
Abstract
Dome formation can occur in cultured tubular epithelial cells originating from various tissues, including the mammary gland and the kidney. The isolation and characterization of normal kidney epithelial stem cells that give rise to dome-forming tubular cells have never been reported. We attempted to isolate and characterize canine kidney epithelial stem cells using a simple cell culture method that we have previously used to isolate other adult human stem cells. Dome-forming kidney epithelial cells were derived from dissociated adult canine kidney tissues that were cultured in a modified keratinocyte serum-free medium supplemented with N-acetyl-
Introduction
The incidence and prevalence of end-stage renal disease (ESRD) are increasing globally [1,2]. Patients with ESRD have been treated by renal replacement therapies including peritoneal dialysis, hemodialysis, and kidney transplantation. These treatments, however, have inherent shortcomings, including comorbidities, poor quality of life, and a shortage of organ donors.
Stem cells are undifferentiated cells with the capacity for unlimited or prolonged self-renewal, and the remarkable potential to give rise to many different cell types in the body. Therefore, stem cells show great potential in regenerative and reparative medicine for the treatment of diseases that are not conventionally curable. There are three major types of stem cells. Embryonic stem (ES) cells might provide an unlimited supply of stem cells due to their infinite lifespan and high differentiation potential. However, drawbacks of ES cells include the potential for teratoma development besides ethical concern. Induced pluripotent stem cells (iPSCs) can be derived after ectopic expression of transcription factors in many different animal and human cell types. Although there are no ethical concerns associated with the development of iPSCs, potential drawbacks of using these cells include mutations, dysplasia, or hyperplasia due to multiple retroviral chromosomal integrations, or aberrant expression of transcription factors [3]. Adult stem cells can be derived from various adult tissues using suitable cell culture media and conditions, without the need for ectopic expression of transcription factors. These adult stem cells may have limited proliferative capacity and differentiation potential compared with ES cells and iPSCs. However, adult stem cells are safer for clinical use and are by far the most common type of stem cell used in clinical trials (34, 52, and 573 clinical trials have used ES cells, iPSCs, and adult stem cells, respectively, as of July 30, 2019;
We previously reported the development of various animal and human adult stem cells from a range of sources (ie, mammary [4,5], adipose tissue [6,7], gastric [8], liver [9], and amniotic fluid [10]). Successful development of these cells was mainly ascribed to the use of a medium containing N-acetyl-
Materials and Methods
Cell culture medium
The medium used to develop putative canine kidney epithelial stem/progenitor cell cultures is a modified MCDB 153 medium (Keratinocyte-SFM; Gibco) supplemented with 2 mM NAC (catalog no. A8199; Sigma) and 0.2 mM Asc-2P (catalog no. A8960; Sigma) (referred to as K-NAC medium) [6]. The calcium concentration of this medium is 0.09 mM. The growth factors/hormones added to this medium include recombinant epithelial growth factor (5 ng/mL), bovine pituitary extract (50 μg/mL), insulin (5 μg/mL), hydrocortisone (74 ng/mL), and 3,3′,5-triiodo-
Isolation of dome-forming canine kidney epithelial cells
A whole canine kidney was excised shortly after euthanasia, which was carried out for reasons unrelated to this study. The kidney was washed with phosphate-buffered saline (PBS), and then the outer fascia and fibrotic tissue were removed and minced into small pieces in a 60 mm plate using scalpels. After washing the minced tissue with PBS to remove the majority of erythrocytes, small pieces were seeded into 60 mm plates and incubated with serum-free K-NAC medium supplemented with 0.1% penicillin–streptomycin (catalog no. 15140-122; Gibco). To facilitate the attachment of suspended cells and tissues to the plate, minimal initial volumes of medium were used. Medium was renewed daily, and cell colonies were developed from pieces of tissue after 7–10 days (Fig. 1a). These colonies were isolated using the glass ring/trypsin method, and were transferred into wells of 12-well plates and incubated with K-NAC medium supplemented with 10% fetal bovine serum (FBS) and 5 mM nicotinamide. These cells continued to grow until confluency and exhibited contact-insensitive growth and dome formation (Fig. 1b). Some cells would detach and float in the medium; these floating cells could be repeatedly transferred to 100 mm plates to give rise to a large number of cells. This procedure was found to be more efficient than trypsinization, which tended to induce cell differentiation. Dome-forming colonies in 100 mm plates could be enriched and purified by removing undesired nondome-forming colonies by scraping with a rubber policeman. Every other day, half of the cell culture medium in plate (conditioned medium) was replaced with fresh medium. Cells in 100 mm plates could then be trypsinized and transferred to T-75 flasks to increase number of cells. All cell cultures were incubated at 37°C in incubators supplied with humidified air and 5% CO2.
