
Abstract
Aberrant microRNA expression correlates with age-related osteoporosis, which impairs bone formation by regulating osteoblastic activity, thus leading to age-related bone loss. In this study, we observed that miR-384-5p was significantly upregulated in bone marrow mesenchymal stem cells (BMSCs) from aged rats compared with BMSCs from young rats. In vitro functional assays revealed that overexpression of miR-384-5p in young BMSCs inhibited osteogenic differentiation and accelerated senescence, whereas knockdown of miR-384-5p in aged BMSCs had the opposite effects. Furthermore, we demonstrated that miR-384-5p inhibited the expression of Gli2 at both the mRNA and protein levels by directly binding to the 3′ untranslated region of Gli2 mRNA. The osteogenic capacity of Gli2-knockdown BMSCs was rejuvenated by miR-384-5p inhibition. Finally, in vivo assays showed that the inhibition of miR-384-5p prevented bone loss and increased the osteogenic capacity in aged rats. Overall, our study suggests that miR-384-5p functions as a negative regulator of osteogenesis, indicating that the inhibition of miR-384-5p may be a therapeutic strategy against age-related bone loss.
Introduction
Aging is accompanied by a progressive loss of bone mass, decreased bone quality, and diminished bone strength, which often result in diseases such as osteoporosis [1]. The decrease in the number and activity of bone-forming osteoblasts is involved in the progression of age-related bone loss [2]. Bone marrow mesenchymal stem cells (BMSCs) as the major source of osteoblasts show downregulation with age in their potential to differentiate into osteoblasts and thus cause the age-related bone loss in bone marrow [3,4]. Cellular senescence is one such aging mechanism that could provide an explanation for the deterioration of bone with age [5]. Senescent BMSCs possess impaired multilineage differentiation potentials that decrease the rate of bone formation and enhance predisposition to osteoporosis [6]. With organismal aging, age-related secretory phenotypic molecules are secreted into the bone microenvironment, inhibit osteogenic differentiation, and promote senescence of BMSCs, thereby causing the osteoporotic bone loss [7]. Thus, to understand the pathological mechanisms of bone loss, it is important to identify the genes associated with age-related changes in the osteogenic behaviors of BMSCs.
MicroRNAs (miRNAs, miRs) are a class of endogenous small (19–25 nt) noncoding RNAs that negatively regulate expression of their target genes by binding to the 3′ untranslated region (3′UTR) of mRNA, thus downregulating gene expression by translational inhibition or destabilization of mRNAs [8]. It has been shown that miRNAs control a wide range of physiological and pathological processes [9,10]. Several miRNAs have been found to be involved in osteogenesis. Davis et al. have reported that miR-183-5p expression increases with age in bone-derived extracellular vesicles, reduces BMSC proliferation and osteogenic differentiation, and promotes senescence [11]. Li et al. have demonstrated that miR-188 regulates the age-related switch between osteoblastic and adipocytic differentiation, namely, that BMSC-specific overexpression of miR-188 in mice reduces bone formation and increases bone marrow fat accumulation [12]. miR-182 is a negative regulator of osteoblastic proliferation, differentiation, and skeletogenesis through targeting FoxO1 [13], whereas miR-216a promotes osteoblastic differentiation and enhances bone formation [14]. Despite these findings, the functions of miRNAs in the impaired potency of osteogenic differentiation in aged BMSCs require further investigation.
In this study, we showed that miR-384-5p was significantly upregulated in BMSCs from aged rats compared with young rats. miR-384-5p suppressed osteoblastic differentiation and accelerated the senescence of BMSCs by targeting Gli2. Additionally, we found that knockdown of miR-384-5p prevented bone loss in aged rats. These findings point to potential therapeutic targets in age-related bone disorders, such as osteoporosis.
Materials and Methods
Osteogenic differentiation
All experimental procedures involving animals were approved by the Animal Care Committee of Naval Medical University. BMSCs from young rats (n = 5, 3 months old) and aged rats (n = 5, 21 months old) were harvested from the femur and tibia, as previously described [15]. For osteogenic differentiation, BMSC cultures were expanded by treatment with an osteogenic induction medium (low-sugar Dulbecco's modified Eagle's medium [DMEM] supplemented with 10% of fetal bovine serum, 100 nM dexamethasone, 50 μM ascorbic acid, and 10 mM β-glycerophosphate). The cells were maintained by refreshment of the osteogenic induction medium every 2 days.
Lentiviral vector preparation and infection
Lentiviruses encoding pre-miR-384-5p, an antisense ribonucleotide toward miR-384-5p, Gli2-specific short hairpin RNA (shRNA; shGli2), or a scrambled control ribonucleotide were obtained from GeneChem, Inc. (Shanghai, China). BMSCs were seeded in six-well plates and infected with the recombinant lentivirus-transducing units plus 5 μg/mL polybrene.
