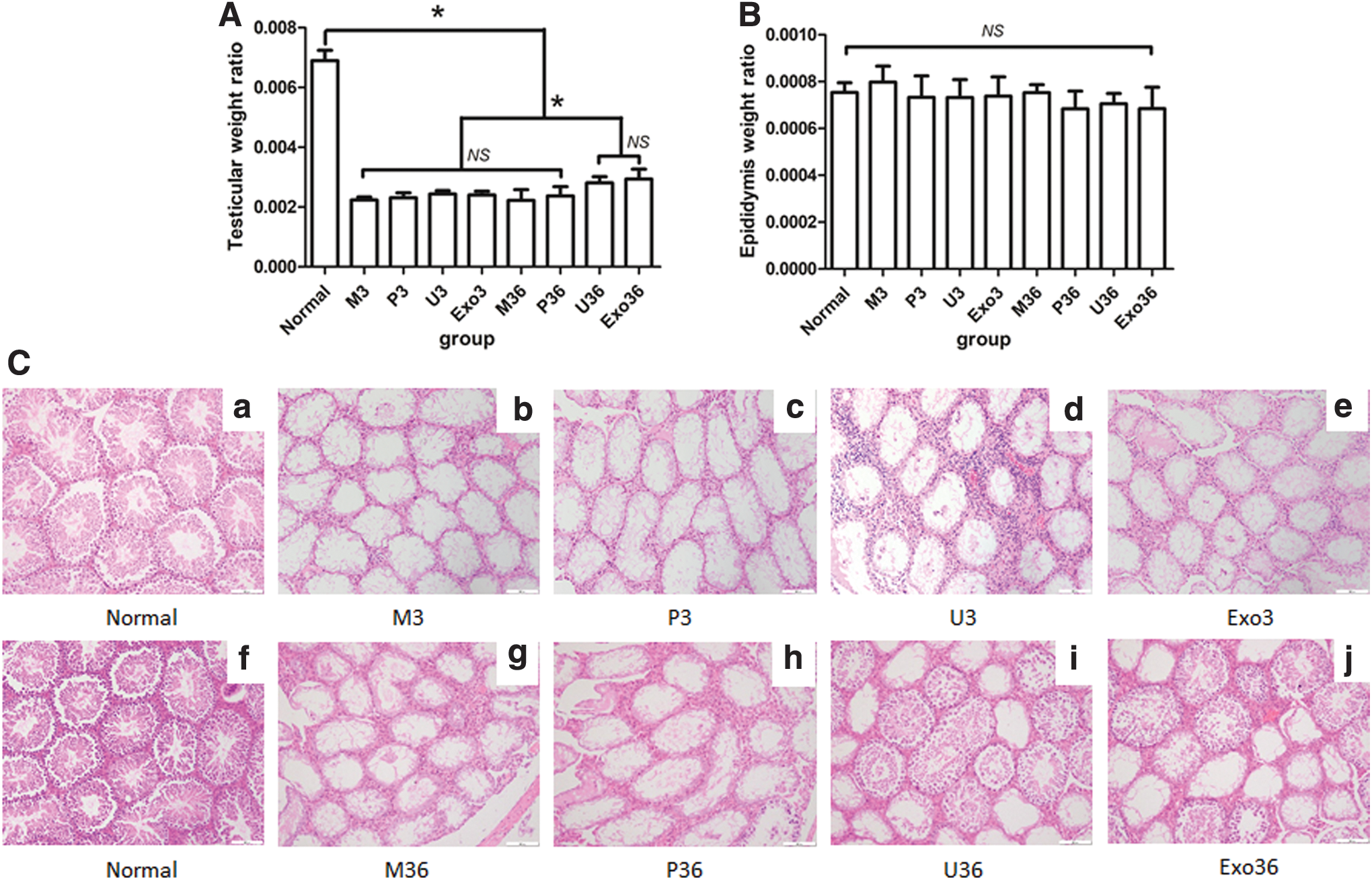
TWR and EWR and H&E staining of testis. Comparison of TWR
TWR and EWR changes
Thirty days after USCs and exosomes injection, all mice were sacrificed for spermatogenesis detection. No immune rejection response occurred in xenotransplantation after human USCs were transplanted into NOA mice.
