
Abstract
Chronic critical limb ischemia (CLI) represents a clinical end stage of peripheral arterial disease. Many CLI patients are ineligible for conventional revascularization therapies; thus, it is urgent to explore an alternative strategy to rescue the ischemic limb. Recent stem cell studies have greatly developed the field of therapeutic angiogenesis, which aims to significantly improve the limb blood supply. In our study, bone marrow mesenchymal stem cells (BMMSCs) served as the control to evaluate the function of umbilical cord mesenchymal stem cells (UCMSCs) in enhancing angiogenesis. We compared gene expression between BMMSCs and UCMSCs, and a bioinformatics analysis indicated that both UCMSCs and BMMSCs could stimulate angiogenesis and angiogenesis-related factors were upregulated in UCMSCs. In vitro assays indicated that both BMMSCs and UCMSCs promoted human umbilical vein endothelial cell proliferation, migration, and tube formation, and the effects of UCMSCs were more obvious. Consistent with in vitro results, both UCMSCs and BMMSCs improved the limb blood supply in a mouse model of hind limb ischemia, in which UCMSCs promoted angiogenesis more significantly. Finally, we found that activation of ERK and PI3K-Akt pathways might be the mechanism by which UCMSCs promote angiogenesis. These results indicate that UCMSCs play an important role in therapeutic angiogenesis to improve limb blood perfusion.
Introduction
Critical limb ischemia (CLI) is defined as ischemic rest pain, tissue loss, or gangrene in the presence of peripheral artery disease (PAD) and hypoperfusion of the lower extremity [1]. CLI, which is commonly classified to Fontaine III and IV, represents the most severe stage of PAD and is associated with significant morbidity and high mortality. The prevalence of PAD, especially CLI, has risen dramatically worldwide [2]. However, despite the technological advances to manage CLI with minimally invasive technologies, current outcomes remain poor, especially in CLI patients [3].
To avoid amputation and rescue the affected limb, the main objective in treating CLI is to increase blood circulation in the hypoperfusion tissue. The goal of restoring blood flow can be achieved by increasing the number of vessels. The use of pharmacological agents to induce new blood vessel growth for the treatment or prevention of pathological clinical conditions has been called therapeutic angiogenesis [4]. Mesenchymal stem cells (MSCs), with characteristics of self-renewal and multilineage differentiation, could help tissue regeneration [5 –7], indicating MSC to be a potential candidate in therapeutic angiogenesis. Increasing studies have revealed the distinction of MSCs in promoting angiogenesis and improving the blood supply in an ischemic limb in vivo [8 –11].
BMMSCs have been validated to be effective in promoting angiogenesis, and early phase clinical trials demonstrated that BMMSC therapies could be safe, feasible, and potentially effective for improving blood perfusion of ischemic limbs [12,13]. Meanwhile, UCMSCs have shown the ability to differentiate into three germ layers, promote tissue repair, and modulate immune responses [14]. UCMSCs were also shown to promote angiogenesis and improve blood supply [10,15,16], and clinical trials revealed their safety and effectiveness at the early phase [17]. Our research used BMMSCs as a control to evaluate the function of UCMSCs in regulating angiogenesis and hind limb ischemia in a mouse model, which aimed to provide a further experimental basis for clinical application of UCMSCs in improving limb ischemia.
Materials and Methods
Cell culture
Human UCMSCs and BMMSCs were purchased from Cyagen (Suzhou, China) and cultured in human MSC complete culture medium with 10% FBS (Cyagen). Human umbilical vein endothelial cells (HUVECs) were purchased from Science Cell (Carlsbad, CA) and cultured in extracellular matrix (ECM) medium with 5% FBS (Science Cell). MSCs and HUVECs at passages 3–8 were used for experiments.
mRNA microarray detection
BMMSCs and UCMSCs were seeded at 2 × 105 cells per cm2 and incubated for 1 day. The attached cells were washed with phosphate-buffered saline (PBS), and total RNA from MSCs was extracted with TRIzol reagent (Invitrogen, Carlsbad, CA). mRNA microarray detection (Aksomics, Shanghai, China) was performed with the samples (BMMSC, n = 3; UCMSC, n = 3).
Preparation of conditioned medium
MSCs were plated at 2.5 × 104 cells per cm2 and incubated for 1 day. The attached cells were washed three times with PBS, and the medium was replaced with serum-free DMEM to generate BMMSC-CM or UCMSC-CM. The conditioned medium (CM) was collected after 48 h of culture, centrifuged at 1500 rpm for 10 min, and filtered through a 0.2-μm filter. The CM was concentrated 30-fold compared with its original concentration by Amicon® Ultra-15 centrifugal filter units (3 kDa; Millipore, Darmstadt, Germany). The concentrated CM was stored at −80°C for future use [18].