Alizarin red S staining and quantification
Mineral deposition was evaluated by Alizarin red S (ARS) staining on day 16 after osteogenic induction. For ARS staining, BMSCs were fixed in 4% paraformaldehyde and washed with distilled water. The cells were incubated with ARS for 10 min and rinsed with distilled water. The stain was eluted with 10% cetylpyridinium chloride (Sigma), and the absorbance was measured at 562 nm on a microplate reader.
Senescence-associated β-galactosidase staining
An assay of cellular senescence-associated β-galactosidase activity (SA-β-gal) was performed in accordance with the instructions of the β-Gal Staining Kit (Cell Signaling Technology, Beverly, MA). Briefly, BMSCs were washed with phosphate-buffered saline and incubated with a staining solution for 12 h. Images of the staining were captured in five randomly selected visual fields, and percentages of positive staining and statistical significance were determined.
Quantitative reverse transcription–polymerase chain reaction analysis
Total RNA was extracted with the TRIzol reagent (Invitrogen, CA) and reverse-transcribed into cDNA using the PrimeScript RT Reagent Kit (TaKaRa, Dalian, China). Quantitative reverse transcription–polymerase chain reaction (qRT-PCR) analysis was performed with the SYBR Premix Ex Taq Kit (TaKaRa). Expression values of genes and miR-384-5p were normalized to Gapdh and U6, and the relative expression levels were calculated by the 2−ΔΔCT method.
Western blot analysis
Cells were obtained by means of a cell scraper and lysed on ice in RIPA buffer supplemented with protease inhibitors. The protein concentration of the samples was determined with the Pierce BCA Protein Assay Kit (Pierce, Rockford, IL), and then, proteins were resolved by sodium dodecyl sulfate-polyacrylamide gel eletrophoresis (SDS-PAGE) in 10% gels and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with 5% fat-free milk and incubated with primary and secondary antibodies. All primary antibodies, that is, those against osteocalcin, osterix, alkaline phosphatase, and p16 were purchased from Abcam, and those against p21 and Gli2 were purchased from Proteintech. The blots were visualized using an ECL Detection Kit (Millipore, Bedford, MA), and quantitative analysis was carried out in the ImageJ software.
A dual luciferase assay
BMSCs infected with the miR-384-5p-overexpressing lentivirus were transfected by means of Lipofectamine 2000 (Invitrogen) with a psiCHECK-2 luciferase reporter plasmid containing either the wild-type (Wt) 3′UTR of Gli2 or the mutant (Mut) 3′UTR of Gli2 with a point-mutated sequence in the binding region for miR-384-5p. After transfection for 48 h, Renilla and firefly luciferase activities were analyzed via a Dual-Luciferase Assay System (Promega, Madison, WI).
Animal experiments
All experimental procedures were approved by the Animal Care Committee of Naval Medical University. Sprague Dawley rats (females, 3 months old) were randomly divided into two groups (n = 5 per group). Each group underwent either sham surgery or bilateral ovariectomy via the dorsal approach. Three months after the surgery, BMSCs in the femur and tibia were harvested from both sham and OVX rats. Sprague Dawley rats (males, 18 months old) were randomly subdivided into two groups (n = 8 per group). The rats of experimental groups received either miR-384-5p-knockdown lentiviral particles (KD-miR-384-5p) or their negative control lentivirus (KD-NC) by periosteal injection into the marrow cavity of the femur. The rats were sacrificed after 3 months, and femur samples were collected and processed for further analysis. Rats were anesthetized with ketamine and xylazine solution intraperitoneally and sacrificed by using thiopental (100 mg/kg) at the end of the experiment.
Microcomputed tomography analysis
The microarchitectural properties of the distal femur were analyzed using a microcomputed tomography (micro-CT) system (SkyScan 1172, Kontich, Belgium). Bones were scanned to obtain high-resolution scanograms, and the selected images of bones were used for three-dimensional reconstruction. The bone volume ratio (bone volume to total volume: BV/TV), Tb.N (trabecular number),
Statistical analyses
Data are presented as mean ± standard deviation. All statistical analyses were performed in GraphPad Prism 5.0 (GraphPad Software, San Diego, CA). Two-tailed Student's t-test was conducted for comparisons between groups, and one-way analysis of variance was carried out for experiments with more than two subgroups. Data with P < 0.05 were considered statistically significant.
