
Abstract
Osteoarthritis challenges traditional therapies and remains a leading cause of lameness in older dogs. Regenerative medicine offers new strategies, typically involving the injection of autologous adipose-derived mesenchymal stem cells (MSCs). Conversely, allogenic MSCs are appealing candidates to palliate patient morbidity and cell preparation time. Regardless of the source of cells, identifying critical donor characteristics, such as age, is essential to obtain the most competent MSCs. The objectives of this study were to determine the influence of donor's age on proliferation, gene expression, and immunomodulatory properties of MSCs in dogs. Canine adipose tissue-derived MSCs (cAD-MSCs) were isolated from the falciform-ligament adipose tissues of nine pairs of gender-matched young (<2 years) or old (>7 years) client-owned dogs undergoing abdominal surgery. Growth kinetics, transcriptome before and after stimulation by tumor necrosis factor alpha and interferon gamma, MSC-induced lymphocyte suppression assay, and secretion of prostaglandin E2 (PGE2) and indoleamine 2,3-dioxygenase (IDO) were compared between cells obtained from young or old dogs. The doubling times at passages 2 and 3 were shorter when MSCs were isolated from young (34.8 ± 1.8 h and 46.3 ± 2.3 h) rather than old dogs (56.5 ± 8.0 h and 123.8 ± 46.7 h, P < 0.05). The MSC transcriptomes from both populations were similar without stimulation, while stimulation resulted in a 3-fold greater expression of osteogenic gene, fibroblast growth factor 10, in cells from old dogs. cAD-MSCs from young dogs suppressed proliferation of activated T cells more strongly (P < 0.05), although secretion of PGE2 and IDO did not differ between groups. In conclusion, donors' age affected proliferation, immunomodulatory properties of cAD-MSCs, and increased expression of osteogenic gene under proinflammatory conditions in our population of dogs. Collectively, our results provide evidence to support further evaluation of allogenic MSC therapies derived from young donors as alternatives to autologous MSC therapy in older dogs.
Introduction
Osteoarthritis (OA) is a leading cause of lameness in dogs, affecting up to 18 million dogs in the United States [1]. This condition is especially prevalent in older dogs [2]. Ultimately, OA results in loss of articular cartilage, and current therapies do not alter the pathologic course of the disease. Mesenchymal stem cell (MSC) therapy has attracted significant attention for treatment of OA because of the potential ability of MSCs to repair damaged tissue by self-renewal and multilineage differentiation [3]. In addition, these cells have immunomodulatory properties that could address the stress-induced and proinflammatory mechanisms involved in the pathogenesis of OA [4].
Immunosuppressive effects of adipose tissue-derived MSCs (AD-MSCs) have been documented in dogs, cats, and horses who received AD-MSCs either systematically or locally [5 –7]. In vitro, canine AD-MSCs (cAD-MSCs) have been found to suppress mitogen-induced T cell proliferation [8 –10]. In this coculture system, concentrations of anti-inflammatory mediators, such as transforming growth factor beta (TGF-β), prostaglandin E2 (PGE2), indoleamine 2,3-dioxygenase (IDO), and interleukin 6 (IL-6), were increased [8 –10]. Inhibition of PGE2 and IDO partially restored T cell proliferation, providing further evidence that these soluble anti-inflammatory mediators may contribute to the immunomodulating properties of stem cells [8]. However, inhibition of PGE2 did not restore T cell proliferation in another study, justifying further investigation [9].
MSC therapy in dogs currently relies primarily on the administration of autologous type II stem cell-based products, usually stromal vascular fraction of fat, because these cells pose less risk than allogenic, ex vivo manipulated cells [11,12]. Canine MSCs are generally administered to manage degenerative diseases, such as OA, most commonly diagnosed in older dogs [2,12,13]. The demographics of dogs are relevant to treatment efficacy, because in humans, the patients' health status and age have been found to affect stem cell properties [14 –16]. In dogs, donor's age has been found to affect MSC proliferation [17,18], number of colony forming units (CFUs) [19], differentiation potential [17,19], and expression of pluripotent genes [17].
However, influence of age on immunosuppressive capacity seems more controversial: age did not correlate with suppression of T cell proliferation by human bone marrow-derived MSCs (hBM-MSCs) in vitro [14]; age affected human MSCs to suppress inflammation and microglia in vivo [20,21]; and age affected rat MSCs to suppress CD4+ T cell proliferation in vitro [22]. The effect of donor's age on immunomodulation capacity of cAD-MSCs has not been investigated. This gap in knowledge is important because it affects our ability to predict the efficacy of autologous stem cell therapy, optimize our selection of treatment modalities for older dogs, and establish criteria for selection of allogenic stem cell source.
Therefore, the objective of this study was to determine the influence of donor's age on proliferation, gene expression, and immunomodulatory properties of cAD-MSCs. Considering published evidence in other species, we hypothesized that donor's age would affect MSCs proliferation and gene expression while its effect on immunomodulation remained an open question.
