
Abstract
Hepatic ischemia reperfusion injury (IRI) remains a major obstacle in liver resection and transplantation surgery, especially in diseased organs. Human mesenchymal stromal cells (MSCs) are reported to acutely alleviate hepatic IRI in mice by releasing bioactive membrane-enclosed extracellular vesicles (EVs), but the long-term effects of MSC-derived EV on hepatic IRI are unknown. Given the considerable differentiation capacity of fibroblasts (FBs) during wound healing and their morphological similarities with MSC, the present study aimed to investigate the potential of these two cell types and their cell-derived EV in attenuating liver damage after IRI. EVs were isolated and purified from the supernatant of MSC and FB cultures and, subsequently, characterized by electron microscopy, nanoparticle tracking analysis, and western blot. Liver injury and organ regeneration in a murine in vivo model of IRI were assessed by serum transaminase levels, histopathology, and immunohistochemistry. Changes in expression of inflammation-associated genes within liver tissue were evaluated by reverse transcriptase quantitative polymerase chain reaction. MSC, MSC-derived EV, FB, and FB-derived EV were systemically administered before hepatic IRI. We found that MSC and MSC-derived EV decreased serum transaminase levels, reduced hepatic necrosis, increased the amount of Ki67-positive hepatocytes, and repressed the transcription of inflammation-associated genes. Although they had no impact on organ damage, FB and FB-derived EV showed some regenerative potential in the late phase of hepatic IRI. Compared to FB, MSC and their derived EV had a stronger potential to attenuate liver damage and improve organ regeneration after hepatic IRI. These results suggest that the key therapeutic factors are located within EV.
Introduction
Hepatic ischemia reperfusion injury (IRI) is a major clinical obstacle in liver transplantation [1] and resection surgery [2], especially in livers burdened by pathological changes such as steatosis, fibrosis, inflammation, and others. Although reperfusion is urgently needed after ischemia, it paradoxically contributes to tissue injury and organ destruction [3]. Complications associated with IRI can result in liver failure following hepatic resection and cause initial graft dysfunction or primary nonfunction [4,5]. One potential option to increase the number of available donor organs in transplantation is to further extend the utilization of organs from expanded criteria donors (eg, livers with macrosteatosis and or long cold ischemia time, donors aged beyond 70 or 80 years), but these organs carry an even higher risk for the development of severe IRI followed by impaired initial graft function [6]. In oncological liver resections, the organ function is increasingly challenged by chemotherapeutic agents in the course of standard multimodal therapies and very often limits major liver resections. Thus, effective strategies to minimize organ damage by preventing IRI will improve the clinical outcome of liver surgery and organ transplantation.
Numerous experimental studies have shown the efficacy of mesenchymal stromal cells (MSCs) in ameliorating IRI and improving liver function during acute hepatic failure [7 –9]. MSCs are of particular interest in cell therapy due to their potential regenerative effects [10]. Furthermore, MSCs are immunoprivileged, which allows their transplantation across major histocompatibility barriers without eliciting an immune response in the recipient. MSCs are readily available from different sources such as bone marrow, adipose tissue, and umbilical cord and are easy to process [11,12]. However, their mode of action is not fully understood [13] and it is still under discussion whether MSCs differentiate and replace injured cells in designated organs or instead mediate their effects through paracrine communication [7,9]. Evidence in support of the latter comes from mice and rats treated with supernatant from MSC cultures in which a beneficial effect against hepatic IRI and a model of acute hepatic failure were found, respectively [8,14,15]. In addition to soluble factors, different studies revealed that extracellular vesicles (EVs) released by MSC play a major role in mediating the protective effects in rodent models of hepatic injury [16 –19]. Importantly, the beneficial effects of EV are not confined to those derived from MSC. Several studies have shown that EV derived from fully differentiated cells such as hepatocytes conferred similar protective effects on the liver against IRI [20,21]. So far, several biological factors responsible for the therapeutic effects of EV have been identified [19,22].
MSCs are characterized by a spindle-shaped fibroblast (FB)-like morphology and were originally referred to as colony-forming unit FBs [23]. FBs are mesenchymal cells with a significant role in tissue development, maintenance, and repair [24] and are found ubiquitously in almost all tissues with differentiation potential [25]. FB-derived EV can deliver various bioactive molecules. For example, EV derived from FB in cardiac tissue shuttle microRNA to neighboring target cells resulting in hypertrophy of cardiomyocytes [22] and EV derived from FB in liver tissue transfected with specific microRNA species decrease tumor growth in a rat model of cholangiocarcinoma [22,26].
Ichim et al. proposed the concept that FB may be more practical for cell therapy than MSC in some cases [24] and that this also applies to FB-derived vesicles. Notably, the protective effects of FB- and MSC-derived EV have not directly been compared so far. Furthermore, little is known about the protective effects of MSC and MSC-derived EV in the late phase (up to 72 h) of hepatic IRI [19,27].
In the present study, we assessed the effects of commercially available bone marrow-derived human MSC and MSC-derived EV compared to commercially available human adult dermal FB and FB-derived EV in the late phase of hepatic IRI in a murine model. We found that preemptive intravenous administration of MSC and MSC-derived EV before hepatic ischemia resulted in a significant reduction of hepatic damage in contrast to mice treated with FB or FB-derived EV. Taken together, our data show that FB or FB-derived EV do not protect against hepatic injury after IRI, whereas the sustained effects of MSC and MSC-derived EV hold promise as future therapies of IRI.
Materials and Methods
Cells and culture
Human bone marrow derived MSCs and adult normal human dermal FBs were obtained from PromoCell (Heidelberg, Germany). MSCs were plated and cultured in stem cell media from PromoCell (C-28009 with supplement and 1% penicillin/streptomycin) according to the recommended protocol. Differentiation to adipocytes, neurocytes, and osteoblasts was performed by the suppliers' instructions. Fixation and staining solutions were obtained from Morphisto (Frankfurt a. M., Germany). FBs were cultured in Fibroblast Growth Medium 2 (PromoCell; C-23020), with supplement and 1% penicillin/streptomycin. All cells were grown until confluence; no passage above seven was used in the experiments.
Characterization of cells by flow cytometry analysis
After incubation with FcBlock™ (BioScience; No. 564220) in Dulbecco's phosphate-buffered saline (PBS) with 2% fetal bovine serum (staining buffer) for 10 min at room temperature, staining of MSC and FB was performed by incubation for 20 min at 4°C using the following conjugated monoclonal antibodies: CD105-PE (BioLegend; AB_755958), CD90-FITC (BioLegend; AB_893429), CD73-PE (BioLegend; AB_2298698), CD45-PE (BioLegend; AB_314396), CD34-PE (Beckman Coulter; A07776), CD14 (Dako; R0864), and HLA-DR-PE (BioScience; No. 347401). Cells were analyzed on a CytoFLEX flow cytometer (Beckman Coulter), and the data were analyzed using CytExpert 2.2 software (Beckman Coulter).