Results
miR-384-5p expression is higher in aged BMSCs
To evaluate the involvement of miR-384-5p in aging, we measured miR-384-5p expression in BMSCs from young (3-month-old) rats and aged (21-month-old) rats. qRT-PCR results revealed that miR-384-5p was significantly upregulated in aged rats compared with the levels in young rats (Fig. 1A). Furthermore, miR-384-5p expression was obviously higher in BMSCs from ovariectomized rats compared with sham control rats (Fig. 1B). These results suggested that the upregulation of miR-384-5p may promote age-related bone loss.
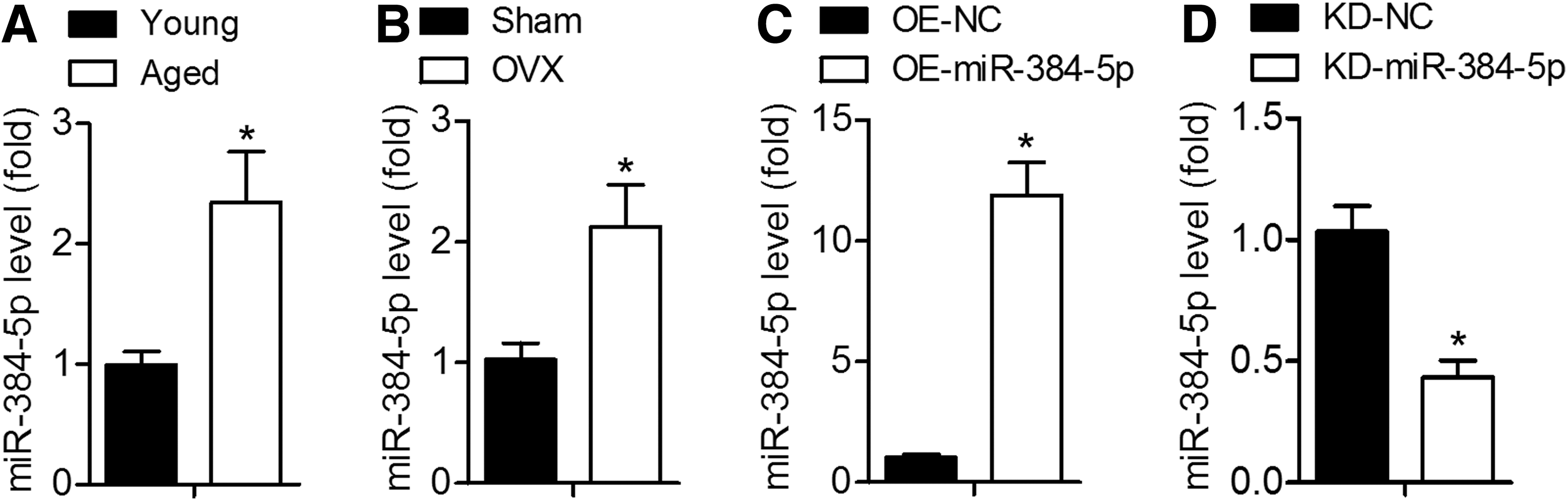
Aging induces miR-384-5p expression in BMSCs.
Overexpression of miR-384-5p inhibits young BMSCs osteogenic differentiation and promotes cell senescence
To determine the role of miR-384-5p in osteogenic differentiation and senescence of BMSCs, miR-384-5p-overexpressing lentiviral particles (OE-miR-384-5p) or their negative control lentiviral particles (OE-NC) were first employed to infect young BMSCs, and qRT-PCR was carried out to confirm miR-384-5p overexpression (Fig. 1C). Calcium deposits were quantified by ARS staining, and the results indicated that calcium deposits were smaller in the OE-miR-384-5p group compared with the OE-NC group (Fig. 2A). We next demonstrated that BMSCs infected with OE-miR-384-5p manifested greater cell senescence compared with OE-NC-infected cells (Fig. 2B). Consistent with these observations, the mRNA and protein expression levels of the osteo-specific genes Ocn, Alp, and Osx were significantly lower, and the expression of senescence-related factors p21 and p16 was higher in BMSCs after miR-384-5p overexpression (Fig. 2C–G). These results indicated that upregulated miR-384-5p contributes to senescence and suppresses osteogenic differentiation of BMSCs.