Materials and Methods
Animals
All procedures in this study were approved by the Institutional Animal Care and Use Committee at the Western University of Health Sciences (Protocol No.: R16IACUC047). Dogs undergoing abdominal surgery for reasons unrelated to the study were enrolled with owners' consent from three local veterinary clinics: Western University of Health Sciences Pet Health Center (Pomona, CA), VCA California Veterinary Specialists (Carlsbad, CA), and VCA Advanced Veterinary Care Center (Lawndale, CA).
Dogs were included in the study if they underwent abdominal surgery that would involve removal of falciform-ligament adipose tissue. Dogs were classified as “young” if they were <2 years of age, and “old” if they were >7 years of age. Exclusion criteria included dogs with intraoperative evidence of inflammatory or neoplastic diseases involving the peritoneal surface or grossly abnormal falciform ligament.
Isolation and maintenance of cAD-MSCs
Approximately 10 g of falciform-ligament adipose tissue harvested during surgery was minced and digested in 15 mL of 0.1% collagenase type IA (Worthington Biochemical Corporation, Lakewood, NJ) and 1% bovine serum albumin (HyClone, Logan, UT) in phosphate-buffered saline on a shaker (180 rpm) at 37°C for 2 h. Cells were resuspended in low-glucose Dulbecco's modified Eagle's medium (HyClone) with 10% fetal bovine serum (HyClone) and 1% penicillin-streptomycin, and cultured in T25 flasks at 37°C, 5% carbon dioxide, and 90% humidity. Cell culture medium was changed every 3rd day until 70%–80% confluence, at which point cells were passaged at 5 × 103 cells/cm2 density 1st time (Passage 1: P1).
Growth kinetics
After 6 days of culture at P1, cells were passaged at the same density. Cells were counted by trypan blue dye exclusion method in triplicate. This process was repeated from P1 to P4. Population doubling (PD) was calculated at each passage as follows: PD = ¼ln(Nf/Ni)/ln2, where Ni and Nf are initial and final cell numbers, respectively, and ln is the natural log. Doubling time (DT) was calculated as follows: DT = culture time (CT)/PD. The PDs of continuous subcultures were added to obtain the cumulative population doubling level (CPDL). Cell morphology was subjectively evaluated at each change in medium.
Transcriptome analysis
Total RNA was isolated from cAD-MSCs at P3 with the RNeasy Mini Kit (QIAGEN, Hilden, Germany). One microgram of total RNA was used for complementary DNA (cDNA) synthesis with the RT2 First Strand Kit (QIAGEN). cDNA samples were mixed with RT2 SYBR Green qPCR Mastermix (QIAGEN) and distributed into each well of RT2 Profiler PCR Array Dog Mesenchymal Stem Cell (QIAGEN) with 84 selected genes. Real-time PCR was performed with CFX-Connect (Bio-Rad, Hercules, CA). Four housekeeping genes (ACTB, HPRT1, PGK1, and RPL13A) were used for normalization. The change in gene expression was calculated by the delta–delta Ct method (2−ΔΔCt) and expressed as fold change.
To test transcriptomic change stimulated by the proinflammatory cytokines, tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ), cells were seeded onto 24-well plates at 1 × 105 cells/cm2 density. Cells were incubated with TNF-α (Kingfisher, Saint Paul, MN) and IFN-γ (RD Systems, Minneapolis, MN) both at 50 ng/mL for 24 h before RNA isolation, as described above.
MSC-induced lymphocyte suppression assay
MSC-induced lymphocyte suppression assay was performed using an exact protocol previously validated [9,10]. Mitotically inactivated cAD-MSCs at P3 and peripheral blood mononuclear cells (PBMCs) were cocultured at 1:5 ratio. PMBCs were activated with 5 μg/mL concanavalin A (ConA; Sigma, St. Louis, MO). After 3 days of coculture, PBMCs were treated with 1 mM bromodeoxyuridine (BrdU; BD Biosciences, Franklin Lakes, NJ). After 24 h incubation, PBMCs were stained with APC anti-BrdU following manufacturer's instructions (APC BrdU Flow Kit; BD Biosciences).
The percentage of BrdU-positive T cells from PBMCs alone served as negative control and that from PBMCs activated by ConA served as positive control (Supplementary Fig. S1). The percentage of BrdU-positive T cells for each sample was calculated as follows: (percentage of BrdU-positive T cells from PBMCs cocultured with cAD-MSCs and activated by ConA – negative control)/(positive control – negative control) × 100. cAD-MSCs from gender-paired young and old dogs were included in the same trial.