Isolation and characterization of EV
EVs were isolated from the supernatant of confluent MSC and FB cultures by sequential ultracentrifugation [28]. In detail, after serum starvation for 48 h supernatant was first centrifuged at 1,600g for 20 min in 4°C. Thereafter, the supernatant was passed through a 0.45 μm filter (Hartenstein, Würzburg, Germany) and ultracentrifuged at 100,000g for 1 h at 4°C (Sorvall Discovery 90SE) to pellet EVs, which were then washed by resuspending in PBS and ultracentrifuged at 100,000g for 1 h at 4°C. The final pellet was resuspended in PBS and EVs counted and sized using NanoSight [Malvern Panalytical GmbH, Kassel, Germany; Software: Nanoparticle Tracking Analysis (NTA) Version 2.3 Build 0033]. Probes were either used directly for experiments or stored at −80°C.
Whole mounts of EV were qualified under the electron microscope (EM900; Zeiss, Oberkochen, Germany) by the Division of Electron Microscopy, Biocenter of the University of Würzburg using 2.5% glutaraldehyde, postfixed with 1% osmium tetroxide, and embedded, as well as stained, with uranyl acetate and lead citrate as previously described [29].
Characterization of cells and EV by western blot analysis
MSC, FB, and vesicles derived from both cell lines were analyzed for surface proteins expressed by MSC, microvesicles, and exosomes. Cells (2 × 105) were homogenized in ice-cold radio immune-precipitation assay lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 1 mM ethylenediaminetetraaceticacid, 0.25% sodium deoxycholate, 1% nonidet P-40 substitute in deionized water, and adjustment of pH to 7.4 with HCl), which contained freshly added protease inhibitor cocktail (5 mg/mL), sodium orthovanadate (200 mM), and sodium fluoride (200 mM) to inhibit protease and phosphatase activity. Next, samples were centrifuged at 14,000g and 4°C for 15 min and stored at −20°C.
Protein concentration was determined by Bio-Rad protein assay (based on Bradford's method). For cell lysates, equivalent amounts of protein were loaded on sodium dodecyl sulfate gels for electrophoresis. Lysates from EV contained an equivalent count of vesicles (5 × 108). After separation, proteins were transferred to polyvinylidene difluoride membrane (0.2 μ), blocked with 5% (w/v) of skim milk powder in tris-buffered saline (TBS)-T buffer [5 mM TRIS, 150 mM NaCl, 0.05% (w/v) Tween-20], and incubated overnight at 4°C with the respective primary antibodies 1:1,000: Rabbit monoclonal antibody CD9 (No. 13174), CD 90 (No. 13801), Histone H2A.X (No. 2595), Calnexin (CANX, No. 2679), signal transducer and activator of transcription 3 (STAT3, No. 30835), Phospho-STAT3 (Tyr705, No. 52075; Cell Signaling Technology Europe, Frankfurt, Germany); CD44 (ab51037), CD63 (ab134045), TSG101 (ab125011), Cytochrome C (CYC1, ab137757; Abcam, Cambridge, United Kingdom); β-actin (AC15); and GAPDH (G9294; Sigma-Aldrich, Munich, Germany) overnight at 4°C. The membrane was washed thrice with TBS and incubated with horseradish peroxidase (HRP) conjugated secondary antibody (1:10,000) for 1 h and then with HRP substrate (ECL substrate; BIO-RAD, Hercules) for 5 min. Proteins were visualized using ChemiDoc Touch Imaging System (Bio-Rad Laboratories GmbH, Munich, Germany).
Murine model of hepatic ischemia reperfusion
All animal studies were authorized by the local ethics committee (Government of Lower Franconia, Germany, project number DMS-2532-2-111), and mice were treated according to institutional and European Union guidelines. Female C57BL/6 mice at 10 weeks of age were purchased from Janvier Labs (Saint Berthevin, France) and were housed in groups of five animals per cage (Makrolon type III) with free access to food and water. The cages were placed in ventilated filter cabinets with humidity and temperature control (Scanbur, Denmark) within the specified pathogen free animal laboratory facility with artificial 12-h dark/12-h light cycle. Mice were anesthetized by intraperitoneal injection of Ketanest (100 mg/kg body weight) and Xylazin (8 mg/kg body weight). For analgesia Carprofen was used at 4 mg/kg body weight.
Surgical procedures
The animals were placed on a heating mat to maintain the body temperature, which was closely monitored by a rectal probe. After a midline laparotomy, MSC, FB (1 × 106 cells), MSC-EV, FB-EV (1 × 109 particles), or PBS (vehicle) in a 200 μL volume was administered through V. cava inferior injection. Five minutes later an atraumatic clip (Yasargil; Aesculap, Tuttlingen, Germany) was placed across the portal vein, hepatic artery, and bile duct just above the branching to the right lateral lobe resulting in deprivation of blood flow to ∼70% of the liver [30]. The clamp was held in place for 45 min. For the time of ischemia, the midline was closed with clips to maintain body temperature and avoid fluid loss by evaporation.
At various time points (24, 48, and 72 h postreperfusion) mice received anesthesia, blood was collected by puncture of the V. cava, and the liver was removed. Parts of all liver lobes were cut in small pieces and stored in RNA later (Qiagen, Hilden, Germany) or left in total and fixed in 3.7% buffered formalin.
Biochemical studies
Blood samples were collected at the end of each experiment and immediately centrifuged at 4°C. Enzyme activities of serum alanine transaminase (ALT) and aspartate transaminase (AST) were determined by standard clinical automated analyses (Roche, Basel, Switzerland), and the results were expressed in international units per liter. The procedures were performed according to the manufacturer's instructions.
Histopathology and immunohistochemistry
Paraffin embedded sections from mouse livers were cut into 1 μm thin slices and mounted onto object slides. After intensive drying, slides were deparaffinized and hydrated with distilled water followed by hematoxylin and eosin staining. Liver sections were scanned at magnification × 20 under a digital microscope. Single shots were merged to one picture; necrotic areas color coded by hand and calculated as the proportion of healthy parenchyma using the BZ-II analyzer software (BZ-H1AE; Keyence, Neu-Isenburg, Germany).
For immunohistochemistry, slides were further washed in TBS and a block of endogenous peroxidase using 3% hydrogen peroxide/PBS solution. Next, slides were soaked in citrate buffer for 40 min at 90°C. After cooling down, immunohistochemistry was performed using a primary antibody against Ki67 rabbit monoclonal antibody (ab16667, dilution 1:1,000; Abcam) and goat-anti-rabbit/HRP secondary antibody (dilution 1:1,000; Life Technologies, Thermo Fisher Scientific, Waltham, MA) along with TBS.