Cell proliferation assay
HUVECs were seeded in a 96-well plate (104 cells/well) and cultured in ECM medium containing 5% FBS. The following day, after washing with PBS, the HUVECs were incubated in MSC-CM for 48 h, which was followed by the cell proliferation assay using a CCK-8 kit (Dojindo, Kumamoto, Japan).
Tube formation assay
HUVEC tube formation on Matrigel was performed as previously described [16]. A 150-μL aliquot of Matrigel (BD Biosciences, San Jose, CA) was added to a chilled 48-well plate and incubated at 37°C for 30 min. HUVECs (3 × 104 cells) were suspended in 500 μL of the MSC-CM and in serum-free DMEM as a control, which were added to the solidified Matrigel. After incubation at 37°C in a 5% CO2 chamber, the tube morphological changes were observed under an inverted microscope (Zeiss, Germany). ImageJ software (NIH; Bethesda) was used to analyze the tube formation, and the master junctions, total mesh areas, and total segment lengths were statistically analyzed.
Wound healing assay
HUVECs were seeded in 12-well plates (2 × 105 cells/well) and incubated in ECM with 5% FBS for 12 h. After washing twice with PBS, they were incubated in ECM with 1% FBS for 24 h. After the cells were grown to confluence, straight lines were scratched using a 10-μL pipette tip. Cells were then cultured in MSC-CM, using serum-free DMEM as a control. Images were taken by an inverted phase-contrast microscope (Zeiss) at 0 and 24 h.
Transwell migration assay
Assays were performed in 24-well cell culture chambers (8.0-mm pores, BD Biosciences). HUVECs were seeded in six-well plates (3 × 105 cells/well), incubated in ECM with 5% FBS for 12 h, followed by washing twice with PBS, and then incubating with MSC-CM and serum-free DMEM as control for 24 h. HUVECs were then plated into the upper chamber at 5 × 104 cells/well in 0.2 mL of serum-free DMEM, and DMEM with 10% FBS was placed in the lower chamber. After 24 h, the cells on the lower surface of the upper chamber were fixed, stained with crystal violet, visualized, and counted, as previously described [19].
Reverse transcription and quantitative polymerase chain reaction assay
Total RNA was extracted from MSCs with TRIzol reagent (Invitrogen) according to the manufacturer's instructions. cDNA was synthesized using a PrimeScript™ RT reagent kit with gDNA Eraser (TaKaRa, Tokyo, Japan). The SYBR Green reverse transcription and quantitative polymerase chain reaction (RT-qPCR) was conducted using SYBR Green I Master (Roche, Basel, Switzerland) in a 10 μL reaction volume on a LightCycler 480 instrument (Roche) according to the manufacturer's instructions. Sequences of primer pairs (GenePharma, Suzhou, China) are detailed in Supplementary Table S1. The expression was calculated normalized to GAPDH (delta CT) and then compared with controls (delta CT).
Animals
Six-week-old male BALB/c nude mice were purchased from the Model Animal Research Center of Nanjing University (Nanjing, China). All animal procedures were approved by the Animal Care and Use Committee of the First Affiliated Hospital of Sun Yat-sen University. Unilateral, hind limb ischemia animal models were established according to previous studies [8,20]. Briefly, mice were anesthetized by isoflurane, and then 7–0 silk sutures were knotted around both the proximal and distal femoral arteries. The right hind limb was the control. Within the first 6 h after resection, 1 × 106 MSCs in 100 μL of PBS (MSC groups) were injected into the adductor muscle of the ischemic hind limb and 100 μL of PBS (PBS group) was the control (n = 5 per group). Measurements of hind limb blood flow were performed using PERIMED PSI-ZR (PERIMED, Sweden) before operation and at days 0, 14, and 21 postinjection. The blood perfusion ratio was defined as the ratio of blood perfusion in ischemic hind limbs to nonischemic hind limbs [16]. Mice were anesthetized and sacrificed by intraperitoneally injecting an overdose of pentobarbital at day 21 postinjection, and the adductor muscle was removed and stored in liquid nitrogen.
Immunofluorescence assay
Frozen muscles were embedded in optimum cutting temperature (OCT) medium compound, and cryosections (3–5 μm) were placed on positively charged glass slides and fixed in acetone (4°C) for 30 min; they were then blocked in 10% goat serum (Solarbio, Beijing, China) for 5–10 min. The tissue samples were then incubated with anti-CD31 antibody (1:400; Abcam, Cambridge, CA) overnight at 4°C. The next day, after gently washing for 10 min three times with PBS, tissues were incubated with secondary antibody (1:400; RD, Minneapolis, MN) for 60 min at room temperature. After washing with PBS, the slides with tissues were dried and mounted with ProLong™ Gold Antifade Mountant with DAPI (Invitrogen). Images were recorded by an inverted fluorescence microscope (Zeiss).
