
Abstract
Periodontium is consisted of root cementum, bone lining the tooth socket, gingiva facing the tooth, and periodontal ligament (PDL). Its primary functions are support of the tooth and protection of tooth, nerve, and blood vessels from injury by mechanical loading. Severe periodontitis induces the destruction of periodontium and results in a significant cause of tooth loss among adults. Unfortunately, conventional therapies such as scaling and root planning are often only palliative. Therefore, the ultimate goal of the treatment for periodontitis is to restore disrupted periodontium to its original shape and function. Tissue engineering refers to the process of combining cells, scaffolds, and signaling molecules for the production of functional tissues to restore, maintain, and improve damaged organs. The discovery of periodontal ligament stem cells (PDLSCs) highlighted the possibility for development of tissue engineering technology-based therapeutics for disrupted periodontium. PDLSCs are a kind of somatic stem cells that show potential to differentiate into multiple cell types and undergo robust clonal self-renewal. Therefore, PDLSCs are considered a highly promising stem cell population for regenerative therapy in periodontium; however, their rarity prevents the progression of basic and clinical researches. In this review, we summarize recent research advancement and accumulated information regarding the self-renewal capacity, multipotency, and immunomodulatory effect of PDLSCs, as well as their contribution to repair and regeneration of periodontium and other tissues. We also discuss the possibility of PDLSCs for clinical application of regenerative medicine and provide an outline of the genetic approaches to overcome the issue about the rarity of PDLSCs.
Introduction
Periodontitis, a pathological inflammatory condition caused by certain bacteria in dental plaque, leads to the destruction of periodontium. Therefore, periodontitis is now a major reason for tooth loss in adults. Tooth loss causes disorder in the individual quality of life by leading to many particularly serious esthetic and functional problems. In addition, recent studies demonstrated positive associations of periodontitis and tooth loss with systemic diseases, including dementia [1], coronary heart disease [2], and oral, lung, and pancreatic cancers [3]. Dentists have succeeded in retarding the progression of periodontitis through conventional therapies such as scaling and root planning. However, once periodontium is disrupted, restoration of its normal structure and function is very difficult.
Regeneration is defined as the reproduction or reconstitution of a lost or injured part of the body in such a way that the architecture and function of the lost or injured tissues are completely restored [4]. Therefore, the ultimate goal of periodontal therapy is to regenerate periodontium injured by periodontitis and to prevent tooth loss.
Tissue engineering is a scientific field mainly focused on restoring damaged organs to their original shape and function through a combination of cells, biological signals, and cell-seeded scaffolds [5]. Stem cells are an unspecialized cell population that possesses four defining characteristics: reproducing themselves (self-renewal), generating progeny destined to differentiate into various functional cell types (multipotency), persisting for a long time, and requiring the immediate environment, the niche [6]. Stem cells play crucial roles in an internal repair system, replacing damaged or dead tissues by reproducing and turning into the specialized cells; therefore, these cells have been regarded as highly promising cells for tissue engineering [7].
There are two main types of stem cells: embryonic stem (ES) cells and somatic stem cells. ES cells are pluripotent and illimitable stem cells originated from the inner cell mass of the blastocyst-stage embryos [8]; therefore, they are one of the most promising cell sources for tissue engineering. However, the ethical controversies and safety concerns associated with ES cells limit their utility in clinical settings. Somatic stem cells, defined as a stem cell population found in various parts of the body in juveniles and adults, are easier to isolate than ES cells [9]. They are attractive for novel approaches to treat various diseases because their primary roles are maintaining and repairing the tissues, and patient-specific somatic stem cells can be transplanted without immune rejection.
Somatic stem cells have been identified in various adult organs, including nerve, skin, pancreas, bone, muscle, heart, and dental tissues [10]. Periodontal ligament (PDL), a highly specialized connective tissue that surrounds the tooth root, plays crucial roles in anchoring tooth to the socket bone and in regulating proper tooth homeostasis, repair, and nutrition [11]. PDL cells were suggested to harbor progenitor cell population that migrated from locations close to blood vessels and endosteal spaces [12]. In 2004, mesenchymal stem cell (MSC) population was first identified in cells derived from human PDL tissue [13]. After that, many researchers also succeeded to isolate periodontal ligament stem cells (PDLSCs) not only from human but also from animals, and characterized them in detail.
The aim of this review article is to summarize the status in PDLSCs, the potential benefits of using PDLSCs to treat damaged PDL and other tissues, and future prospective of cell reprogramming application for PDLSC-based regenerative therapies for periodontium.
Self-Renewal Capacity of PDLSCs
Stem cells exhibit asymmetric cell division, which produces one daughter cell of equivalent fates (self-renewal) and one daughter cell that is committed to differentiate into nonstem cell [14]. Basically, self-renewal is a cell division process in which the capacity of progeny cells to proliferate and differentiate is closely similar to that of parental cells. As a result, stem cells could help maintenance of the undifferentiated state and regenerate tissues throughout the life of each individual.
Many researchers reported the self-renewal capacity of PDLSCs as well as other types of stem cells. Surprisingly, human PDLSCs revealed higher growth potential than human bone marrow-derived MSCs (BMMSCs) and human dental pulp stem cells (DPSCs) [15]. In addition, human BMMSCs stopped proliferation by about 50 population doublings; however, human PDLSCs maintained the proliferative capacity beyond 100 population doublings [16]. The high self-renewal potential of PDLSCs would be associated with mechanical loading; PDL tissue is continually exposed to mechanical force caused by mastication or occlusion and PDLSCs that undergo static mechanical strain promote their proliferation rate [17].
Recently, interleukin (IL)-11, angiotensin II (Ang II), and Ang II receptor type 2 (AGTR2) were suggested to be involved in the mechanical loading-induced proliferation in PDLSCs; human PDL cells exposed to mechanical loading promoted the production and secretion of IL-11 through the Ang II/AGTR2 signaling pathway, and PDL cell-derived IL-11 exerted stimulating effects on proliferation of human PDLSCs [18]. Many other factors such as hypoxia condition [19] and Rho-kinase signaling pathway [20] were reported to regulate proliferation of human PDLSCs.
In addition, young donor-derived PDLSCs exhibited higher colony-forming unit and proliferation activity than aged donor-derived PDLSCs [21]. High S and G2/M phase activities of the cell cycle have been known to correspond to rapid cell proliferation and PDLSCs from young donors showed a higher percentage of cells in S + G2/M phases compared with PDLSCs from aged donors [22]. These results suggested that cell senescence also affected the proliferative ability of human PDLSCs.