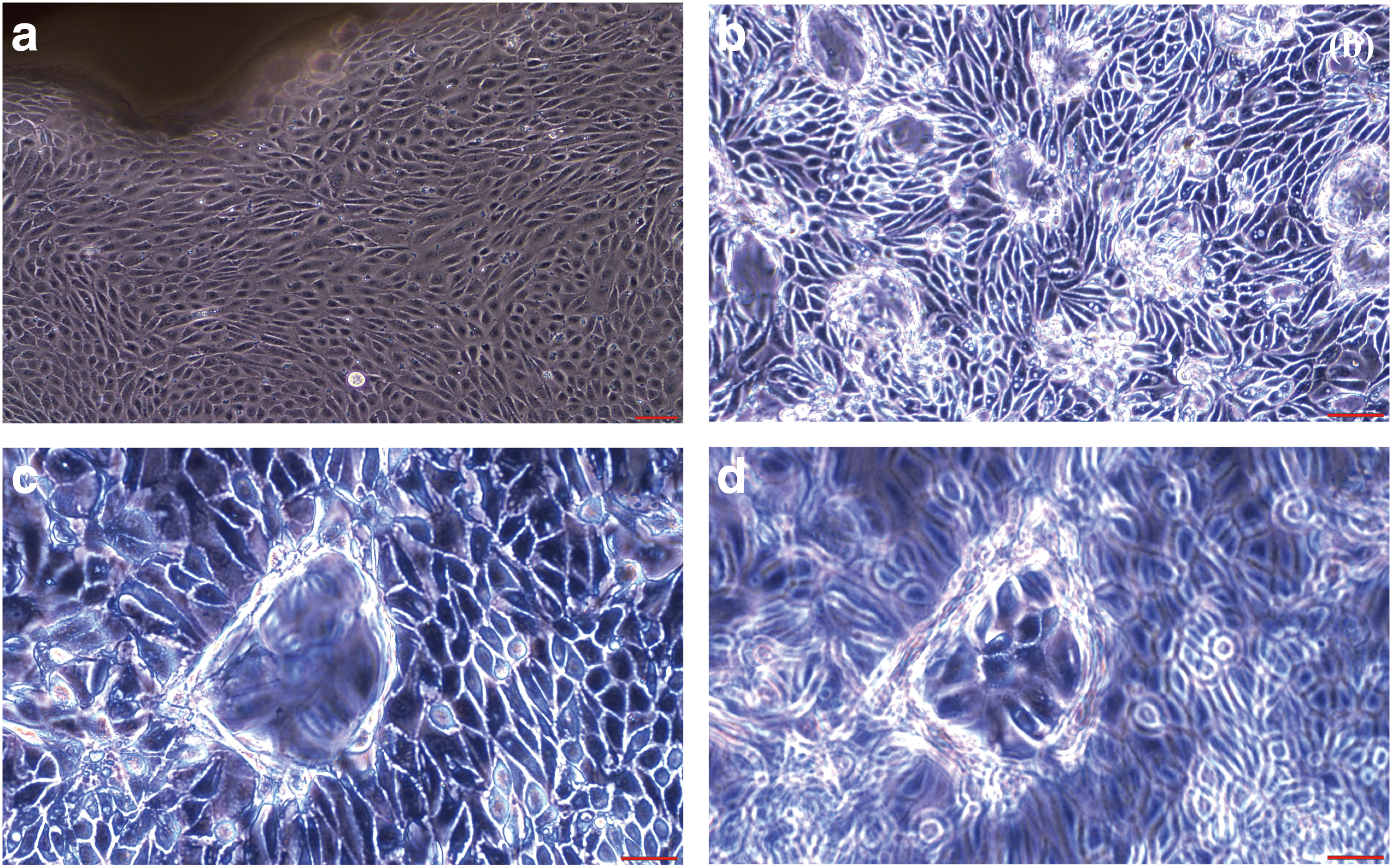
Primary canine kidney epithelial cell cultures were successfully established from canine kidney tissue. Initial cell growth from minced canine kidney tissue is shown
Cumulative population doubling levels
The cumulative population doubling level (cpdl) was calculated to determine the proliferation capacity of cells using the equation: ln (N f/N i)/ln2, where N i and N f are initial and final number of cells, respectively, and ln2 is the natural log. The initial number of cells was 2 × 105 cells in a 100 mm dish for each propagation. To obtain the final cpdl, cpdl in continuous subculture periods was added.
Anchorage-independent growth assay
To determine anchorage-independent growth (AIG), agarose (0.5%, Type I; Sigma) was prepared in K-NAC medium containing 5% FBS at 39°C, and was added to 60 mm dishes and allowed to solidify in a 37°C incubator. A total of 50,000 cells suspended in medium containing 0.33% agarose were overlaid on top of the solidified 0.5% agarose layer. The plates were incubated at 37°C, and K-NAC medium containing 5 mM nicotinamide and 10% FBS was added 3 days after cell inoculation and renewed every 3 days. After 4 weeks, colonies were stained and scored using a microscope.
Gap junctional intercellular communication
Gap junctional intercellular communication (GJIC) was studied by the scrape loading/dye transfer technique reported previously [22]. In brief, this technique involves (1) the growth of a confluent cell culture in 35 mm plates over a period of 1–2 days; (2) the addition of 2 mL Lucifer Yellow (catalog no. L0259, 0.5 mg/mL in PBS; Sigma); (3) scraping of the cell monolayer with a surgical blade to allow entry of the dye into damaged cells, which transfer the dye to neighboring cells within 3 min if the cells are GJIC competent; and (4) removal of the dye solution and washing of the cell monolayer with PBS. Dye transfer between cells was visualized with a Nikon Eclipse TE300 UV fluorescence microscope and recorded using a digital camera connected to a computer.
Flow cytometry
Subconfluent canine kidney epithelial cells (clone DK-701) were grown as described previously in 60 mm dishes and dissociated from the dishes using a nonenzymatic dissociation solution (Cellstripper; Corning). Fluorescence-activated cell sorting (FACS) buffer (Hank's balanced salt solution, 1% bovine serum albumin, 0.1% NaN3) was used to wash cells between staining and fixing steps. Cells were incubated with FACS buffer containing 20% fetal calf serum and antimouse CD16/CD32 (BD Biosciences) to help block Fc receptors, if present. Cells were then incubated with antibodies CD24 (clone M1/69; Biolegend) and CD133 (clone 13A4; eBiosciences) and washed three times with FACS buffer. BD Cytofix™ (BD Biosciences) was used to fix cells per the manufacturer's protocol. Cells were analyzed on an FACS BD Canto II™ (BD Biosciences). After acquisition, data analysis was performed using FLOWJO v10 software.