Body weight and testis weight of groups were showed in Supplementary Figure S3. TWR and EWR were detected to evaluate the effect of busulfan on NOA mice. TWR was significantly decreased in the blank group at day 36 after busulfan administration compared with normal mice (Fig. 2A and Supplementary Figs. S1A, S2A, P < 0.05). TWR was significantly increased in the U36 and Exo36 group compared with negative control mice (P < 0.05). The EWR was not significantly different in all eight groups (Fig. 2B and Supplementary Figs. S1B, S2B, P = 0.24).
Histological variation after USC and USC-exos transplantation
Compared with normal mice (Fig. 2C[a]), the seminiferous epithelium was thinner and lacked spermatogenic cells in the M3, P3, U3, and Exo3 groups, and there were no differences between the M3, P3, U3, and Exo3 groups (Fig. 2C[b–e]). After treatment with USCs and USC-exos, the seminiferous epithelium was thicker in the U36 (Fig. 2C[i]) and Exo 36 (Fig. 2C[j]) groups than in the M36 group (Fig. 2C[g]) and P36 (Fig. 2C[h]) group, which was the negative control (Supplementary Figs. S1C, S2C). However, the seminiferous epithelium in the U36 and Exo 36 groups did not recover to the same thickness as that in normal mice.
Expression of human nuclei in vivo
To track the USCs transplanted into mice testes, human nuclei (red fluorescence) were assessed to detect human USCs. No obvious expression of human nuclei was observed in the testes of the U3 group or the U36 group 30 days after transplantation (Fig. 3).
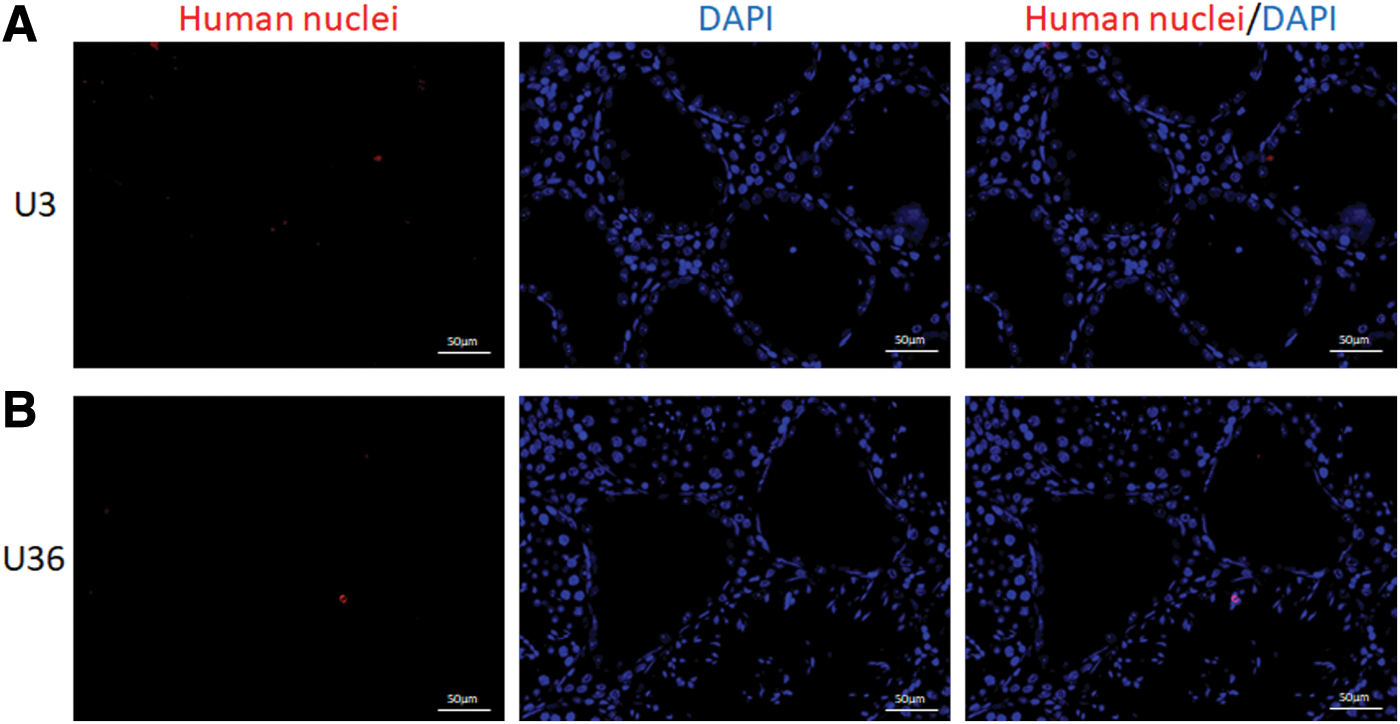
Tracking human nuclei in the testes after USCs transplantation. No obvious expression of human nuclei (red fluorescence in nuclear) was observed in the testes of U3 group and U36 group 30 days after transplantation.
