
Abstract
Hepatic stellate cells (HSCs) are liver-specific cells playing critical roles in liver physiological and pathophysiological processes. Transforming growth factor-β1 (TGF-β1) is an inflammatory cytokine secreted by both hepatocytes and HSCs. We have previously shown that microvesicles (MVs) derived from quiescent HSCs protect hepatocyte functions. In this study, we investigated the effects of MVs released from TGF-β1-stimulated HSCs (HSC-MVs) on xenobiotic-injured hepatocytes. Two hepatocyte cell lines (BRL-3A and HL-7702) were treated with N-acetyl-p-aminophenol or H2O2 to build the injury models. Different concentrations of HSC-MVs were used to coculture with injured hepatocytes. MTT, Hochest33258 staining, and flow cytometry were used to determine their effects on the viability and apoptosis of hepatocytes. Liver injury indicators, alanine aminotransferase (ALT) and aspartate amino transferase (AST), were assessed by enzyme-linked immune sorbent assay kits. The phosphoinositide 3-kinase (PI3K) activator (740Y-P) and extracelluar signal regulated kinase (Erk)1/2 activator (platelet-derived growth factor-BB) were used for pathway analysis. The expression levels of p-PI3K/PI3K, p-Akt/Akt, and activated caspase-3 were measured by western blot. Results showed that (i) HSC-MVs dose dependently impaired the viability of hepatocytes in both injury models, (ii) moreover, HSC-MVs dose dependently increased the apoptosis in those cell models, (iii) HSC-MVs also elevated the levels of ALT and AST in the coculture media, and (iv) these effects were accompanied by a decrease in p-PI3K/PI3K and p-Akt/Akt, which could be partially abolished by 740Y-P. Meanwhile, the proapoptotic effect of HSC-MVs was associated with p-Erk1/2/Erk1/2 downregulation and activated caspase-3 upregulation, and could be inhibited by Erk1/2 activation. Our findings demonstrate that HSC-MVs are involved in inflammatory hepatocytes injury probably through the PI3K/Akt, Erk1/2, and caspase-3 pathways.
Introduction
Hepatic stellate cells (HSCs) are major resident nonparenchymal liver-specific mesenchymal stem cells [1]. In a normal liver, HSCs are in the quiescent stage for storing vitamin A [2]. Upon liver injury, HSCs will be activated and transdifferentiated from vitamin A-storing cells into myofibroblast-like cells. Afterward, the activated HSCs produce mediators including tumor necrosis factor-α and interferon-β, which induce liver cell apoptosis [3]. Therefore, HSCs play an important role in the pathogenesis of liver injury.
Currently, extracellular microvesicles (MVs) are believed to convey the effects of parent cells [4]. MVs are lipid membrane vesicles with diameter of 100–1000 nm and are released when cells undergo stress, activation, or apoptosis [5,6]. They are recognized to play a pivotal role in cell-to-cell communication and to serve as a novel player in tissue microenvironment [7,8]. In liver, MVs derived from HSCs or hepatocytes have been shown to be implicated in inflammation that is associated with liver diseases such as liver fibrosis, cholangiopathies, and hepatobiliary malignancies [9,10].
The functions of MVs are highly correlated with the status of their mother cells. We have previously shown that MVs derived from starving stress or apoptotic stress-stimulated endothelial progenitor cells exerted different effects on endothelial cell functions [11]. Acetaminophen [N-acetyl-p-aminophenol (APAP)] and H2O2 are well-defined xenobiotics that have been widely used to build hepatocytes injury models [12,13]. Our recent study has demonstrated that MVs derived from HSCs under normal environment can protect APAP/H2O2-injured hepatocytes [12]. Of note, the liver environment elements including cytokine paracrine, reactive oxygen species, and MVs could be altered upon injury. MVs released from different status of cells may vary in functions [11,14]. Nevertheless, the effects of MVs released from activated HSCs on hepatocytes remain unknown.
Various cytokines, such as transforming growth factor (TGF)-β1 and platelet-derived growth factor (PDGF), play an important role in HSCs activation [15]. As an inflammatory and profibrotic cytokine secreted by both hepatocytes and HSCs, TGF-β1 has been shown to have regulatory roles in cell viability, differentiation, and immune function [16,17]. TGF-β1 released from injured hepatocytes contributes to HSCs activation [18,19], whereas TGF-β1 derived from activated HSCs could induce apoptosis of hepatocytes [20]. However, whether the MVs of TGF-β1-activated HSCs could modulate the function of hepatocytes remains an open question. Understanding this question could help to develop therapeutic approaches for liver injury.
Phosphoinositide 3-kinase (PI3K)/Akt and extracelluar signal regulated kinase (Erk) 1/2 pathways are known to participate in various cellular processes including cell proliferation, migration, and apoptosis [11,21]. Indeed, recent studies have shown that the PI3K/Akt pathway attenuates APAP-induced hepatotoxicity in vivo and in vitro [22]. Erk1/2 pathway has also been shown to be disrupted during hepatotoxicity [23]. Erk1/2 activation exerts protecting effect on menadione-induced apoptosis of hepatocytes [24]. Caspase-3 is an important regulator implicated in apoptosis of hepatocytes [12]. Thus PI3K/Akt and Erk/1/2 pathways and caspase-3 protein are important factors for the regulation of hepatocyte functions.
