
Abstract
Cell–cell communication plays a central role in the guidance of migrating neurons during the development of the cerebral cortex. Neuregulins (NRGs) are essential mediators for migration and maintenance of the radial glial scaffold. We show, in this study that soluble NRG reduces neuronal motility, causes transition of bipolar cells to multipolar ones, and induces neuronal mitosis. Blocking the NRG receptor, ErbB4, results in reduction of neuron–neuron and neuron–radial glial contacts and causes an increase in neuronal motility. Blocking the radial glial metabotropic glutamate receptor 5 (mGluR5), the nonselective cation channel transient receptor potential 3 (TRPC3), or matrix metalloproteinases (MMPs) results in similar effects as ErbB4 blockade. Soluble NRG counteract the changes in motility pattern. Stimulation of other radial glial G-protein-coupled receptors (GPCRs), such as muscarinic acetylcholine receptors or endothelin receptors counteract all the effect of mGluR5 blockade, but not that of ErbB4, TRPC3, and MMP blockade. The results indicate that neurotransmitters and endothelins acting on radial glial GPCRs are, through proteolytic NRG/ErbB4 activation, able to modify the migratory behavior of neurons.
Introduction
Control of neuronal migration through cell–cell interaction is essential during the development of cerebral cortex. Of particular importance is the neuronal guiding by the radial glial scaffold [1 –5]. This process is dynamic since the neurons detach from radial glial processes and may move independently for considerable distances and reattach or switch to move on other processes [6]. Transient interactions between neurons themselves have also been shown to be of importance in neuronal guidance and synaptogenesis [7]. A variety of factors has been implicated in glial–neuron interactions [8,9] and neuron–neuron interaction [7].
Several studies have shown that a family of growth factors, known as the neuregulins (NRG1–4) play a major role in neuronal migration [10 –13]. NRGs have a particular role in maintenance of radial glial integrity [10,12,14] and radial glial-assisted migration [11]. Neuregulins (1 and 3) and their receptors (ErbB3/4) have received a considerable attention due to the strong linkage of variants to schizophrenia [15,16]. Each member of this structurally related polypeptide family contain an epidermal growth factor (EGF)-like domain that can activate the ErbB family of tyrosine kinase receptors [17 –19].
The role of NRG in cell–cell interaction is controversial. In one study, interneurons moved toward NRG-producing cells [20], whereas in another study, NRG-producing cells exerted a repulsive effect [21] under apparently identical conditions. NRGs are activated by proteolytic cleavage leading to soluble or membrane-bound active forms. Consequently, NRGs may thus signal in a paracrine or juxtacrine manner [22,23]. G-protein-coupled receptors (GPCRs) and in particular, those coupling to Gq-type G-proteins have been demonstrated to cause transactivation of ErbB receptor family members through activation of proteolytic cleavage of the membrane-anchored ligand [24 –27]. ErbB transactivation by GPCRs seems to require protein kinase C (PKC) [28] and/or elevations in intracellular Ca2+ concentrations ([Ca2+]i) [29,30]. Activity-dependent proteolytic release of NRG1 has also been demonstrated [31 –33], and metabotropic glutamate receptors promote shedding of the NRG ectodomain [34].
The metabotropic glutamate receptor 5 (mGluR5) has emerged as being essential for normal neurogenesis ([35 –37], reviewed in Jansson and Akerman [38]). This receptor is predominantly located on radial glial processes and promotes process extension [14,39,40]. The mGluR5 is coupled to the activation of a nonselective cation channel, canonical transient receptor potential 3 (TRPC3) [40 –42], which has previously been shown to act as a signal for chemotaxis [43,44]. Inhibition of mGluR5, loss of TRPC3, or its inhibition by the selective blocker pyr3, leads to a reduction in radial process length, detachment of neurons from these processes, and an increase in neuronal motility [40].
Considering the important role of neuregulins in neuronal development and neuropsychiatric disease, it is of importance to delineate the actions of neuregulins at the cellular level. The aim of the present study was to investigate a possible role of neuregulins acting on ErbB receptors as possible mediators of mGluR5/TRPC3-promoted cell–cell contact and regulation of neuronal motility.
Materials and Methods
Cell culture and neuronal differentiation
Neural progenitor cells (NPCs) were isolated from the walls of the lateral ventricles of embryonic day 14.5 129SvJ/C57Bl6/N mice, as described previously [45]. Mice were group housed in standard laboratory conditions in qualified animal facilities, in accordance with the guidelines of the National Institute of Health. All animal experiments were performed according to the Guide for the Care and Use of Laboratory Animals and European Economic Community Council Directive. The protocols were approved by the Experimental Animal Ethics Committee of the National Laboratory Animal Center, Finland. Cells were cultured as neurospheres in proliferation media: Dulbecco's modified Eagle's medium: Ham's Nutrient Mixture F-12 media (1:1) containing B27 supplement (both from Gibco, Life Technologies Ltd., Paisley, UK), 2 mM
Immunocytochemistry
NPCs were differentiated for 3 days on poly-
Primary antibodies used were mouse monoclonal anti-microtubule-associated protein (MAP)-2 (MAB364, 1:500; Millipore), mouse monoclonal anti-ErbB4 (HFR-1, Ab19391, 1:100; Abcam) received as a kind gift from Dr. Klaus Elenius, rabbit polyclonal anti-EAAT1 (excitatory amino acid transporter 1, also known as glial glutamate–aspartate transporter (GLAST) (ab416, 1:500; Abcam), and rabbit polyclonal anti-HER4/ErbB4 (clone E200, 1:100; Merck Millipore, Germany). Secondary antibodies used were Alexa Fluor 488 donkey anti-rabbit IgG (A11008, 1:400), Alexa Fluor 488 donkey anti-mouse IgG (R37114, 1:400), Alexa Fluor 546 donkey anti-goat IgG (A11056, 1:400), and Alexa Fluor 546 donkey anti-mouse IgG (A10036, 1:400; Life Technologies Ltd.). Cells were photographed using an Olympus AX70 Provis microscope (Olympus, Espoo, Finland), equipped with fluorescence optics and a charge-coupled device camera (PCO AG, Kelheim, Germany). Brightness and contrast of the obtained images were adjusted with Corel Draw Graphics Suite X7 (Corel Cooperation, Ontario, Canada).
Time-lapse imaging
Time-lapse imaging of cellular movement was performed in a real-time cell monitoring system, Cell-IQ® system (Chip-Man Technologies Ltd., Tampere, Finland), which is equipped with a phase-contrast microscope, camera (Nikon, Japan), and Cell-IQ cell-culturing platform equipped with two plate holders. The equipment (including automation and environmental regulation) was controlled by Imagen software (Chip-Man Technologies). The time-lapse images were analyzed with ImageJ and its plugin MJtracker and the data quantified with Excel (Microscoft, Redmond, WA) and Origin 6.0 (OriginLabCorp., Northampton, MA). After initializing the imaging sequence, a minimum of a 10-h time period was used before calculating the motility index. The motility index parameter of migratory cells was defined as the number of time points the cell moved with a speed greater than 30 μm/h divided by the number of time points the cell speed was below this value.
Calcium imaging
Ca2+ imaging experiments were performed by plating ∼20–30 neurospheres on poly-
Gene expression analysis
Cell type-specific gene expression was performed on our genes of interest using a whole-genome mouse embryonic single cell RNA sequencing dataset (GEO Accession No. GSE119945) recently published by Cao et al. [46]. From the dataset, the unique molecular identifiers' (UMIs) count as well as the filtered high-quality cells clustered according to cell type and subtype (ie, after removal of cells with <400 mRNAs and detected doublets) were downloaded (
where Xi
equals the UMI count of each cell in the subtype i,
Statistical analysis
Before performing the statistical tests, the Shapiro–Wilks test was applied to assess the normality of the data distributions, and equal variance using Levene's test. Data were analyzed using either Student's unpaired t-test or a one-way ANOVA (analysis of variance) with Tukey's post hoc analysis, or Mann–Whitney U test. The Bonferroni correction was applied to maintain an overall type I error rate of 0.05 against multiple comparisons. Data are presented as mean ± standard error of the mean. Significances are as follows: ns (not significant) P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001.