Immunohistochemical assay
The mouse muscle sections (3 μm) were processed with hydrogen peroxide blocker and incubated with a primary antibody: anti-CD31 antibody (Abcam). Following overnight incubation and washing, muscle sections were incubated with the secondary antibody (Proteintech, Rosemont, IL) and developed using 3,3-diaminobenzindine reagent (DAB; Solarbio). Hematoxylin was used for nucleus staining. The muscle sections were observed and imaged using an Olympus BX51 microscope (Olympus, Tokyo, Japan).
Western blotting
The HUVEC total protein was extracted using RIPA lysis buffer (Beyotime, Shanghai, China) with 1% PMSF (Beyotime), and the protein concentration was measured using a BCA kit (Beyotime). Proteins were separated by SDS-PAGE, transferred to PVDF membranes (Millipore, Bedford, MA), and incubated with primary antibodies. The primary antibodies were anti-GAPDH (Proteintech), anti-MEK1/2 (CST; Danvers, MA), anti-MEK1/2 (phospho Ser 217/221; CST), anti-ERK1/2 (CST), anti-ERK1/2 (phospho Thr202/Tyr204; CST), anti-Akt (CST), anti-Akt (phospho Thr308; Abcam), and anti-Akt (phospho Ser473; CST). IgG-horseradish peroxidase secondary antibody (Proteintech) was used. Protein bands were exposed to the ECL luminol reagent (Millipore) using an Amersham Imager 600 instrument (GE, Fairfield, CT).
Statistical analyses
Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses were performed using the online database DAVID 6.8 (
Results
BMMSCs and UCMSCs could promote angiogenesis in GO analysis