In this study, we investigated the potential effects of HSCs under TGF-β1 stimulation on the viability and apoptosis of drug-injured hepatocytes. The liver injury indicators, alanine aminotransferase (ALT) and aspartate amino transferase (AST), in the medium of the injured hepatocytes were measured. Signaling pathway proteins including activated Caspase-3, PI3K, phosphorylated PI3K, Akt, and phosphorylated Akt were examined. PI3K activator and Erk1/2 activator were also used to explore the underlying mechanisms.
Materials and Methods
Cell lines and culture conditions
Human HSCs (LX-2) and Rat HSCs (HSC-T6) were obtained from Jennio Biological Technology and used to prepare MVs. The normal human hepatic cells (HL-7702) and rat hepatic cells (BRL-3A) were purchased from Guangdong Joycer Biotech Co. Ltd and used to establish hepatic injury model. All types of cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Hyclone), supplemented with 10% fetal bovine serum (FBS; GIBCO) and 1% penicillin/streptomycin in a 37°C incubator with humidified atmosphere of 5% CO2/95% air.
Preparation and analysis of HSC-MVs
HSC-MVs were generated from human or rat-derived TGF-β1-treated LX-2 and HSC-T6, which were, respectively, defined as LXHSC-MVs and T6HSC-MVs. In brief, LX-2 and HSC-T6 cells were cultured in 100-mm cell culture dishes. When cells grown to 80% confluence, LX-2 and HSC-T6 cultures were washed with phosphate-buffered saline (PBS) and replaced with fresh culture medium containing 10 ng/mL TGF-β1 [25]. After 24 h incubation, the culture medium was collected and centrifuged at 300 g, for 15 min, and then centrifuged at 2,000 g, for 30 min, to remove cells and cell debris. Then the cell-free culture medium was centrifuged at 20,000 g, for 2 h, to pellet HSC-MVs. The pellet of MVs was resuspended with 20 nm-filtered PBS (Whatman, Pittsburgh, PA).
The size and amount of HSC-MVs were determined by nanoparticle tracking analysis (NTA; Malvern, Britain). In addition, MVs-specific marker Annexin V expression was measured by western blot analysis. The MVs were defined as Annexin V positive vesicles with the size of 100–1000 nm.
Injury models of hepatocytes
The injury models of hepatocytes were established as we previously described [12]. In brief, after confluence, HL-7702 cells were treated with 8 mM APAP or 600 mM H2O2. As the injury models in another cell line of hepatocytes, BRL-3A cells were treated with 6.4 mM APAP or 480 mM H2O2.
Coculture assay of HSC-MVs with hepatocytes
To study the incorporation of HSC-MVs with hepatocytes, HSC-MVs were labeled with a red fluorescence dye PKH26, resuspended with DMEM and added to the culture medium as we previously described [26]. In brief, HSC-MVs were labeled with PKH26 (2 × 10−6 M; Sigma-Aldrich) for 5 min at 37°C, followed by wash with 1 × PBS and ultracentrifuged at 20,000 g for 90 min. The concentration of HSC-MVs for coculture experiments was 2 × 107/mL. Cells treated with HSC-MVs free medium that have received the same treatment as HSC-MVs solution were set as control. After 24 h of coculture, the incorporation of HSC-MVs into hepatocytes was observed under a fluorescence microscope (TCS SP5II; Leica, Germany). Hepatocytes were then used for various analyses as already described.
MTT assay
Cell viability of hepatocytes was measured by MTT (3-[4,5-dimethylthiazyol-2yl]-2,5-diphen-yltetrazolium bromide, 5 mg/mL; Sigma) assay [27]. In brief, HL-7702 and BRL-3A were seeded into a 96-well plate at a density of 2 × 103/well with 100 μL DMEM (supplemented with 10% FBS). To determine the effect of LXHSC-MVs on injured hepatocytes viability, HL-7702 cells were cocultured with different concentrations of LXHSC-MVs (2 × 106/mL, 2 × 107/mL, 2 × 108/mL, respectively) when APAP/H2O2 was administered, and BRL-3A cells were cocultured with different concentrations of T6HSC-MVs (2 × 106/mL, 2 × 107/mL, 2 × 108/mL, respectively) when APAP/H2O2 was added. PBS-treated cells were used as normal control groups. Treatment and control groups were set as hepatocytes treated with APAP or H2O2 without HSC-MVs incubation.
After incubation, MTT solution (20 μL) was added and incubated with cells for 4 h at 37°C, then 150 μL dimethyl sulfoxide was added to each well and incubated for 20 min at 37°C. The optical density value of cells was read at 490 nm in a microplate reader (BioTek). Measurement was carried out at 24 h. Cells in triplicate wells were examined at each time point. Results were calculated from the values obtained in three independent experiments. For pathway analysis, cells were preincubated with 740Y-P (Selleckchem; 20 μg/mL) for 2 h.
Hochest 33258 staining assay
Cell apoptosis was tested by Hochest 33258 staining as previously described [27]. In brief, HL-7702 and BRL-3A cells were seeded at 2 × 105/well in a six-well plate and cultured in 2 mL DMEM (supplemented with 10% FBS). After treatment with APAP/H2O2 and HSC-MVs for 24 h, cells were fixed in 4% paraformaldehyde, washed with PBS, and stained with Hochest 33258 staining solution according to the manufacturer's instructions (Beyotime) and observed under a fluorescence microscope. Five fields were randomly selected from the staining of each well for each group, and the average number of positive cells and total cells per field were determined. The apoptosis rate of cells was defined as the ratio of positive cells versus total cells. For pathway analysis, cells were preincubated with 740Y-P (Selleckchem; 20 μg/mL) or PDGF-BB (R&D; 25 ng/mL) for 2 h.