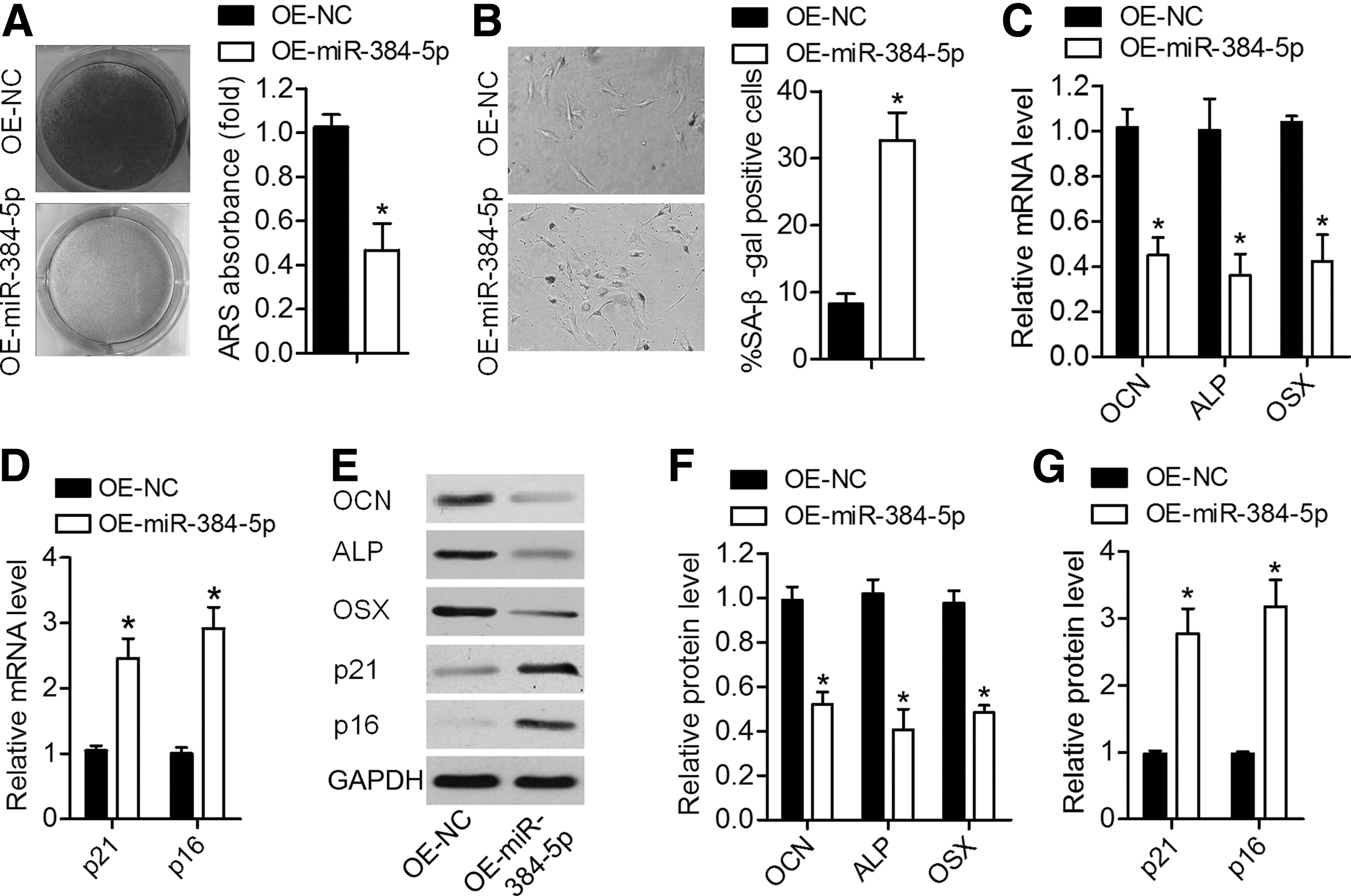
Overexpression of miR-384-5p suppressed osteogenic differentiation of young rat-derived BMSCs and promoted their senescence.
The knockdown of miR-384-5p in aged BMSCs promotes osteogenic differentiation and suppresses cell senescence
We next confirmed that miR-384-5p expression was significantly lower in aged BMSCs infected with KD-miR-384-5p compared with BMSCs infected with KD-NC (Fig. 1D). As presented in Fig. 3A and B, the suppression of miR-384-5p significantly increased mineral deposition detected by ARS staining and decreased cell senescence as determined by SA-β-gal staining. qRT-PCR and western blotting analyses suggested that the miR-384-5p knockdown resulted in significant upregulation of the osteo-specific genes (Ocn, Alp, and Osx) and downregulation of the senescence-related factors (p21 and p16; Fig. 3C–G). These results indicated that the knockdown of miR-384-5p promotes osteogenic differentiation of aged rat-derived BMSCs.