Anti-inflammatory mediator production
During quantification of proliferated T cells in MSC-induced lymphocyte suppression assay, supernatants were collected, centrifuged, and stored at −80°C until processed. PGE2 was quantified with Prostaglandin E2 Express EIA Kit (Cayman Chemical Company, Ann Arbor, MI) according to manufacturer's instructions. IDO enzymatic activity was measured through quantification of its metabolite, kynurenine, as previously described [8].
Statistics
Results are presented as mean ± standard error of the mean. Cells from nine pairs of gender-matched young and old dogs were compared. Normality of data was examined with the Kolmogorov–Smirnov test.
DT and CPDL in growth kinetics were compared with the nonparametric Wilcoxon matched-pairs signed rank test. All Ct values obtained from transcriptome analysis were compared using RT2 Profiler PCR Array Data Analysis v3.5 (QIAGEN). To evaluate transcriptome changes by proinflammatory cytokines, cells from the same donors were compared between pre/poststimulation. Percentages of BrdU-positive T cells for each sample in MSC-induced lymphocyte suppression assay were compared with a paired t-test. Concentrations of anti-inflammatory mediators were compared with Friedman's test as nonparametric repeated-measures analysis of variance among 4 groups (1. PBMCs alone; 2. PBMCs activated by ConA; 3. PBMCs activated by ConA and cocultured with cells from young dogs; 4. PBMCs activated by ConA and cocultured with cells from old dogs) with the Bonferroni correction when differences were detected.
Statistical analysis was performed using SAS® 9.4 (SAS Institute, Cary, NC). The level of significance was set at P < 0.05, and marginal significance (tendency) was set at P < 0.1.
Results
Animals
The 18 dogs enrolled in this study were paired by gender (9 pairs; Supplementary Table S1). Young dogs included 5 intact females (3 Boxer, 1 Golden Retriever, and 1 Siberian Husky), 2 spayed females (1 Bull Terrier and 1 Border Terrier), and 2 neutered males (1 German Shepherd and 1 Labrador Retriever Mix). Old dogs included 1 intact female (1 Chihuahua), 6 spayed females (2 Labrador Retriever Mix, 1 German Shepherd, 1 American Eskimo, 1 Dalmatian, and 1 Mongrel), and 2 neutered males (1 Terrier Pitbull Mix and 1 Great Pyrenees Mix). The average age of young dogs was 13.0 ± 2.3 months (range: 5–23 months), while that of old dogs was 112.6 ± 9.7 months (range: 84–163 months). Body weight did not significantly differ between young (18.6 ± 2.1 kg) and old dogs (24.0 ± 2.8 kg, P = 0.16).
The indications for abdominal surgeries included 6 exploratory celiotomies for gastrointestinal diseases (1 pyloric obstruction, 1 intussusception, 4 negative explorations), 5 ovariohysterectomies, 3 splenectomies (1 hyperplastic nodule, 2 hematomas), 3 foreign bodies, and 1 gastric dilation and volvulus. Adipose tissue was consistently harvested immediately upon celiotomy.
Growth kinetics
The DTs of cells isolated from young (38.3 ± 1.3 h) and old dogs (42.9 ± 3.3 h, P = 0.6) did not differ at P1. The DTs at P2 and P3 were shorter when cells were isolated from young (34.8 ± 1.8 h and 46.3 ± 2.3 h) rather than old dogs (56.5 ± 8.0 h and 123.8 ± 46.7 h, P < 0.05; Fig. 1A). Similarly, the CPDLs of cells from young (3.8 ± 0.1) and old (3.7 ± 0.2) dogs did not differ at P1. However, CPDLs of cells isolated from young dogs tended to be higher (7.9 ± 0.2) than those of cells from old dogs at P2 (7.3 ± 0.2, P = 0.05) and were higher (11.2 ± 0.3) than those of cells from old dogs at P3 (9.5 ± 0.3, P < 0.05; Fig. 1B).