Annexin V-PE/7-AAD staining and flow cytometry analysis
HL-7702 and BRL-3A cells were treated as already mentioned. The apoptosis assay was carried out by using an annexin V-PE/7-AAD apoptosis detection kit (BD Biosciences) as we previously described [21]. In brief, cells were washed with PBS, resuspended with 100 μL 1× annexin-binding buffer, incubated with 5 μL PE-conjugated annexin V and 5 μL 7-amino-actinomycin (7-AAD) for 15 min in the dark, and then analyzed by flow cytometry. Cells stained with both annexin V-PE and 7-AAD were considered as late apoptotic cells, and cells stained only with annexin V-PE were considered to be early apoptotic cells. Each measurement was analyzed three times, and three plates per measurement were conducted in each group. For pathway analysis, cells were preincubated with 740Y-P (Selleckchem; 20 μg/mL) or PDGF-BB (R&D; 25 ng/mL) for 2 h.
AST and ALT analysis
The levels of AST and ALT in the cell-cultured media were measured by the respective detection assay kits (Nanjing Jiancheng Bioengineering Institute, China) [28]. As we already described, HL-7702 and BRL-3A cells were cultured in six-well plates. After growing to 80% confluence, cells were treated as we already described. After 24 h treatment, culture medium of each group was collected for measurement according to the manufactory instructions. For pathway analysis, cells were preincubated with 740Y-P (Selleckchem; 20 μg/mL) for 2 h.
Western blot
Proteins from cells and HSC-MVs were extracted with lysis buffer [29]. Protein lysates were electrophoresed through sodium dodecyl sulfate-polyacrylamide gel elctrophoresis gel and transferred onto polyvinylidene fluoride membranes. The membranes were blocked for 1 h and incubated with primary antibodies against cleaved caspase-3, PI3K, p-PI3K, Akt, p-AktSer473, Erk1/2, p-Erk1/2, Annexin V, and β-actin (all from CST) at 4°C overnight. After washing three times for 30 min with Tris-buffered saline Tween-20 (TBST), the immunoreactivity was visualized by ECL solution (Amersham, Sweden).
Statistical analysis