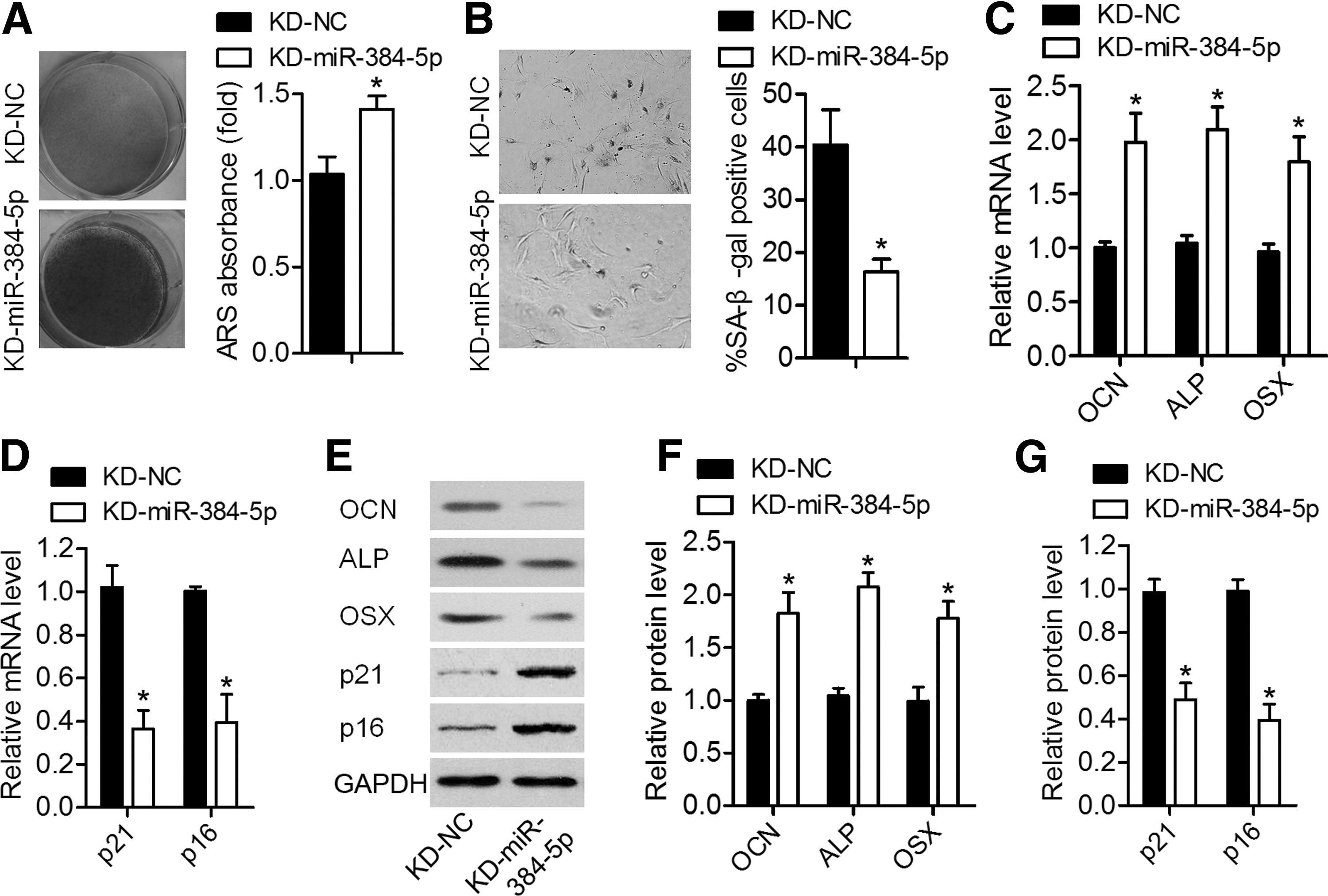
The knockdown of miR-384-5p promoted osteogenic differentiation of aged rat-derived BMSCs and inhibited their senescence.
miR-384-5p binds to the 3′UTR of Gli2 and inhibited its expression
To further elucidate the potential molecular mechanisms involved, target prediction software TargetScan was utilized to predict the possible target genes of miR-384-5p. Among the predicted candidates, we were particularly interested in Gli2 because of its positive roles in osteogenic differentiation [16]. To determine whether Gli2 could be downregulated by miR-384-5p, we constructed luciferase reporter vectors containing the Wt or Mut miR-384-5p target sequences of the Gli2 3′UTR (Fig. 4A). Overexpression of miR-384-5p remarkably suppressed the luciferase activity of the Wt Gli2 3′UTR reporter gene but not of the Mut reporter gene (Fig. 4B). In addition, overexpression of miR-384-5p in young BMSCs significantly decreased Gli2 expression at both the mRNA and protein levels (Fig. 3C, E, F), whereas the knockdown of miR-384-5p in aged BMSCs increased GLI2 expression (Fig. 3D–F). These results indicated that Gli2 mRNA is a direct target of miR-384-5p in BMSCs.