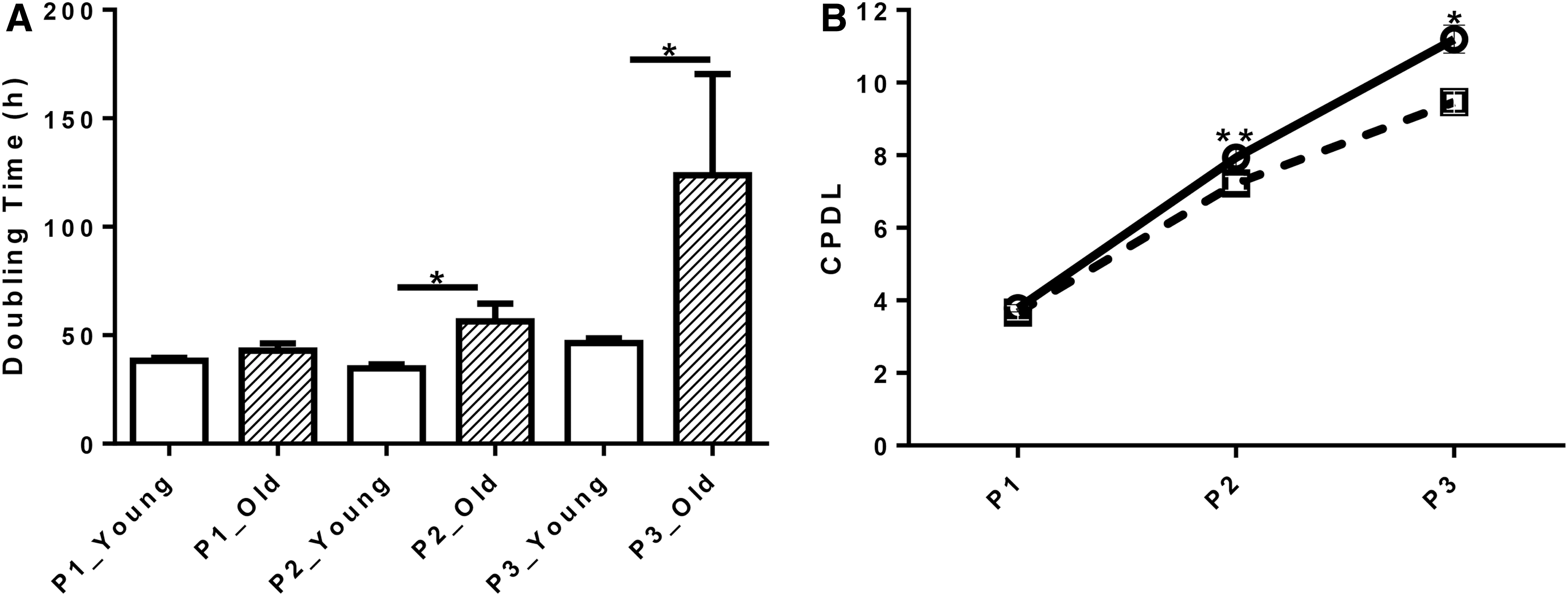
Growth kinetics of cAD-MSCs from young and old dogs. Doubling time
Changes in cellular morphology consistent with senescence corroborated the results above (Fig. 2). At P1, cellular size and confluency after 6 days of culture were similar between cells from young and old dogs. However, at P2, cells appeared larger and less confluent in wells containing cells from old dogs compared with those from young dogs. At P3, signs of senescence became prominent in cells from old dogs.
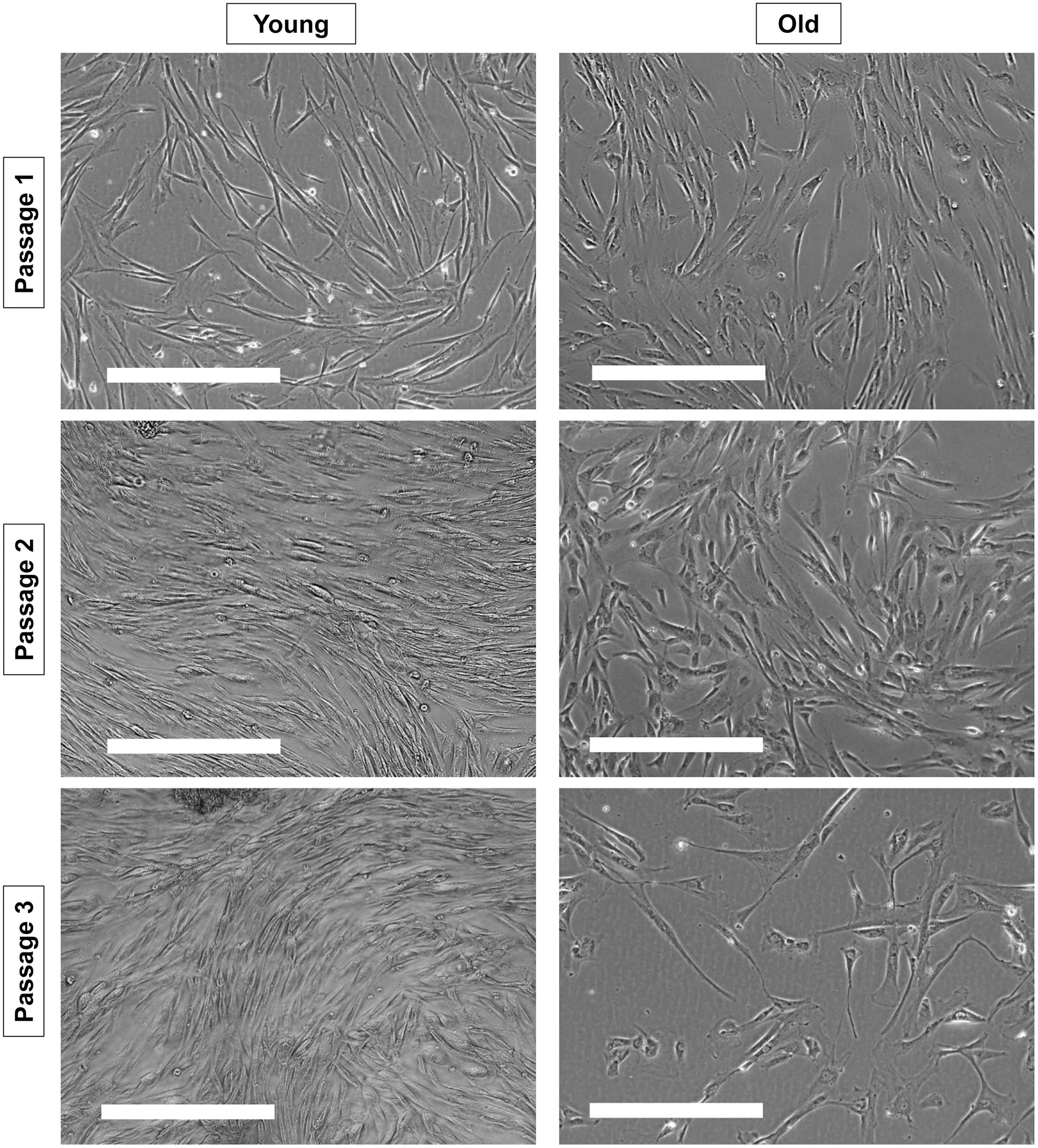
Representative morphology of cAD-MSCs from young and old dogs. Morphology of cAD-MSCs from representative young dogs (left panels) and old dogs (right panels) after 6 days of culture at passages 1, 2, and 3 is shown. Scale bars = 400 μm.
Transcriptome analysis
The expressions of PPARG (P = 0.09), ADIPOQ (P = 0.06), and SOX9 (P = 0.08) were increased by at least twofold in cells from young dogs compared with old dogs without stimulation (Fig. 3A; Table 1A). After stimulation with TNF-α and IFN-γ, the expression of FGF10 was increased by 2.87-fold (P < 0.05) in cells from old dogs compared with young dogs (Fig. 3B; Table 1B).

Volcano plot of the transcriptome of cells isolated from young and old dogs, with or without stimulation by tumor necrosis factor alpha and interferon gamma. Each dot represents a gene plotted on an x-axis representing log2 (fold change) against y-axis representing—log10 (P value). Expression levels of each gene were compared between cells from old dogs/cells from young dogs without stimulation
List of Genes with More Than Twofold Change Between Cells from Old Versus Young Dogs (A and B) and Between Cells Stimulated Versus Unstimulated Conditions (C and D)