Data are all expressed as the mean ± standard error of the mean. Multiple comparisons were performed by two-way analysis of variance. Comparisons for two groups were performed by using a Student's t-test (Graph Pad Prism 5 software). A P < 0.05 was considered statistically significant.
Results
NTA of HSC-MVs
NTA showed that HSC-MVs were in size of 100–400 nm (Fig. 1a, b). There were no significant differences in size and concentration between LXHSC-MVs and T6HSC-MVs (Fig. 1c). The western blot analysis results showed that both MVs were Annexin V positive vesicles (Fig. 1d). These results demonstrated that the HSC-MVs were successfully isolated from HSCs culture medium.

Analysis of HSC-MVs by NTA and western blot.
HSC-MVs merged with hepatocytes after in vitro coincubation
After coincubation of PKH 26-labeled HSC-MVs with hepatocytes for 24 h, we observed granular red fluorescence in the cytoplasm of hepatocytes (Fig. 2). The red fluorescence was not found in control group (Fig. 2). These data suggested that HSC-MVs can effectively merge with hepatocytes.
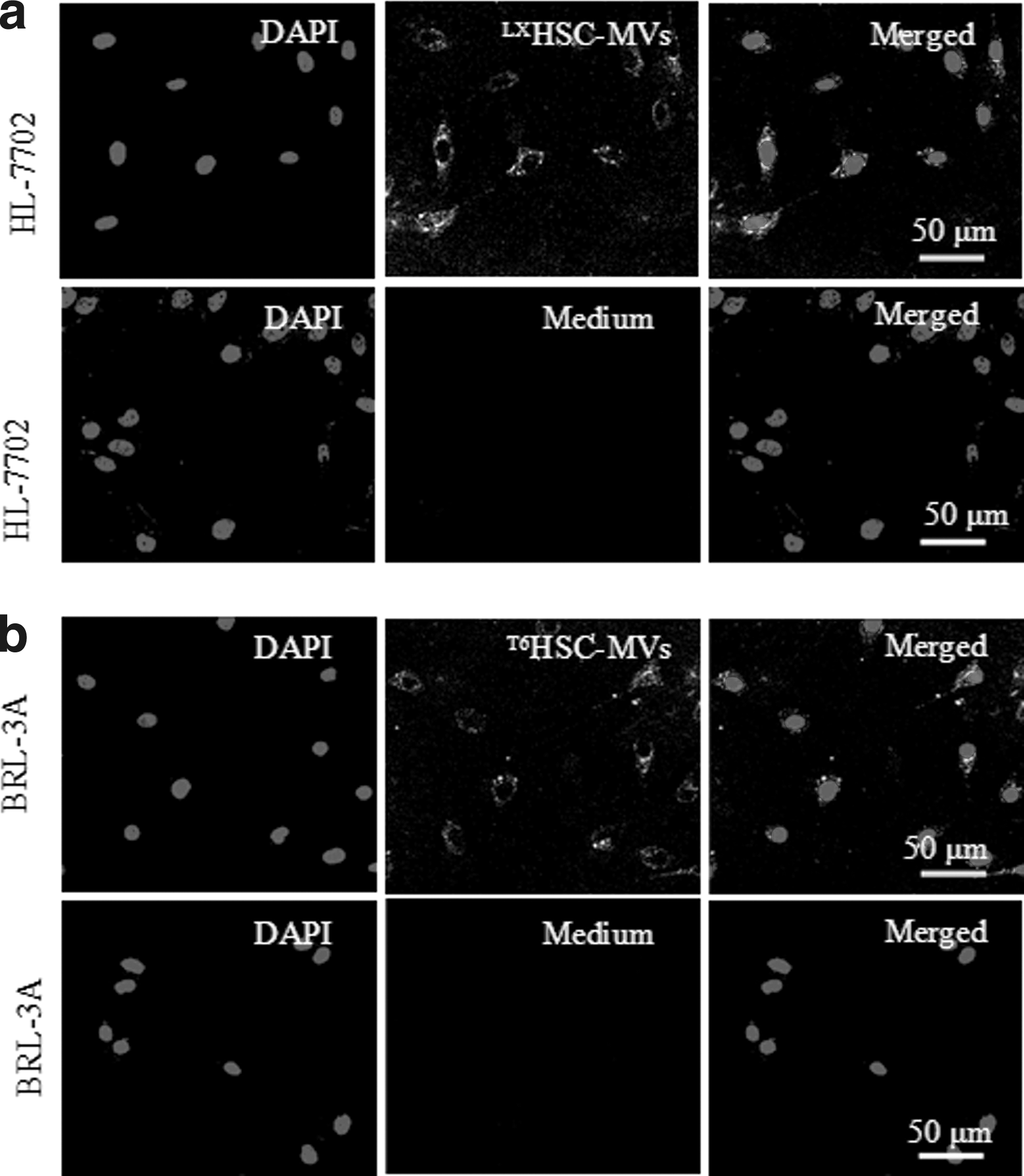
The incorporation of HSC-MVs with hepatocytes after coculture. Representative images showing the merging of PKH26-labeled HSC-MVs with HL-7702 cells
HSC-MVs dose dependently decreased the viability of hepatocytic injury models through inhibiting PI3K/Akt pathway
MTT assay showed that APAP or H2O2 decreased the viability of BRL-3A and HL-7702 hepatocytes (vs. normal control; Fig. 3a, b). HSC-MVs dose dependently aggravated the inhibitory effect of APAP/H2O2 on BRL-3A and HL-7702 viability (vs. treatment control; Fig. 3a, b). The viability of HSC-MVs-treated cells was decreased at a higher dose as compared with those at a lower dose (vs. HSC-MVs at 2 × 107/mL or 2 × 106/mL; Fig. 3a, b). In addition, preincubation of hepatocytes with the PI3K activator could attenuate this effect (vs. HSC-MVs at 2 × 108/mL; Fig. 3a, b). These data indicate that MVs from TGF-β1-stimulated HSCs could aggravate the APAP/H2O2-injured hepatocytes viability by inhibiting PI3K pathway.
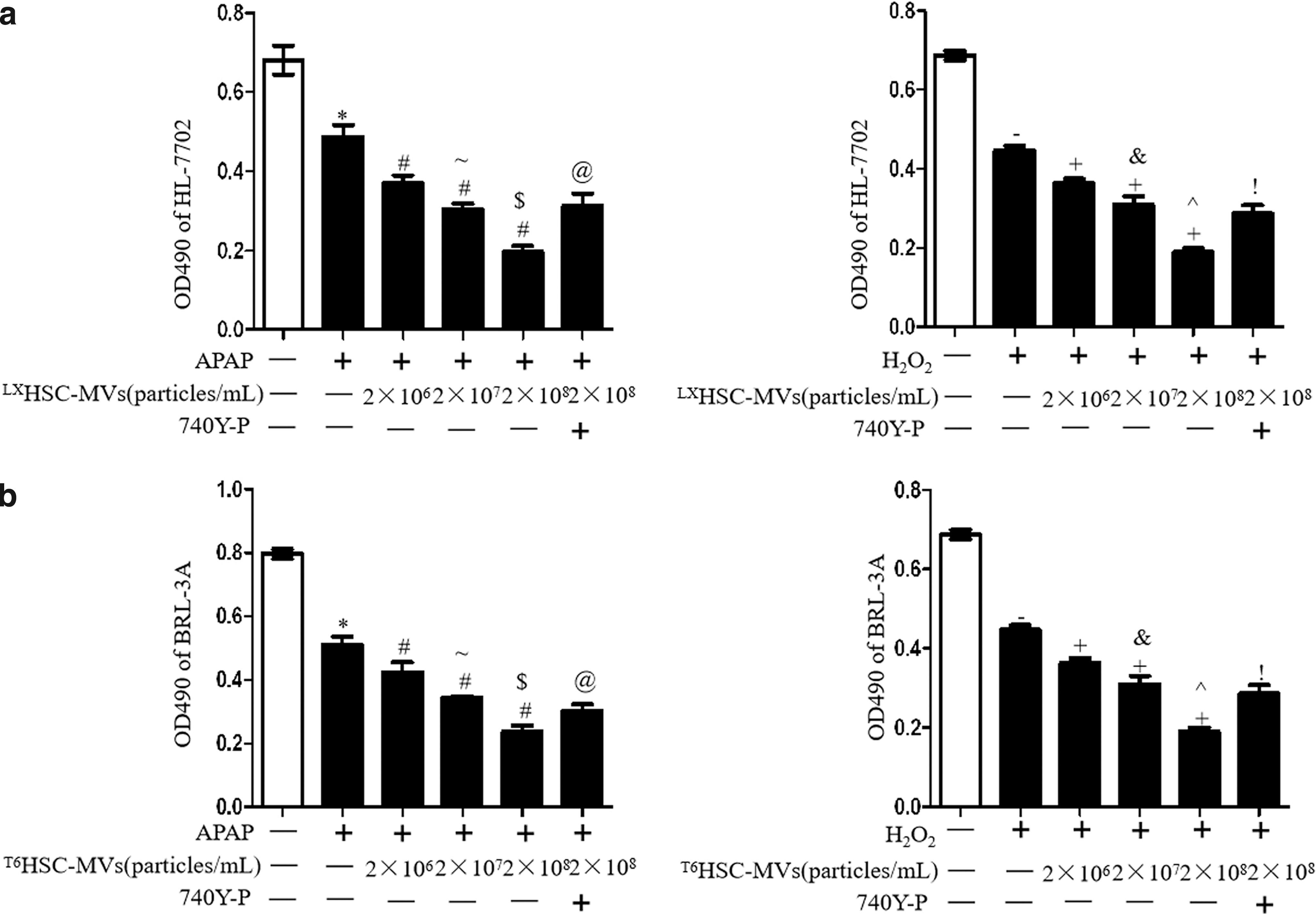
HSC-MVs dose dependently decreased viability and expression of p-Akt/Akt in injury models of hepatocytes.