Reverse transcription polymerase chain reaction
Total RNA was extracted from cells using a Versagene RNA purification kit (Gentra) and treated with DNase I to remove contaminating DNA. cDNA was synthesized from 1 μg total RNA using random hexamer primers and Superscript III reverse transcriptase (Invitrogen). For amplification of cDNA, we used primers specific for OCT4, SOX-2, and NANOG (stem cell transcription factors) that were derived from coding regions of the corresponding genes [7]. A total of 25 μL polymerase chain reaction (PCRs) were prepared with 2 μL cDNA, 5 pmol of each primer, 0.5 U of Taq polymerase (Invitrogen), and final concentrations of 40 μM deoxyribonucleotide triphosphate, 2 mM MgCl2, 20 mM Tris–HCl, and 50 mM KCl. Cycling conditions were as follows: 94°C for 4 min; 30–35 cycles at 94°C for 1 min, annealing for 1 min, and 72°C for 1 min; followed by 72°C for 5 min. PCR products were separated on 2% agarose gels by electrophoresis, stained with ethidium bromide, and visualized under UV light. The digital images were captured using AlphaImager software. PCR products were gel purified using a Qiaex II Gel Extraction Kit (Qiagen), sequenced with an automated sequencer, and verified by alignment to the canine genome.
Multipotent differentiation
Adipogenesis
A total of 4 × 105 cells were plated in 35 mm dishes and treated with rosiglitazone (5 μM), 3-isobutyl-1-methylxanthine (IBMX; 500 μM), dexamethasone (1 μM), indomethacin (100 μM), insulin (10 μg/mL), glucose (4.5 g/L), and 5% rabbit serum for 21 days. The induction medium with supplementations was renewed every 3 days. Oil red O staining was performed to reveal lipid droplet formation.
Osteogenesis
A total of 4 × 105 cells were plated in 35 mm dishes and treated with dexamethasone (0.1 μM), Asc-2P (50 μM), and β-glycerophosphate disodium (10 mM; DAG cocktail) in D-medium containing 10% FBS for 6–8 weeks, with medium change every 3 days. Alizarin red staining was carried out to detect calcified extracellular matrix deposit.
Chondrogenesis
Micromass cultures of cells (1 × 105 cells/10 μL) were allowed to form in 24-well plates for 2.5 h before treatment with transforming growth factor-β1 (10 ng/mL), Asc-2P (50 μM), and insulin (6.25 μg/mL) (TAI cocktail) for 14 days, with medium change every 3 days. The micromass was stained with Alcian blue for the presence of sulfated proteoglycan-rich matrix.
Three-dimensional growth on Matrigel
Liquid Matrigel was added to 35 mm dishes (0.5 mL/plate) and allowed to solidify in an incubator at 37°C. A total of 1 × 107 cells in 0.5 mL K-NAC medium containing 10% FBS and 5 mM nicotinamide were then added onto the Matrigel for three-dimensional cell growth.
Regulation of self-renewal and dome-forming tubular differentiation of canine stem/progenitor cells by STAT3
For this study, 4 × 104 cells were plated in 35 mm dishes for 1 day and then treated with AG490 (2.5, 5, or 10 μM and dimethyl sulfoxide [DMSO] as vehicle control) or lipopolysaccharide (LPS) (0.1, 0.2, or 0.4 μg/mL). The medium was renewed once every 3 days. Both dome-forming and nondome-forming colonies were counted after 4 weeks.
Statistical analysis
Mean ± SD data were analyzed using one-way analysis of variance followed by post hoc Dunnett's multiple comparison test (GraphPad Prism).
Results
Canine kidney epithelial cell cultures were successfully established from canine kidney tissues
Canine kidney cell colonies were initially derived from minced kidney tissues. We used the serum-free K-NAC medium [6] to develop cell populations with characteristics of stem cells. The use of this cell culture medium and the methods described previously yielded colonies displaying epithelial cell morphology after 7–19 days, as shown in Fig. 1a. These colonies were isolated by glass ring/trypsin method and propagated for further characterization.
Canine kidney epithelial cells that were capable of forming dome-like structures can be continuously derived from single cells of the initial dome-forming cell culture
Although canine kidney cell colonies were initially derived in serum-free K-NAC medium [6], we observed that these cells grew more efficiently in K-NAC medium supplemented with 5 mM nicotinamide and 10% FBS. We, therefore, used this serum-containing medium to determine the cells' proliferation potential and colony-forming capacity on plastic or soft agar.
The isolated kidney epithelial cell colonies proliferated in wells and displayed contact-insensitive growth (Fig. 1b). Dome-like structures appeared in confluent cell cultures (Fig. 1c, d), similar to those reported for the canine MDCK cell line [11]. These cells showed density-dependent growth after subculture, that is, optimum cell proliferation density >4,000/cm2. Furthermore, confluent dome-forming canine kidney epithelial cell cultures in 12-well plates could be maintained >13 months with medium change. Floating cells that detached from these adherent cultures could also be repeatedly transferred to new wells or plates to initiate new dome-forming cultures.
Canine kidney epithelial cells express stem cell markers on the cell surface
To examine whether the population of canine kidney epithelial cells express the well-known cell surface markers for renal stem/progenitor cells (see references in “Discussion” section), we performed flow cytometry to analyze the expression of CD24 and CD133 on the surface of normal canine cells (clone DK-701). We found that 22.2% of the cells express CD24 on the surface (Fig. 2a, c) and 5.13% of the cells expressed CD133 on their surface (Fig. 2b, c). In addition, double positive cells expressing both CD24 and CD133 were present at 2.66% (Fig. 2c).