Spermatogenic marker expression in mice testes
First, to evaluate the effect of USC and USC-exos transplantation, the expression of the undifferentiated spermatogonial marker DAZL (red fluorescence) in testes was determined by immunofluorescence staining. In normal mice, DAZL was expressed around the seminiferous epithelium. After busulfan treatment, the expression of DAZL was significantly decreased in the M3 group. The expression of DAZL in the U3 group or Exo3 group showed no increase compared with the M3 and P3 groups (Fig. 4A). Immunofluorescence staining of DAZL in USCs transplantation at day 0 was showed in Supplementary Figure S3.
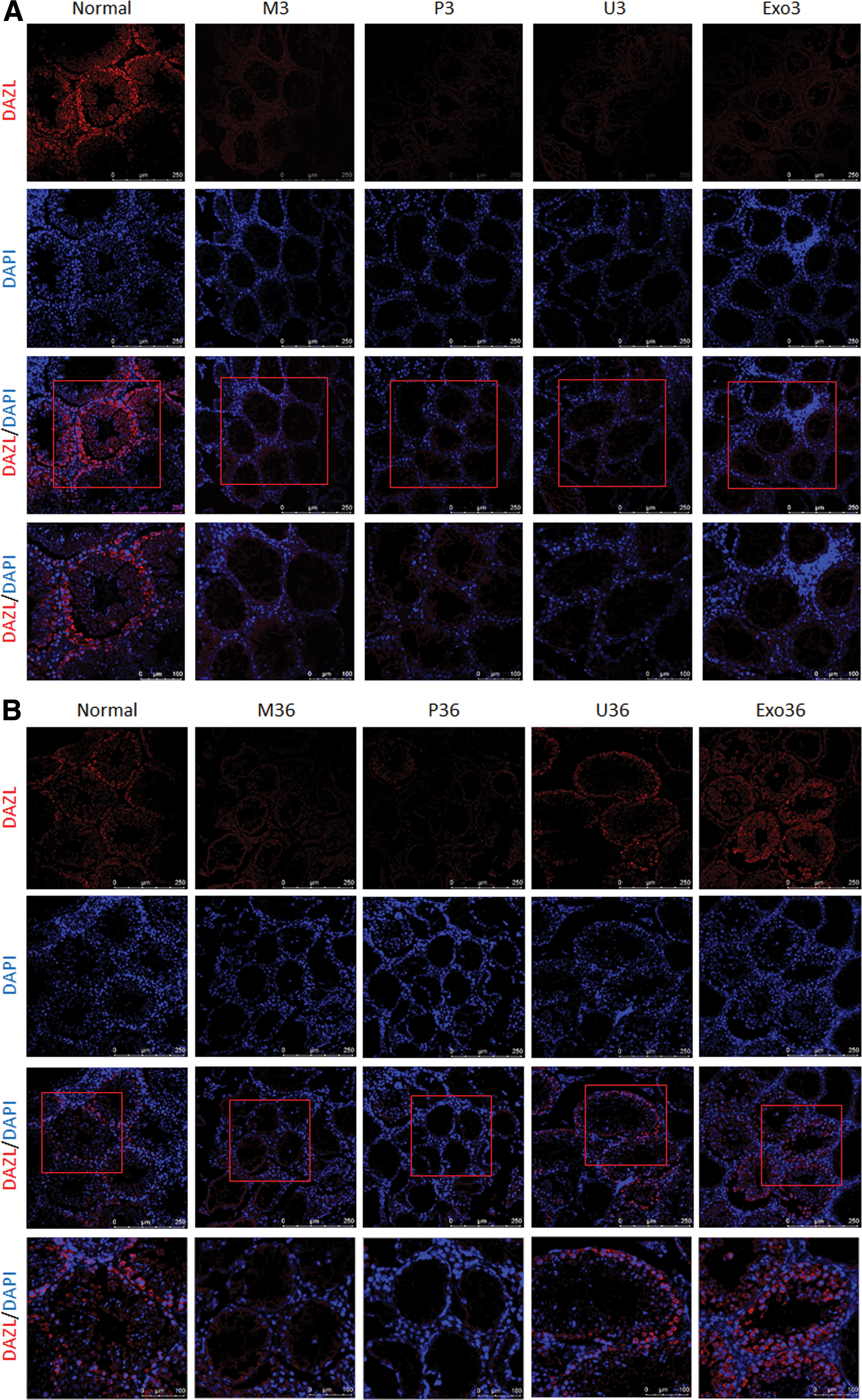
When the NOA mice received USCs and USC-exos transplantation after 36 days of busulfan administration, the expression of DAZL was significantly increased compared with the M36 and P36 groups (Fig. 4B and Supplementary Fig. S3).