Fold regulation of each gene expression was compared between cells from old and young dogs without (A) and with (B) stimulation. Fold regulation of each gene expression was compared between stimulated and unstimulated conditions in cells from young (C) and old (D) dogs. Positive values indicate upregulation and negative values indicate downregulation.
Suggested gene functions based on criteria from RT2 Profiler™ PCR Array Dog Mesenchymal Stem Cell (QIAGEN) and in the context of immunomodulation.
MSC, mesenchymal stem cell.
Within cAD-MSCs isolated from each age group, stimulation with TNF-α and IFN-γ resulted in dramatic changes in transcriptome patterns represented by cluster 1 (with stimulation) and cluster 2 (without stimulation; Supplementary Fig. S2). In cAD-MSCs isolated from young dogs, stimulation upregulated 9 genes (IL6, VCAM1, NGFR, ICAM1, B2M, LIF, ABCB1, CEBPD, and FGF2) by more than twofold, while stimulation downregulated 18 genes (ANPEP, BMP4, FGF10, TGFB3, NT5E, PPARG, LPL, NES, PROM1, MMP2, TBX5, KAT2B, ADIPOQ, KITLG, SOX9, INSR, MITF, and FASN) by more than twofold (Fig. 3C; Table 1C). In cAD-MSCs isolated from old dogs, stimulation upregulated 10 genes (IL6, VCAM1, ICAM1, B2M, ABCB1, BDNF, HGF, LIF, CEBPD, and FGF2) by more than twofold, while stimulation downregulated 10 genes (TGFB3, ANPEP, BMP4, LPL, FGF10, NES, ACTA2, MMP2, FASN, and KAT2B) by more than twofold (Fig. 3D; Table 1D).
MSC-induced lymphocyte suppression assay
As a measurement of T cell proliferation, percentages of CD3+ T cells incorporated BrdU within PBMCs cocultured with cAD-MSCs isolated from young and old dogs were normalized to positive control (PBMCs + ConA) and compared. cAD-MSCs isolated from young dogs inhibited activated T cell proliferation (54.6% ± 5.4%) to a greater percentage than cAD-MSCs isolated from old dogs (64.7% ± 8.2%, P < 0.05; Fig. 4).