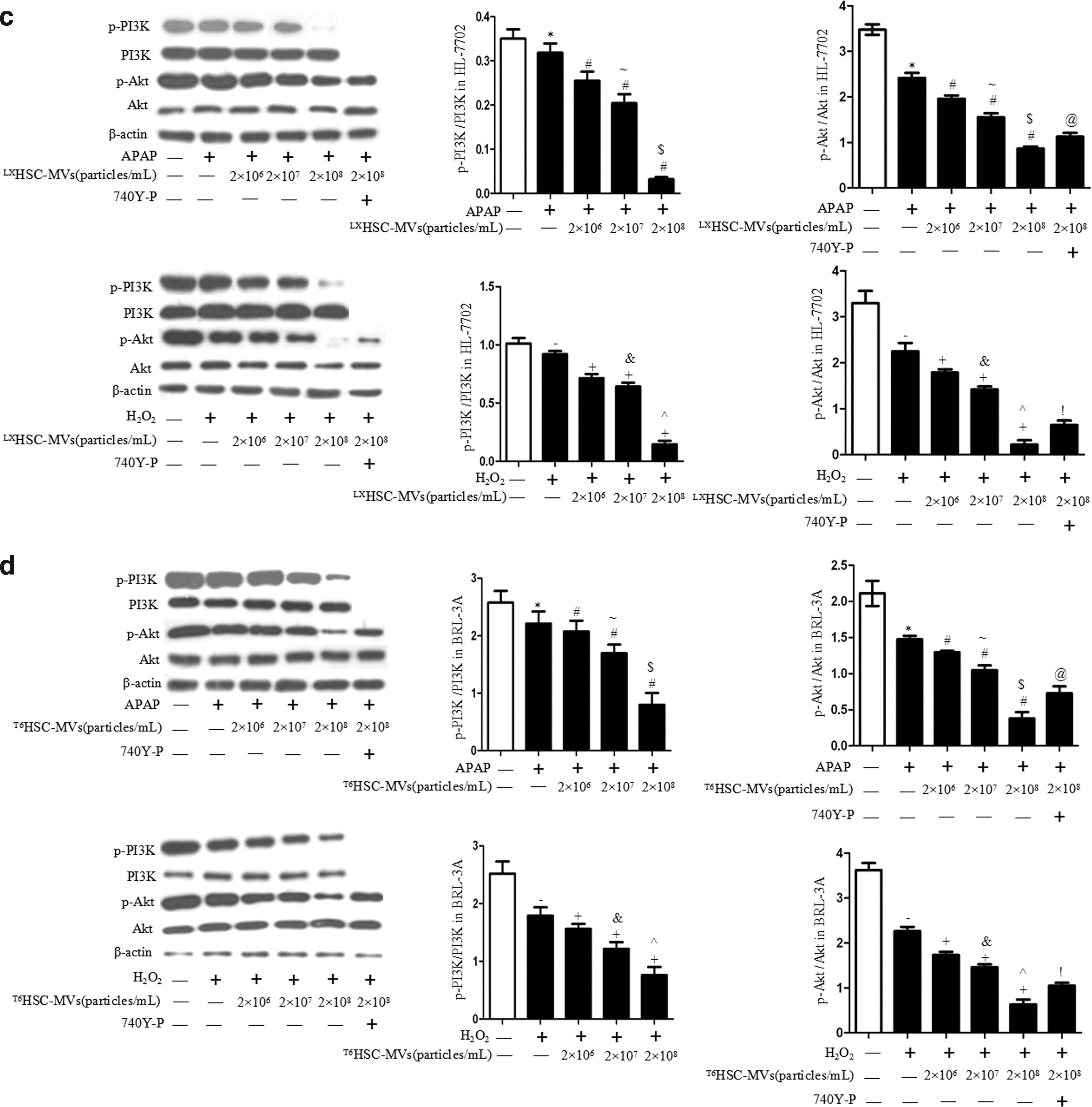
Meanwhile, HSC-MVs dose dependently downregulated the levels of p-PI3K/PI3K and p-Akt/Akt in APAP/H2O2-treated BRL-3A and HL-7702 (vs. treatment control; Fig. 3c, d). The levels of p-PI3K/PI3K and p-Akt/Akt in HSC-MVs-treated cells were lower at a higher dose as compared with those at a lower dose (vs. HSC-MVs at 2 × 107/mL or 2 × 106/mL; Fig. 3c, d). Again, preincubation of hepatocytes with 740Y-P attenuated these effects (vs. HSC-MVs at 2 × 108/mL; Fig. 3c, d). These data indicate that the inhibitory effect of HSC-MVs on viability of hepatocytes is closely related to the PI3K/Akt pathway.