Second, to further determine the spermatogenesis at different stages, spermatogenic gene expression levels (Pou5f1, Prm1, SYCP3, and DAZL) were detected in the testes 30 days after transplantation. As shown in Fig. 5, the expression of spermatogenic genes was significantly decreased in the model mice and PBS-treated mice compared with normal mice. Expression of spermatogenic genes in the U3 group or Exo3 group was not significantly upregulated compared with the M3 and P3 groups (Fig. 5A–D; P = 1.00). However, these spermatogenic genes were significantly increased in the U36 group and Exo36 group compared with the M36 and P36 groups (Fig. 5E–H and Supplementary Fig. S4; P < 0.05).
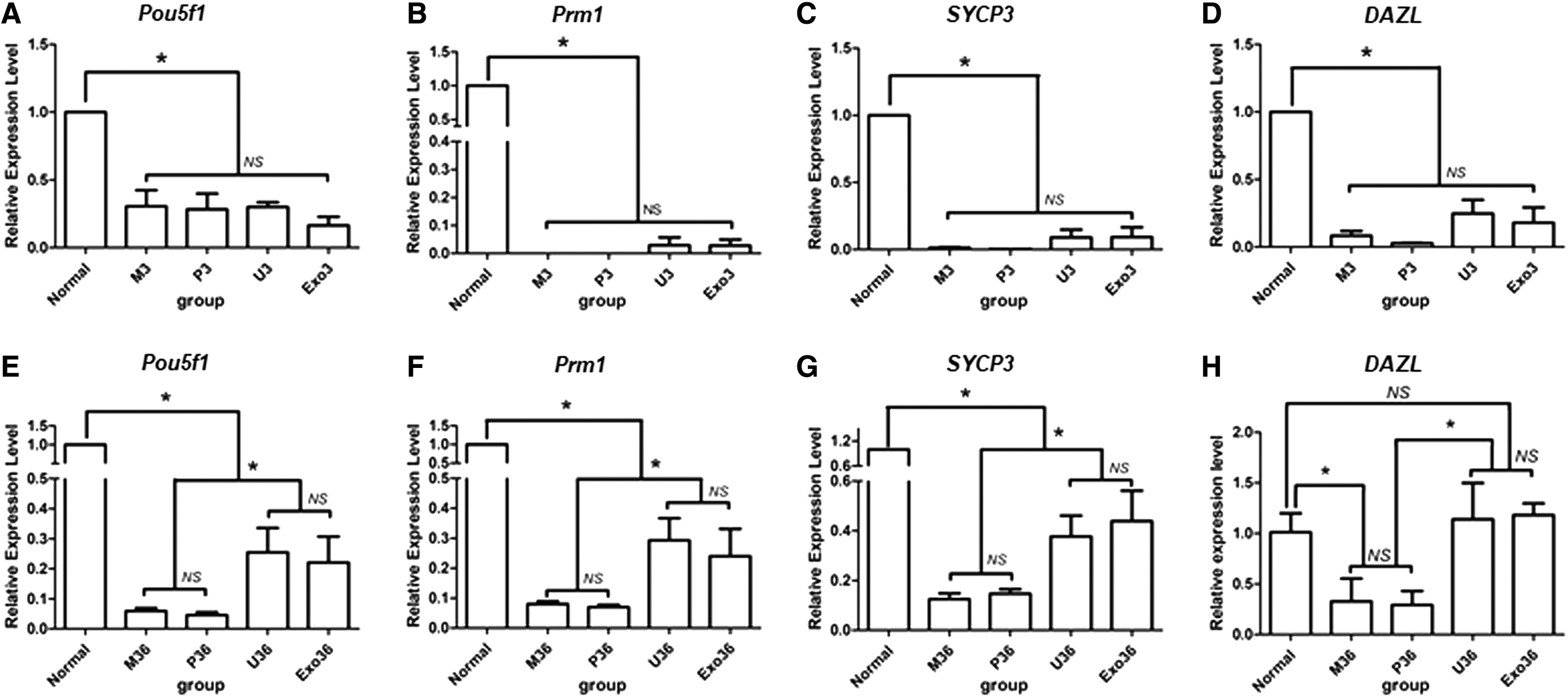
Quantitative real-time PCR analysis of germ cell genes (Pou5f1, Prm1, SYCP3, and DAZL).