Results
ErbB4 immunostaining in E14.5 neural progenitor cells (NPCs)
To visualize the migratory behavior of NPCs, time-lapse microscopy was used to document the behavior of NPCs as they migrated out from neurospheres. Thick radial processes extended from the neurosphere forming a dense layer, consisting of a mixture of radial glial type processes and developing neuronal cells (Fig. 1A). The radial processes have previously been shown to stain with markers for radial glia (GLAST, BLPP), mGluR5, and ErbB1 (EGFR) [48 –50]. The individual neuron-like bipolar cells (marked as N) frequently exit from this layer, move in an apparent random fashion. We have shown previously [40] that even if the “free” cells appear to move randomly, they tend to move close to the radial layer (marked by dotted line) and make frequent contacts with the tips of radial processes. Contacts are shown with arrows in Fig. 1A.
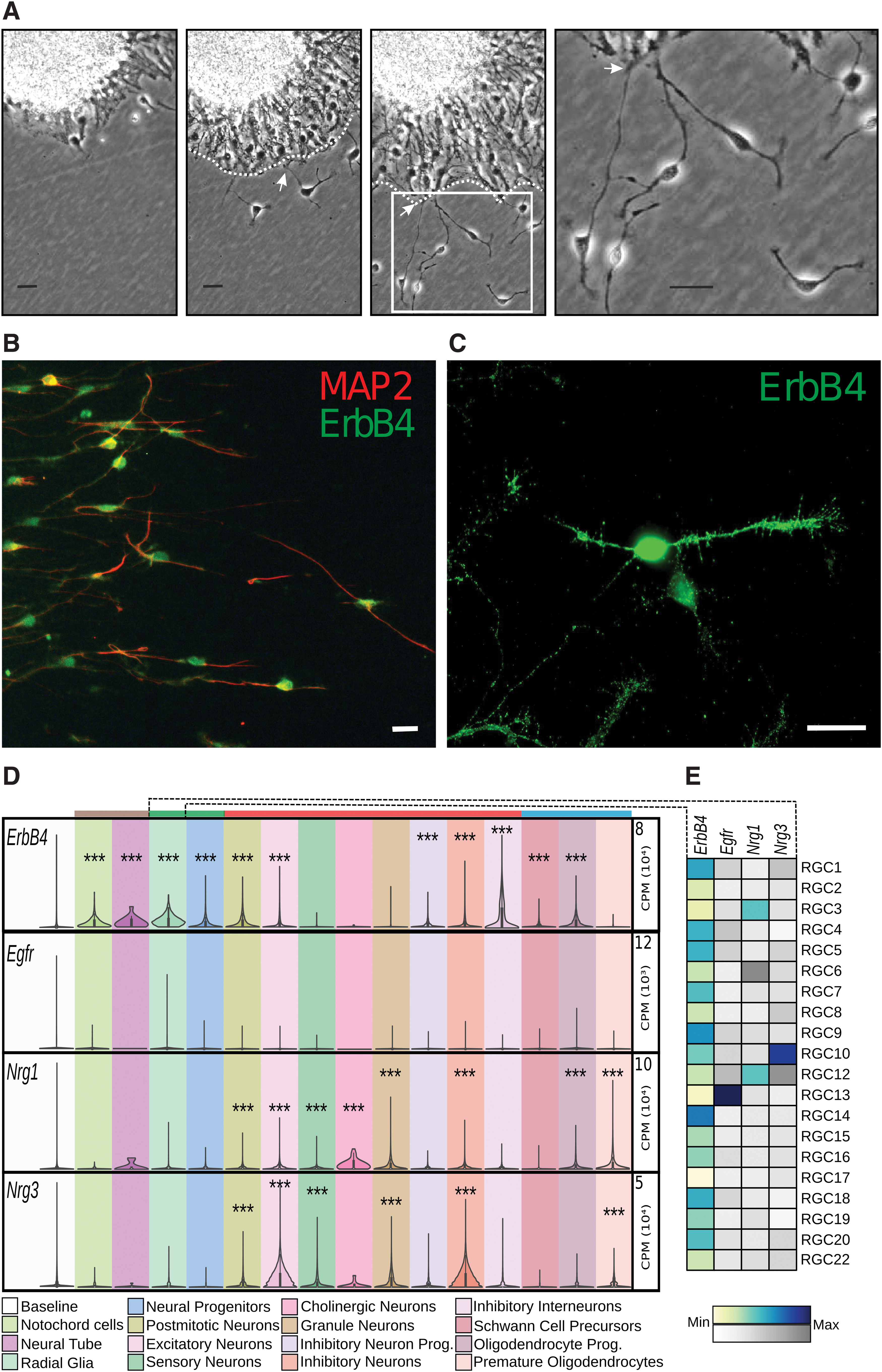
Morphology and ErbB4 immunostaining of neurosphere-derived cells. In
We have previously shown that mRNA of Erbb4 but not that of Erbb3 is expressed in both proliferating and differentiated neurosphere-derived NPC and that Nrg3 is the most prevalent neuregulin [14]. Antibodies directed against the ErbB4 receptor were used to localize receptor expression. ErbB4 receptor colocalized in cells expressing the neuronal marker MAP-2 as seen in Fig. 1B. As seen in the higher magnification image in Fig. 1C, cell neurites as well as soma were positively stained. No detectable signal was seen in negative controls without primary antibody when fluorescence was observed at the same exposure times as with the primary antibody.
Using publicly available single cell RNA-sequencing datasets, we set to analyze the cell type-specific gene expression of ErbB4 as well as Nrg1 and Nrg3. Recently, the transcriptomic profiling of the developing whole mouse embryo (E9.5–E13.5) was released [46]. Given the time point of our in vitro experiments, we chose to focus on E13.5 as it was closest temporally. From the E13.5 dataset, the clustered annotations (hereby marked in quoted italics) of the cleaned high-quality cells and their UMIs were downloaded and normalized. This resulted in a dataset of roughly 270,000 cells containing 37 main cell types and multiple subtypes, ranging from 3 to 27 per main cell type, from which we extracted our genes of interest and analyzed all cells that had been annotated as belonging to a cell type that is directly related to the nervous system.
The mRNA baseline expression distributions were generated by pooling the expression values for each cell type contained in the dataset. To determine if gene expression was significant, the main cell types were compared with the baseline expression. As seen in Fig. 1D, ErbB4 was significantly expressed in most of the main cell types with the exception of the “Sensory,” “Cholinergic,” and “Granule” neurons as well as the “Premature oligodendrocytes.” Nrg1 and Nrg3 showed similar patterns in their expressions, with the developed neuronal cell types and some glial cells showing significant levels of expression (with the exception of the “Cholinergic neurons” for Nrg3 and “Inhibitory interneurons” for both). Interestingly, the expression of Egfr was found to be nonsignificant for all main cell types, although some main cell types, such as “Radial Glia,” showed high variations in their expressions, suggesting that Egfr may be highly expressed in a subset of the samples and masked by the low expressions in other subsets.
As such, we split each main cell type according to their subtypes and repeated the analysis for the aforementioned genes, to assess if some subtypes significantly expressed the genes of interest. Consequently, we found that 1 out of 15 subtypes of “Premature oligodendrocytes” significantly expressed ErbB4, while none of the “Sensory” (0 out of 16 subtypes), “Cholinergic” (0 out of 4 subtypes) or “Granule” (0 out of 16) neuron subtypes did.
This is contrasted by all of the “Radial Glia” subtypes (21 out of 21 subtypes), which significantly expressed ErbB4, as shown in Fig. 1E, suggesting that ErbB4 is abundantly expressed in RGCs during development. Furthermore, as shown in Fig. 1E, one subtype of “Radial glia” significantly expressed Egfr. This was matched by only two other subtypes in total, from “Notochord cells” (1 out of 15 subtypes) and “Oligodendrocyte precursor cells” (1 out of 19 subtypes), and in none of the neuronal cell types.
For Nrg1 and Nrg3, “Radial glia” were the only main cell type to have subtypes that significantly expressed both despite not being significant at the pooled main cell-type level (Fig. 1E). The others either expressed one of the genes, or none. For example, the “Neural progenitor cells” had 3 out of 27 subtypes that significantly expressed Nrg1, whereas none expressed Nrg3. “Neural tube,” “Schwann cells,” and “Notochord cells” did not display any significant levels of Nrg1 or Nrg3. A similar pattern is seen in the analysis of single cell RNA sequencing data (see Supplementary Data) from the developing mouse midbrain transcriptome [51] indicating that ErbB4 mRNA, and to a lesser extent Nrg3 mRNA, are present in radial glia and both RNAs abundantly expressed in neurons while the EGF receptor (EGFR) ErbB1 was expressed exclusively in radial glia (Supplementary Fig. S1).