Moreover, Martinez et al. demonstrated that human PDLSCs treated with sonic hedgehog (Shh) showed higher proliferation rate than untreated cells [23]. Interestingly, they also revealed that differentiated PDL cells exhibited no change in their proliferation after Shh treatment, suggesting that Shh signaling pathway acted specifically on the self-renewal of PDLSCs.
These results exhibited the high proliferative potential of PDLSCs using simple assays such as MTT assay colony-forming assay, and immunocytochemical staining for proliferation markers; however, these assays could not assess the clonogenic capacity of stem cells. In addition, a lot of reports have evaluated the self-renewal capacity of stem cells by in vitro studies; however, stem cells are thought to reside in “stem cell niche” in vivo.
Stem cell niche is an area that provides a specific microenvironment where stem cells remain in undifferentiated and self-renewable state. Its activity is suggested to depend on cell–cell and/or cell–matrix interactions; adhesion of germline stem cells to the extra cellular matrix prevented their loss in the Drosophila ovary [24]. Furthermore, niche supporting cells and surrounding tissues has been known to secrete multiple signaling factors that regulate stem cell behavior; Shh secreted from a neurovascular bundle stem cell niche controlled MSC homeostasis in the adult mouse incisor [25] and cyclic dermal bone morphogenetic protein (BMP) signaling from the bulge stem cell niche controlled hair stem cell numbers during hair follicle regeneration in the mouse skin [26].
An in vitro stem cell culture correlates poorly with an in vivo stem cell growth; therefore, it may be difficult to assess the precise clonogenic and self-renewal capacity of PDLSCs using in vitro assays.
On the other hand, a recent report demonstrated that expanded human skeletal muscle stem cells in vitro showed the potential not only to generate chimeric muscles but also to engraft in the sublaminar stem cell niche with a great frequency when they were transplanted into immunodeficient mice [27]. Menicanin et al. subcutaneously transplanted in vitro cultured ovine PDLSCs into the dorsal surface of immunodeficient mice and re-isolated them after 8 weeks of transplantation [28]. Surprisingly, these cells could survive, proliferate, and express MSC-related markers, and differentiate into several types of mesenchymal cells in vitro. Moreover, their secondary transplants gave rise to mineralized matrices with Sharpy's fibers. Based on these in vivo and in vitro results, PDLSCs were suggested to show a high self-renewal potential both in vivo and in vitro.
Multipotency of PDLSCs
PDL tissue is mainly consisted of dental follicle-derived mesenchymal cells; therefore, PDLSCs have been reported to possess the ability to differentiate into various types of mesenchymal-lineage cells (Fig. 1). Seo et al. could differentiate human PDLSCs into osteoblast-like cells that formed Alizarin Red-positive mineralized nodules and highly expressed bone-related marker genes [13]. They also revealed the differentiation potential of human PDLSCs into adipocyte-like cells that contained lipid droplets in their cytoplasm and highly expressed adipose-related marker genes.
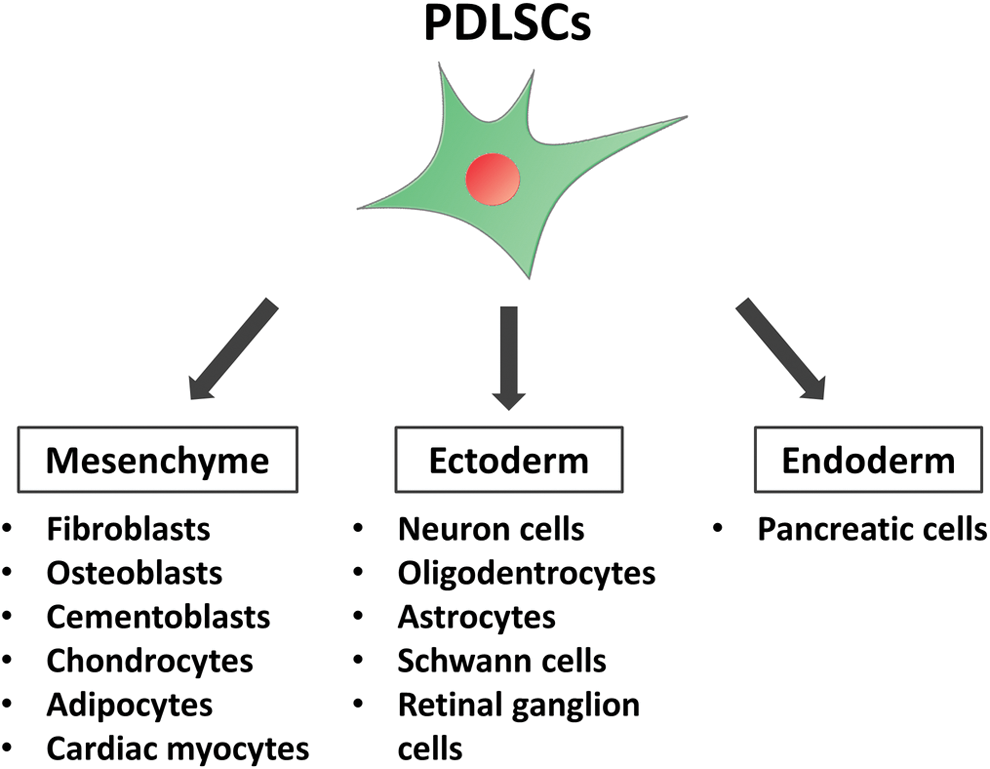
Multipotency of PDLSCs. PDLSCs were reported to differentiate into mesenchymal-, ectoderm-, and endoderm-lineage cells in vitro, suggesting their multipotency. PDLSC, periodontal ligament stem cell. Color images are available online.
Xu et al. succeeded to generate chondrocyte-like cells from human PDLSCs as well as osteoblast- and adipocyte-like cells [29]. In addition, Pelaez et al. exhibited that human PDLSCs exposed to the short-term mechanical strain could differentiate into cardiac myocytes expressing cardiac cell markers, sarcomeric actin, and cardiac troponin T proteins [30].
Interestingly, recent reports demonstrated that PDLSCs could differentiate into not only mesenchymal-lineage cells but also other lineage cells (Fig. 1). Neural cells such as neuron cells, oligodendrocytes, astrocytes, and Schwann cells are derived from ectoderm-derived cells. Kadar et al. established a new neurodifferentiation protocol applying epigenetic reprogramming, PKC/PKA activation, and neurotrophic factor stimulation [31]. Importantly, their neurodifferentiation protocol succeeded in differentiating more human PDLSCs into a neural fate than two previous protocols for human BMMSCs, suggesting that PDLSCs and their new induction protocol would be a breakthrough in the treatment of patients with damaged neuron.