To identify the differentially expressed genes (DEGs) between BMMSCs and UCMSCs, we performed mRNA microarray detection using MSC specimens (BMMSC, n = 3; UCMSC, n = 3). The screening standard for DEGs was defined as P < 0.05, log (fold change) >2. We found that 394 DEGs were upregulated in BMMSCs compared with UCMSCs, while 355 DEGs were upregulated in UCMSCs compared with BMMSCs (Fig. 1A).

BMMSCs and UCMSCs promote angiogenesis in GO analysis.
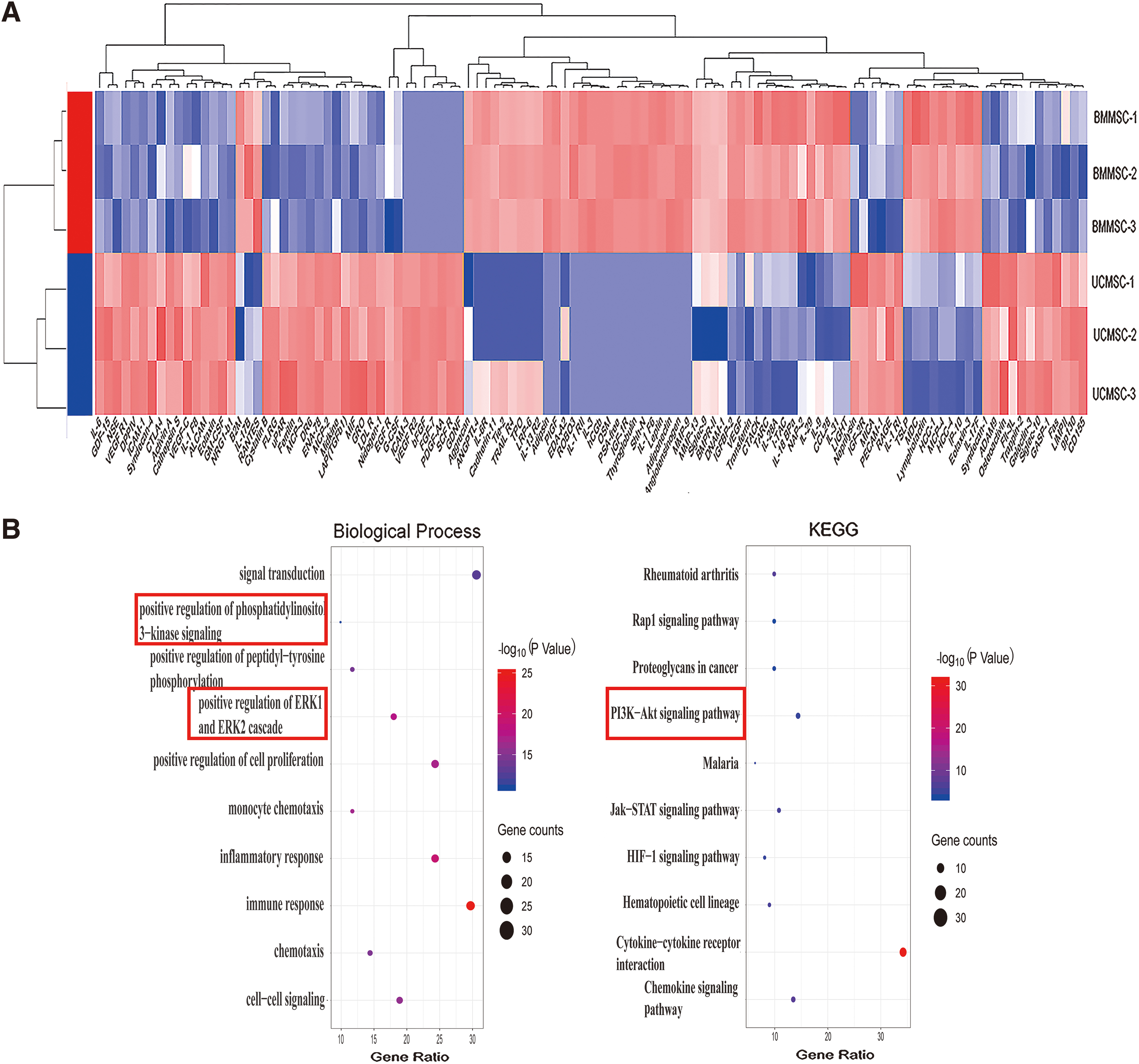
To further explore the biological functions of these DEGs, GO and KEGG analyses were conducted. Fifteen items were enriched in the biological process classification of GO analysis (FDR <0.05, Supplementary Table S2), among which two items were related to angiogenesis: vasculogenesis and positive regulation of angiogenesis (red box, Fig. 1B). Items related to positive regulation of endothelial cell migration and positive regulation of cell proliferation were also enriched (green box, Fig. 1B). As reported, MSCs can promote endothelial cell proliferation and migration [8,9,20], which is closely related to angiogenesis. Five items were enriched in the KEGG analysis (Supplementary Table S2), and the PI3K-Akt signaling pathway was reported to be involved in promoting endothelial cell proliferation, migration, and angiogenesis [23,24].
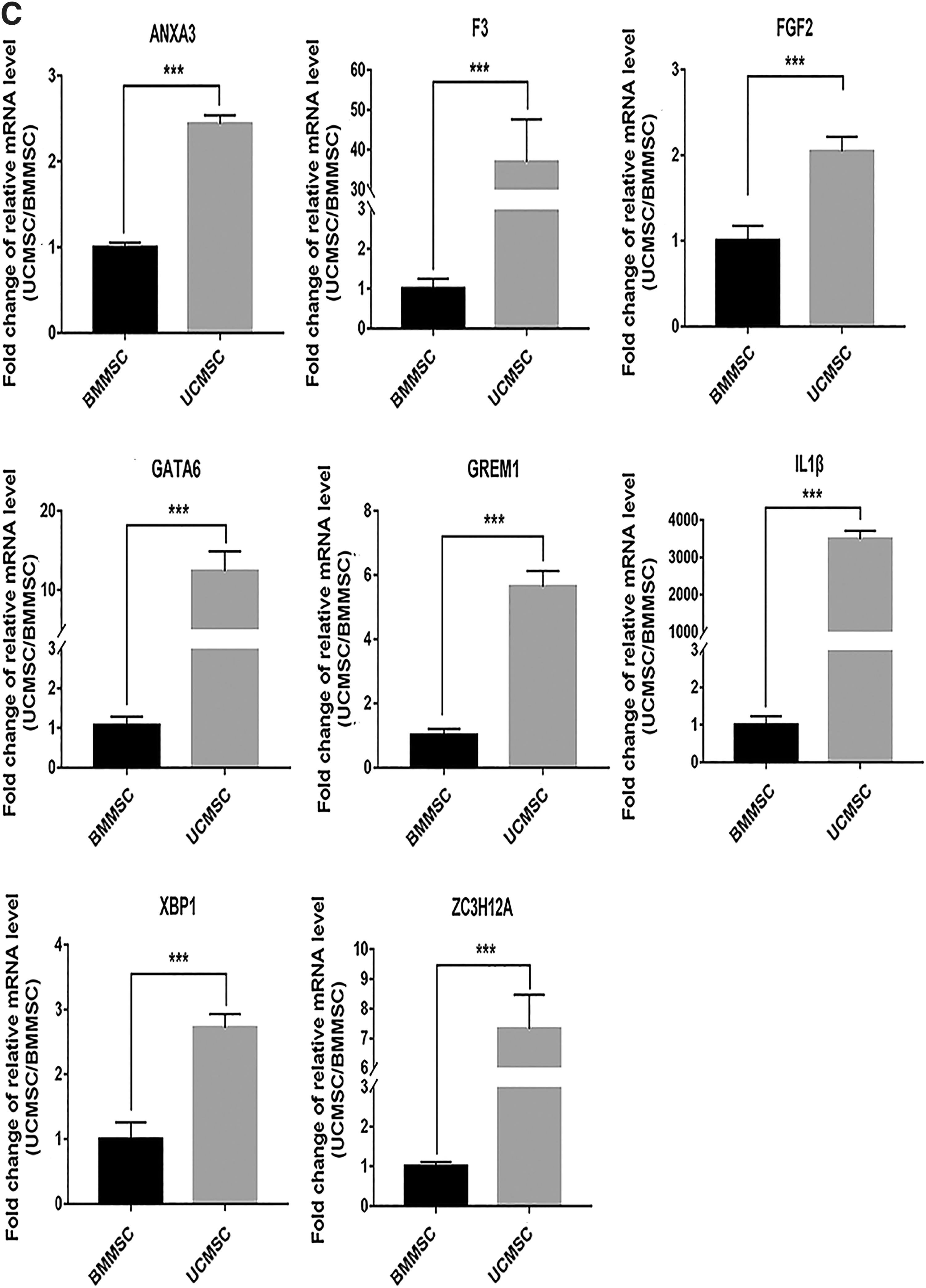
The DEGs involved in vasculogenesis and positive regulation of angiogenesis in the GO analysis and upregulated in UCMSCs from the mRNA microarray were validated using the RT-qPCR assay. The results indicated that mRNA expression levels of ANXA3, F3, FGF2, GATA6, GREM1, IL1β, XPB1, and ZC3H12A (Fig. 1C) and mRNA expression levels of FOXF1, HHEX, KDR, MYOCD, PIK3R3, PTGS2, SOX17, and ZFPM2 (Fig. 2B) were significantly upregulated in UCMSCs compared with BMMSCs.