ErbB4 plays a role in neuronal–radial glial contacts
Time-lapse images of control cultures, NRG-treated cultures, and cultures treated with afatinib (ErbB4 receptor blocker) were analyzed to assess the role of NRG in neuron–radial glia interactions. Fewer contacts were seen in the presence of the soluble EGF-like domain of NRG1 (sNRG1), seen as an increase in the number of cells detached from the radial processes (denoted * Fig. 2A). Treatment with afatinib also increased the number of cells outside the radial layer (Fig. 2A). The number of freely moving cells detached from radial processes were quantified over a time period of 25 h. Both soluble sNRG1 and afatinib caused a significant increase in nonattached cells during this time period (Fig. 2B). The 2-methyl-6-(phenylethynyl)pyridine (MPEP), a selective blocker of mGluR5, also caused detachment of neuronal cells from radial glia (Fig. 2B) as shown earlier [40].

Effect of NRG/ErbB4 on the spatial distribution of neuronal cells with respect to processes of radial glial type cells. In
The culture medium (see Materials and Methods section) generally used for neuronal progenitor cell culture contains about 25 μM glutamate as an essential component (according to the manufacturer's information). This concentration should be enough to cause a considerable activation of mGluR5 [52], which explains the constitutive activity and effects of MPEP. Mouse and human neural progenitor cells show robust Ca2+ responses also to muscarinic receptor stimulation in the same cells that also respond to mGluR5 stimulation [14,53,54].
It was therefore of interest to test whether stimulating with a muscarinic receptor agonist would be able restore the radial process interaction in MPEP-inhibited cells, thus replacing mGluR5 with respect to motility. As shown in Fig. 2B, oxotremorine M (oxoM) caused a significant decrease in the number of unattached neuronal cells in MPEP-treated cultures to the same level as in control cells.
The data presented above suggest that interaction with radial glia might interfere with the free movement of neuronal cells. To verify this, the spatial distribution of neuronal cells with respect to radial glial processes was determined. Figure 2C shows the spatial distribution of 25 control, 25 sNRG1-treated, and 25 afatinib-treated cells during a period of 10 h. Note the much wider area of distribution of the afatinib-treated cells. The maximal distance each cell had moved from the radial layer after exiting from the neurosphere was then determined.
As seen in Fig. 2D, both control and sNRG1-treated cells moved to an average distance of about 150 μm from the radial processes, while afatinib-treated cells moved much further (on average close to 400 μm). MPEP-treated and cells treated with pyr3, a blocker of nonselective cation channels TRPC3, behaved like afatinib-treated cells. They distributed maximally to a distance of 300–400 μm from the radial tips. In the additional presence of sNRG1, the cell moved closer to the radial processes (around 200 μm). sNRG1 did not affect the distribution of afatinib-treated cells.
Multipolar transition in NRG-treated cells
A closer inspection of the morphology of sNRG1-treated cells revealed a frequent transition of bipolar neuronal cells into cells with multiple processes. In vivo multipolar cell morphology represents a transit state of migration. Multipolar cells accumulate at the intermediate zone, stop migrating, divide, and finally after a time period, proceed migration leaving an axon behind ([55,56], reviewed in Cooper [57]). Under the control conditions, multipolar cells could usually be observed close to the border of radial process tips (Supplementary Video S1). Time-lapse images showing the transition of bipolar cells near the radial layer into multipolar ones (denoted *) and one of them finally dividing to two daughter cells (denoted #) is shown in Fig. 3A. Statistical analysis confirmed that addition of sNRG1 induced considerable increase in the number of multipolar cells (defined as neuron-like cells with four or more processes) (Fig. 3B). Afatinib caused a significant reduction in multipolar cells (Fig. 3B). Since Nrg3 is the dominant neuregulin in these cultures [14], its effect was tested. A soluble analog of NRG3 caused a modest increase in the number of multipolar cells (Fig. 3C). Blocking mGluR5 with MPEP considerably reduced the multipolar transition. The number of multipolar cells, in the presence of MPEP, was restored by the soluble analog of NRG3. In addition, stimulation of muscarinic receptors with oxoM had a restoring effect.
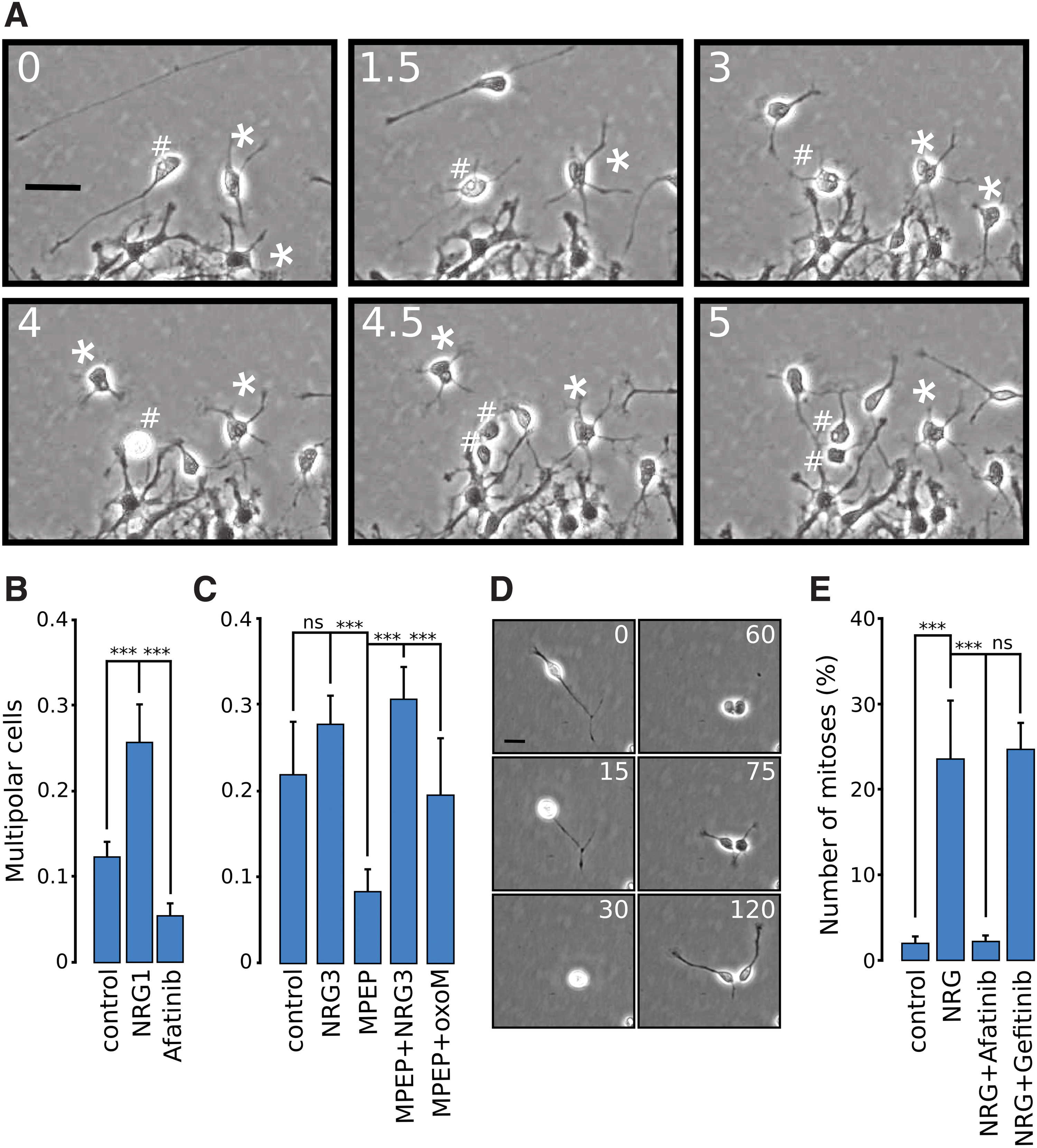
Multipolar transition and mitosis in the presence of NRG. In
Neuronal division in NRG-treated cells
Mitoses of neuronal cells were seldom seen in control conditions in the time-lapse image sequences. In NRG-treated cultures, however, dividing bipolar neuronal cells were frequently observed. A division of a typical bipolar cell resulting in two bipolar daughter cells is shown in Fig. 3D. Statistics on the number of neuronal divisions indicated near 10-fold increase in mitoses, whose effect was blocked by the ErbB4 blocker afatinib, but not by ErbB1 blocker gefitinib (Fig. 3E).