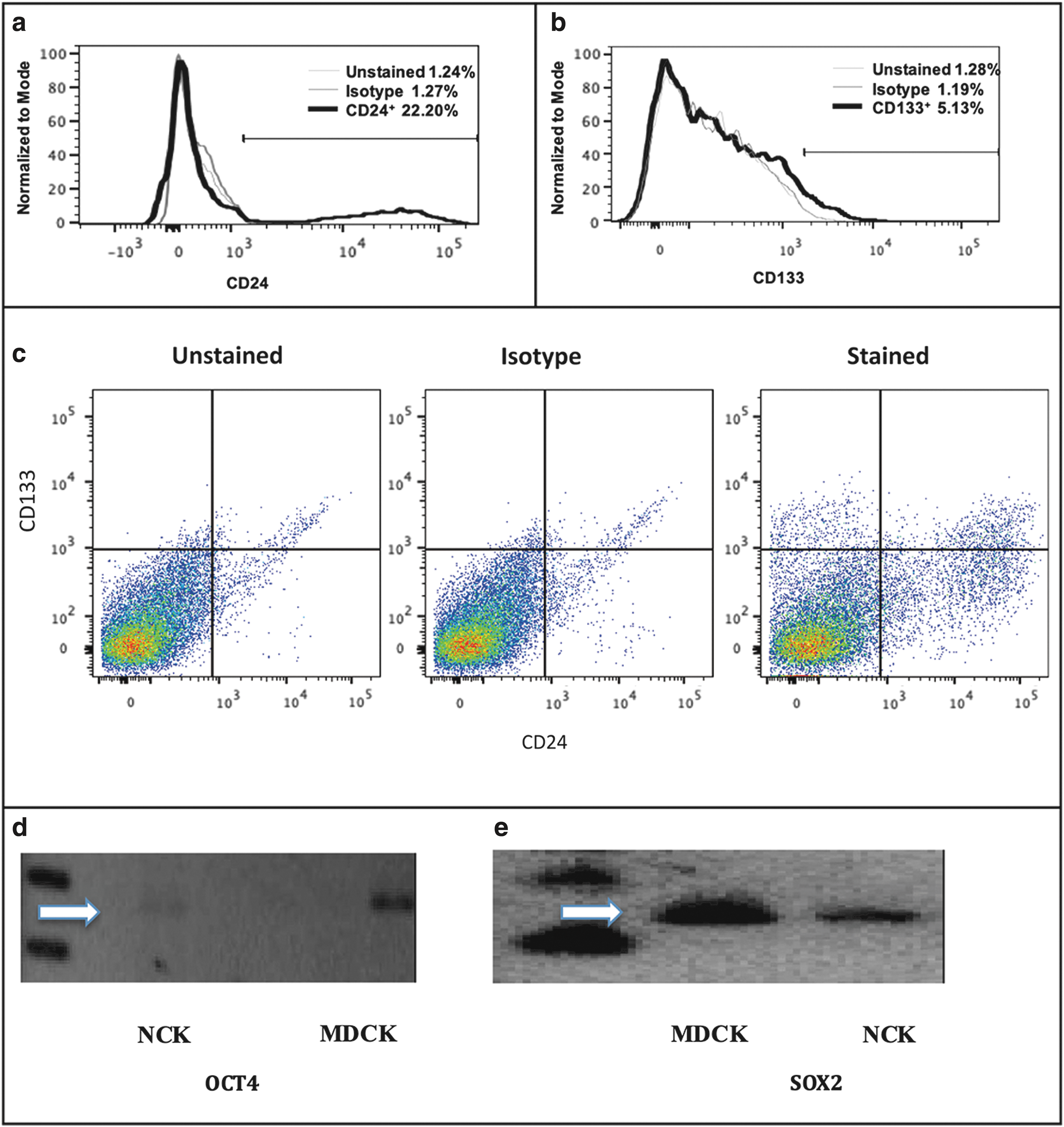
Canine kidney epithelial cells express kidney stem cell markers. Flow cytometry analysis shows that canine kidney epithelial cells (clone DK-701) contain cell populations that express CD24 and CD133 on the surface. Histogram plots for CD24
Canine kidney epithelial cells express pluripotency-associated transcription factors
Experiments were carried out to detect the expression of important stem cell markers by reverse transcription PCR. Both the canine kidney epithelial cells developed in this study, and MDCK cells, expressed OCT4 and SOX-2 as shown by the expression of mRNA encoding OCT4 in Fig. 2d (274 bp product) and SOX-2 in Fig. 2e (142 bp product) in normal canine kidney (NCK) cells and MDCK cells. However, NANOG expression was not detected in these cells (data not shown).
Canine kidney epithelial cells proliferate in culture for finite time and display AIG in soft agar
The proliferation potential of dome-forming canine kidney epithelial cells was determined by estimating their cpdl in continuous culture. We found that these cells had a finite lifespan, with ∼30 population doublings in a period of 42 days. These cells were also capable of colony formation in soft agar (ie, AIG) (Fig. 3a). The frequency of AIG was found to be ∼4% (Fig. 3a) compared with an AIG frequency of ∼8% for MDCK cells (Fig. 3b).