Whole tissue sections were scanned at ultraresolution on the digital microscope (BZ-9000; Keyence). Ki67 positive hepatocytes were quantitated by pixel counting using BZ-II analyzer software (BZ-H1AE; Keyence) followed by manual correction to exclude other Ki67 positive cell types in 10 high power fields (HPFs) per section. This was done in a standardized manner for each liver section: six HPF throughout the peripheral liver tissue and four within the central part. Necrotic areas were excluded from this analysis.
Reverse transcriptase quantitative polymerase chain reaction analysis
Total RNA was extracted from liver tissue with TRIzol, as recommended by the manufacturer (Invitrogen Life Technologies, Germany). Samples were quantified by Nanodrop ND-1000 Spectrophotometry (Thermo Fisher Scientific) and RNA integrity assessed using the Experion automated electrophoresis station (Bio-Rad Laboratories, Inc.).
RNA (0.7 μg) was reverse transcribed using the iScript cDNA Synthesis Kit from Bio-Rad. The cDNA synthesis of 1:2 diluted cDNA was performed by heating at 25°C for 5 min, at 42°C for 30 min, and at 85°C for 5 min. Quantitative PCR (qPCR) was performed with MESA GREEN qPCR MasterMix Kit for SYBR Green containing Meteor Taq hotstart polymerase (Eurogentic GmbH, Germany). qPCRs were performed on a CFX96 real-time PCR system (Bio-Rad) operated by CFX Manager Software (version 3.0). The cycler protocol was 5 min at 95°C, 40 cycles of 15 s at 95°C, 60 s at 60°C, and 5 min at 72°C. Postamplification melting curves were controlled to exclude primer-dimer artifacts and contaminations.
mRNA expression levels of interleukin beta (Il1β), interleukin 6 (Il6), intercellular adhesion molecule 1 (Icam1), vascular cell adhesion molecule 1 (Vcam1), high mobility group box 1 (Hmgb1), and heme oxygenase (Ho1) were normalized to those of the reference gene B2m (beta-2-microglobulin), calculated with the ΔΔCq method and displayed relative to PBS control. The primer pairs used are listed in Supplementary Table S1.
Statistical analysis
Data are presented as mean ± standard error of the mean, with each dot representing individual animals. Statistical analysis was performed using Prism 8 software (GraphPad Software, San Diego, CA). Differences between groups were assessed using Mann–Whitney U test. Results with P values <0.05 were considered as statistically significant.
Results
Characterization of MSC and FB in vitro
MSCs were commercially acquired and cultured using optimized medium for cell proliferation provided by the manufacturer. MSCs showed a FB-like morphology, were plastic adherent, and expressed CD105, CD90, and CD73 (>95% positivity) but lacked the expression of CD45, CD34, CD14, and HLA-DR (<1% negativity) in flow cytometry phenotyping (Fig. 1A). Flow cytometry analysis of commercially purchased FB revealed an identical surface marker profile showing positive expression of CD105, CD90, and CD73 (>95%), while lacking the expression of CD14, CD34, CD45, CD105, and HLA-DR (<1% negativity) (Fig. 1B).
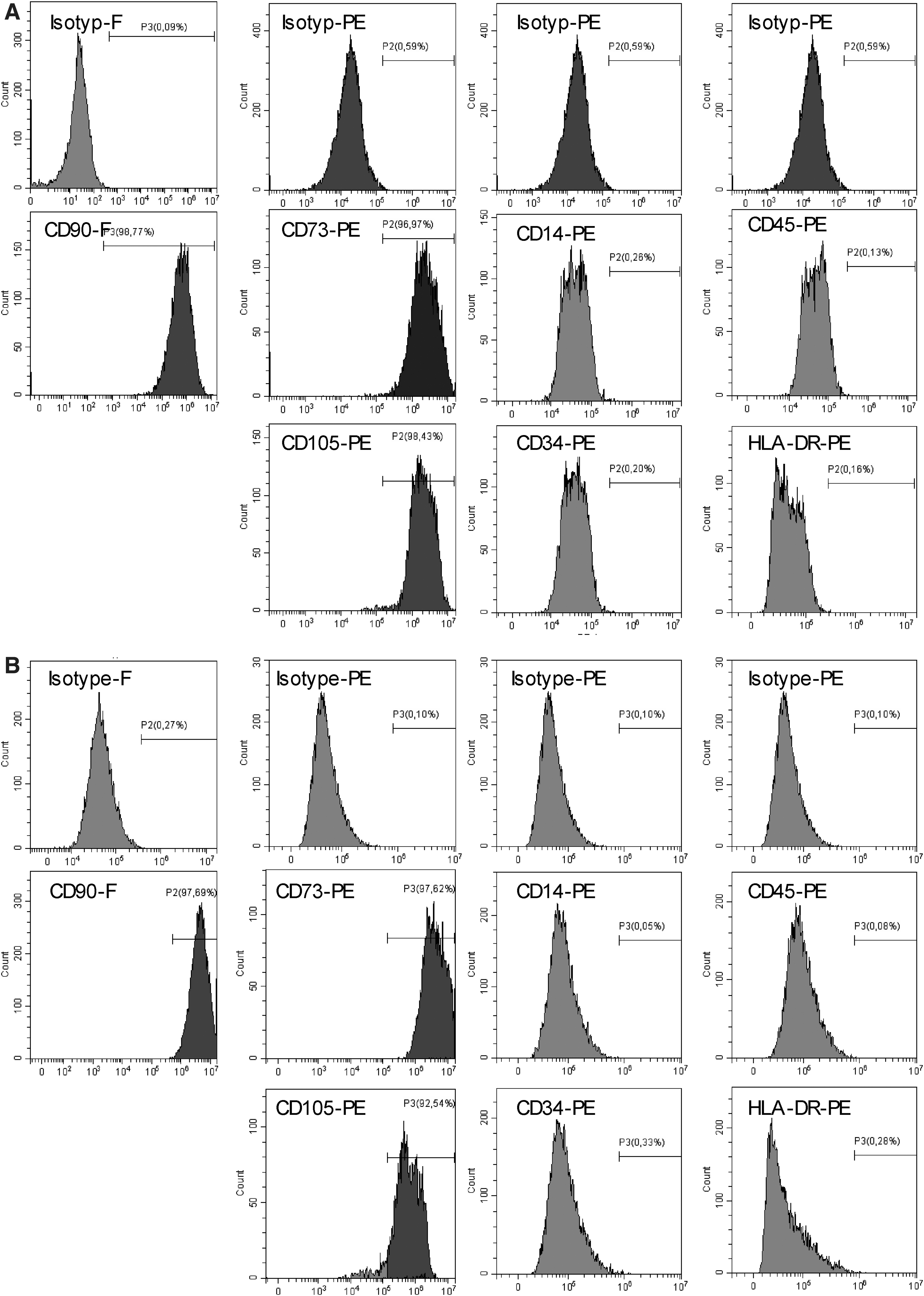
Immunophenotype of bone marrow-derived human MSC and normal human dermal FBs. Representative flow cytometry analyses of defining positive MSC markers CD105, CD90, and CD73 and negative markers CD45, CD34, CD14, and HLA-DR on MSC and FB cells.