Blocking ErbB4 increased the motility index of migrating NPCs
Since NRG is essential for neuronal migration [11], we proceeded to determine its effect on neuronal motility using time-lapse imaging [40]. Images were taken every 15 min for a time period of 48 h. Figure 4A shows the spatial pattern of movement for a representative cell. The time points from the start of the analysis (h) are shown on the images. As seen, the cell moved in phases of rapid linear progression with alternating slow phases during which it frequently changed direction. During these slow phases, the bipolar cells retract their long processes, extend short processes in many directions, and rotate in a slow fashion (Fig. 4A images on the right). The speed as a function of time for this cell is shown in Fig. 4B. Note the phasic motility pattern with phases of high speed and alternate slow phases.
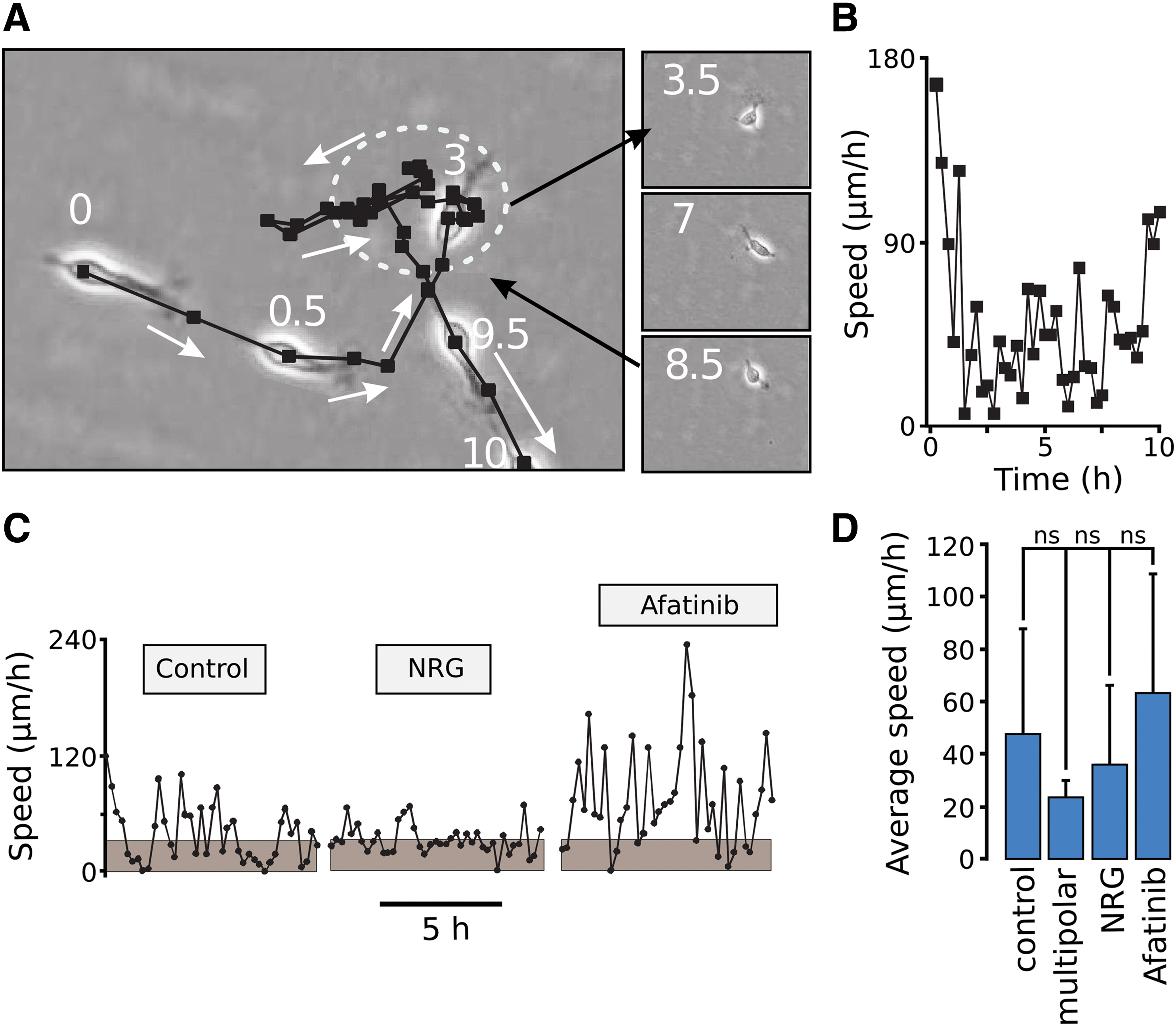
Effect of NRG/ErbB4 on the motility of neuronal cells. In (
These observations are in agreement with previous work done on the pattern of migration of subventricular zone neurons in slices [58,59]. The speed as a function of time of a control cell, sNRG1-treated cell and a cell treated with afatinib are shown in Fig. 4C. Note the frequent periods of stalling in control cells (gray areas denote speed below 30 μm/h) and increased stalling in NRG-treated cells. The afatinib-treated cell stalled less frequently and showed an overall higher speed. Statistics for the average speed of movement for control (bipolar) cells, multipolar cells (ie, cells with four or more processes during the period of tracking), NRG-treated cells, and afatinib-treated cells was then determined. The average speed of the multipolar cells was low, NRG treatment showed a tendency of a reduction in speed while afatinib showed an apparent increase (Supplementary Video S1). No significant differences between these groups could, however, be observed.
Since the type of the cellular movement is phasic in nature, the average speed of movement is not an appropriate measure of cell motility [40]. Therefore, the motility was quantified as the motility index, defined as the frequency by which cells move with a speed above 30 μm/h divided with the frequency by which the cells move below this value [40]. As shown in Fig. 5A, NRG treatment caused a small reduction in the motility index, while afatinib considerably increased it. NRG did not affect the afatinib-induced increase in motility index as expected. Compared with control cells, treatment with 100 nM gefitinib did not produce significant difference in the motility index in the neuronal cells outside the radial glial network (Fig. 5A). This is in stark contrast to the effect of blocking with afatinib. Adding NRG with gefitinib, if anything, showed a tendency of an increase in neuronal motility.
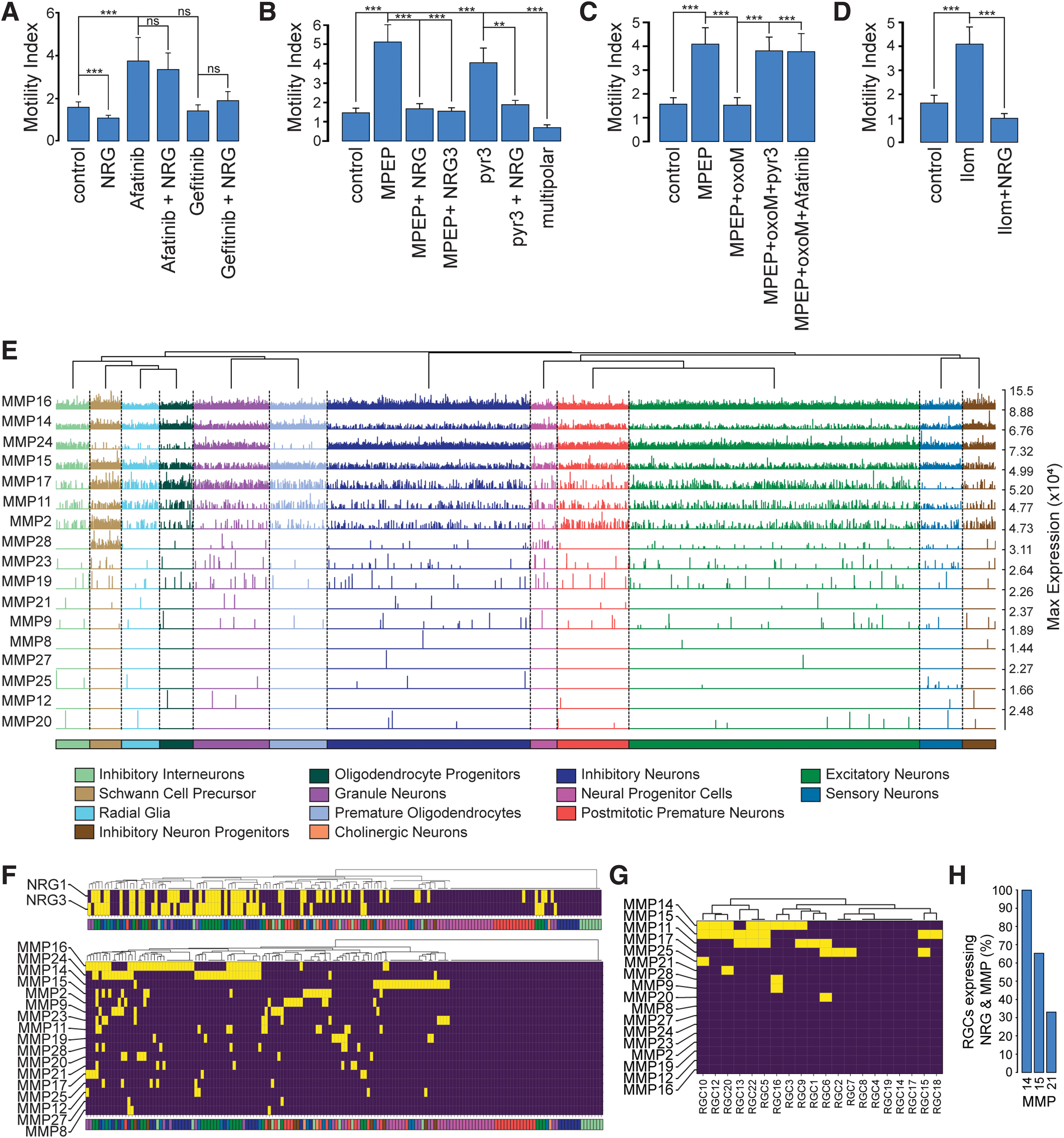
Effect of NRG/ErbB4 on the motility index of neuronal cells and MMP expression. In (
Role of NRG in the motility pattern changes brought about by blocking mGluR5/TRPC3
The increased motility index seen in the presence of afatinib is reminiscent of the effect seen in cells when mGluR5 receptors is blocked by MPEP [39] and in cells lacking functional nonselective cation channels TRPC3, or in the presence of the TRPC3 channel blocker pyr3 [40]. It was therefore of interest to determine whether NRG would affect the motility pattern caused by these treatments. sNRG1 treatment indeed reversed the MPEP and pyr3 induced increase in motility to control values (Fig. 5B; see Supplementary Video S1). As already shown above in Fig. 5A, sNRG1 did not affect the motility increase seen with afatinib. The soluble analog of NRG3 had a similar effect as sNRG1. sNRG1 also had a similar effect on the increased motility of cells from TRPC3/6 knockout mice (data not shown). The motility index of multipolar cells was significantly lower than control bipolar cells.