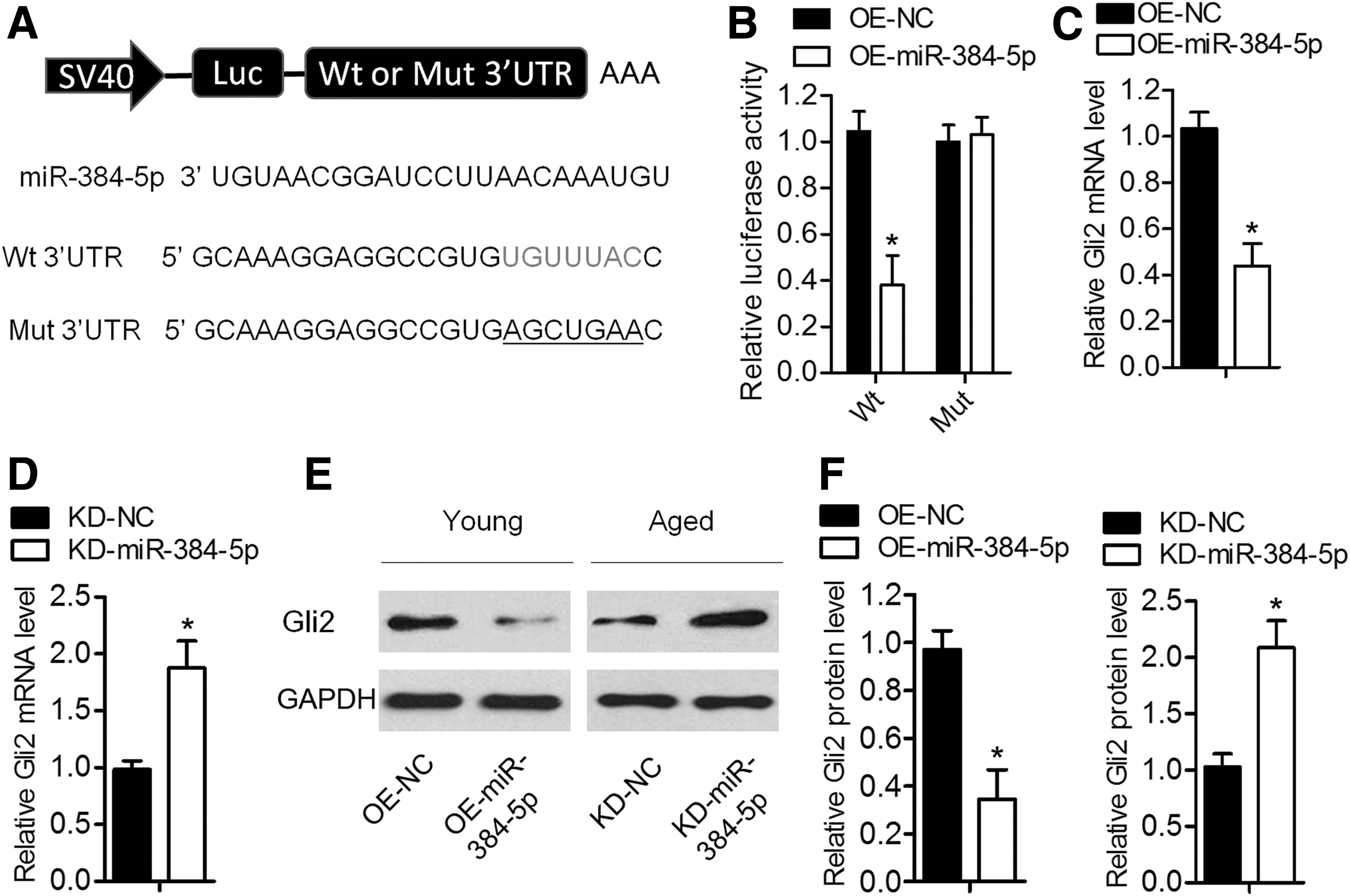
Gli2 mRNA is a direct target of miR-384-5p.
The knockdown of miR-384-5p promotes osteogenic differentiation of BMSCs by upregulating Gli2
To determine the role of GLI2 in the miR-384-5p-mediated BMSC differentiation and senescence, BMSCs were coinfected with shGli2 and KD-miR-384-5p. qRT-PCR and western blotting analyses revealed that mRNA and protein expression levels of Gli2 were significantly lower in BMSCs infected with siGli2, and this suppression was attenuated by the miR-384-5p knockdown (Fig. 5A, B). The shGli2-infected BMSCs manifested lower mineral deposition and higher cellular senescence, and these effects were reversed by the miR-384-5p knockdown (Fig. 5C, D). Likewise, the decreased expression levels of osteo-specific genes and the increased amounts of senescence-related factors were observed in shGli2 BMSCs; these effects were next attenuated by the miR-384-5p knockdown (Fig. 5E–G). These results suggested that miR-384-5p inhibited osteogenic differentiation of BMSCs and induced their cellular aging by suppressing Gli2 expression.