The MSC could be differentiated into adipocytes, neurons, and osteoblasts by culture (Fig. 2A). Therefore, we used MSC in this study that fulfilled a set of minimal criteria proposed by the International Society of Cellular Therapy to determine bone marrow-derived MSC [31]. FB showed only a single-lineage differentiation potential into neuron-like cells under the same conditions (Fig. 2E).
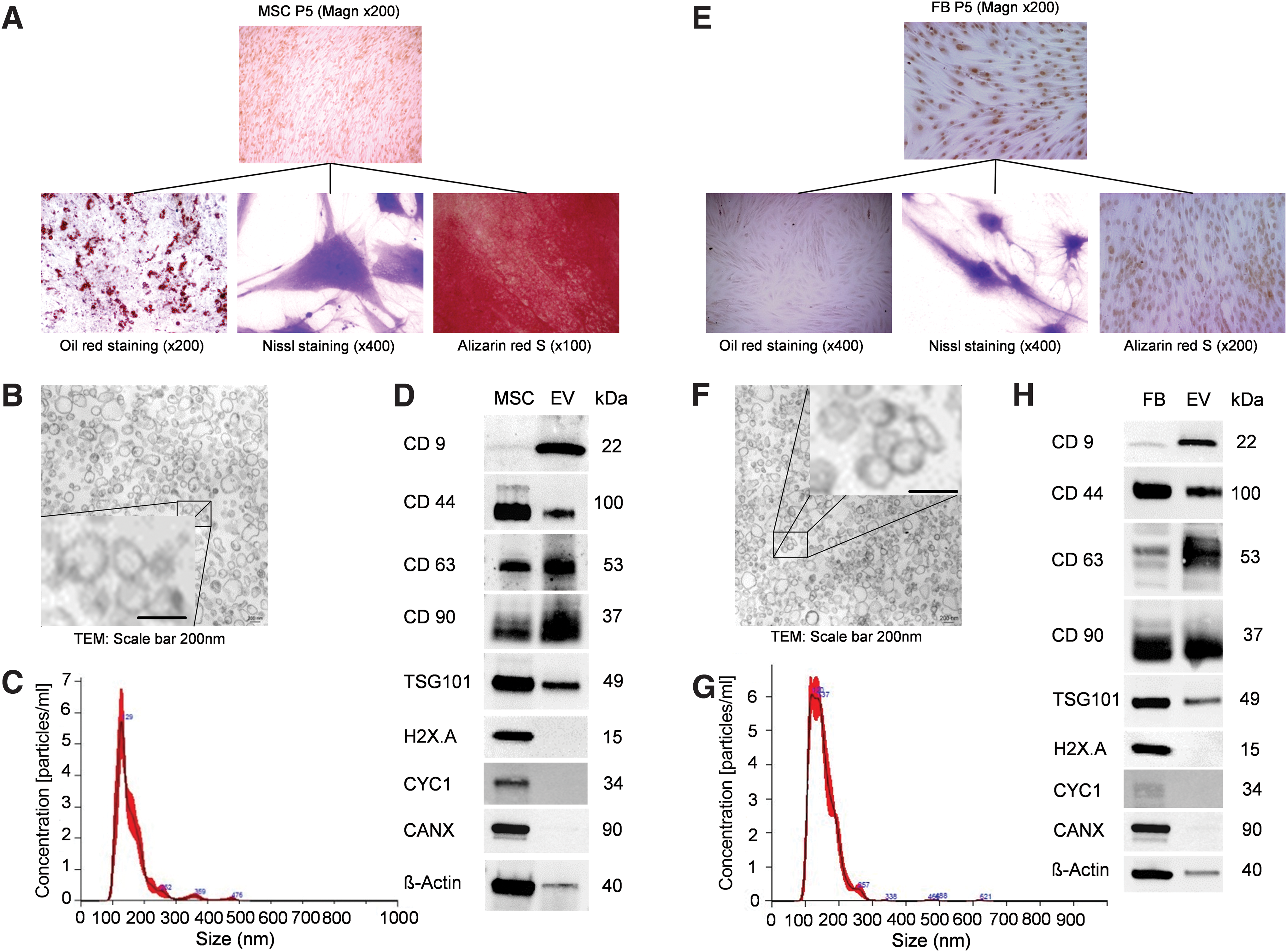
Characterization of MSC, FB, and isolated vesicles.
Isolation and characterization of MSC-derived and FB-derived EV
EVs were isolated from serum depleted culture media of MSC and FB using an ultracentrifugation-based protocol [28] and presented as membrane-bound spherical particles (Fig. 2B, F). Nanoparticle analysis revealed a comparable size distribution for MSC-derived and FB-derived EV (respective mean sizes of 160 ± 57 nm vs. 153 ± 42 nm and peak sizes of 129 nm vs. 120 nm) (Fig. 2C, G). The protein composition of MSC-derived and FB-derived EV was analyzed by immunoblotting and revealed plasma membrane proteins (CD44, CD90), membrane proteins commonly found on exosomes (CD9, CD63), and an endosome-associated protein (TSG101) (Fig. 2D, H) [32,33]. In addition, we excluded contaminations of EV with proteins derived from subcellular compartments, for example, nucleus (H2X.A), mitochondria (cytochrome C), and endoplasmic reticulum or Golgi apparatus (calnexin). These results confirmed that EV derived from MSC and FB displayed typical features of EV in terms of size and protein marker profile and therefore fulfilled the minimum criteria proposed by the International Society of Extracellular Vesicles [34].
MSC and MSC-EV reduce hepatic IRI in mice
The modulatory effects mediated by MSC, FB, MSC-derived, and FB-derived EV were evaluated in a murine model of warm hepatic IRI. MSC, EV, or PBS (vehicle) was injected into the V. cava inferior 5 min before 45 min of ischemia (Fig. 3A). Hepatic injury was assessed by consecutive measurements of liver serum transaminases (AST and ALT) and histopathological evaluation of parenchymal necrosis 24, 48, and 72 h after IRI. MSC-derived EV significantly reduced ALT (P < 0.05) and AST (P < 0.05) levels after reperfusion in contrast to FB-derived EV and vehicle control 24 h after reperfusion (Fig. 3B, C). This effect could also be seen in animals treated with MSC, as ALT (P < 0.05) and AST (P < 0.05) were significantly reduced compared to animals treated with FB or vehicle control (Fig. 3D, E).