Finally, to further confirm the expression levels of spermatogenic cell-specific markers, the expression of the spermatogenic markers UCHL1 and DAZL was detected by western blotting. The expression of spermatogenic markers UCHL1 and DAZL in the U3 group and the Exo3 group is not significantly different from the M3 and P3 groups (Fig. 6A–C and Supplementary Fig. S5; P < 0.05).
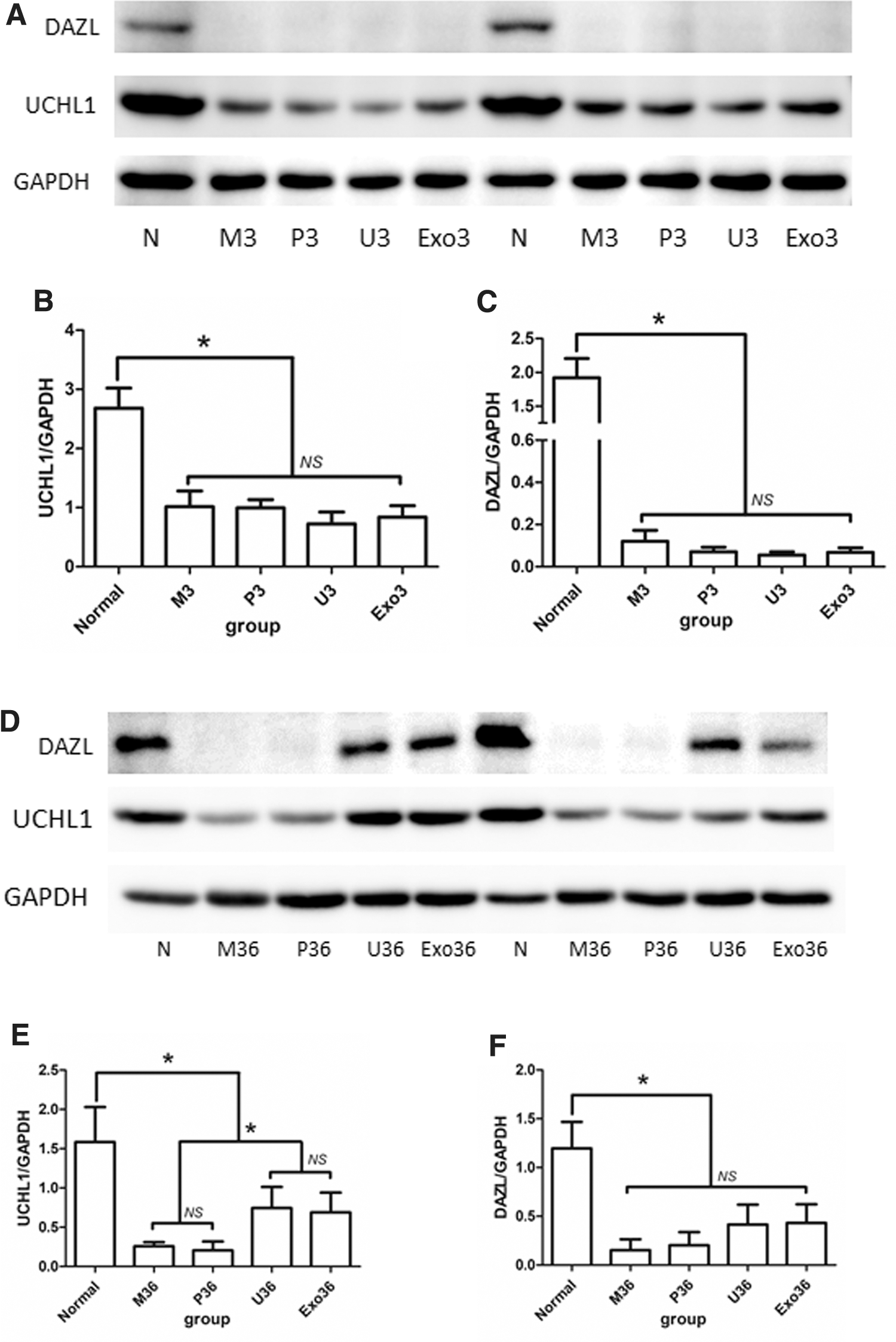
Western blot analysis of germ-cell-specific genes UCHL1 and DAZL in Normal, M3, P3, U3, Exo3 group and M36, P36, U36, Exo36 group.