Techawattanawisal et al. also developed neural cell-, oligodendrocyte-, and astrocyte-like cells using free-floating culture of rat PDLSCs [32]. Dapeng et al. demonstrated that dog PDLSCs could be induced to differentiate into Schwann cell-like cells by activation of Erk1/2 signaling pathway [33]. PDLSCs derived from adult human also showed the ability to differentiate into retinal ganglion-like cells that could form synapses and respond glutamate-induced calcium [34].
Neural crest has been known to emerge at the junction of the epidermal and neural ectoderm during development, and neural crest-derived cells then begin to migrate into their destination in the embryo and generate various types of cells, including neural cells, melanocytes, and endocrine cells [35]. PDL is derived from the cranial neural crest-derived dental mesenchyme and PDLSCs originate from a migratory population of neural crest-derived cells [36]. As both PDLSCs and neural cells are derived from the same embryologic neural crest, PDLSCs would be more likely to differentiate into neural cells than other types of stem cells.
Moreover, Lee et al. reported the differentiation of PDLSCs into endoderm-lineage cells; three-dimensional cell culture allowed human PDLSCs to differentiate into pancreatic cell-like cells that expressed pancreas-related genes and secreted insulin in response to high concentrations of glucose [37]. These results suggested that PDL would contain extremely immature stem cell population that could differentiate not only into mesenchymal-lineage cells but also into other lineage cells. Therefore, PDLSCs would be an attractive cell source for regenerative medicine in various tissues.
Immunomodulatory Effects of PDLSCs
Immunomodulatory functions of somatic stem cells have been reported in many different cell types, including dental tissues such as PDL, dental pulp, root apical papilla, and gingiva [38,39]. Their immunomodulatory properties are not constitutive and are affected by surrounding microenvironment; quiescent stem cells revealed low immunomodulation activities; however, inflammatory cytokines markedly promoted their immunomodulatory functions [40]. Activated immune cells mainly produce inflammatory cytokines; therefore, immune cells and somatic stem cells reciprocally control each other and this interaction is considered to be of importance for tissue homeostasis as well as in the processes of inflammation and repair.
Periodontitis is a highly prevalent chronic inflammatory disease that results in the destruction of tooth-supporting tissues. Alveolar bone loss is a common characteristic associated with periodontitis, which is induced by host innate and adaptive immune response to various biofilm-associated multiple bacteria [41]; bacterial accumulation elicits migration and activation of polymorphonuclear neutrophils and monocytes that secrete various cytokines such as ILs, tumor necrosis factor-α, osteoclast differentiation factor, and macrophage colony-stimulating factor. In response to these cytokines, hematopoietic precursor cells differentiate into osteoclast precursor cells, which finally fuse into mature multicellular osteoclasts that are related to alveolar bone resorption.
Peripheral blood mononuclear cells (PBMNCs) originate from hematopoietic precursor cells and contain osteoclast precursor cells that have the potential to generate osteoclasts. These cells are also critical components of the innate and adaptive immune system. Intriguingly, PDLSCs were reported to suppress proliferation of PBMNCs (Fig. 2); activated PBMNCs are known to secrete interferon-gamma (IFN-γ) and human PDLSCs treated with IFN-γ inhibited PBMNC growth through the upregulation of indolamine 2,3-dioxygenase-1 (IDO-1), a molecule that shows an immunosuppressive effect [42]. This result suggested that the immunomodulatory abilities of PDLSCs for PBMNCs would depend on the IFN-γ signaling pathway.
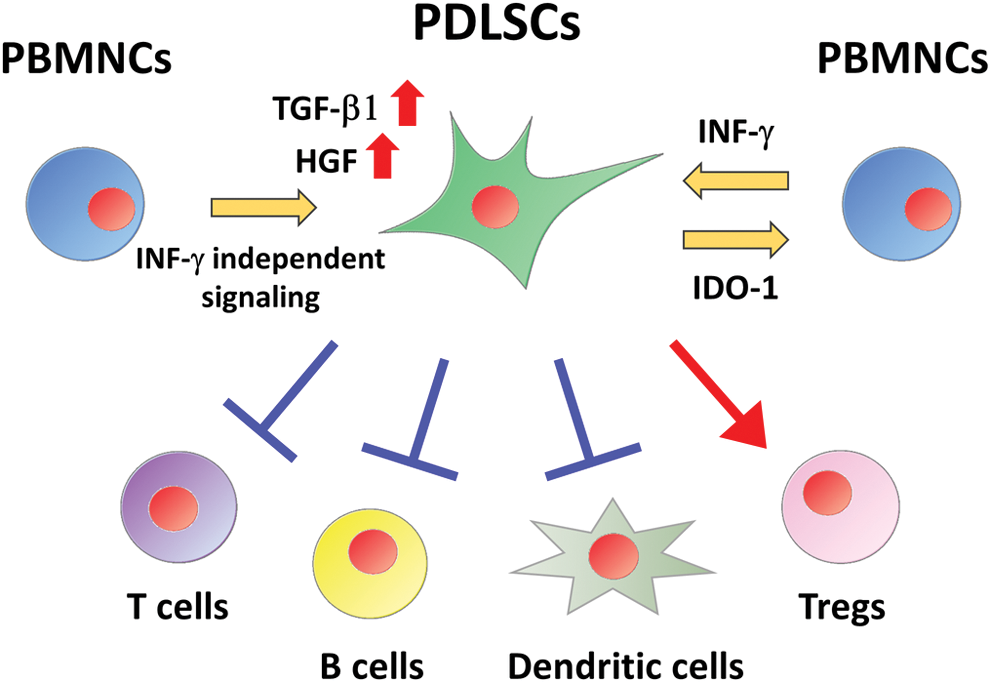
Immunomodulatory effects of PDLSCs. Activated PBMNCs secrete IFN-γ and PDLSCs exposed to IFN-γ inhibit PBMNC proliferation through the upregulation of IDO-1. PDLSCs co-cultured with PBMNCs increase the expression of TGF-β and HGF through IFN-γ independent signaling pathway. PDLSCs suppress activation of T cells, B cells, and dendritic cells; however, they promote generation of Tregs. HGF, hepatocyte growth factor; IDO-1, indolamine 2,3-dioxygenase-1; IFN-γ, interferon-gamma; PBMNCs, peripheral blood mononuclear cells; TGF-β, transforming growth factor-β; Tregs, regulatory T cells. Color images are available online.
Toll-like receptor (TLR)2 and IL-12 were demonstrated to enhance IFN-γ-dependent immunomodulatory properties; TLR2 agonist-treated human PDLSCs promoted IFN-γ-induced IDO-1 gene and protein expression [43], and IL-12-treated human PDLSCs significantly increased the expression of IFN-γ, as well as IDO-1 and human leukocyte antigen G [44].