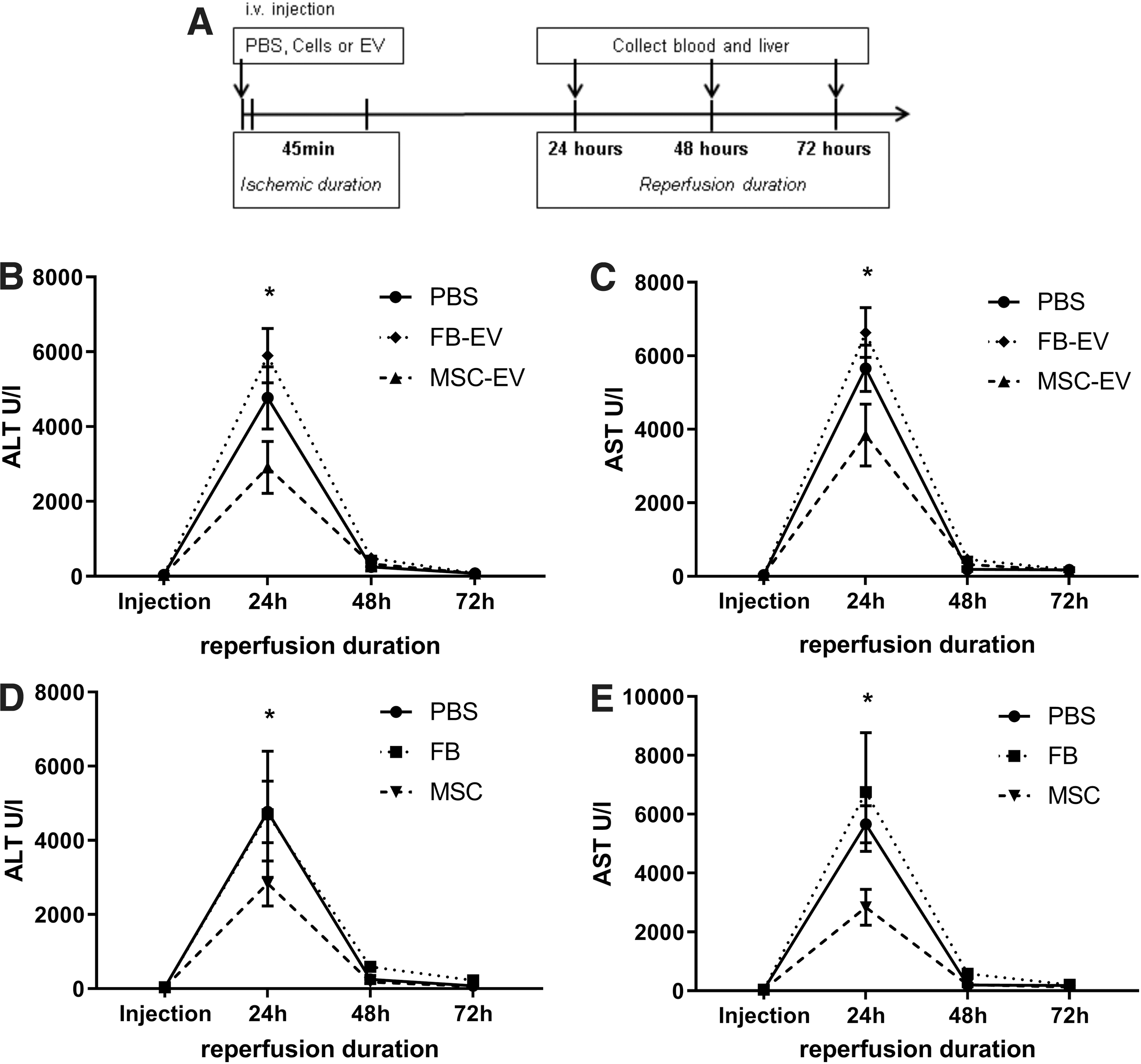
MSC and MSC-derived EV decrease ALT and AST levels after hepatic IRI.
The extent of necrosis was determined in the median liver lobe (Fig. 4). Control animals receiving PBS (vehicle) developed severe necrosis of liver parenchyma of 46.8 ± 1.9% 24 h after IRI (Fig. 5A). The beneficial effect of MSC and MSC-derived EV on the extent of tissue necrosis (28.4 ± 15.7% and 27.6 ± 14.1%, respectively, at 24 h after reperfusion) was detectable at each time point after IRI (Fig. 5A). In contrast, FB and FB-derived EV failed to reduce necrosis, as well as vehicle control.
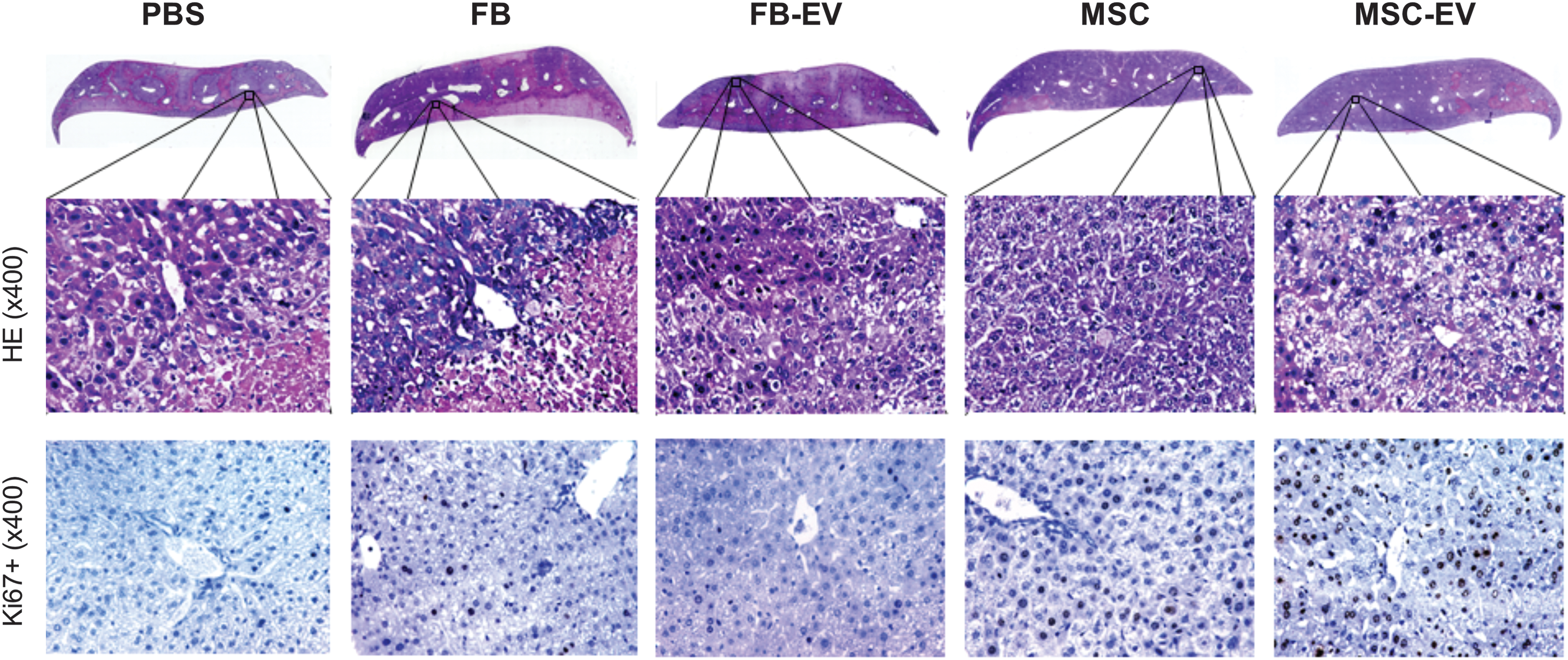
MSC and MSC-EV reduce liver necrosis and accelerate liver regeneration after hepatic IRI. H&E staining and immunohistochemistry staining for Ki67 in representative liver sections of mice treated with MSC, FB, MSC-EV, FB-EV, or PBS 48 h after hepatic IRI. For H&E staining full sections and representative HPFs are shown. In immunostaining for Ki67 brown cells in HPF are considered positive. H&E, hematoxylin and eosin; HPF, high power field. Color images are available online.