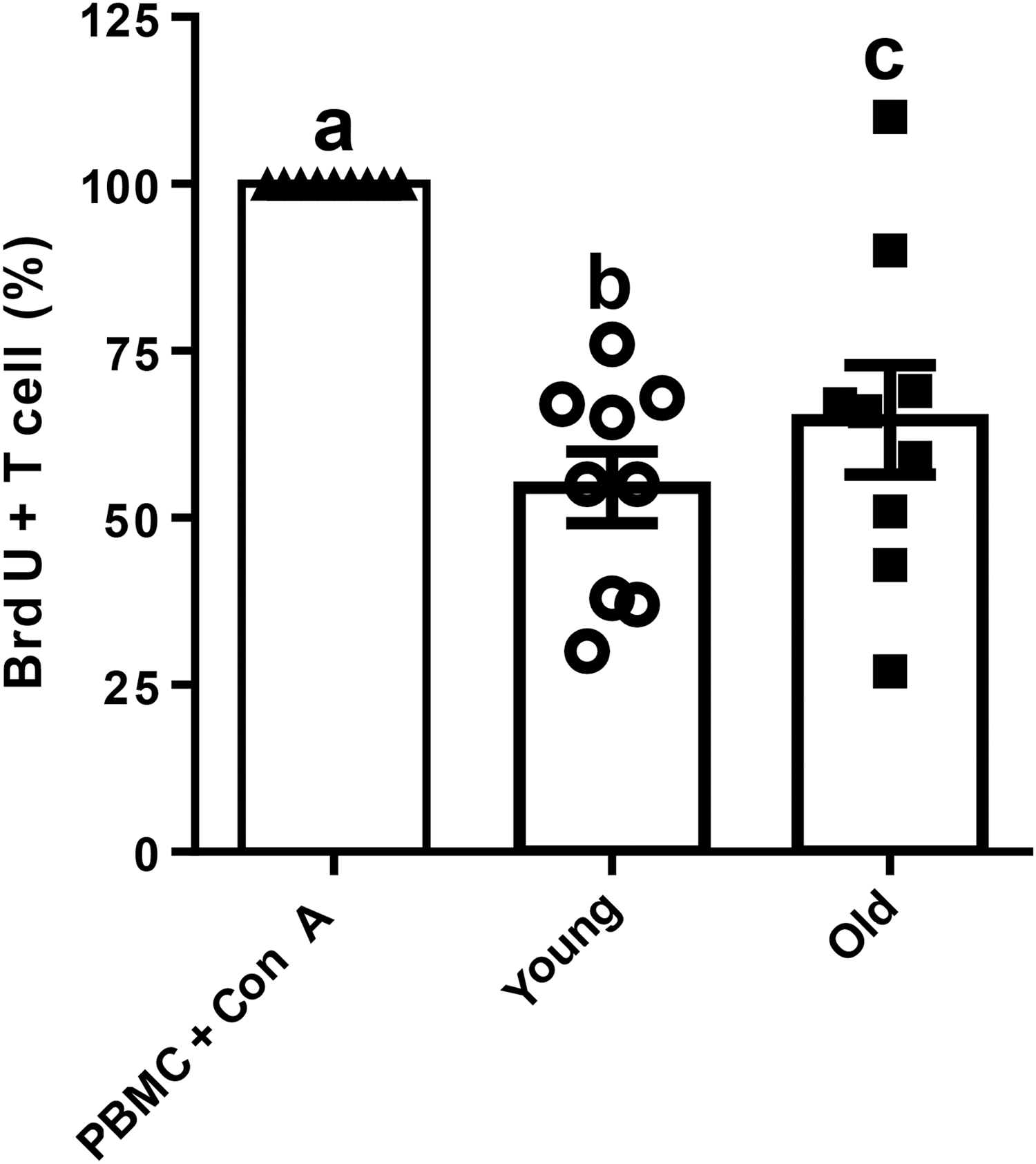
MSC-induced lymphocyte suppression assay. Percentages of BrdU+ T cells cocultured with cAD-MSCs isolated from young (white circle) and old (black square) dogs were normalized with those of control group (PBMCs + ConA, black triangle). Different lowercase letters indicate P < 0.05. BrdU, bromodeoxyuridine; ConA, concanavalin A; PBMCs, peripheral blood mononuclear cells.
Anti-inflammatory mediators
PGE2 concentration was higher in supernatants when PBMCs were cocultured with cAD-MSCs compared with cultures of PBMCs alone or PBMCs stimulated with ConA. However, PGE2 concentration did not differ between supernatants collected from coculture of PBMCs with cAD-MSCs isolated from young or old dogs (P = 0.096). IDO concentration was higher when PBMCs were stimulated with ConA regardless of coculture with cAD-MSCs than PBMCs alone. However, IDO concentration did not differ when stimulated PBMCs were cocultured with young or old dogs (P = 0.096; Fig. 5).