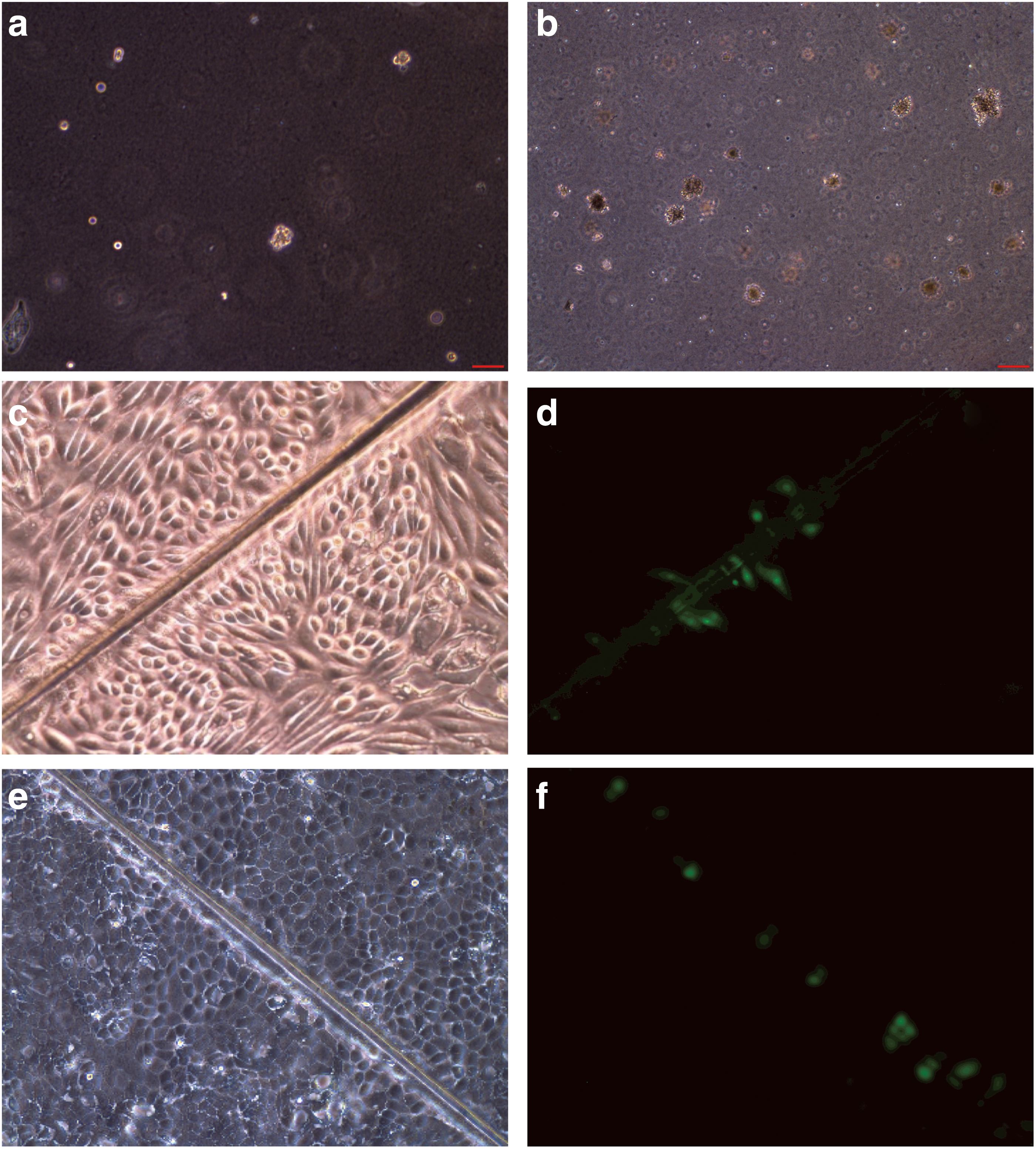
Canine kidney epithelial cells exhibit several features of stem cells. The AIG frequency of dome-forming canine kidney epithelial cell cultures on Matrigel
Canine kidney epithelial cells are deficient in GJIC
The lack of GJIC is a common property of stem cells and cancer cells [23]. Therefore, we carried out experiments to determine whether the canine kidney epithelial cells developed in this study, and immortalized canine MDCK cells, were capable of GJIC. Experiments using the scrape-loading dye transfer technique [22] revealed that both of these cell types are GJIC deficient as shown in Fig. 3c and d for normal canine epithelial cells and in Fig. 3e and f for MDCK cells. As shown, the dye retained in the damaged cells without transferring to neighboring cells as shown by GJIC-deficient cells [22].
Canine kidney epithelial cells are capable of multilineage differentiation
We have previously reported reliable protocols for osteogenic, adipogenic, and chondrogenic differentiation for human and canine adipose-derived mesenchymal stem cells [6,7]. Applying these protocols, we found that canine kidney epithelial cells were capable of osteogenic, adipogenic, and chondrogenic differentiations as revealed by positive Alizarin red (Fig. 4b), Oil red O (Fig. 4d), and Alcian blue staining (Fig. 4f), respectively, as compared with the control treatment shown in Figs. 4a, c, and e.
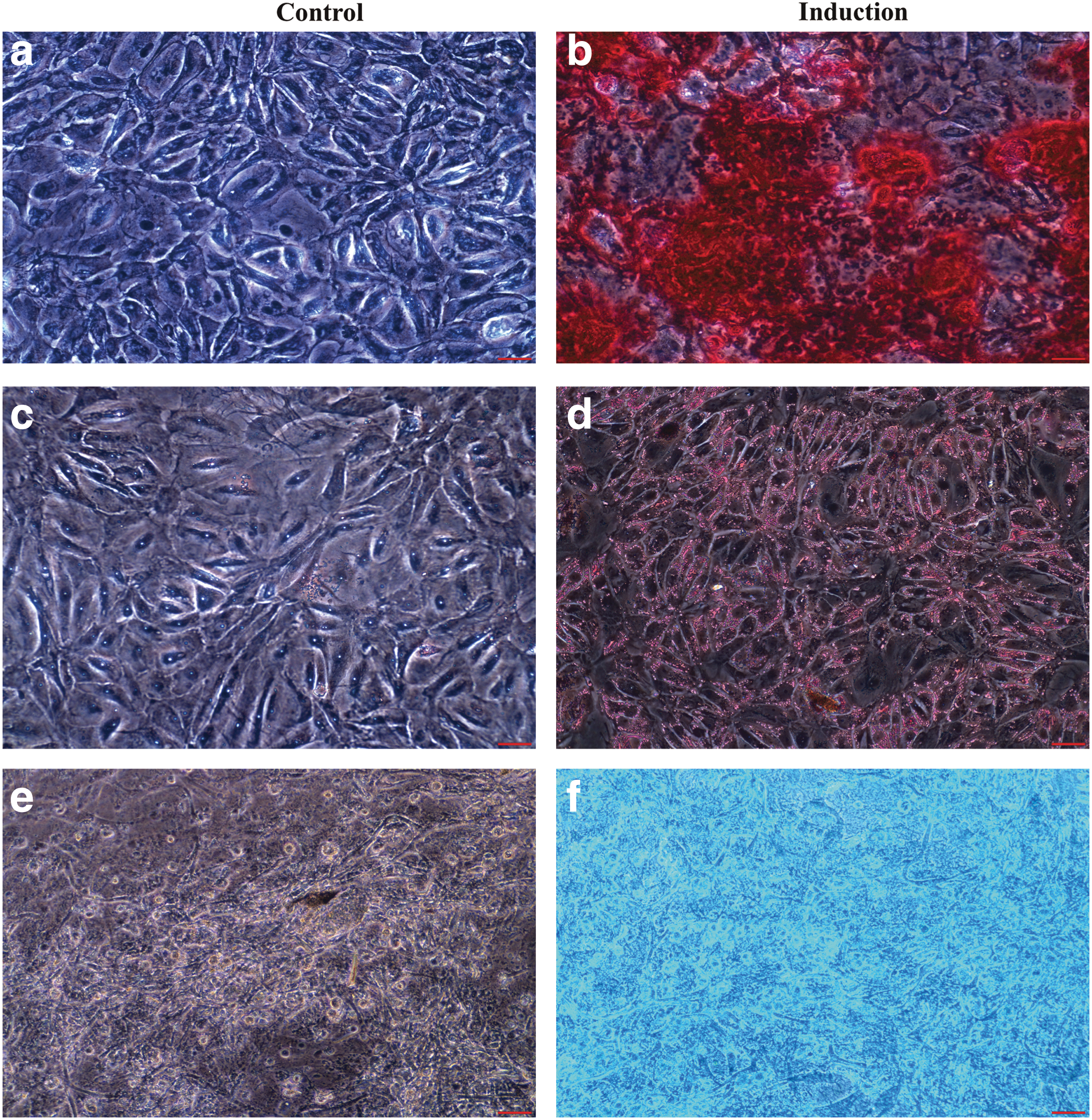
Canine kidney epithelial cells can differentiate into mesenchymal lineages. NCK epithelial cells underwent osteogenic, adipogenic, and chondrogenic differentiation after treatment with respective induction cocktails
Canine dome-forming kidney cells form ductal tubule-like organoids on Matrigel