In addition, transforming growth factor-β1 (TGF-β1) and hepatocyte growth factor, potential mediators of immunomodulatory effects in MSCs, were upregulated in human PDLSCs co-cultured with PBMNCs despite the presence of an IFN-γ neutralizing antibody [42]. TLR4 agonist could not enhance IFN-γ-related IDO-1 expression in human PDLSCs; however, it induced the upregulation of proinflammatory mediators such as IL-6, IL-8, and monocyte chemotactic protein-1 [43]. ERK inhibitor significantly attenuated lipopolysaccharide-induced inhibitory effect of human PDLSCs on PBMNC migration [45]. These results suggested the involvement of IFN-γ-independent signaling pathway in immunomodulatory functions of PDLSCs on PBMNCs.
PBMNCs consist of various lymphocytes, including T cells, dendritic cells, and B cells. Recent studies demonstrated the immunomodulatory effects of PDLSCs on each cell type of PBMNCs (Fig. 2); allogeneic transplantation of human PDLSCs suppressed proliferation of T cells in a prostaglandin E2 (PGE2)-dependent manner in periodontitis swine models [46]. In addition, human PDLSCs significantly reduced the expression of nonclassical major histocompatibility complex glycoprotein, CD1b, on mature dendritic cells, resulting in decreased proliferation of T cells [47]. Human PDLSCs also inhibited activation of B cells through programmed death-1 and its ligand interaction in cell-to-cell contact-dependent manner [48].
On the other hand, PBMNCs treated with human PDLSCs promoted the generation of CD4+ CD25+ Foxp3+ regulatory T cells (Tregs), although Tregs were a component of PBMNCs [49]. Tregs were critical for maintaining self-tolerance and immune homeostasis, and intriguingly, they were reported to be involved in the regulation of immune response in periodontitis [50]; inhibition of Treg function promoted the production of inflammatory cytokines and increased the number of leukocytes and bone loss, suggesting that enhancing Treg activation would inhibit the progression of periodontitis.
Previous studies demonstrated that human BMMSC-derived IDO-1, TGF-β1, and PGE2 induced their activation and maturation [51,52]. PDLSCs have been known to express these soluble factors, suggesting that PDLSC-derived several soluble factors would play important roles in protecting PDL from periodontitis by the activation of Tregs. Based on these results, it is indicated that the immunomodulatory functions of PDLSCs may provide a novel therapeutic approach not only for periodontitis but also for immune-related disorders.
Stem Cell-Related Marker Expression of PDLSCs
Human PDLSCs were first identified using two early MSC-related cell surface molecules, STRO-1 and CD146/MUC18 [13] (Table 1). We also used other cell surface molecules associated with stromal cells and endothelial cells, including CD44, CD90, CD105, CD166, and Stro-3 to identify human PDLSCs [42]. Trubiani et al. also exhibited that human PDLSCs highly expressed MSC-related markers such as CD10, CD26, CD29, CD73, and CD349/FZD9 [53]. Iwasaki et al. demonstrated that CD44, CD73, and CD90 were detected not only in human PDLSCs but also in human BMMSCs and dermal fibroblasts; however, CD146 was strongly expressed only in human PDLSCs [54], indicating that CD146 may be an important cell surface molecule to identify human PDLSCs.
Mesenchymal Stem Cell-, Neural Crest Cell- and Embryonic Stem Cell-Related Marker Expression in Periodontal Ligament Stem Cells
Neural crest cells are a transient multipotent stem cell population that initially emerges at the junction of the epidermal and neural ectoderm, and migrates into their destination in the embryo and generates various types of cells [35].
Huang et al. reported the high expression of neural crest markers, including Nestin, Slug, p75NTR, and SOX10 in human PDLSCs, and suggested the presence of neural crest cell-like cells (NCLCs) in human PDL tissue (Table 1) [55]. Kaku et al. investigated the localization of NCLCs in PDL tissue using WNT1-CRE/ZEG mice [56]. They demonstrated that mouse NCLCs located in the middle of PDL tissue and constituted 1.4% of all PDL cells. Consistent with these results, recent studies reported the isolation of NCLCs expressing p75NTR, HNK-1, SOX10, and Nestin from human PDL cells by selecting connexin-43-expressing cells [57] and culturing in low serum conditions [58]. We also developed a multipotent clonal human PDL cell line that highly expressed Slug, SOX10, Nestin, p75NTR, and CD49d [59].
Surprisingly, NCLCs isolated from PDL cells did not express MSC-related cell surface markers [56], indicating that NCLCs in PDL tissue would be distinguished from PDLSCs that express MSC-related markers.
In addition, strong expression of OCT4 and NANOG, the core components of a complex network of transcription factors to maintain the pluripotent state in ES cells, was also observed in human PDLSCs (Table 1) [53]. Consistent with this result, Kawanabe et al. identified ES cell-related antigens, including SSEA-1, SSEA-3, SSEA-4, TRA-1–60, TRA-1–81, ALP, SOX2, and REX1, in human PDLSCs [60]. Interestingly, human PDLSCs strongly expressed NANOG, SOX2, OCT4A, and OCT4B compared with human MSCs derived from peripheral blood and umbilical cord [61], and the expression level of SSEA-4 was significantly higher in human PDLSCs than that in human BMMSCs and DPSCs [15].
These results indicated that the majority of PDLSCs show MSC-like properties and some cells may have the characteristics of more immature cells such as neural crest cells and ES cells. However, most studies about stem cell-related marker expression of PDLSCs were performed in vitro and their marker expression was assessed without stem cell niche. Stem cells are thought to maintain undifferentiated state when they locate in stem cell niche; therefore, further in vivo studies should be essential to evaluate of MSC-, neural crest-, and ES cell-related marker expression in PDLSCs.
Involvement of Semaphorin 3A in Stemness Regulation of PDLSCs
Semaphorin 3A (Sema3A), a member of the semaphoring family secreted protein, was reported to perform important roles in the development of various tissues such as blood vessels, peripheral nerves, and skeletal tissues [62]. Moreover, it functions as a potent osteoprotective factor through the promotion of bone formation and the inhibition of bone resorption [63].
Our previous study demonstrated Sema3A and its receptor expression in mouse PDL tissue and human PDL cells [64]. Interestingly, a multipotent human PDL cell line highly expressed Sema3A compared with a low-potent human PDL cell line, indicating the importance of Sema3A in regulating the stem cell features of PDLSCs. Then, we developed a Sema3A-transfected low-potent human PDL cell line (2-52sema3a) and an empty vector-transfected low-potent human PDL cell line (2-52empty). The expression level of MSC- and ES cell-related markers was upregulated in 2-52sema3a compared with that in 2-52empty (Fig. 3). After the culture in osteogenic and adipogenic induction media, the formation of mineralized nodules and lipid droplets, and the increase of bone- and adipose-related marker gene expression were confirmed in 2-52sema3a, but not in 2-52empty (Fig. 3).