Concentrations of PGE2 (left panel) and IDO (right panel) in culture supernatant during MSC-induced lymphocyte suppression assay. Supernatants were collected from coculture of PBMCs alone (PBMC, white triangle), PBMCs activated by ConA (PBMC + ConA, black triangle), PBMCs activated by ConA and cocultured with cAD-MSCs isolated from young dogs (young, white circle), and PBMCs activated by ConA and cocultured with cAD-MSCs isolated from old dogs (old, black square). Different lowercase letters indicate P < 0.05. IDO, indoleamine 2,3-dioxygenase; PGE2, prostaglandin E2.
Discussion
The main findings of this study are that in our population of dogs, donor's age affected the ability of cAD-MSCs to proliferate and suppress T cell proliferation, as well as influenced gene expression related to multipotentiality under inflammatory microenvironments in vitro.
The shorter DT of cAD-MSCs isolated from young dogs was consistent with a more robust cell proliferation in this population compared with cAD-MSCs isolated from old dogs. This finding was in general agreement with previous reports, where age had a negative effect on the DT and the number of CFUs in cAD-MSCs [17,18], canine BM-MSCs [19], and hBM-MSCs [14,23,24]. This influence had been attributed to the predisposition of MSCs isolated from older donors to senescence [25]. For example, the lower yield and prolonged DT of hBM-MSCs isolated from older donors were associated with shortened telomere restriction fragment [26]. In addition, hBM-MSCs isolated from older donors had a prolonged DT, increased apoptosis, and senescence-associated β-galactosidase-positive staining [15,27].
Age seemed to affect the immunosuppressive capacity of cAD-MSCs because cells from young dogs suppressed T cell proliferation more strongly than those from old dogs. Our finding was in agreement with several studies on rat MSCs in vitro [22] and human MSCs in vivo [20,21], but differed from another study in humans, where donors' age did not correlate with the immunosuppressive capacity of hBM-MSCs in vitro [14]. This discrepancy likely reflects differences of species, cell sources, and experimental conditions in mechanisms involved in the immunomodulation provided by MSCs.
In our study, cAD-MSCs were cocultured with allogenic PBMCs at a ratio of 1:5, and cells from young and old dogs reduced T cell proliferation by 45% and 35%, respectively. In previous reports, cAD-MSCs reduced T cell proliferation by ∼25% and 75%, when they were cocultured with allogenic PBMCs at 1:1 and 1:10, respectively [8,28]. The relatively high T cell proliferation and variation among studies may stem from diverse major histocompatibility complex (MHC) class II expression profiles of MSCs. Indeed, allogenic equine MSCs expressing haplotype-mismatched MHC II allowed a stronger proliferation of T cells than those expressing haplotype-matched or nonexpressing MHC II [29].
The immunogenicity of allogenic MSCs has been mainly attributed to this MHC II mismatching, and has been associated with a short survival of implanted cells [30,31]. The superiority of cAD-MSCs from young dogs on immunosuppressive capacity in our study was subtle and could be negated if MHC II-mismatched allogenic MSCs are administered. However, the influence of MHC II haplotype matching on immunosuppressive capacities of canine allogenic MSCs has not been established and warrants further studies.
The mechanism involved in the decline of immunosuppressive capacity of cAD-MSCs obtained from old dogs does not seem related to PGE2 and IDO, because the concentrations of these anti-inflammatory mediators did not seem affected by the age of the donor. This apparent lack of influence of age on the secretion of PGE2 and IDO by cAD-MSCs was consistent with findings reported on hBM-MSCs tested for expression of PGE2 and IDO during MSC-induced lymphocyte suppression assay [14], even after stimulation by IFN-γ [32,33]. Collectively, these data provide evidence to suggest that donor's age does not have a profound effect on the capacity of MSCs to produce PGE2 and IDO, and may instead act on other mechanisms.
The mechanisms of immunosuppression by cAD-MSCs are thought to rely on direct contact with T cells and secretion of several factors, including PGE2, IDO but also others, such as TGF-β and adenosine [8,28,34]. In hBM-MSCs, donor's age affected direct contact mechanisms, including expression of vascular cell adhesion molecule 1 (VCAM1), activated leukocyte cell adhesion molecule, and programmed death ligand 1 [14].
From the transcriptome analysis in our study, the proinflammatory cytokines TNF-α and IFN-γ upregulated the expression of immunosuppressive genes, IL6, VCAM1, and ICAM1, in cAD-MSCs from both young and old dogs. Although expression levels did not statistically differ between these populations upon stimulation, IL6 and VCAM1 were more likely to be upregulated in cAD-MSCs from young dogs, with 102 to 104-fold lower P values. Another immunosuppressive gene, NGFR, was upregulated only in cells from young dogs.