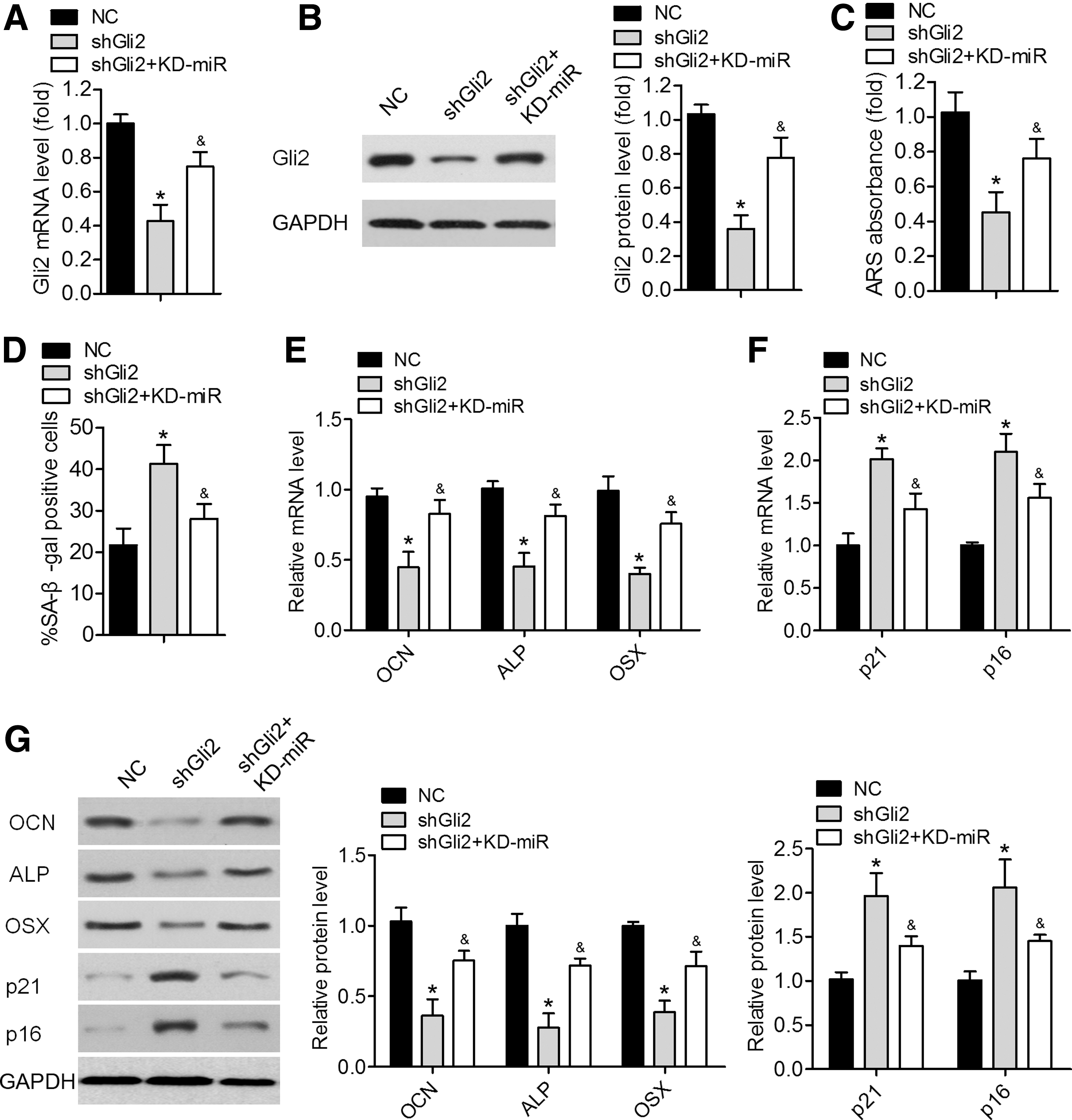
miR-384-5p inhibited BMSC differentiation and induced cellular aging by suppressing Gli2.
miR-384-5p knockdown prevents bone loss in aged rats
On the basis of the above observations, we hypothesized that the inhibition of miR-384-5p prevents bone loss in aged rats. To test this hypothesis, 18-month-old rats were administered with KD-miR-384-5p or KD-NC into the femoral bone marrow cavity. Three months after the administration, qRT-PCR analysis confirmed that BMSCs from KD-miR-384-5p-treated rats showed a significant decrease in the expression of miR-384-5p compared with control cells (Fig. 6A). Furthermore, the micro-CT analysis suggested that the miR-384-5p knockdown remarkably increased trabecular bone volume and number and decreased trabecular separation compared with that in the control group (Fig. 6B–G). These results indicated that the miR-384-5p knockdown prevented bone loss in aged rats.