Enrichment of angiogenesis-related items is upregulated in UCMSCs in the GSEA analysis.

Enrichment of angiogenesis items was upregulated in UCMSCs in the GSEA analysis
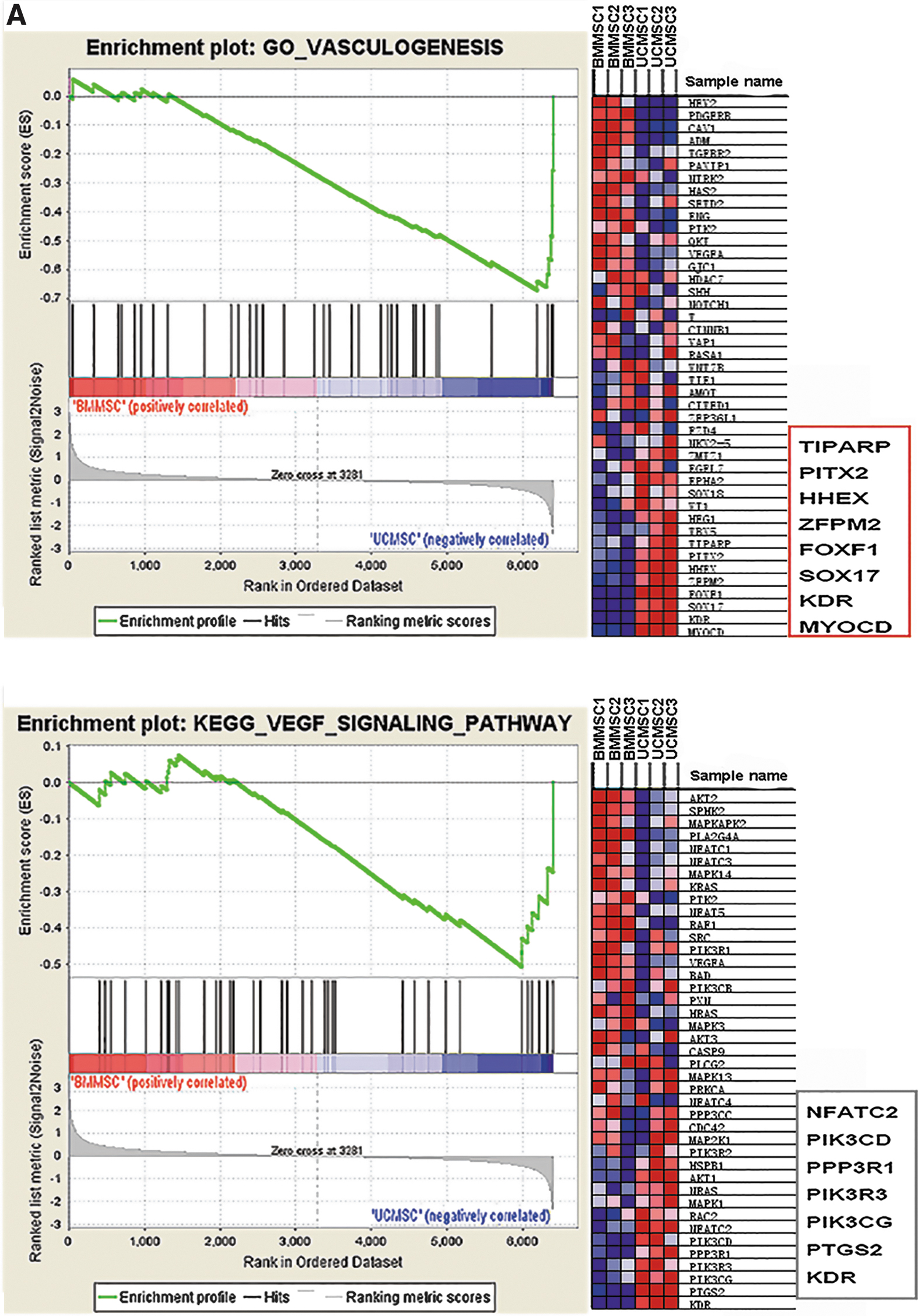
Both UCMSCs and BMMSCs could promote angiogenesis, which was supported by our GO analysis. To further explore whether UCMSCs or BMMSCs exert stronger effects on angiogenesis, GSEA analysis was conducted between the two MSCs. We found that vasculogenesis and the vascular endothelial growth factor (VEGF) signaling pathway were enriched in UCMSCs (Fig. 2A, P < 0.05), which indicated that UCMSCs could exert stronger proangiogenic effects compared with BMMSCs.