After canine kidney epithelial cells were plated on Matrigel as described in the Materials and Methods section, tubule-like structures formed in 10–14 days and ductal tubule-like organoids emerged 4–6 weeks later. Tubule-like structure and tubular budding from ductal tubule organoids formed by NCK epithelial cells are shown in Fig. 5a and b, respectively. Figure 5c and d shows the ductal tubule-like organoids formed by NCK epithelial cells on Matrigel.
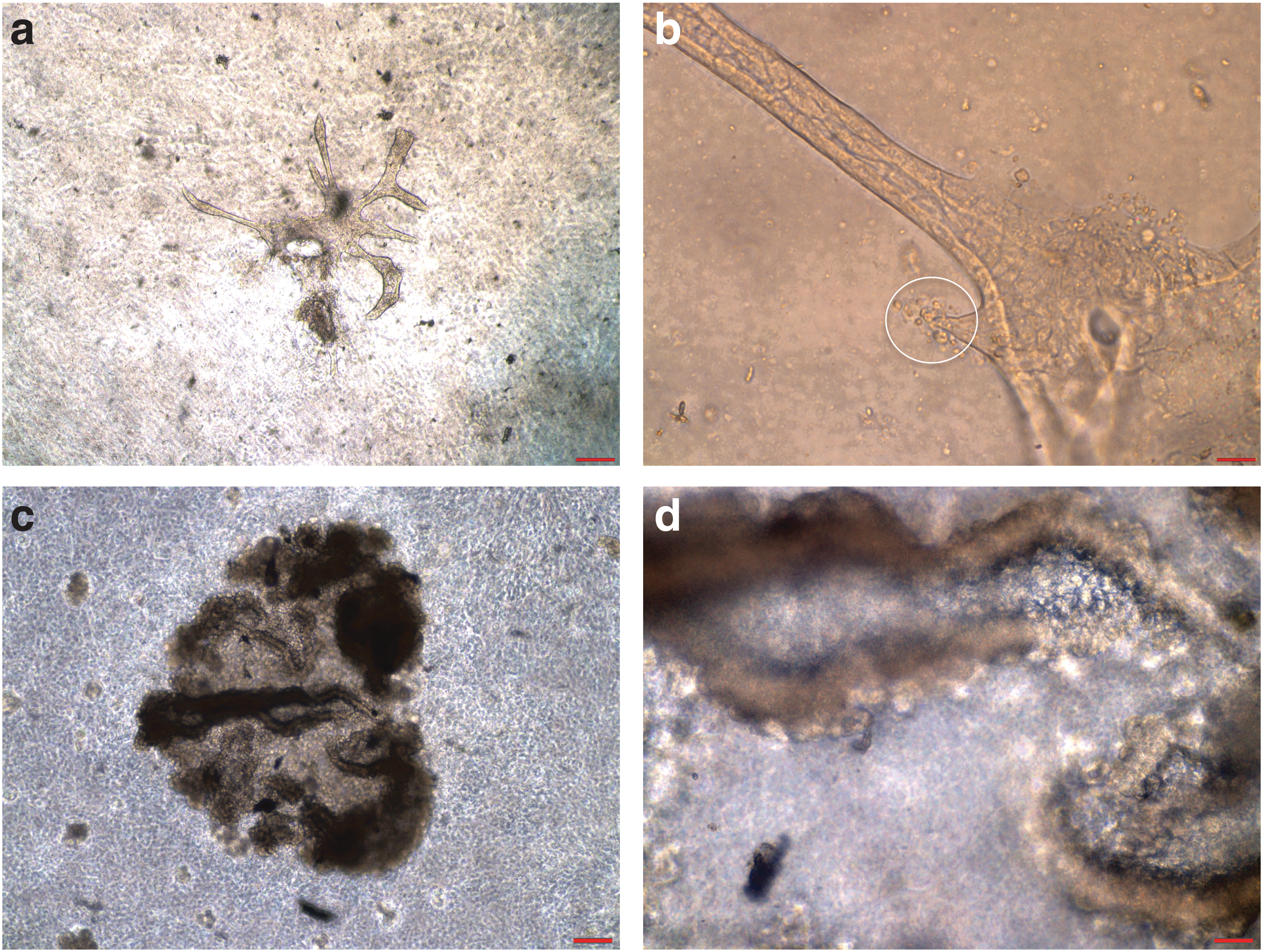
Canine kidney epithelial cells form ductal tubule-like structures. Ductal tubule-like organoids formed by NCK epithelial cells on Matrigel are shown
Regulation of self-renewal and dome-forming tubular differentiation of canine stem/progenitor cells is STAT3 dependent
The transcription factor STAT3 is required for the maintenance and self-renewal of ES cells [24]. In canine MDCK cells, cell confluence has been reported to activate STAT3 and trigger epithelial dome formation [13]. Therefore, we tested the effect of AG490, a STAT3 phosphorylation inhibitor, or LPS, an activator of STAT3, on self-renewal and dome formation. Experiments to determine the effect of STAT3 phosphorylation inhibitor AG490 on colony-forming efficiency were carried out. As shown in Fig. 6a, AG490 inhibited the formation of colonies in a dose-dependent manner. An average of 99, 81, 71, and 22 colonies per plate were found for DMSO vehicle control and treatments with 2.5, 5, or 10 μM AG490, respectively (the respective P values were 0.0942, 0.0251, and 0.0005). AG490 treatment significantly affected dome formation only at concentrations >5 μM. In contrast, treatment of these cells with LPS, which activates STAT3, increased colony-forming frequencies in a dose-dependent manner (an average of 13, 15, 23, and 26 colonies per plate were found for the control and treatments with 0.1, 0.2, or 0.4 μg/mL LPS, respectively (the respective P values were 0.5542, 0.0201, and 0.0079) (Fig. 6b).