The miR-384-5p knockdown prevents bone loss in aged rats.
Discussion
Several researchers have found that various miRNAs participate in BMSC osteogenesis and in bone integrity via gene expression regulation and suggested that targeting these miRNAs might be a promising remedy for osteoporosis [17,18]. In our study, we found that the miR-384-5p expression is significantly higher in BMSCs from aged and ovariectomized rats. In addition, we indicated that miR-384-5p obviously suppressed osteogenic differentiation of BMSCs and accelerated their senescence by downregulating Gli2. Moreover, we demonstrated that the knockdown of miR-384-5p prevents bone loss in aged rats. These results improve our understanding of aging-related miRNAs and support the development of miR-384-5p-based therapies for age-related bone loss.
Increasing evidence suggests that many miRNAs become dysregulated in BMSCs with age and play a crucial part in determining the differentiation potential of BMSCs, such as miR-183-5p, miR-342, and miR-188 [11,12,19]. Nonetheless, the regulation of BMSC differentiation involves multiple miRNAs. To study the molecular mechanism of the osteogenic differentiation potential of BMSCs, we identified an upregulated miRNA, that is, miR-384-5p, in the BMSCs of aged and ovariectomized rats compared with BMSCs from young and sham rats, thus indicating that miR-384-5p may affect the process of senescence of BMSCs.
In recent years, there were several reports of miR-384-5p's biological functions. It has been revealed that the inhibition of miR-384-5p alleviates osteoarthritis through suppressive effects on apoptosis of cartilage cells via the nuclear factor κB (NF-κB) signaling pathway and targeting of SOX9 [20]. The downregulation of miR-384-5p attenuates rotenone-induced neurotoxicity in dopaminergic SH-SY5Y cells by inhibiting endoplasmic reticulum stress [21]. Several studies have shown that the upregulation of miR-384-5p by a high-fat diet may impair the beclin-1-mediated protection of macrophages through autophagy to accelerate the development of atherosclerosis and that the downregulation of miR-384-5p in the inflammatory macrophages may enhance macrophage autophagy and contribute to the development of diabetic encephalopathy [22,23]. Besides, miR-384-5p regulates ischemia-induced cardioprotection [24]. Nonetheless, there have been no studies about the action of miR-384-5p on the regulation of BMSC differentiation. In this study, overexpression of miR-384-5p in young BMSCs inhibited osteogenic differentiation and accelerated senescence, whereas the knockdown of miR-384-5p in aged BMSCs had the opposite effects. Moreover, lentiviruses encoding an antisense ribonucleotide toward miR-384-5p were injected into the bone marrow microenvironment, and the results showed that the knockdown of miR-384-5p prevented bone loss in aged rats.
Identification of genes targeted by miRNAs is widely believed to be a key step toward understanding the role of miRNAs in gene regulatory networks [25]. As part of the effort to understand the interactions between miR-384-5p and its target, a publicly available database (TargetScan) was searched here. Gli2 was identified as a potential target gene of miR-384-5p and is associated with osteoblastic differentiation. The hedgehog (Hh) pathway is a major regulator of cellular differentiation, proliferation, and stem cell maintenance [26]. One study indicates that the Hh pathway activates a positive feedback mechanism involving insulin-like growth factors to induce osteoblastic differentiation [16]. GLI2 predominantly exists in the full-length form that becomes a transcriptional activator in response to Hh [27]. Miao et al. have shown that Gli2-null embryos exhibit deficiencies in both bone formation and cartilage angiogenesis [28], and Joeng and Long have revealed that the GLI2 transcriptional activator is a crucial effector for Indian Hh signaling in osteoblast development and cartilage vascularization [29]. That is why in this study, we chose Gli2 as a target of miR-384-5p for further research. In our study, we verified whether Gli2 mRNA is a direct target of miR-384-5p in BMSCs by a luciferase reporter gene assay. Moreover, miR-384-5p overexpression significantly decreased the expression of Gli2 at both the mRNA and protein levels in BMSCs. The osteogenic capacity of siGli2-treated BMSCs was rejuvenated by miR-384-5p inhibition. These results suggest that miR-384-5p decreases osteogenic differentiation of BMSCs and increases senescence by downregulating Gli2.
In conclusion, this study for the first time indicates that miR-384-5p negatively regulates osteogenic differentiation of BMSCs and accelerates cell senescence by directly targeting Gli2 mRNA. To treat bone aging, an antisense approach targeting miR-384-5p may be of therapeutic value.
Footnotes
Author Disclosure Statement
No competing financial interests exist.