We validated the core-enriched genes of UCMSCs that were involved in vasculogenesis and the VEGF signaling pathway in the GSEA analysis (Fig. 2A, red box) through RT-qPCR. The results indicated significant upregulation of 8 genes, including FOXF1, HHEX, KDR, MYOCD, PIK3R3, PTGS2, SOX17, and ZFPM2, in UCMSCs compared with BMMSCs (Fig. 2B). These 8 genes were also the DEGs that were involved in vasculogenesis and positive regulation of angiogenesis in the above GO analysis (Fig. 1C).
These results supported the conclusion that UCMSCs can promote angiogenesis better than BMMSCs. This was further supported by the following in vivo and in vitro assays.
UCMSCs promoted HUVEC proliferation, migration, and tube formation in vitro
We further investigated the effects of UCMSCs and BMMSCs on HUVEC proliferation, migration, and tube formation. The CCK-8 analysis showed that both UCMSCs and BMMSCs significantly enhanced the proliferative ability of HUVECs (Fig. 3A), while HUVECs cultured in UCMSC-CM showed a stronger proliferative ability than those cultured in BMMSC-CM (Fig. 3A). In the scratch wound assay, both UCMSCs and BMMSCs significantly enhanced the migration ability of HUVECs, while HUVECs cultured in UCMSC-CM showed stronger migration ability than those cultured in BMMSC-CM (Fig. 3B). This was further verified by the Transwell migration assay (Fig. 3C). Furthermore, we employed the tube formation assay to examine the potential effects of MSCs on angiogenesis, finding that HUVECs cultured in both UCMSC-CM and BMMSC-CM formed many capillary-like structures, whereas HUVECs cultured in the control medium rarely formed tube structures. Moreover, the master junctions, total mesh areas, and total segment lengths in the UCMSC-CM group were significantly increased, larger, and longer compared with the BMMSC-CM group (Fig. 3D). Our results demonstrated that both UCMSCs and BMMSCs could promote angiogenesis, while UCMSCs exerted a stronger effect compared with BMMSCs.

UCMSCs promote HUVEC proliferation, migration, and tube formation in vitro.
UCMSCs promoted blood perfusion and attenuated hind limb ischemia in vivo
To investigate the biological functions of UCMSCs and BMMSCs in vivo, we used a mouse hind limb ischemia model by ligating the left femoral artery and injecting UCMSCs and BMMSCs directly into the left adductor muscle. The control group was ligated and injected with PBS. Tissue blood perfusion was measured by laser Doppler at days 0, 14, and 21 postinjection (Fig. 4A). We found that both UCMSCs and BMMSCs promoted blood perfusion in the ischemic limb. At day 0 postinjection, the UCMSC group, BMMSC group, and control group showed low levels of blood perfusion. At day 14, blood perfusion in MSC groups was gradually recovered, whereas in the control group, blood perfusion was still poor. At day 21, blood perfusion in MSC groups was significantly better than in the control group, which remained at a low level. Moreover, blood perfusion in the UCMSC group was significantly improved compared with that in the BMMSC group (Fig. 4A).

UCMSCs promote blood perfusion and attenuate hind limb ischemia in vivo.
Tissue blood perfusion recovery from hind limb ischemia primarily depends on angiogenesis in muscles because neovascularization provides nutrients and oxygen to ischemic muscles. Thus, to explore whether UCMSCs and BMMSCs can stimulate angiogenesis in vivo, we stained the endothelial marker CD31 in ischemic adductor muscles through immunofluorescence (IF) and immunohistochemical (IHC) staining. In the IF assay, we found that UCMSCs and BMMSCs significantly increased the average microvessel density (MVD), which is the number of microvessels per square millimeter [20]), compared with the control group (Fig. 4B). UCMSCs significantly increased the average MVD compared with BMMSCs. In IHC staining, the increase of average optical density (AOD) of CD31 represented an increase in small blood vessels. The results indicated that the AOD dramatically increased in both UCMSC and BMMSC groups compared with the control group (Fig. 4C), while the AOD dramatically increased in the UCMSC group compared with the BMMSC group.