Next, we tested whether muscarinic receptor stimulation would be able to replace mGluR5 with respect to motility. As shown in Fig. 5C oxoM caused a significant decrease in the motility index of MPEP-treated neuronal cells to the same level as in control cells. The reduced motility was increased back again by pyr3 and afatinib to about the same level seen in cells treated with MPEP, pyr3, or afatinib alone.
The results above indicate that GPCRs (mGluR5 and muscarinic acetylcholine receptors) modify neuronal motility by a mechanism involving NRG. GPCRs causes transactivation of ErbB receptors through Ca2+ and PKC-mediated activation of matrix metalloproteinases (MMPs). MMPs subsequently activate membrane-bound, inactive forms of ErbB receptor ligands (NRGs) through proteolysis, thus causing receptor transactivation [25,60]. This transactivation can be inhibited by broad-range MMP blockers such as ilomastat [61]. Figure 5D shows the effect of ilomastat on neuronal motility. A considerable increase in motility reminiscent of that seen upon blocking mGluR5/TRPC3 and ErbB4 was observed. This effect was reversed by sNRG1, suggesting that MMPs are involved in the observed motility changes.
Using the publically available whole mouse brain embryo dataset, we set out to identify likely MMP candidates by analyzing the cell type distribution of MMP expression from the E13.5 developing whole mouse embryo [46]. In this study, we took a similar approach as for Erbb4, Nrg1, and Nrg3. However, as we were primarily interested in the patterns of expression in the neuronal cell types, we excluded the non-neuronal cell types from the baseline expression (ie, “White Blood Cells,” “Osteoblasts,” etc…).
The MMP family consists of a total of 28 family members [62], 17 of which were present in the E13.5 RNA-seq dataset. The expression levels of all 17 Mmps and their cell type distribution is shown in Fig. 5E. Of the 17 Mmps, Mmp16, Mmp24, Mmp14, Mmp15, Mmp17, Mmp11, and Mmp2 showed a relatively higher expression across all central nervous system (CNS) cell types in comparison to the other remaining Mmps, which had less frequent expression across and within the cell types (Fig. 5E). Shown in Fig. 5F, using P < 0.05 as a cutoff filter, is a binarized heatmap of the expression levels of Mmps shown on a cell subtype-specific level for all the neuronal subtypes, with “Radial Glia” subtypes and MMP expression shown in Fig. 5G (see Fig. 1E for Nrg expression in “Radial Glia” subtypes). As seen in Fig. 5F, Mmp16 and Mmp24 were the dominant Mmps found in the neuron subtypes (ie, “Excitatory neurons” and “Inhibitory neurons”). While in contrast, Mmp14 and Mmp15 were the dominant Mmps for “Radial Glia” subtypes, with no significant expression above the baseline distribution for Mmp16 or Mmp24 (Fig. 5G). Of the “Radial Glia” cell subtypes, which significantly expressed Nrg (Fig. 1E), all expressed Mmp14 (three out of three subtypes), with Mmp15 expressed in two out of three subtypes and Mmp21, one out of three “Radial Glia” subtypes (Fig. 5H). Interestingly, we observed that out of all the cell subtypes (Fig. 5F, G) that significantly expressed Nrg also had a significant expression of at least one member of the MMP family, with only 4% expressing Nrg1 or Nrg3 with no significant expression of any member of the MMP family, supporting the important role of MMPs for NRG function.
Effect of endothelin on the motility of neuronal cells
The endothelin B (EDNRB) receptor has been used as one of the markers for radial glia [51]. Supplementary Figure S2 shows the distribution of the mRNA for endothelin 1 (Edn1), Ednra, and Ednrb receptors in different subpopulations of radial glia and neurons. The mRNAs of both receptors are exclusively expressed in radial glia and no signal in neurons is observed, while Edn1 was exclusively expressed in pericytes. Like mGluR5 (and M1, M3, and M5 muscarinic receptors), EDNRB receptors are Gq-coupled and their activation causes mobilization of intracellular Ca2+ in Bergmann glia, a cerebellar analog of radial glia [63].
It was therefore of interest to see whether endothelin receptor stimulation would replace mGluR5 in MPEP-treated cells. Ca2+ imaging was initially used to characterize endothelin-responding cells. The cells were challenged with 10 nM endothelin, 2 ng/mL EGF, and 50 μM kainate (an agonist of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)/kainate receptors, which fails to desensitized response AMPA receptors resulting in prolonged responses), as a marker of neuronal cells [39]. Ten nanomolars of endothelin caused a robust Ca2+ response in most cells and the same cells responded to EGF but not to kainate (Fig. 6A). Kainate-responding cells on the other hand did not respond to endothelin or EGF. The radial glial network appeared distorted upon prolonged treatment with 10 nM endothelin. Therefore, lower concentrations were tested. Figure 6B shows that the cells responded to concentrations as low as 0.1 nM endothelin. Treatment of cells with 0.1 and 1 nM endothelin had no obvious effect on cell morphology. A concentration–response curve for the effect of endothelin on intracellular Ca2+ is shown in Fig. 6C with comparison to the response of stimulating the same cellular batch cells with 10 μM DHPG, an agonist of mGluR5, and with 2 ng/mL EGF.