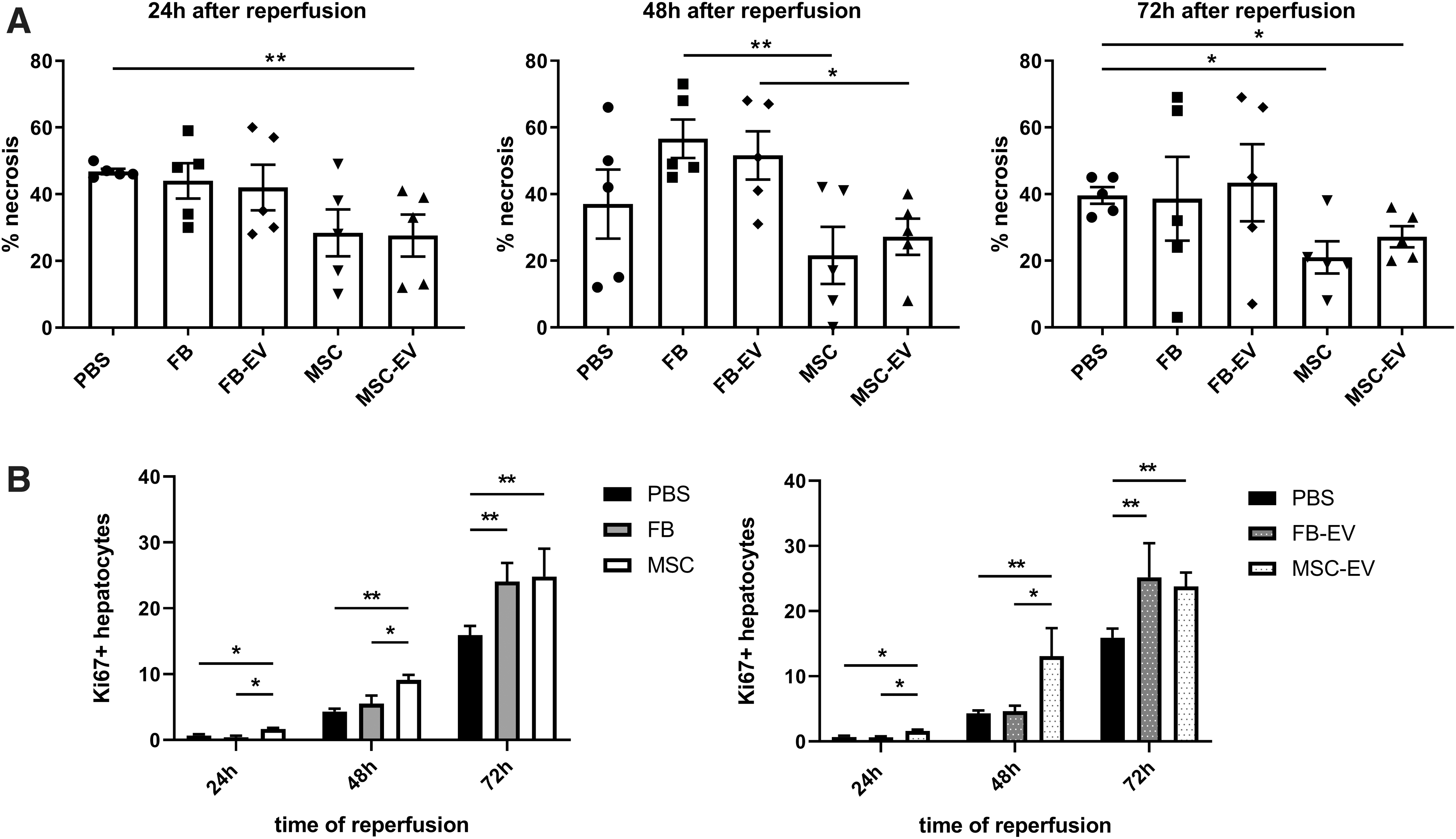
MSC-EV improves resolution of tissue necrosis and liver regeneration after hepatic IRI. Whole liver section scans (Keyence microscope) were quantified for necrotic areas and Ki67 staining using BZ-II-Analyzer software.
MSC and MSC-derived EV promote early hepatocyte proliferation after hepatic IRI
Next, we analyzed the pro-regenerative effects of MSC, MSC-derived EV, FB, and FB-derived EV by staining liver sections with the proliferation marker Ki67 (Fig. 5B). We found a significantly stronger increase in Ki67-positive hepatocytes in animals treated with MSC 24, 48, and 72 h after ischemia and reperfusion (24 h: P < 0.05 vs. FB and PBS, respectively; 48 h: P < 0.05 vs. FB, P < 0.01 vs. PBS; 72 h: P < 0.01 vs. PBS) and MSC-derived EV (24 h: P < 0.05 vs. FB-EV and PBS, respectively; 48 h: P < 0.05 vs. FB-EV, P < 0.01 vs. PBS; 72 h: P < 0.01 vs. PBS) compared to animals treated with FB, FB-derived EV, and vehicle control (PBS). Interestingly, the amount of K67-positive cells 72 h after reperfusion was comparable in all treatment groups (MSC 24.8 ± 4.2%, FB 24.1 ± 2.8%; MSC-derived EV 23.8 ± 2.2%, FB-derived EV 25.2 ± 5.2%). However, the PBS-control group showed significantly less Ki67-positive hepatocytes compared to treatment groups at 72 h after reperfusion (P < 0.01 vs. FB, FB-derived EV, MSC, and MSC-derived EV) (Fig. 5B).