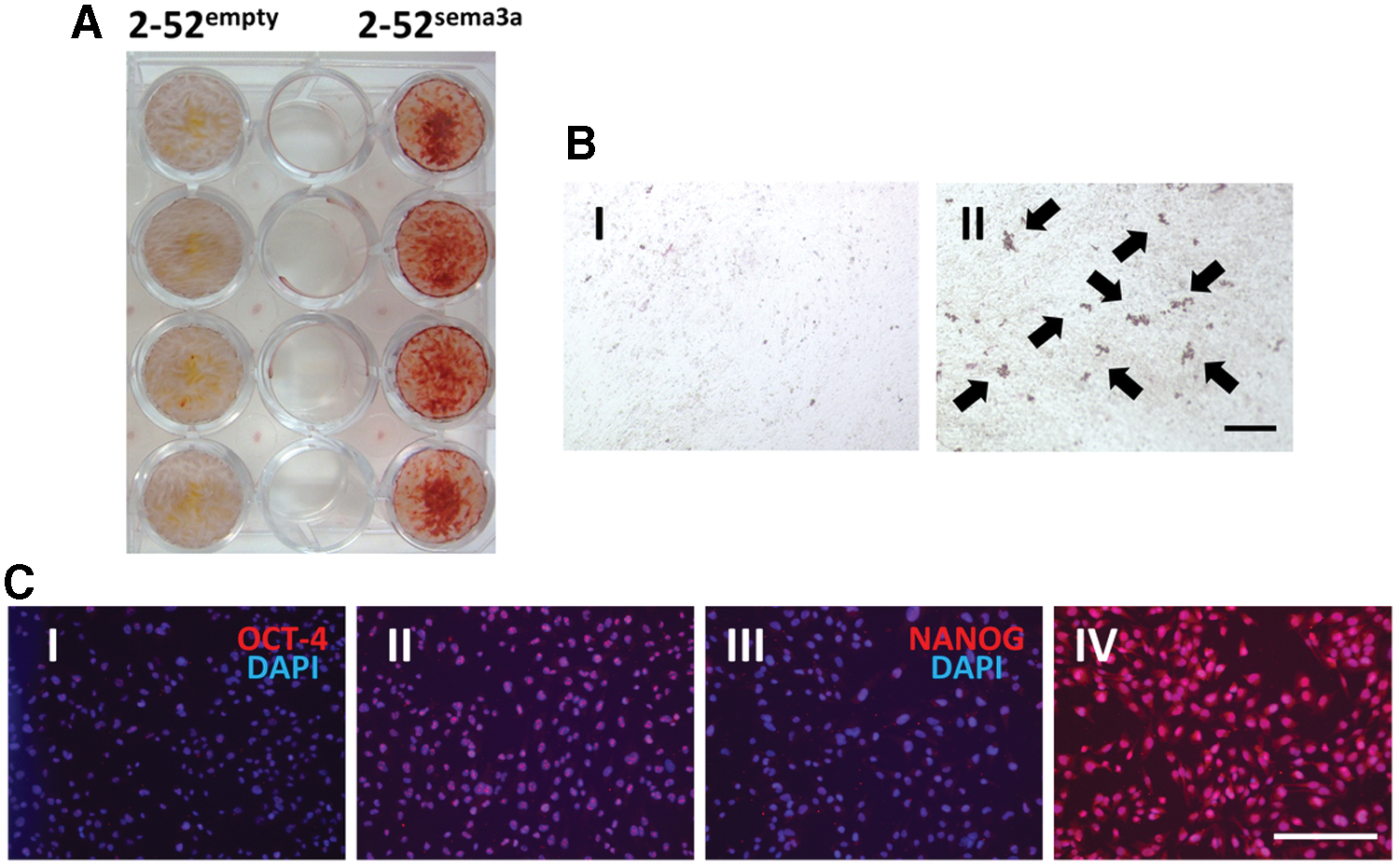
Effects of Semaphorin 3A on osteoblastic and adipogenic differentiation and embryonic stem cell-related protein expression in a low-potent human PDL cell line.
These findings suggested that the induction of Sema3A gene provided multipotency for a low-potent human PDL cell line. This result is important because Sema3A reprogrammed PDL cells have the potential to become an easily available cell source with PDLSC-like properties. Yamada et al. also revealed that Sema3A knockdown significantly decreased stem cell marker expression and completely abolished tumorigenicity, sphere formation, and self-renewal ability in mouse lung cancer stem cells [65]. Our recent results also demonstrated that Sema3A treatment promoted chemotaxis, proliferation, and odontoblastic differentiation of human DPSCs [66]. The result of these studies strongly supported our hypothesis that Sema3A would play crucial roles in maintaining stemness of PDLSCs.
At the present time, we are planning to confirm the safety of Sema3A reprogrammed PDL cells and evaluate their potential to regenerate PDL tissues in vivo.
Regeneration of Periodontium Using PDLSCs
Tissue engineering is the field of life science to promote regeneration of damaged or diseased tissues and organs using stem cells, biological signals, and cell-seeded scaffold. Therefore, a number of researchers have tied to regenerate periodontium based on the tissue engineering technology.
Seo et al. first transplanted human PDLSCs (4 × 106 cells/carrier) with a hydroxyapatite (HA) scaffold into artificially formed periodontal defects in the mandibular molars of immunodeficient mice [13]. After 8 weeks, they confirmed the reconstitution of cementum- and PDL-like structures in the transplant sites, suggesting the significance of PDLSCs to accomplish the regeneration of periodontium.
Ninomiya et al. also performed allogeneic transplantation of rat PDLSCs or differentiated PDL cells (5 × 105 cells/disk) with HA disks under the fascia of the dorsal muscles of wild-type Lewis rats [67]. At 12 weeks, new bone-like tissues were formed around HA disks with rat PDLSCs, whereas no mineralized tissue formation was observed around HA disks containing differentiated rat PDL cells. Immunohistochemical staining revealed that the matrices generated in the HA disks with rat PDLSCs were positive for bone-related marker proteins, including ALP, OPN, BSP, and OCN.