HSC-MVs dose dependently increased cell apoptosis through modulating PI3K/Akt and Erk1/2 pathway in hepatocytic injury models, paralleling with caspase-3 upregulation
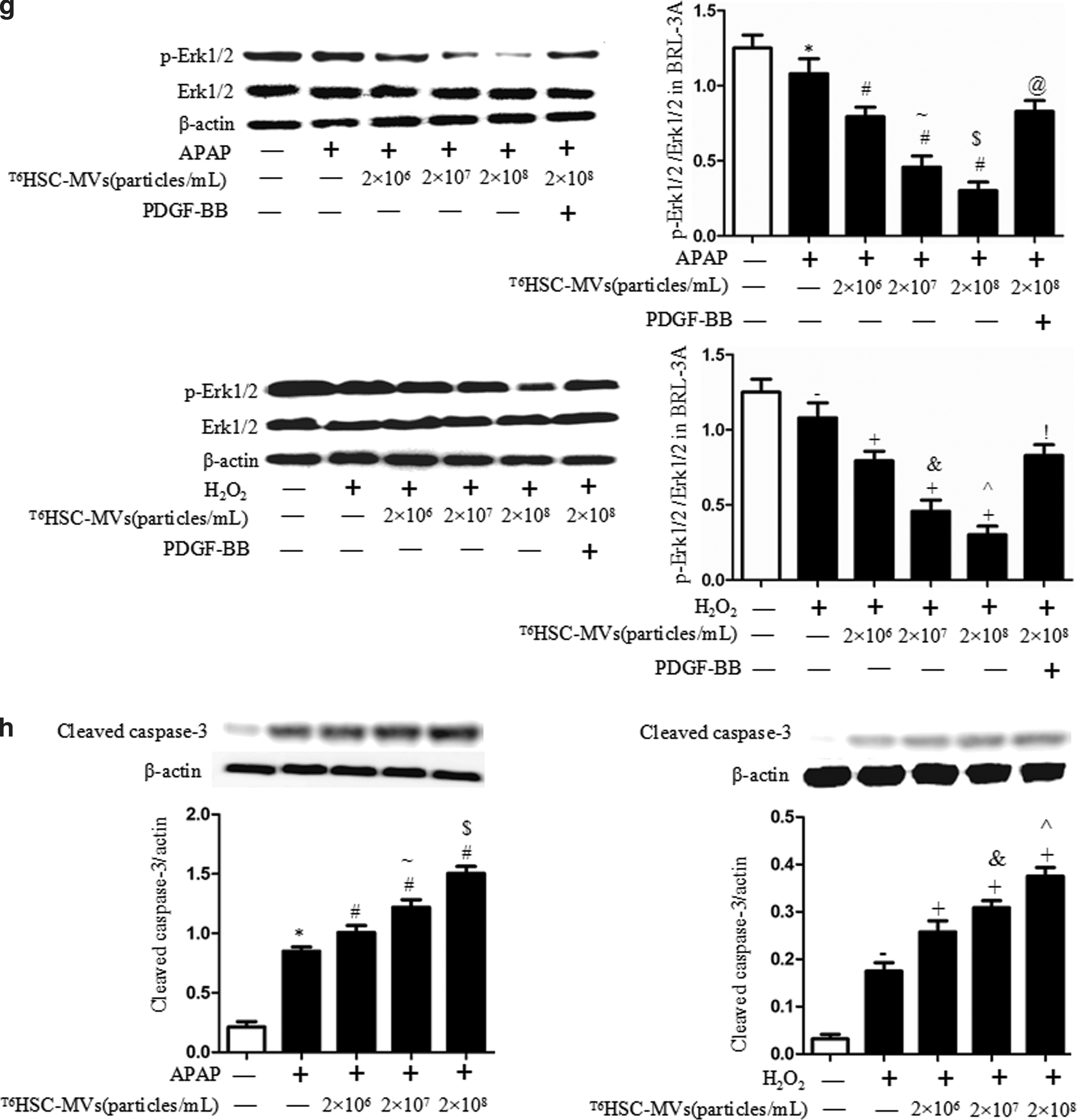
Annexin V-PE/7-AAD analysis and Hochest 33258 staining revealed that APAP or H2O2 increased apoptosis of BRL-3A and HL-7702 cells (vs. normal control; Fig. 4a–d). HSC-MVs dose dependently increased the cell apoptotic rate and cleaved caspase-3 protein level, while decreased the phosphorylation of Erk1/2 when compared with untreated group (vs. treatment control; Fig. 4a–h). These effects were more obvious at a higher dose than at a lower dose (vs. HSC-MVs at 2 × 107/mL or 2 × 106/mL; Fig. 4a–h). In addition, preincubation of hepatocytes with the PI3K activator or Erk1/2 activator could attenuate the proapoptotic effect of HSC-MVs (vs. HSC-MVs at 2 × 108/mL; Fig. 4a–h). These data indicate that the proapoptotic effect of HSC-MVs on apoptosis of hepatocytes is related to PI3K/Akt, Erk1/2 pathway, and cleaved caspase-3 expression.