MSC and MSC-derived EV change the intrahepatic transcription of inflammation-associated genes after hepatic IRI
The direct effects of MSC, MSC-derived EV, FB, and FB-derived EV on inflammation-associated genes in liver tissue were evaluated 24 h after IRI by reverse transcriptase quantitative polymerase chain reaction. The mRNA expression of the late-acting inflammatory mediator Hmgb1 was significantly reduced in groups receiving MSC or MSC-derived EV (Fig. 6A). Similar results were obtained for Icam1 (Fig. 6A). The most pronounced decrease in expression of inflammation-associated genes was noted for Ho1 mRNA, which was decreased >3-fold by MSC and MSC-derived EV (P < 0.01) compared with animals receiving PBS (Fig. 6A). However, it must be stated that the mRNA expression of Icam1 and Ho1 was also reduced in animals receiving FB-derived EV, although statistical evaluation remained below the level of significance. These data indicate an anti-inflammatory effect of MSC, MSC-derived EV, and to a much lesser degree of FB-derived EV. In comparison to the PBS group, a trend toward reduced mRNA expression levels of the pro-inflammatory cytokine Il1β, as well as Vcam1, was seen in groups treated with MSC and MSC-derived EV (data not shown).
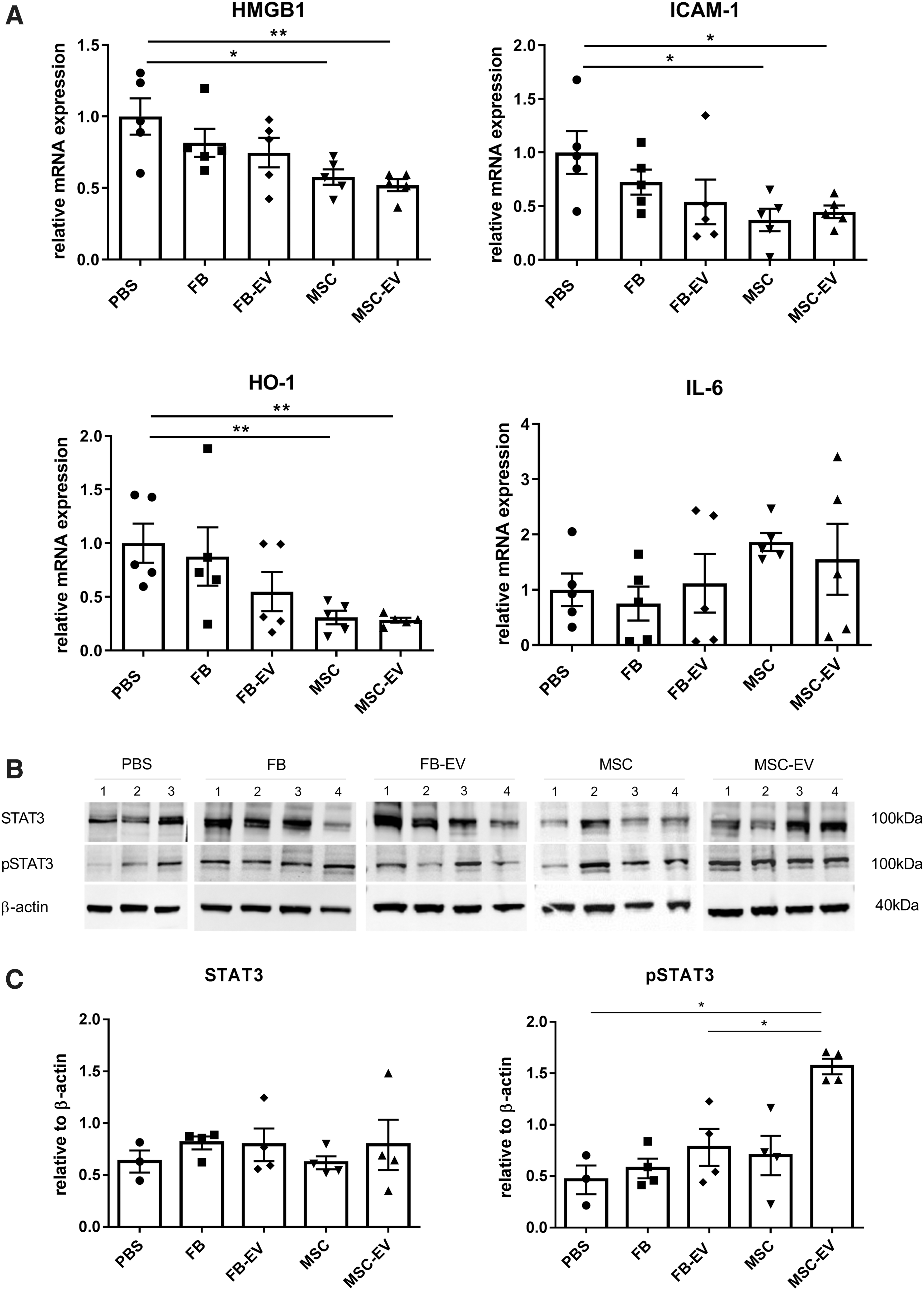
MSC and MSC-derived EV change mRNA and protein expression of inflammation and regeneration associated genes. mRNA and protein expression were assessed in liver tissue from mice of each treatment group 24 h after reperfusion.
The multifunctional cytokine IL-6, which is involved in triggering hepatocyte proliferation during liver regeneration after IRI showed a trend toward higher expression levels in groups treated with MSC and MSC-derived EV. Phosphorylated and thereby activated STAT3, a downstream target of IL-6 contributing to hepatocyte proliferation, increased significantly by MSC-derived EV (Fig. 6B, C). FB had a markedly lesser effect on expression of the analyzed genes and proteins. However, our data underscore a repression of inflammation-related genes in liver tissue mediated by MSC and MSC-derived EV during the early phase of IRI.
Discussion
In the present study, we demonstrated that MSC and MSC-derived EV reduced tissue injury significantly and accelerated organ regeneration in a murine model of hepatic IRI in the resolution phase up to 72 h after reperfusion. Livers treated with MSC or MSC-derived EV showed a decrease in hepatic necrosis together with lower hepatic expression of inflammation-associated mediators and an increased number of Ki67-positive hepatocytes. This effect appeared to be more pronounced with MSC and MSC-derived EV because commercially available FB and FB-derived EV had no effect on organ protection and early initiation of liver regeneration after hepatic IRI. While previous studies have already demonstrated beneficial effects of MSC in the early phase of hepatic IRI up to 24 h, the present study is the first to show effects of MSC-derived EV in comparison to FB-derived EV during the resolution phase of hepatic IRI.