The expression of spermatogenic marker UCHL1 was significantly increased in the U36 group and the Exo36 group compared with the M36 and P36 groups (Fig. 6D, E; P < 0.05), which indicated the restoration of spermatogenesis. Moreover, the expression of DAZL was increased in the U36 group and Exo36 group compared with the M36 and P36 groups, but this difference was not significant (Fig. 6D, F and Supplementary Fig. S6; P = 0.10).
Discussion
Herein, we report for the first time that USCs and USCs-exos were able to promote the spermatogenesis recovery of NOA mice induced by busulfan. Our study demonstrated that early treatment of USCs was not beneficial to busulfan-treated mice, but USCs and USC-exos could promote long-term spermatogenesis repair of NOA mice. USCs may facilitate endogenous spermatogenesis restoration of busulfan-treated NOA mice through paracrine exosomes.
Busulfan, which is an anticancer drug and myeloablative alkylating agent, has been widely used to establish an NOA azoospermia mice model [26]. Endogenous spermatogenic cells were eliminated by busulfan treatment through alkylation of DNA, disrupting distribution of vimentin filaments, impeding SSCs differentiation, promoting SSCs dormancy, and generating oxidative stress [27]. Although busulfan destroyed the vast majority of spermatogenic cells and finally resulted in azoospermia, there were still a few residual spermatogonial cells remaining, which was the foundation of endogenous spermatogenesis restoration. Fertility of the mice was lost within 4 weeks after busulfan treatment, and fertility might be restored after >30 weeks [26].
Several studies have indicated that stem cell therapy is a promising treatment for NOA. Many stem cells, such as ESCs [28], iPSCs [29], and SSCs [30], are candidates for differentiation into germ cells. However, the sources of ESCs, iPSCs, and SSCs are limited. Ethical problems and tumor formation risk should also be taken into consideration.
To date, several studies have demonstrated that transplantation of MSCs, including BMSCs and adipose-derived stem cells (ADSC), is able to restore the fertility of busulfan-treated azoospermia rodents [9,31 –33]. However, these MSCs must be obtained by invasive methods, thus limiting broad usage in the clinic. In our previous study, we obtained a new type of MSCs, named USCs, which could be isolated noninvasively from human urine and possessed a high proliferation rate, multipotent differentiation capacity, and secretion of growth factors [11]. In this study, after USCs were xenotransplanted into the testes of busulfan-treated mice, characteristic restoration of spermatogenesis was found in these mice. However, no recovery of spermatogenesis in testes was seen when USCs were transplanted immediately on the same day (U0 group) of busulfan administration. The half-life of busulfan is 3.0 ± 0.7 h [34]. In our study, we transplanted USCs into testes after this point, when the concentration of busulfan declined significantly. Residual busulfan may influence USCs directly or by acute stress response. The underlying mechanism remains to be further elucidated.