In addition, Park et al. formed a complex using human PDLSCs (6 × 106 cells/carrier) and HA and β-tricalcium phosphate (HA/β-TCP) scaffold, the most common calcium phosphates clinically applied for replacing and augmenting bone tissue, and transplanted it into the dorsal region of immunodeficient mice [68]. The complex successfully generated PDL-, bone-, and cementum-like tissues after 8 weeks of transplantation. Surprisingly, Sharpey's fiber-like tissue formation was also identified between PDL and cementum under similar conditions, indicating the complete generation of PDL tissues.
Furcation defect is defined as a loss of attachment and bone in the inter-radicular area of multirooted teeth and represents one of the major complications of periodontal disease. Several studies used a fenestration defect model to assess the regenerative potential of PDLSCs.
A previous study performed autologous transplantation of PDL cells (2 × 105 cells) in blood coagulum into surgically created fenestration defects on the maxillary canine and first molar teeth at a spacing of 5–5 mm in dogs [69]. After 42 days, the amount of new bone formation was greater in the cell-seeding group (84%) compared with the control group (39%); however, the amount of new cementum formation was limited (9%–12%).
Iwasaki et al. transplanted human PDLSCs with a decellularized amniotic membrane into surgically created furcation defects with a buccopalatal depth of 2 mm and a horizontal depth of 1.5 mm at the media furcation in nude mice [70]. After 4 weeks of transplantation, the defect was filled mainly with fibroblastic cells, collagen fibers, and blood vessels in the control group, while the formation of new bone (86.43% ± 7.50%) and cementum (3.441 ± 0.485 μm) was confirmed at the defect in the PDLSC transplantation group. These results suggested that the new periodontal tissue formation was enhanced not only in subcutaneous tissues but also in the defects of periodontium after transplantation of PDLSCs.
On the other hand, Nunez et al. revealed no significant difference in the amount of periodontal regeneration at the furcation area and new cementum formation between PDLSC transplantation group and control group [71]. They transplanted a bone grafting material into critical-size (6 mm) supra-alveolar periodontal defects with or without canine PDLSCs (1.4 × 106 cells/carrier) and performed histological evaluation after 3 months postsurgery. Importantly, they did not use a barrier membrane for the surgery and the defects that they surgically created were not limited to the furcation area, indicating that stabilization of grafted PDLSCs during healing and the balance between the defect size and grafted PDLSC number may be important for PDLSC-based therapy for damaged periodontium.
It is of importance for the stimulation of PDLCs to differentiate into the cells that constitute periodontium in the regeneration processes. The treatment with growth factors and morphogens is known to be efficient for stimulating PDLSCs. Yu et al. cultured human PDLSCs with or without insulin-like growth factor 1 (IGF-1) and revealed that IGF-1 promoted their proliferation and osteoblastic differentiation in a dose-dependent manner [72]. They also transplanted human PDLSCs (1 × 106 cells/carrier) treated with or without IGF-1 under renal capsules of immunocompromised mice. At 21 days, human PDLSCs treated with IGF-1 generated more mineralized bone-like tissues and showed higher expression of bone-related marker proteins such as RUNX2, Osterix, and OCN than untreated cells.
Jung et al. developed human PDLSCs genetically modified to express the gene encoding BMP2 and subcutaneously transplanted them (1 × 106 cells/carrier) with HA/β-TCP scaffold into the dorsal area of immunocompromised mice [73]. After 8 weeks, the BMP2-overexpressing human PDLSCs generated significantly more mineralized tissues highly expressing bone- and cementum-related markers- including ALP, OCN, BMP2, and CEMP1, than untreated cells.
Jin et al. treated human PDLSCs (1 × 107 cells/carrier) treated with or without plasminogen activator inhibitor-1 (PAI-1) and mixed with HA/β-TCP and human tooth root dentin matrix [74]. Then, they subcutaneously transplanted the complexes into the dorsal region of immunocompromised mice. At 8 weeks after transplantation, PAI-1-treated human PDLSCs revealed greater amount of new cementum formation on the surface of HA/β-TCP compared with untreated cells. In addition, newly formed cementum area in the PAI-1-treated group was 2.13-fold larger than that in the control group.
Lim et al. exposed human PDLSCs (1 × 106 cells/carrier) to harmine, a kind of β-carboline alkaloid, and subcutaneously transplanted them with a biphasic calcium phosphate scaffold into immunocompromised mice [75]. After 8 weeks, harmine-treated human PDLSCs showed greater mineralization than untreated cell. Interestingly, they also promoted the formation of Sharpey's fiber-like collagen that perpendicularly inserted into mineralized tissues.
Along with growth factors and morphogens, cell sheet transplantation shows promise in engineering three-dimensional biological tissues. PDL cell sheets (3 × 104 cells/dish) developed using temperature-responsive culture dishes were transplanted with β-tricalcium phosphate into three-wall infrabony defects on the mesial side of bilateral mandibular first molars in dogs [76]. After a 6-week period of healing, the cell sheet group revealed significantly higher bone mineralization (76.95% ± 5.08%), cementum regeneration (78.22% ± 5.50%), and periodontal score (4.63 ± 0.25) than the negative control group.
In addition, several studies tried to regenerate PDL tissues using PDLSC sheets with several scaffolds and growth factors. Gao et al. generated cell sheets using human PDLSCs (1 × 106 cells/well) treated with osthole, a coumarin-like derivative extracted from Chinese herbs, and subcutaneously transplanted them with HA/β-TCP scaffolds into the dorsal region of nude mice [77]. After 4 weeks of postsurgery, the osthole-treated group formed larger amount of mineralized tissues on the surface of HA/β-TCP scaffolds than the untreated control group.
Wang et al. developed three types of cell sheets (5 × 104 cells/well) that were derived from human PDLSCs, jaw bone MSCs (JBMSCs), and the combination of PDLSCs and JBMSCs (PDLSCs + JBMSCs) [20]. They were treated with platelet-rich fibrin and mixed with HA/β-TCP and human-treated dentin matrix, and subcutaneously transplanted into the dorsal region of nude mice. Intriguingly, three cell sheets produced very different structures after 8 weeks of transplantation; PDLSC sheets tended to generate fiber tissues, while JBMSC sheets were prone to form mineralized tissues. Moreover, PDLSC + JBMSC sheets exhibited a balance between newly formed PDL- and bone-like tissues.
Based on these results, several clinical trials that used autologous human PDLSCs have been performed. Feng et al. reported autologous human PDLSC transplants into the intrabony defects of deep periodontal pockets (from 4.8 to 10 mm) of three male patients [78]; human PDLSCs isolated form third molars of each patient were mixed with a bone grafting material (4 × 106 cells/carrier). Following a mucoperiosteal flap reflection and a removal of inflammatory periodontal tissues, a PDLSC-grafting material complex was inserted into periodontal defect areas. After 72 months postsurgery, the surgical reentry to the implanted area and X-ray analysis of two patients exhibited definitive regeneration of periodontium. Furthermore, at 3, 6, and 42 months, all three patients showed the significant decrease of tooth movement, probing depth (PD), and clinical attachment level (CAL).