MSC- and FB-derived EV show comparable morphological characteristics
In general, our characterization data for the EV derived from MSC or FB show typical features of EV in terms of size and protein marker profile and, therefore, fulfill the minimum criteria proposed by the International Society of Extracellular Vesicles [34]. EVs, secreted by many different cell types as an integral component of intercellular communication [32], represent a heterogeneous population of membrane-enclosed particles consisting of exosomes (40–120 nm diameter), EV (50–1,000 nm diameter), and apoptotic bodies (500–2,000 nm diameter). In addition to differences in size, exosomes and EV are different in their biological function and in the mechanism by which they are secreted by cells [28,35]. While the biogenesis of exosomes initiates with endocytosis and the formation of early endosomes maturating to multivesicular bodies which finally release their content as exosomes, EVs directly shed from the plasma membrane by a process referred to as budding [32]. We compared MSC-derived EV and FB-derived EV and found no differences in morphology, size distribution, and protein markers. The detected protein marker profile was comparable to that previously described for exosomes [32,36,37]. However, the mean size of 160 ± 57 nm for the MSC-derived EV and 153 ± 42 nm for the FB-derived EV both slightly exceeded the size range of 40–120 nm considered characteristic for exosomes. At present, there is no molecular marker to strictly separate endosome-derived exosomes from small-sized microvesicles that have budded from the plasma membrane [38,39]. However, both MSC- and FB-derived EV displayed comparable features.
MSC and MSC-EV but not FB and FB-EV reduce hepatic IRI in mice
Our data show that MSC-derived EV significantly reduced the extent of liver necrosis and promoted liver regeneration in a rodent model of hepatic IRI. We obtained similar results from animals treated with MSC, whereas administration of FB and FB-derived EV had no detectable effect on liver injury. However, both FB and FB-derived EV increased hepatocyte proliferation at a later time point and FB-derived EV lowered the expression of certain inflammation associated genes in hepatic tissue 24 h after reperfusion. Previously, EVs derived from differentiated cells such as hepatocytes have been recognized as important players for liver regeneration after hepatic IRI [20]. The results we obtained with MSC-derived EV and FB-derived EV on liver tissue repair after hepatic IRI could be explained by differences in the content of bioactive molecules. This might be due to a different gene expression in MSC and FB [40] resulting in a stronger regenerative potential in MSC-derived EV compared to FB-derived EV [41]. The use of EV in vivo is generally considered safe in contrast to whole MSC [42,43], which has been demonstrated with spontaneous malignant transformation in vitro [42,43]. Consequently, a potential risk of neoplastic transformation by MSC in vivo may exist and needs to be examined. In addition, EVs endure storage at −80°C without measurable changes in biological function and thereby would be readily available in the clinical setting of liver resection or organ transplantation [44].
Hepatic IRI is a pathophysiologic phenomenon, in which hypoxic liver damage is enhanced after the restoration of blood flow and reoxygenation of the liver. This phenomenon exhibits a triphase pattern of injury followed by liver regeneration: an early phase that occurs immediately after reperfusion, an intermediate phase within 2–6 h of reperfusion, and a late phase that occurs from 6 to 48 h after ischemia. The initial phase is characterized by the pH paradox, in which the return from acidotic to normal pH after reperfusion leads to mitochondrial dysfunction and cell death. Subsequently, the intermediate phase reflects oxidative stress and the uncontrolled release of different cytokines. As a result, adhesion molecules are upregulated promoting the tethering and diapedesis of neutrophils into the liver parenchyma during the late phase [45]. These neutrophils then induce hepatocyte necrosis through the release of cytotoxic proteases and oxygen-derived radicals [46] and reduce tissue regeneration by the resulting inflammation. Published studies have focused mostly on the intermediate phase [19,27], describing damage control rather than organ regeneration by MSC-derived EV. An increase in hepatocyte viability and proliferation rate could be shown in vitro [17,19] but was missing in vivo. In this study, animals that received a preemptive injection of MSC and MSC-derived EV immediately before the ischemia presented earlier hepatocyte proliferation after hepatic IRI.
Anti-inflammatory effects and induction of IL-6-dependent regeneration may explain the beneficial effects following MSC and MSC-derived EV
In this study, expression of Hmgb1 mRNA in all cells within the liver tissue was significantly reduced in both MSC and MSC-derived EV treated animals 24 h after reperfusion. Furthermore, MSC and MSC-derived EV lowered Hmgb1 and Il1β mRNA expression in liver tissue. Finally, animals treated with MSC and MSC-derived EV showed increased levels of hepatic Il6. Until now, the triggering factor for liver regeneration after IRI remains not completely understood [47]. One effect caused by IRI and important for organ regeneration is the release of HMGB-1 into the extracellular space, where it acts as an essential damage-associated molecular pattern molecule activating pro-inflammatory signaling and cytokine release through Toll-like receptors [48]. In a rodent model of hepatic IRI, administration of neutralizing antibodies against HMGB-1 protected liver tissue from reperfusion injury [49]. Directly reducing HMGB-1 release by downregulation of nuclear Hmgb1 expression through siRNA significantly protected the liver against warm IRI [50]. The expression of the adhesion molecule ICAM-1 is induced by IL-1 on hepatocytes [5,51] and improves the transmigration of neutrophils, whereas antibodies against ICAM-1 prevent neutrophil extravasation and substantially decrease liver injury [52]. ICAM-1 is also expressed constitutively on vascular endothelial cells such as liver sinus endothelial cells and upregulated during inflammatory responses [53]. Targeting ICAM-1 could therefore serve to protect the liver during inflammatory stress. Consequently, attenuation of inflammatory responses associated with tissue injury could be a mechanism for the beneficial effects of MSC and MSC-derived EV during hepatic IRI seen in our experiments. As a result, Ho1 mRNA expression was significantly lower in mice treated with MSC and MSC-derived vesicles.
IL-6 is a well-known pro-inflammatory cytokine released within minutes of reperfusion [54]. However, it is also known as one of the key players in liver regeneration. In rodent studies, IL-6 knockout mice showed impaired liver regeneration and reconstitution of IL-6 signaling with recombinant IL-6 accelerated restoration of functional liver mass after major hepatic resection [55]. IL-6 binds to gp130 and activates downstream targets such as STAT3, which promotes cell-cycle entry through cyclin and cyclin dependent kinase [45,55]. Consequently, phosphorylated STAT3 expression was significantly increased in animals treated with MSC-derived EV, driving accelerated cell proliferation in the liver. This suggests a potential mechanism for the beneficial effects we observed following the administration of MSC-derived EV.
In conclusion, our study supports the use of MSC-derived EV as a preemptive strategy to alleviate hepatic IRI and improve liver regeneration following hepatic IRI. In this context, the regenerative capacity of liver tissue plays a pivotal role in liver transplantation and resection surgery. Further studies are underway to explore the therapeutic factors located within MSC-derived EV essential for mediating anti-inflammatory and pro-regenerative effects.
Footnotes
Acknowledgments
The authors thank Prof. Dr. J.W. Lee, University of California, San Francisco, CA for methodical advice, Sabine Gahn and Manuela Hofmann for technical assistance, Andrea Prappacher for the management of the mouse colonies, the Laboratory of Prof. Dr. Krohne, Head of the Division of Electron Microscopy, Biocenter of the University of Würzburg for providing transmission electron microscopy, and the Department for Functional Materials in Medicine and Dentistry, University of Würzburg, Germany for providing nanoparticle tracking analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Interdisciplinary Center of Clinical Research (grant no. Z-3/67), Clinical Faculty of the University of Würzburg.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.