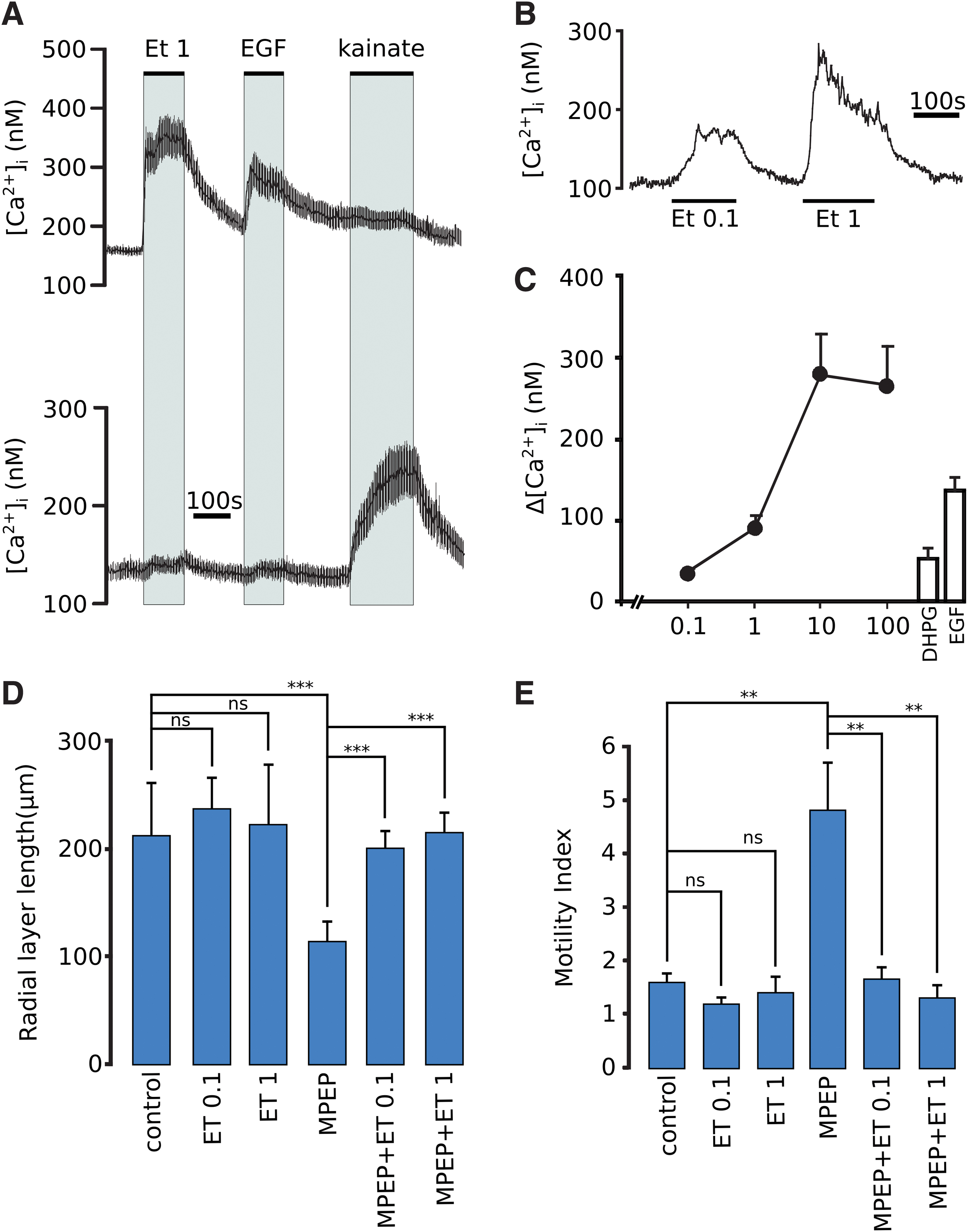
Effect of endothelin on intracellular Ca2+, radial process extension and the motility of neuronal cells. In
We have previously shown that MPEP reduces the length of radial glial processes and that muscarinic stimulation restores process elongation [14]. As shown in Fig. 6D, MPEP reduced the length of radial processes and this effect was reversed by 0.1 and 1 nM endothelin. Furthermore, the increased motility in MPEP-treated cells was reduced to the control level by endothelin (Fig. 6E).
Effect of NRG on the distribution of stalling cells
As shown above, sNRG1 treatment has a tendency to promote phases of slow motility or stalling. Our earlier studies indicate that stalling periods frequently coincide with interactions of neuronal cells with radial glial processes [40]. An image sequence of a neuronal cell making contact with radial processes and finally being released after 5 h is shown in Fig. 7A. As shown in Fig. 7B, the majority of stalling control cells were distributed very close to the tips of radial processes and seldom found further than 150 μm from these processes indicating that radial processes have an impact on the motility of the cells. In the presence of NRG, stalling cells accumulated at a distance around 100 μm from the radial processes while stalling afatinib, MPEP and py3-treated cells showed a diffuse distribution at distances between 0 and >400 μm with no clear peaks. With additional sNRG1 treatment, the stalling cells showed a similar distribution as in the presence of NRG alone (about 100 μm from the radial processes). Interestingly, in the presence of oxoM, MPEP-treated cells showed a similar distribution as control cells with the majority of stalling cells residing very close to the radial processes. There was, however, no change in the distribution of pyr3-treated cells in the presence of oxoM (data not shown).
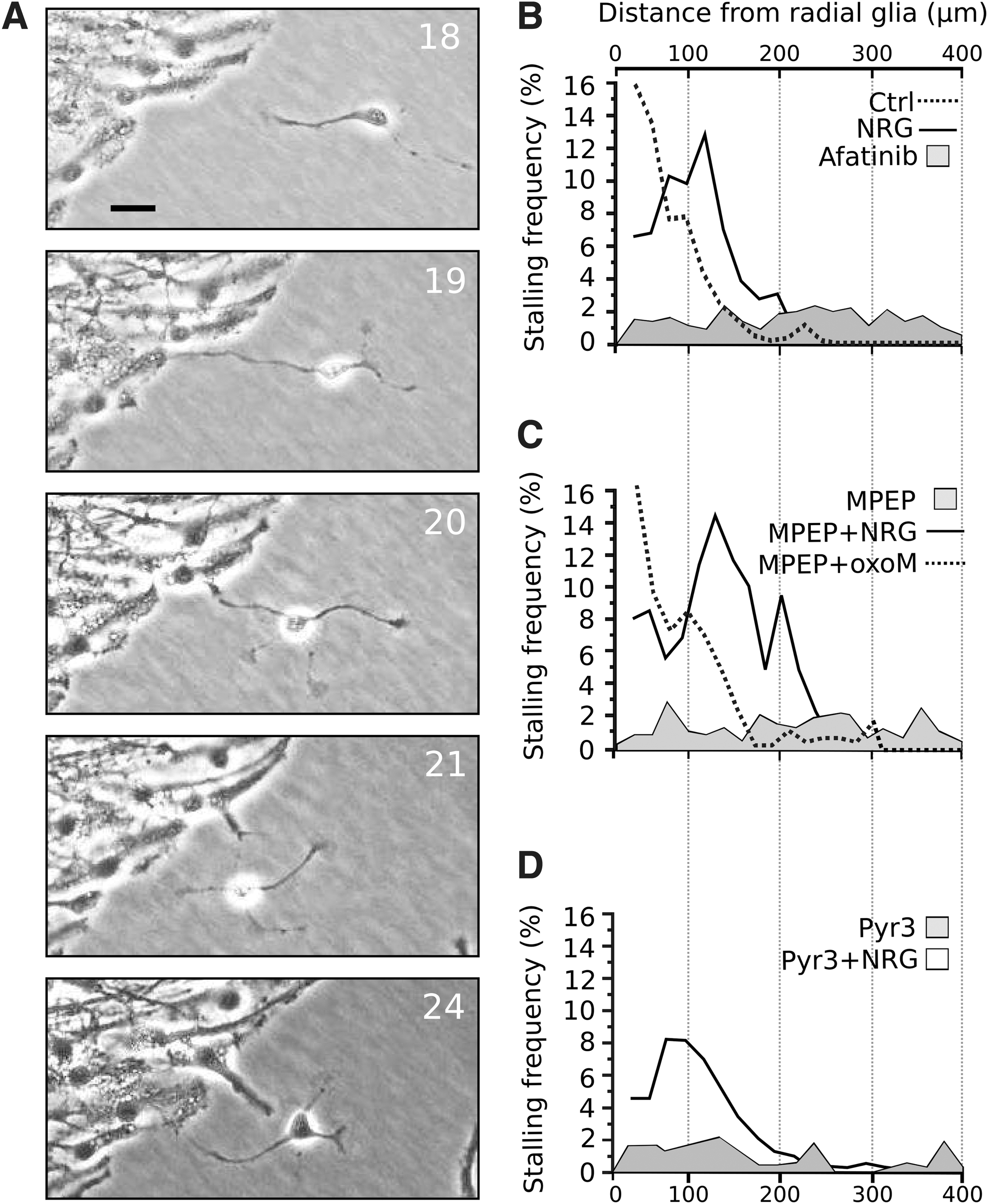
Effect NRG/ErbB4 on the spatial distribution of stalling cells. In
Effect of NRG on neuron–neuron interaction
The reason for the high incidence of stalling cells around 100–200 μm from radial processes is not obvious. Interactions between neuronal cells exiting from the radial glial layer could be one of the factors restricting the motility. An image sequence of a contact between two neuronal cells is shown in Fig. 8A. The duration of each contact is 1.0 ± 0.4 h (± SD n = 23 neurospheres). The number of contacts per total number of “free” neuronal cells was then quantified. NRG did not significantly affect the number of contacts between cells in control conditions (Fig. 8B). Afatinib, MPEP, and pyr3 significantly reduced the number of neuron–neuron contacts. Soluble NRG did not restore neuron–neuron contacts in any of these conditions. The contacts in the presence of MPEP were however, restored in the presence of oxoM.