Shalini et al. compared 28 periodontitis patients treated with open flap debridement (OFD) or OFD followed by autologous human PDLSCs and their niche transplants [79]; PDLSCs along with their niche were gently removed from extracted wisdom teeth and immediately mixed with gelatin scaffolds. After OFD, the mixture was placed in the periodontal defects. At 3, 6, 9, and 12 months of transplantation, OFD with human PDLSC treatment revealed a reduction in PD, CAL, and defect area, and an increase in bone density in defect area compared with OFD treatment.
Iwata et al. formed autologous human PDLSC sheets in vitro and transplanted them to the 10 periodontitis patients with PDs of more than 4 mm [80]; human PDLSCs derived from wisdom teeth were cultured on temperature-responsive culture dishes to generate cell sheets. After 2 weeks of culture, the generated cell sheets (from 0.55 to 2.00 × 106 cells/sheet) were autologously transplanted with a β-TCP scaffold and biodegradable polyglycolic acid mesh into the periodontal defects. At 6 months postsurgery, all cases showed a reduction of PD and CAL, and an increase of bone height; the mean CAL gains were 2.5 ± 2.6 mm, the mean PPD reductions were 3.2 ± 1.9 mm, and the mean gain of computed tomography image-based linear bone height was 2.3 ± 1.8 mm.
These studies suggested that using autologous human PDLSCs to treat periodontal defects was safe and effective; however, multicenter and randomized controlled studies with an increased sample size should be essential to examine the efficacy of human PDLSC-based periodontal therapy.
Regeneration of Organs Using PDLSCs
PDLSCs have been reported to show the potential for regeneration of not only periodontium but also other organs. Human PDLSCs (1 × 106 cells/carrier) mixed with a nanohydroxyapatite-coated genipin-chitosan conjugation scaffold (HGCCS) or genipin-chitosan framework (GCF) were transplanted into surgically created full-thickness defects (5 mm in diameter) in the parietal bones in adult male Sprague-Dawley rats [81]. After 12 weeks of transplantation, both human PDLSCs with HGCCS group and human PDLSCs with GCF group promoted the formation of a thick layer composed of fibrous connective tissue and newly formed bone. Interestingly, human PDLSCs with HGCCS group showed higher volume of new bone with infiltrated blood vessels than human PDLSCs with HGCCS group.
After the transplantation into tendon defects of nude mice, human PDLSC (4 × 106 cells)-encapsulated RGD-coupled alginate microspheres loaded with TGF-β3 could differentiate into tenocytes that expressed tendon markers, including Tenomodulin, Eya1, Eya2, and Scleraxis [82]. They also induced tendon regeneration in vivo, and surprisingly, the amount of collagen fibers with a yellow color was greater in the human PDLSC group than in the BMMSC and gingival MSC groups.
Cell suspension of human PDLSCs (1 × 105 cells/μL) was injected into the brain of immunocompromised mice [83]. After 4 weeks postinjection, human PDLSCs were detected in dorsal lamina of the dentate gyrus, the dorsal cortex and CA1, the corpus callosum, fimbria, neighboring epithelium of lateral ventricles, and the caudal pole of subventricular zone. In addition, these cells highly expressed neural (Dcx, NF-M, and GABA) and glial (GFAP and NG2) markers, suggesting that transplanted human PDLSCs could integrate, migrate, and differentiate into mature neural cells in the brain.
Human PDLSCs (5 × 104 cells) with saline were also injected into the vitreous chamber of adult F344 rats after optic nerve crush [84]. At 3 weeks, the majority of injected cells maintained in the vitreous chamber and localized in the nerve fiber layer and the retinal ganglion cell layer; however, some cells migrated into the host retina. Moreover, grafted PDLSCs enhanced survival of retinal ganglion cells and induced generation of new axons located in the damaged area. The findings from these studies suggested that PDLSCs would perform crucial roles in the homeostasis and regeneration not only in PDL tissues but also in other various tissues.
Importantly, two different mechanisms have been suggested in PDLSC-mediated regeneration of a variety of tissues. First is the direct differentiation of transplanted PDLSCs into the mature somatic cells. Many studies demonstrated the survival of subcutaneously transplanted PDLSCs into the dorsal region of mice and the formation of bone-, cementum-, and Sharpy's fiber-like tissues in the proximity of the transplanted site [13,21,28,70,73 –75,77]. Menicanin et al. also confirmed human PDLSCs within the newly formed mineralized tissues, suggesting their differentiation into mature osteocytes [28]. In addition, human PDLSCs transplanted into tendon and brain could engraft and strongly express tenocyte- and neural cell-related markers, respectively [82,83]. These results strongly support that grafted PDLSCs can differentiate into functional specialized cells and contribute to regenerate damaged tissues.
The second is the indirect effects through PDLSC-derived paracrine factors. PDLSCs have been known to produce various signaling molecules, including growth factors, cytokines, and extracellular matrices. Nagata et al. injected conditioned medium collected from human PDLSCs or dermal fibroblasts into surgically created periodontal defects in Sprague-Dawley rats [85]. At 4 weeks after injection, the PDLSC group revealed more new mineralized tissues with thick collagen bundles in the defects than the dermal fibroblast group.
Our previous report also suggested the neuroprotective roles in human PDLSCs in a paracrine manner [59] (Fig. 4). A multipotent human PDL cell line (1–17 cells) promoted neural differentiation and migration of neural progenitors. Intriguingly, a nerve growth factor (NGF) neutralization antibody and a specific inhibitor of NGF receptor tyrosine kinase activity (K252a) significantly inhibited these effects of 1–17 cells, suggesting that 1–17 cell-derived NGF could bind its receptor of neural progenitors and induced their neural differentiation and migration. Moreover, conditioned medium form 1–17 cells (1–17CM) protected apoptotic cell death of neural progenitors, and interestingly, neural progenitors treated with 1–17CM and recombinant NGF revealed the same expression level of antiapoptotic genes.