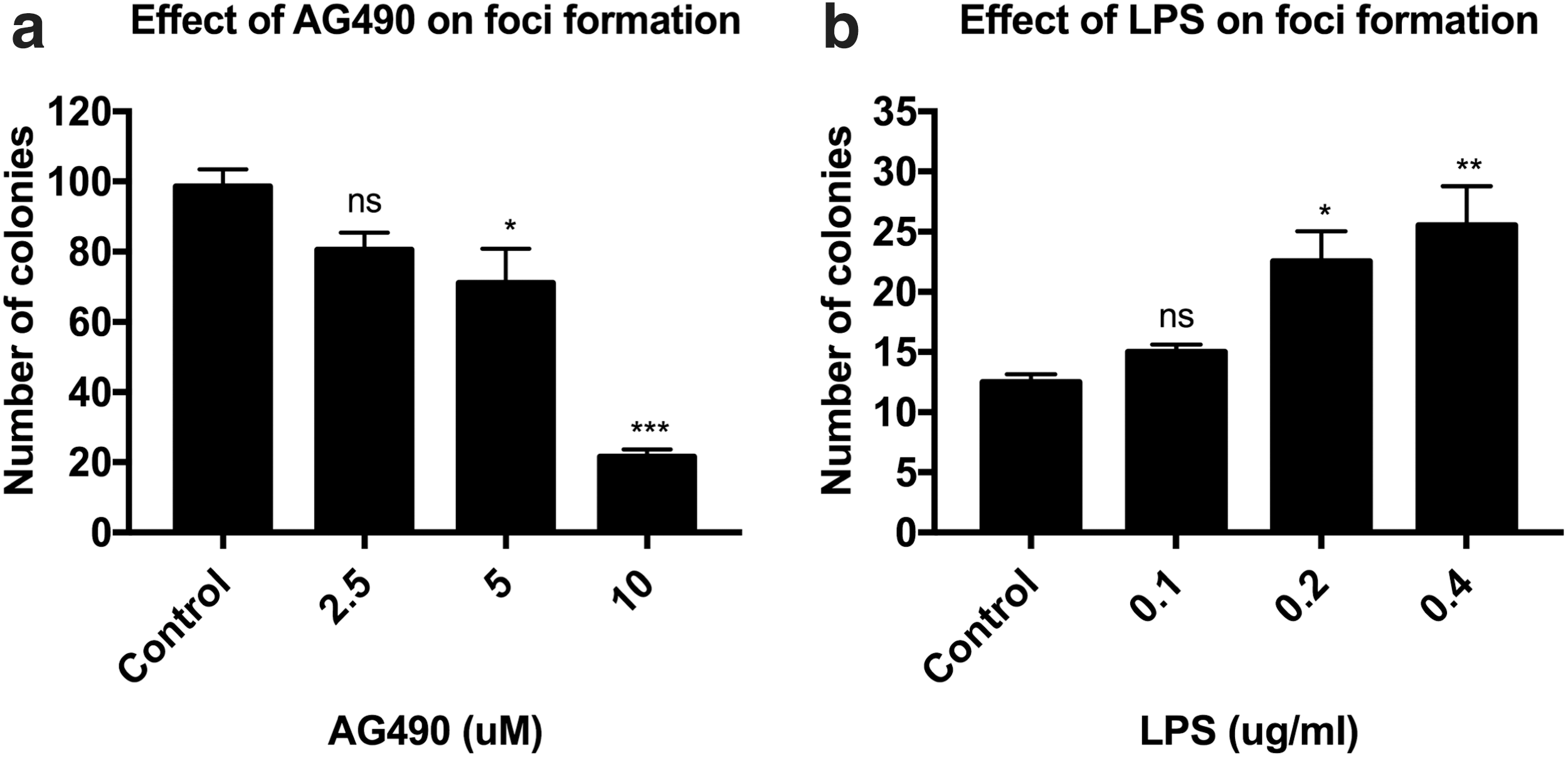
STAT3 modulators can alter the ability of NCK epithelial cells to form colonies. The STAT3 phosphorylation inhibitor AG490 inhibited colony formation by NCK epithelial cells in a dose-dependent manner
Discussion
Dome formation can occur in cultured tubular epithelial cells originating from various tissues including the mammary gland [25] and the kidney [11]. This study demonstrates that dome-forming cells can be isolated from adult canine kidney tissues. Other than the well-known immortal canine kidney epithelial MDCK cell line, the isolation of dome-forming normal canine epithelial cells has not been reported. Furthermore, we show that these cells possess several characteristics of stem cells, that is, (1) high self-renewal potential; (2) the ability to differentiate into multiple cell lineages; (3) expression of the stem cell markers OCT4 and SOX-2; (4) expression of cell surface markers CD24 and CD133; and (5) deficiency in GJIC.