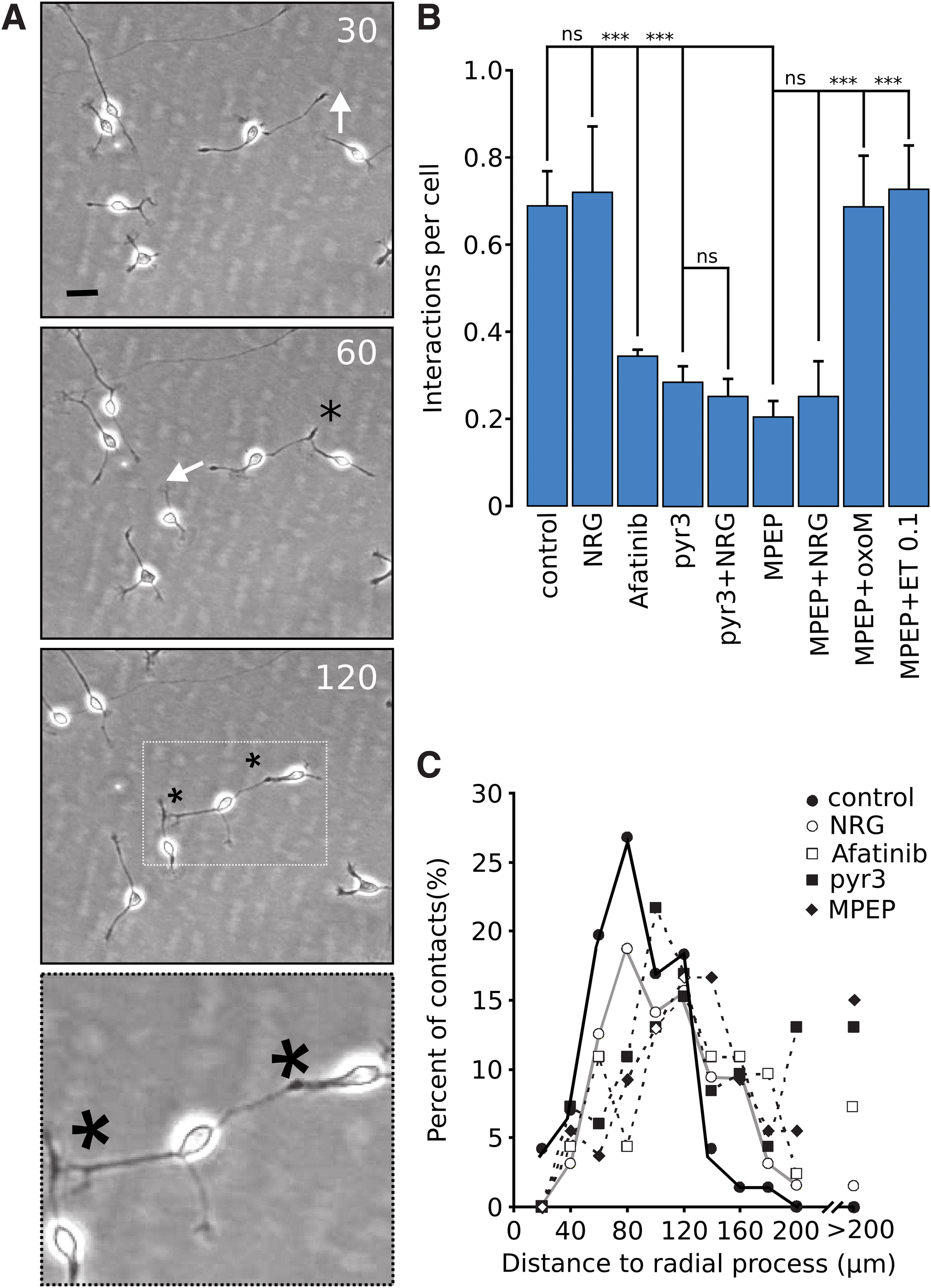
Role of mGluR5/TRPC3, NRG/ErbB4 in neuron–neuron contact. In
The number of neuronal-neuronal contacts were then quantified as a function of distance to radial glial processes in control and NRG-treated cells. Most contacts occurred between 50 and 100 μm from nearest radial processes (Fig. 8B). In afatinib-treated cells and in MPEP/pyr3-treated cells, the neuron–neuron contacts show a peak around 100 μm (Fig. 8C) while some contacts could be seen at a distance above 200 μm with these treatments.
Discussion
The exact positioning of neuronal cells in the developing cerebral cortex requires communication between neurons and their radial glial scaffold [64] on the one hand and neuron–neuron contact on the other [7]. The NRG/ErbB4 signal pathway is essential as a mediator of communication between radial glia and neuronal cells [11,65]. Previous studies have shown that also in vivo, neurons detach and reattach to radial processes during migration [6]. Our data suggest that cell–cell contacts of this type are controlled by GPCRs through proteolytic activation of NRG/ErbB4 signaling. As such, our results favor a scenario in which activation of the dominant mGluR5 in the early stages of NPC migration and differentiation is a crucial step in the release and processing of NRG, which in turn directs and manages neuronal–radial glial interaction through ErbB4 [14]. The abundant presence of ErbB4 receptors and the lack of ErbB3 expression in cortical progenitor cells suggest that the actions of NRG in developing cortical neural progenitor cells occur through ErbB4 [14]. This is in agreement with previous ErbB receptor expression studies on mice showing that ErbB3 mRNA is hardly detected in cortical regions during early brain development, whereas ErbB4 expression was most prominent, becoming more restricted postnatally [66,67].
NRG has previously been shown to be essential for correct neuronal migration [11,65]. Neuregulins exist in several different membrane-bound forms [13]. Some forms of NRG (type 1/2) are substrates for cleavage by sheddases leading to the release of soluble forms of NRG, whereas some cleaved forms remain in the membrane (type 3). Thus NRG-ErbB interactions are mediated either through a soluble or membrane-bound forms of NRG, for example, interactions can also occur within the framework of cell-to-cell contact [22,23].
Although NRG is essential for correct neuronal migration, the mechanism by which NRG controls migration remains controversial. In one study, interneurons moved toward NRG-producing cells [20], whereas in another study, NRG-producing cells exerted a repulsive effect [21] under seemingly identical conditions.
In this study, addition of the soluble EGF domain of NRG caused neurons to detach from radial processes. Blocking of ErbB4 receptors with afatinib also resulted in detachment of neurons. This apparent discrepancy that both stimulation and inhibition of ErbB4 results in detachment would be that added soluble NRG1 competes with membrane-associated forms of NRG for docking at ErbB4. The distribution of the neuronal cells with respect to radial glia was, however, different, while sNRG1 promoted stalling and cells resided close to radial processes, afatinib treatment caused an increase in motility and a distribution of the neuronal cells away from the radial glial processes.
The balance between secreted and membrane-bound forms of NRG in addition to motogenic factors would thus determine the distance between radial processes and neurons. This may explain those differences concerning chemoattraction/repulsion mediated by NRG discussed above. The main effects of soluble NRG observed in this study were a reduced neuronal motility by increasing the frequency of slowly moving phases or stalling. Studies done in vivo suggest that stalling phases in NPC migration represent periods of exploration [68]. In this phase of migration, cells often extend several processes and rotate slowly [39,40,55,69]. The most prominent change seen in the presence of NRG was the multipolar transition. In vivo multipolar cells have been described as a transient progenitor population. Multipolar cells accumulate in the intermediate zone, move slowly and randomly, divide, and finally resume a bipolar migration leaving the axon terminal behind ([3,55], reviewed in Cooper [57]). The role of NRG as a mediator of this transition in vivo according to the authors' knowledge has not been explored. In addition, NRG promoted symmetric neuronal divisions. Symmetrical divisions are of importance in expanding and producing diversity in neuronal populations [70]. Several factors, including ligands of GPCRs, have previously been shown to promote neuroblast divisions.