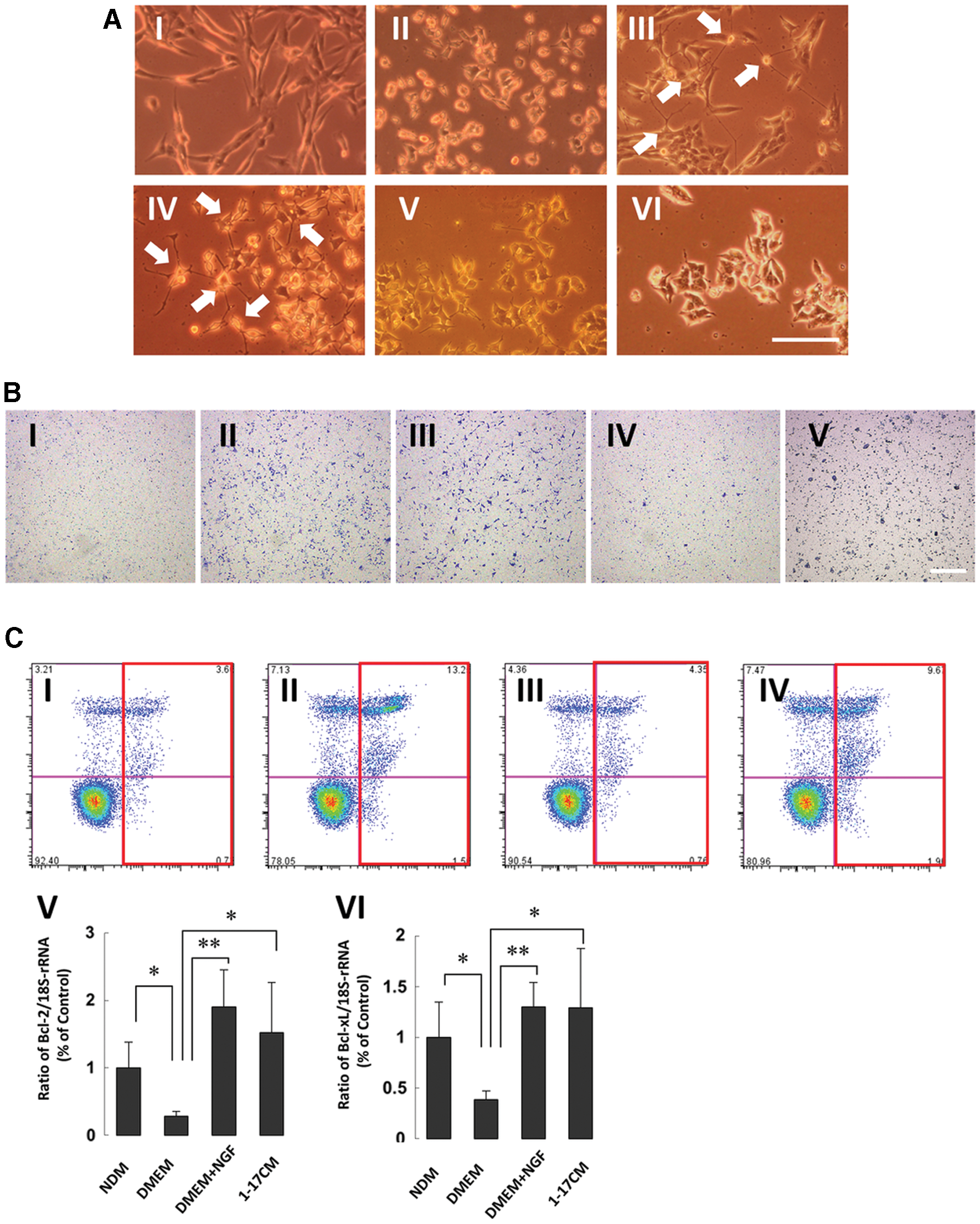
Neuroprotective roles in a clonal human multipotent PDL cell line.
Based on these results, regenerative potential of PDLSCs should be mediated by both direct and indirect mechanisms.
Future Prospective
As reviewed in this article, PDLSCs show self-renewal, multipotency, and immunomodulatory properties; therefore, they should hold great promise for the treatment of destructed periodontium. However, the rarity of human PDLSCs prevents their application for studies of regeneration in periodontium.
Cell reprogramming, a process to convert cells from one particular type to another, holds great promise to overcome this issue. Takahashi et al. first developed induced pluripotent stem cells (iPSCs) from adult dermal fibroblasts by cell reprogramming with four genes, Oct4, Sox2, Klf4, and c-Myc [86]. iPSCs showed ES cell-like properties, including unrestricted proliferative potential and differentiation ability into three primary germ layer-derived cells, suggesting reprogramming somatic cells by gene transfer would provide the huge number of immature cells required for cell therapy.
Recently, several studies succeeded in controlling the differentiation of iPSCs into PDLSCs. Yin et al. demonstrated that human gingival cell-derived iPSCs treated with enamel matrix derivative or growth/differentiation factor-5 strongly expressed bone-, cementum, and PDL-related markers compared with untreated iPSCs [87]. Our recent study also reported the establishment of PDLSCs from iPSCs using the extracellular matrix [88]; following the induction of human skin fibroblast-derived iPSCs to differentiate into NCLCs, these cells were further cultured on the extracellular matrix produced by human PDL cells. They revealed higher differentiation potential into osteoblasts and adipocytes along with stronger expression of MSC- and PDL-related markers compared with NCLCs seeded on fibronectin or skin fibroblast-derived extracellular matrix.
These methods of establishing PDLSCs by applying iPSCs may help to acquire a great amount of PDLSCs from iPSCs to clarify the behavior of PDLSCs during healing process of periodontium. Importantly, transplantation of iPSCs harbors the risk of tumorigenesis; therefore, the development of safe nontumorigenic iPSC-derived PDLSCs should be essential to achieve their clinical application.
Direct cell reprogramming is a kind of cell reprogramming; however, it can convert cells into another without passage through an intermediate or pluripotent state [89]. Previous reports demonstrated the generation of NCLCs and functional osteoblasts from human fibroblasts with direct cell reprogramming with FOXD3 and Oct4, L-Myc, Runx2, and Osterix, respectively [90,91]. These cells may be promising for the research about regeneration of periodontium because PDL tissue is originated form neural crest and consists of various types of cells, including osteoblasts.
On the other hand, cell reprogramming factors that induce mature specialized cells to turn into PDLSCs have not been identified. These factors may enable us to acquire a large amount of PDLSCs more simply, quickly, and efficiently; therefore, further research will probably be necessary to figure them out. Based on recent progress in cell reprogramming technology, PDLSCs generated by the application of iPSCs and direct cell reprogramming may be used as a valuable and unlimited cell source for tissue engineering study in periodontium.
Footnotes
Acknowledgment
This project was supported by the Grant-in-Aid for Scientific Research (grant nos. JP 17H04385, JP17H01598, and JP18K19651) from the Japan Society for the Promotion of Science, Japan.
Author Disclosure Statement
No competing financial interests exist.