IL-6, a pleiotropic cytokine, is known to upregulate during MSC-induced lymphocyte suppression assay and function as anti-inflammatory cytokine of canine MSCs [8 –10]. VCAM1 and intercellular adhesion molecule 1 (ICAM1) are both implicated in direct contact mechanisms of MSCs with T cells that are essential in immunosuppression and inducible by proinflammatory cytokines [35,36]. A recent study reported that MSCs cause cell cycle arrest of activated T cells at G0–G1 through ICAM1 [37]. Nerve growth factor receptor (NGFR), a specific marker of MSCs [38], plays an essential role in suppression of T cell activation by melanoma cells [39]. These immunosuppressive molecules may contribute to the stronger immunosuppressive capacity of cAD-MSCs from young dogs.
Age did not alter the transcriptomes of cAD-MSCs under normal conditions, confirming the results of previous studies on cAD-MSCs using identical assay [40] and human MSCs using microarrays [41,42]. The only influence of age on gene expression consisted of an overexpression (2.87-fold) of the osteogenic gene, fibroblast growth factor 10 (FGF10), in cAD-MSCs from old dogs, under inflammatory conditions. Although its exact role has yet to be elucidated, FGF10 has been reported to regulate the epithelial expression of sonic hedgehog, which enhances osteogenesis [43,44].
In general, donor's age is considered to reduce the osteogenic potential of MSCs [17,19,45 –47]. The apparent discrepancy with our findings may stem from differences in environment, our study simulating MSCs' response under inflammatory microenvironment rather than the normal microenvironment used in previous reports. MSCs may respond differently under normal and inflammatory conditions. For example, the negative effects of amyotrophic lateral sclerosis and chronic obstructive pulmonary disease on immunosuppression of MSCs were detectable only under an inflammatory microenvironment [48,49]. To our knowledge, our findings are the first to support the concept of enhanced osteogenicity of cAD-MSCs from old dogs under inflammatory microenvironment. Such activity could increase unregulated bone formation, such as osteophytes.
The main limitations of our study are inherent to its clinical nature, affecting our ability to control variables between canine populations. Indeed, adipose tissue was harvested during abdominal surgeries unrelated to the study, creating a bias in the indications for surgeries and health status of dogs. The most common indication for surgery consisted of elective neutering in young dogs, whereas old dogs were more commonly affected with comorbidities. This bias was mitigated by exclusion of dogs with intraoperative evidence of inflammatory or neoplastic diseases involving the peritoneal surface as well as those with grossly abnormal falciform ligament. In addition, adipose tissue was harvested before any manipulation of intra-abdominal organs.
The second limitation deals with our classification of dogs based on age. While dogs <2 years of age are considered juvenile in all breeds, dogs >7 years of age may be considered as adult in small (<9.1 kg) breeds, senior in medium to large (9.5–54.5 kg) breeds, and geriatric for very large (>54.5 kg) breeds [50]. A cutoff of 7 years was selected for inclusion as “old dog” in our study as a compromise between these classifications. We did not restrict size, breeds, or body weight in our study as we are not aware of any publication documenting an influence of these variables on stem cell behavior in dogs. Instead, we aimed to expand our pool of candidates to all dogs meeting our age requirements. Although unintentional, our population of old dogs was fairly homogeneous, with no giant breed dog, and only dog weighing <9 kg (Chihuahua). This inclusion would not affect our conclusions regarding differences identified between groups but could have contributed to type II errors, if this dog's cells behave similarly as those obtained from young dogs.
Small sample size could also have led to type II error, affecting our mechanistic analysis on secretion of PGE2 and IDO. Similarly, gene expression levels of TGF-β1 and TGF-β3 both with and without stimulation were not different between cAD-MSCs from young and old dogs. Since multiple mechanisms, including PGE2, IDO, TGF-β, adenosine, direct contact, and potentially more, are thought to contribute to immunosuppressive capacity of cAD-MSCs, further investigations may require much larger sample size to detect subtle age effects on each mechanism.
In conclusion, donor's age affected the proliferation of cAD-MSCs and their ability to suppress T cell proliferation in our population of dogs undergoing abdominal surgery for reasons unrelated to the study. Donor's age also increased the expression of osteogenic genes by MSCs under inflammatory conditions. Combined, these results justify further investigation of allogenic cAD-MSCs from young dogs as a source of cells for regenerative medicine.
Footnotes
Acknowledgments
We acknowledge Ayman A. Mostafa (Cairo University), Kaitlin C. Clark, and Naomi J. Walker (University of California, Davis) for their technical help with experiments. We thank Drs. David Clark, Naomi Barnes, Omar G. El-Taliawi, and their staff, Leslie Tortez and others, at the three veterinary clinics which recruited donors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by AKC Canine Health Foundation Grant 2326-E and Western University of Health Sciences Intramural Grant.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.