We have previously shown that blocking the glutamate receptor mGluR5 or the nonselective cation channel TRPC3 with pyr3 or deletion of the Trpc3 gene [40] results in neuronal detachment from radial processes and an increase in neuronal motility in a similar way as blocking ErbB4 as shown in this study. Although the effect of NRG on neuronal motility was modest under basal conditions, the increased neuronal motility seen by blocking mGluR5 with MPEP or interference with TRPC3 function was completely reversed to control levels in the presence of NRG. The low motility and modest effect of NRG in control conditions is thus probably due to endogenous activation of NRG/ErbB4 through inherent mGluR5/TRPC3 activity.
One likely mechanism by which Gq-coupled mGluR5 could activate NRG/ErbB4 is by transactivation through a cleavage of membrane-bound NRG isoforms by sheddases to release membrane-bound and soluble active NRG locally. This has been described for other ErbB receptor ligands [25,60]. Transactivation is sensitive to blockers of MMPs such as ilomastat. The ilomastat-increased motility seen in this study, and the reduction of the ilomastat-increased motility in the presence of soluble NRG, supports the mechanism involving cleavage of membrane-bound NRG.
Furthermore, stimulation of muscarinic receptors with oxoM, when mGluR5 was blocked with MPEP, restored the low motility index seen in controls and even restored neuron–neuron contacts. This suggests that any Gq-coupled receptor, which is abundantly expressed, would be able to replace mGluR5 and cause ErbB4 transactivation in a pyr3 and afatinib-sensitive manner. Muscarinic receptor-mediated Ca2+ responses are seen in neural progenitor cells in the same cells that respond to mGluR5 activation [53,54]. These results strongly suggest a scenario where neurotransmitters released from developing neurons would, through radial glial GPCRs, activate NRG through proteolytic cleavage. This in turn would induce the stalling of nearby neurons and cause them to remain in close proximity to the radial glial process, possibly attaching to membrane-bound forms of NRG [71]. Since soluble NRG was unable to restore neuron–radial glial and neuron–neuron interactions in MPEP-treated cells, it appears likely that these interactions are mediated by membrane-bound forms of NRG. OxoM by signaling through muscarinic receptors to MMPs, on the other hand, would be expected to restore functions of both soluble and membrane-bound NRG.
The analysis of the transcriptomic data from radial glia and neurons suggests that these cell types express different subtypes of membrane-bound matrix metalloproteases, which have been shown to be involved as sheddases (reviewed in Refs. [72 –74]). Mmp14, Mmp15, and Mmp21 seemed to show higher frequency of expression in radial glia, whereas Mmp24 and Mmp16 showed higher frequency in neuronal cells. MMP14 (and possibly MMP15) is the most likely candidate involved in the results presented in this study, since its role in activation of ErbB receptors is well known [72,75]. MMP14 also called MT1-MMP is activated by GPCRs [76] intracellular calcium [77] and PKC [78,79]. As shown in this study, most of the cells expressing neuregulins also expressed Mmp14 or Mmp15, which makes their involvement in GPCR-mediated transactivation of ErbB4 receptors likely. A hypothetical scheme describing the mechanism by which mGluR5 and other radial glial GPCRs would affect neuronal motility through NRG/ErbB4 signaling is shown in Fig. 9.

Simplified scheme to describe presented results. Glutamate released from neurons acting on radial glial mGluR5 [38,39] leads to activation of TRPC3 channels [39,40] causing a proteolytic activation of membrane-bound NRG [14] through MMP. Since NRG3 expression appears to dominate in neurons it is uncertain whether soluble NRG is released from radial glia or close by neurons. Soluble NRG acting on ErbB4 promotes stalling, multipolar transition, division of nearby neurons, and radial process extension [14], while active membrane-bound NRG additionally initiates cell–cell contacts. These effects are counteracted by blocking of mGluR5, TRPC3, MMP, or ErbB4. The mGluR5 block is circumvented by activation of other Gq-protein-coupled receptors such as ET or muscarinic (M) receptors. ET, endothelin. Color images are available online.
Interestingly, endothelin acting on Gq-coupled endothelin receptors was also able to counteract the effects of mGluR5 blockade at very low concentrations (0.1 nM). Endothelins have an important role in the regulation of glial function and pathology [80]. Endothelin is produced by endothelial cells and the endothelin A receptor is expressed exclusively in pericytes, whereas the endothelin B receptor is expressed in radial glia but not neurons [51]. Endothelin1 has previously been shown to activate Ca2+ signals in Bergmann glia, which are the cerebellar analogs of cortical radial glia [63].
Additionally, as shown in this study, endothelin stimulated the elongation of radial glial processes as has been shown earlier for muscarinic stimulation [14]. Endothelin has previously been shown to stimulate growth of spinal cord glial cells [81], suggesting that endothelin mediates communication between the vascular system and nervous system.
Thus, the present results indicate that endothelin produced by the vascular endothelium in vivo would exert a chemotactic influence on radial glia with NRG acting downstream of the endothelin receptor, which increase process growth and influence neuronal behavior in close apposition to radial glial processes (Fig. 8). Endothelin 3 has previously been shown to inhibit neuronal migration, which is in agreement with the results presented in this study [82].
Our results indicate that proteolytic activation of NRG acts downstream of Gq-coupled receptor activation. Most of the studies on ErbB receptor transactivation by GPCRs have been done on the EGFR ErbB1. There is, however, evidence that similar mechanisms are involved in activation of NRG/ErbB4. Activity-dependent release of NRG has been shown both in vivo and in vitro [31 –33], by mechanisms involving glutamate receptors [34], PKC, and Ca2+ ions [33]. Both mGluR5 and muscarinic receptor are Gq-coupled and induce Ca2+ mobilization.
The increased motility caused by blocking the mGluR5/TRPC3 or ErbB4 signaling is thus probably due to reduced NRG-dependent stalling, which would unleash endogenous motogenic mechanisms. Endogenous motogenic factors include BDNF and glutamate acting on AMPA receptors. AMPA receptor blockers considerably reduce neuronal motility [48,83]. In addition, cannabinoid receptor agonists have been shown to increase the motility of postnatal migrating neurons ([84], reviewed in Maccarrone et al. [85]). Interestingly, in this context blocking of the endocannabinoid synthesis counteracts the increased motility and reduced neuron–neuron interaction caused by ErbB4, mGluR5, or TRPC3 blockade [50]. Part of the effects seen in this study would thus be explained by interactions between the NRG/ErbB4 system and other diffusible factors like endocannabinoids. This would explain why radial glial mGluR5 (and muscarinic/endothelin receptors) appear to affect the motility of neurons at some distance from radial glial processes.
As discussed above, our results also suggest that mGluR5/TRPC3 and NRG/ErbB4 promote radial glial chemoattractive effects on surrounding neurons and induce stalling. Chemotaxis has previously been shown to limit the migration and positioning of transplanted neural progenitor cells [86]. In light of the results presented in this study, NRG seems to influence neurons in several ways, which would lead to restriction of their free motility and migration. It promotes neuronal attachment to radial glia and neuron–neuron contacts (probably membrane attached isoforms). Interference with mGluR5/TRPC3 or NRG/ErbB4 signaling might thus be a therapeutic means of aiding progenitor-based regenerative therapy.
Taken together, the results presented suggest that NRG/ErbB4 involved limits of neuronal motility and distribution by promoting neuron–radial glia and neuron–neuron attraction. As suggested, earlier cell–cell attraction limits the spread of grafted progenitors in brain tissue. It should be noted that most of the ligands used in this study, which increase neuronal motility, such as MPEP and afatinib, have been tolerated in clinical trials and could thus be used as adjuncts in neural progenitor cell transplantations to enhance migration of grafted cells into host tissue. This study therefore supports the notion that a signaling complex involving mGluR5/TRPC3 in close association with NRG/ErbB4 and pathway plays an integral role in neuron–radial glial interaction that together dictate the migratory behavior of cortical precursor cells. Neuregulins and their receptors have received a considerable attention due to the strong linkage of variants to schizophrenia [15,16]. The mGluR5 receptors have also been linked to schizophrenia and autism, and mGluR5 modulators are in clinical trials for the treatment of certain forms of schizophrenia [87].
Footnotes
Acknowledgment
The authors would like to thank Mr. Jarno Hörhä for his laboratory assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Sigrid Juséliuksen Säätiö, Magnus Ehrnroothin Säätiö, Finska Läkaresällskapet, the Swedish Brain Foundation (grants FO2017-0107 and FO2018-0209), and the Swedish Research Council (grants 2009-3364, 2013-3189, and 2017-00815).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Video S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.