Spermatogenesis occurs in continuous mitotic, meiotic, and postmeiotic phases [35]. Large numbers of stage-specific genes were identified, and these genes played important roles in germ cell development. Pou5f1 is a pluripotency gene in germ cells. SYCP3, a component of transverse elements of the synaptonemal complex, is positive in spermatocytes. Prm1 is a haploid spermatid marker [36]. In our study, we found that the expression of these three genes was upregulated when the NOA mice were transplanted with USCs and USC-exos for 30 days. Furthermore, UCHL1 is expressed in undifferentiated spermatogonial cells, and DAZL is the marker of SSCs and spermatogonial cells [37]. At both the gene and protein levels, we demonstrated that USCs and USC-exos transplantation could promote spermatogenesis in all phases, in accordance with a previous study [33].
MSCs may restore fertility by two possible mechanisms. First, MSCs were transdifferentiated into germ cells through suitable induction. Weak evidence has shown that MSCs differentiate into germ cells [10]. However, in theory, the differentiation ability of MSCs is much lower than that of ESCs or iPSCs, making it difficult for MSCs to differentiate into germ cells. In our study, no human nuclei-labeled cells were found in NOA mouse testes, which indicated that no differentiation of germ cells from implanted USCs occurred. Second, MSCs could secrete trophic factors to stimulate the proliferation and differentiation of endogenous SSCs or recovery of spermatogenesis microenvironments. As described in previous reports, MSCs promoted tissue repair by secreting cytokines such as glial cell derived neurotrophic factor (GDNF), fibroblast growth factor 2, vascular endothelial growth factor [38], and exosomes carrying a variety of miRNAs [39]. Human umbilical cord MSCs were likely to promote spermatogenesis by indirect paracrine mechanisms [25]. Similar to previous studies [25,33], the data in this study showed that both xenotransplantated USCs and USC-exos promoted spermatogenesis recovery of NOA mice, which indicates that USCs may promote endogenous spermatogenesis recovery through paracrine effects instead of differentiation.
Moreover, exosomes-based cell-free therapy has raised great interest in recent years. Compared with cell therapy, exosomes have many advantages, including nontumorigenesis, nonimmune rejection, ease of reaching the wound sites, high stability, and no concern of vascular obstruction [40,41]. Microvesicles derived from MSCs may activate a proliferative program in surviving tubular cells after injury by carrying mRNA [42]. MSCs reduced myocardial ischemia/reperfusion injury by secreting exosomes [43]. Recent studies demonstrated that MSC exosomes may regulate protein expression of recipient cells and modify cell characteristics by miRNA transfer [44]. To our knowledge, our study is the first to explore the effect of USCs-exos on NOA mice. Our data demonstrated that transplantation of USC-exos was able to promote spermatogenesis in the testes of busulfan-induced NOA mice. The possible mechanism might lay in the abundant miRNAs carried in USC-exos, and miRNAs play an important role in the maintenance and differentiation of SSCs [45]. Although there were few residual spermatogonia in the seminiferous tubules, the transplantation of USC-exos may deliver abundant miRNAs to regulate differentiation of endogenous spermatogoniums in busulfan-induced NOA mice and finally result in the restoration of spermatogenesis. Further study is needed to further analyze the detailed mechanism of USC-exos on NOA.
Conclusion
Our study demonstrated that USCs and USC-exos promoted the restoration of endogenous spermatogenesis in busulfan-treated NOA mice, while USCs could not prevent damage caused by busulfan in the early stage. This study provides potential treatment for NOA patients, especially for NOA patients, with partial residual spermatogenic cells. The mechanism and safety of USCs and USC-exos therapy on azoospermia need further study.
Footnotes
Acknowledgments
We thank Deying Zhang for his kind help in building the busulfan-induced azoospermia mouse model, Prof. Baisong Lu and Dr. Ning Li for their kind help in the usage of the hollow fiber bioreactor, and Dr. Huashan Liu and Dr. Ru Deng for their kind help in the culture and identification of USCs. This research was funded by the following grants: the Natural Science Foundation of China (nos. 81571489, 81671834, 81671449, and 81871110), the Science and Technology Planning Project of Guangdong Province, China (no. 2016A020214004), the Natural Science Foundation of Guangdong Province (2014A030310359), the Science and Technology Planning Project of Guangdong Province (2016B030230001), the Key Scientific and Technological Program of Guangzhou City (201604020189), the Guangdong Special Support Plan-Science and Technology Innovation Youth Top Talents Project (2016TQ03R444), the Guangzhou Pearl River Science and Technology Nova Project (201806010089), and the Youth Teacher Training Project of Sun Yat-sen University (no. 17ykpy68).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.