In continuous subculture, these normal kidney epithelial cells were able to undergo ∼30 population doublings. In confluent cultures in 12-well plates, floating cells that detached from the cell monolayer could be continuously collected and transferred into new plates to establish new cultures for >13 months. These new cell cultures maintained their dome-forming abilities. This appears to be a novel method for the long-term growth of stem cells that are sensitive to terminal differentiation after trypsinization. These NCK epithelial cells were capable of multilineage differentiation into adipocytes, osteoclasts, and chondrocytes, as well as tubular structure-like organoids on Matrigel. Whether these cells can also differentiate into other cell types remains to be examined.
The canine kidney epithelial stem cells derived in this study express OCT4 and SOX-2, but not NANOG. This does not cast doubt on their qualification as stem cells, since the reprogramming of human renal proximal tubular cells into iPSCs required only OCT4 and SOX-2 [26]. Compared with the immortal canine MDCK cell line, NCK cells expressed lower levels of OCT4 and SOX-2. This is likely due to the lower frequency of stem cells in the population as indicated by their lower colony-forming efficiency (0.18%). We found that ∼22% cells in the canine kidney epithelial cell population expressed CD24, ∼5% cells expressed CD133, and ∼2.7% cells expressed both CD24 and CD133. It was previously shown that CD24 was found to be highly expressed by renal progenitor cells [27]. Similarly, CD133+ kidney cells expressed several stem cell markers and integrated into developing tubules [28]. In fact, kidney cells expressing both CD24 and CD133 exhibited stem cell features such as self-renewal, multilineage differentiation, and expression of stem cell transcription factors including OCT4 [29 –33]. The lower percentage of cells expressing both CD24 and CD133 compared with cells expressing only CD24 or CD133 found in this study may indicate that CD24+/CD133+ cells are indeed the original stem cells that could undergo self-renewal or give rise to cells expressing only one of these two markers. This can be tested in a future study.
The canine kidney epithelial cells developed in this study share many stem/progenitor cell characteristics with the well-known immortal MDCK canine kidney cell line, that is, dome formation, deficiency in GJIC, expression of OCT4 and SOX-2, and the ability of AIG. The frequency of AIG of the cells described here was ∼4%, which is higher than the colony-forming efficiency from single cells on plastic surfaces (0.18%), but lower than the frequency of AIG (∼8%) for the MDCK cell line.
The mentioned lines of evidence strongly suggest that both the adult canine kidney epithelial cells developed here, and the MDCK cell line, are canine kidney stem/progenitor cells. MDCK cells at confluency were found to activate STAT3 transcription and to trigger dome formation [13]. For our NCK epithelial cells, we observed that sustained cell growth required a high cell density (>4,000/cm2) and that dome formation was found in high cell density areas. We, therefore, suspect that, in both cell types, high cell density induces STAT3 expression, which promotes both stem cell self-renewal and differentiation into tubular cells. In agreement with the mentioned hypothesis, the STAT3 inhibitor AG490 was found to reduce colony-forming efficiency (Fig. 6a), whereas the STAT3 activator LPS was found to increase colony-forming efficiency (Fig. 6b). Putative adult stem cells were found at multiple locations in the kidney (proximal tubule, renal papilla, interstitium, Bowman's capsule, and glomerular parietal epithelium) of mice, rats, and humans [34,35]. Various methods have been used to isolate these putative renal stem cells, including retention of dyes, side populations, specific markers, and cell culture methods [35]. These renal/progenitor stem cells may have therapeutic potential [36] as exemplified by the three-dimensional reconstruction of kidney structures in vitro [37] and the repair of renal damage in mice with severe combined immunodeficiency [30].
Human kidney stem/progenitor cells can be developed from embryonic kidneys [30], tubular renal cells after iPSC reprogramming with OCT4 and SOX-2 [26], and fetal [38] or adult [29,39 –46] kidneys. The isolation of kidney stem/progenitor cells from embryonic and adult tissues relies primarily on the expression of cell surface markers, that is, CD133/CD24 [30,39 –46]. After isolation, the potential therapeutic use of these cells depends on identifying proper media for long-term growth of a large number of cells. Human fetal kidney stem cells were found to be contact insensitive (ie, colonies formed on a layer of human fibroblasts that had been lethally irradiated with X-rays), GJIC deficient [38], and positive for OCT4 expression [46]. To our knowledge, ours is the first study to demonstrate that kidney stem/progenitor cells can give rise to dome-forming tubular cells. We believe that our cell culture method will be useful in the development of human stem cells from adult kidneys. This method is straightforward and requires relatively little tissue to develop sufficient quantities of stem cells for potential therapeutic applications.
Conclusion
We have isolated and characterized the adult canine kidney epithelial cells with stem cell characteristics that can give rise to dome-forming tubular cells. Our results are consistent with the hypothesis that high cell density induces STAT3 expression, which promotes both stem cell self-renewal and differentiation into tubular cells. Our cell culture method should be useful for the development of normal human kidney stem cells for clinical applications and for studying mechanisms of nephrotoxicity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
T.C. and S.J. were supported by Kaohsiung Chang Gung Memorial Hospital CMRPG8G1201. Research materials were supplied by Dr. C.-C. Chang's laboratory. M.N. was supported by the Comparative Medicine and Integrative Biology graduate program, and the Post DVM fellowship from Michigan State University.