HSC-MVs dose dependently increased apoptosis and expression of cleaved caspase-3 in injured hepatocytes models.


HSC-MVs dose dependently increased the release of AST and ALT of hepatocytic injury models through modulating PI3K/Akt pathway
Our result showed that APAP or H2O2 promoted the leakages of AST and ALT in BRL-3A and HL-7702 hepatocyte (vs. normal control; Fig. 5a, b). After treated with HSC-MVs, the leakages of AST and ALT in BRL-3A and HL-7702 cells were significantly increased (vs. treatment control; Fig. 5a, b). We also observed that the deleterious effect of HSC-MVs was dose dependent. We found that the leakages of AST and ALT in HSC-MVs-treated cells were higher at a higher dose than at a lower dose (vs. HSC-MVs at 2 × 107/mL or 2 × 106/mL; Fig. 5a, b). Again, preincubation of hepatocytes with the PI3K activator could attenuate this effect (vs. HSC-MVs at 2 × 108/mL; Figs. 5a and 4b). These data indicate that MVs derived from TGF-β1-activated HSCs do dependently promote the leakage of ALT and AST of hepatocytes through inhibiting PI3K/Akt pathway.
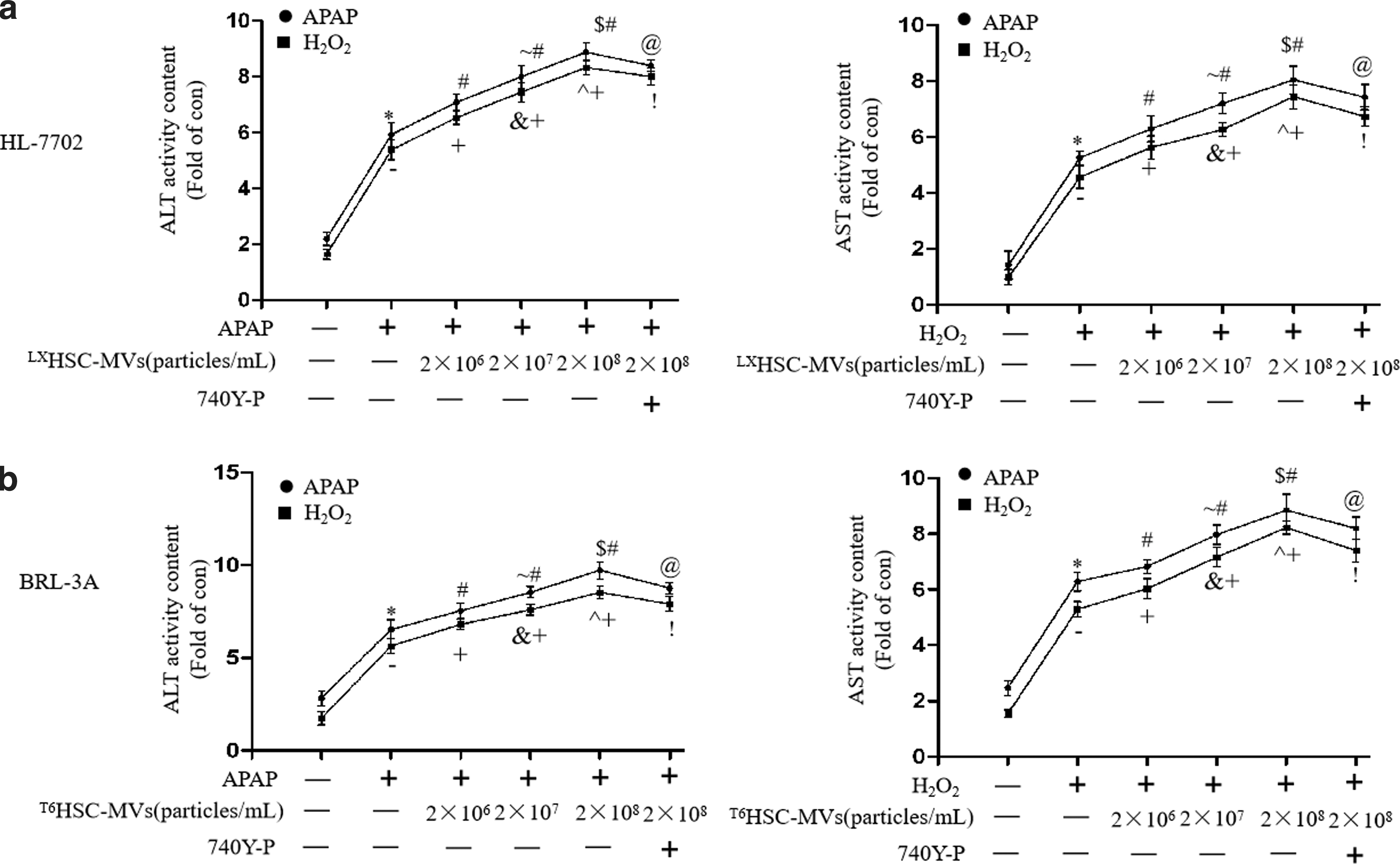
HSC-MVs dose dependently increased the leakage of AST and ALT in injured hepatocyte models.