These results suggested that transplantation of MSCs can stimulate angiogenesis in the ischemic muscle. UCMSCs played a more significant proangiogenic role than BMMSCs, which was in accordance with the in vitro assays.
UCMSCs promoted angiogenesis through the ERK and PI3K-Akt pathways
As reported, the paracrine activity of stem cells is an important mechanism in promoting angiogenesis [20,25,26]. To identify the differentially expressed proteins (DEPs) between BMMSC-CM and UCMSC-CM, we performed cytokine protein microarray detection (RayBiotech) using MSC-CM specimens (BMMSC-CM, n = 3; UCMSC-CM, n = 3). The screening standard for DEPs was defined as P < 0.05, log (fold change) >1.5. We found that 57 DEPs were upregulated in BMMSC-CM compared with UCMSC-CM, while 57 DEPs were upregulated in UCMSC-CM compared with BMMSC-CM (Fig. 5A).
UCMSCs promote angiogenesis through the ERK and PI3K-Akt pathways.
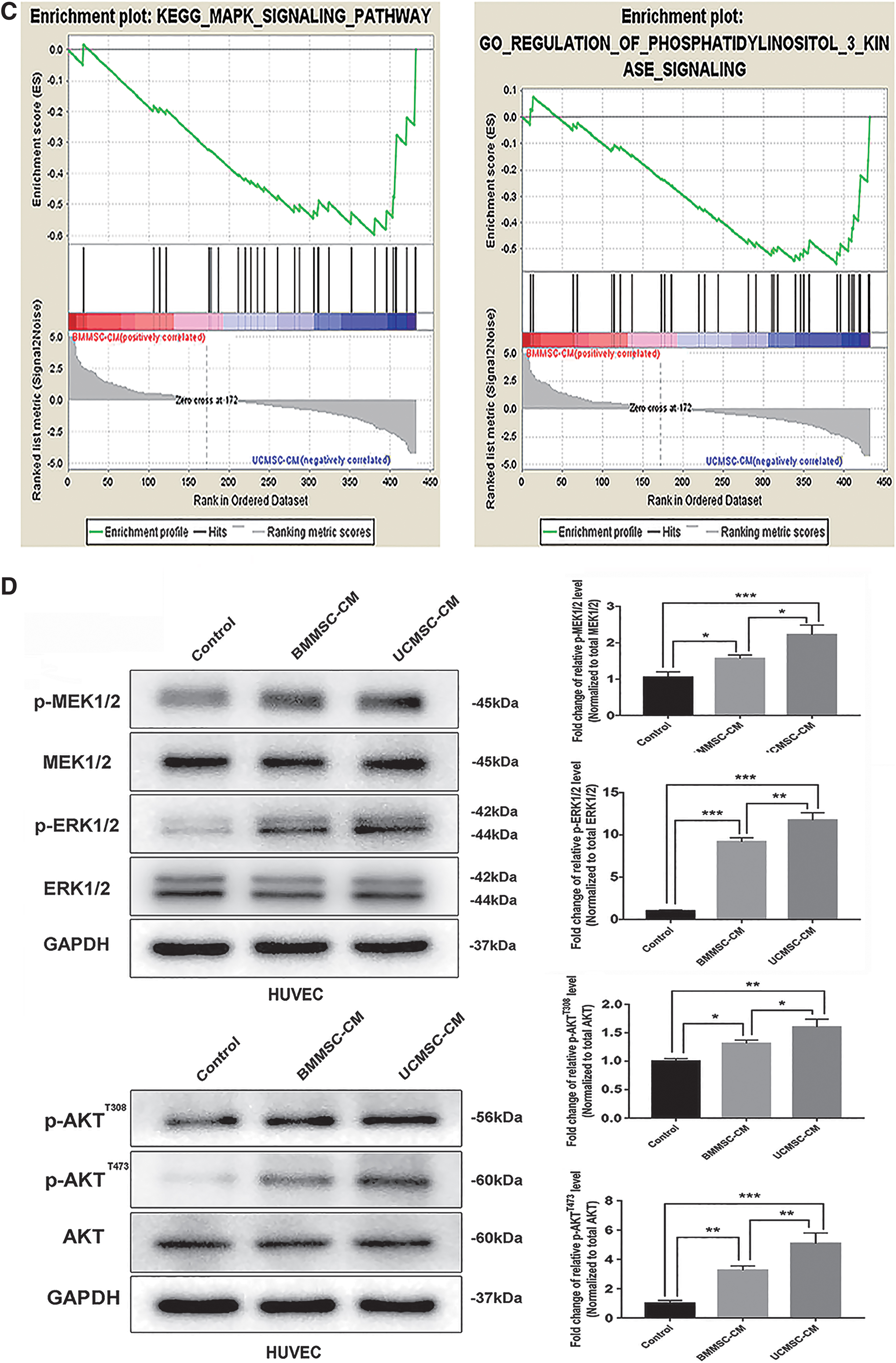
To further explore the biological functions of these DEPs, GO and KEGG analyses were conducted. Positive regulation of phosphatidylinositol 3-kinase (PI3K) signaling and positive regulation of the ERK1 and ERK2 cascade were enriched in the GO analysis biological process classification (FDR <0.05; Fig. 5B and Supplementary Table S3). The PI3K-Akt signaling pathway was enriched in the KEGG analysis (Fig. 5B and Supplementary Table S3). In the meantime, GSEA analysis was conducted between the two MSCs. We found that the MAPK signaling pathway and regulation of PI3K signaling were enriched in UCMSC-CM (Fig. 5C, P < 0.05), which indicated that UCMSCs could exert a stronger activation of ERK (MAPK) and PI3K pathways compared with BMMSCs.