Discussion
In this study, we found that MVs secreted from HSCs challenged with TGF-β1 stimulation could merge with hepatocytes, which significantly aggravates APAP or H2O2-induced hepatocytic injury, as indexed by cell viability and apoptosis and the leakage of ALT and AST. The effects of HSC-MVs on hepatocytes were associated with a decrease in phosphorylation of PI3K and Akt, which could be abolished by 740Y-P. The proapoptotic effect could also be inhibited by PDGF-BB, which was accompanied with upregulation of cleaved caspase-3. Our findings indicate that MVs from TGF-β1-activated HSCs can mediate HSCs–hepatocytes communication for aggravating the injury of hepatocytes.
It has been shown that activation of HSCs contributes to the pathogenesis of liver injury [30]. TGF-β1 is a key factor involved in the pathological basis of activation of HSCs [2]. APAP and H2O2 have been used to induce apoptosis of hepatocytes, oxidative stress, and dysfunction [12,31]. APAP causes liver injury by both covalent modification of protein targets and oxidative stress-mediated injury pathway, whereas H2O2 induces liver injury through the oxidative stress pathway [13]. Thus, we used APAP and H2O2 to build two models of hepatocytic injury in this study. We found APAP or H2O2 could significantly induce the apoptosis of hepatocytes, suggesting the successful construction of hepatocytic injury model. MVs can play functional roles in target cells, and their effects are mainly dependent on the stimuli to the source cells [11,14,32]. We have previously demonstrated that MVs from HSCs under normal environment exerts a protective role in hepatocytes [12]. In this study, we generated MVs from TGF-β1-activated HSCs and examined their functional role in these two cell models. We found that MVs released from TGF-β1-activated HSCs dose dependently aggravated cell viability and apoptosis in these cell models. Interestingly, our recent study has demonstrated that MVs released from HSCs at the quiescent stage showed protective effects on APAP/H2O2-induced hepatocytic injury [12]. Taken together, these results indicated that MVs released from different status of HSCs might exert different effects on hepatocytic injury. These results are similar to our previous finding that MVs derived from endothelial progenitor cells under starving stress or apoptotic stress play diverse effects on hypoxia/reoxygenation-induced EC apoptosis [14]. Our findings add new evidence to the paracrine effects of activated HSCs [33,34]. It is known that the injured hepatocytes can produce a mass of TGF-β1, which, in turn, promotes the activation of the HSCs [2,35]. In this case, MVs derived from TGF-β1-activated HSCs represent a novel mechanism involved in the pathological process of hepatic injury. These observations also suggest that targeting the MVs released from TGF-β1-activated HSCs could serve as a therapeutic target for liver injury. However, our study was merely carried out in vitro. Whether MVs secreted from TGF-β1-activated HSCs could merge with hepatocytes in liver tissue and the functions in vivo needs further exploration.
The serum levels of ALT and AST are highly correlated with the integrity of the liver cells. It is well known that extensive increase of ALT and AST in serum has been considered as important indicators for liver injury [33]. Previous study reported that APAP/H2O2 increased the level of ALT and AST in hepatocytes [12]. In this study, we confirmed that APAP or H2O2 increased the leakage of ALT and AST in hepatocytes. As expected, MVs released from TGF-β1-activated HSCs dose dependently increased the medium level of ALT and AST in APAP or H2O2-treated hepatocyte. These findings provide further evidence to support the deleterious effects of HSC-MVs on APAP or H2O2-induced hepatocytic injury.
To further understand the underlying mechanisms of HSC-MVs-induced hepatocyte dysfunction, we examined the PI3K/Akt pathway that has been reported to involve in liver function [36]. We found that HSC-MVs dose dependently decreased p-PI3K/PI3K and p-Akt/Akt in APAP or H2O2-induced injury in HL-7702 and BRL-3A hepatocytes, which could be partially abolished by 740Y-P. In addition, activation of the PI3K/Akt pathway with 740Y-P abolished the deleterious effects of HSC-MVs on the viability, apoptosis, and leakage of ALT and AST. Erk1/2 pathway has also been reported to be closely related to the hepatocyte functions [37]. Therefore, we analyzed the effect of Erk1/2 activator on apoptosis of hepatocytes. Results showed that treatment with Erk1/2 activator markedly reduced the APAP/H2O2-induced apoptosis of hepatocytes, paralleled with upregulation of Erk1/2 phosphorylation. Moreover, cleaved caspase-3 is an important proapoptotic factor, playing a key role in the execution phase of cell apoptosis [38]. Herein, our results demonstrate that the HSC-MVs induced apoptosis of hepatocytes coupled with cleaved caspase-3 upregulation. These data indicate that HSC-MVs worsen APAP or H2O2-induced hepatic injury through the inhibition of PI3K/Akt, Erk1/2 pathways, and promotion of cleaved caspase-3. Nevertheless, more studies on the exact functional contents of HSC-MVs and detailed mechanisms are needed.
Conclusion
Our data demonstrate that MVs derived from HSCs under TGF-β1 stimulation exert deleterious effects on toxicant-induced hepatocytes injury through the PI3K/Akt, Erk/1/2, and caspase-3 pathways.
Footnotes
Acknowledgments
This study was supported by National Natural Science Foundation of China (NSFC, #81670578) and Guangdong provincial natural science fund subject (2017A030313688).
Author Disclosure Statement
No competing financial interests exist.