The ERK [27,28] and PI3K-Akt signaling pathways [23,24] have been reported to be involved in promoting endothelial cell angiogenesis. The above results indicated that UCMSCs may promote endothelial cell angiogenesis through a paracrine mechanism, which activates the ERK and PI3K-Akt signaling pathways in endothelial cells. Thus, we cultivated HUVECs in CM from BMMSCs and UCMSCs and then examined the related pathway protein levels of incubated HUVECs.
We detected the phosphorylation levels of MEK1/2, ERK1/2, and Akt in CM-cultivated HUVECs to evaluate activation of the ERK and PI3K-Akt pathways. The results indicated significantly increased phosphorylation levels of MEK1/2, ERK1/2, and Akt in HUVECs cultivated in the CM of BMMSCs and UCMSCs compared with the control (Fig. 5D), while phosphorylation levels of MEK1/2, ERK1/2, and Akt in HUVECs cultivated in UCMSC-CM were significantly upregulated compared with HUVECs cultivated in BMMSC-CM (Fig. 5D). These findings demonstrated that UCMSCs exerted stronger effects on angiogenesis in HUVECs through the ERK and PI3K-Akt pathways.
Discussion
In this research, our results indicated that both UCMSCs and BMMSCs can significantly promote HUVEC proliferation, migration, and tube formation, as well as improve blood perfusion of ischemic limbs, which is supported by previous studies [10,12,13,15,16]. The results lead us to conclude that UCMSCs have a stronger angiogenic effect than BMMSCs, which is supported by our GSEA analysis and in vitro and in vivo assays, especially the ERK and PI3K-Akt pathway analyses. These results indicate that UCMSCs play an important role in therapeutic angiogenesis.
Combined with previous studies [20,25,26], stem cells have been shown to promote angiogenesis through paracrine activity, which was validated in our study by cultivating HUVECs in CM from UCMSCs and BMMSCs. Paracrine factors from MSCs may stimulate and activate the related receptors on HUVEC membranes [8,29], and activation of receptors further phosphorylates and activates the ERK and PI3K-Akt signaling pathways [23,24,27,28]. Phosphorylation and activation of MEK1/2 can phosphorylate and activate ERK1/2, and this process indicates activation of the ERK pathway. Meanwhile, phosphorylation of Akt indicates activation of the PI3K-Akt pathway. Activation of these two signaling pathways leads to nuclear transcription to promote HUVEC proliferation, differentiation, and migration, with which angiogenesis is stimulated [29]. In our research, HUVECs cultured in MSC-CM showed significant phosphorylation of MEK1/2, ERK1/2, and Akt. Phosphorylation of MEK1/2, ERK1/2, and Akt was more significant in HUVECs cultured in CM from UCMSCs compared with BMMSCs. Therefore, activation of the ERK and PI3K-Akt pathways may be the mechanism by which UCMSCs promote angiogenesis (Fig. 6).

Schematic diagram summarizing the signaling pathways through which UCMSCs promote angiogenesis.
As reported, MSCs from bone marrow and peripheral blood have been validated for their safety and effectiveness in patients with CLI [30] and have been investigated in phase II clinical trials [13]. Human umbilical cord blood-derived MSCs and adipose-derived stem cells have also been studied in phase I trials of patients with limb ischemia; however, their safety and effectiveness need further assessment [31,32]. UCMSCs have shown the ability to differentiate into three germ layers, promote tissue repair, and modulate immune responses [14]. In our research, UCMSCs significantly stimulate angiogenesis, which was concordant with previous reports [10,15,16]. Furthermore, our study indicated that UCMSCs can significantly activate the ERK and PI3K-Akt pathways in HUVECs, which confirmed the paracrine mechanism of UCMSCs in promoting angiogenesis. In addition, UCMSCs can modulate the immune response because of their low immunogenicity [14]. Thus, UCMSCs play an excellent role in rescuing CLI patients and are worthy of further study.
Footnotes
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This work was supported by the National Natural Science Foundation of China [nos. 81070258 and 81270378].
Supplementary Material
Supplmentary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.