
Abstract
Septic patients often die in a context of multiple organ dysfunction syndrome (MODS), despite the macro-hemodynamic parameters being normalized and after the onset of antibiotic therapy. Microcirculation injury during sepsis affects capillary permeability and leukocyte–endothelium interactions and is thought to be instrumental in organ injury. Several studies have demonstrated a beneficial effect of mesenchymal stromal cells (MSCs) injection on survival and organ dysfunctions in sepsis models. In vivo activity of MSCs also appears to be very much dependent on the information provided before injection. Indeed preconditioning by interferon γ (IFNγ; MSC-IFNγ) increases immunosuppressive capacity of MSCs in vitro and in vivo. Therefore, the objective was to evaluate the effect of MSC naive or IFNγ preconditioned on leukocyte–endothelium interactions in a polymicrobial sepsis model by intraperitoneal feces injection. Six hours (H6) after this induction, we used intravital microscopy in mice cremaster muscle venules to study the flow behavior of leukocytes. Plasmas were harvested to evaluate inflammation level and endothelial activation. We showed that MSC-IFNγ have a beneficial effect on microcirculation, by increasing the flow of white blood cells (WBCs) and the percentage of venules containing flowing WBCs, by significantly reducing the adhesion of WBCs and by increasing the average red blood cell velocity (VRBC). In conclusion, our results suggest that intravenous injection of preconditioned MSC-IFNγ improves microvascular hemodynamics in early phases of sepsis.
Introduction
Sepsis is a complex disease state characterized by a life-threatening organ dysfunction caused by a dysregulated host response to infection [1]. Its pathophysiology is still partly misunderstood. Patients often die in a context of multiple organ dysfunction syndrome (MODS), despite macro-hemodynamic parameters having been corrected and early antibiotic therapy being administered.
Several studies argue for microcirculation playing a key role in the pathophysiology of sepsis and show that persistence of microcirculatory disturbances are related to MODS and death [2 –5]. Microcirculation injury affects the three levels of microvascular network (arterioles, capillaries, and venules) impairing vascular autoregulatory mechanisms, capillary permeability, and leukocyte–endothelium interactions [6 –8]. Leukocytes activation and recruitment are essential for an effective host defense against infection. However, leukocyte–endothelial cell interactions have been implicated in inflammation-mediated vascular injury, which leads to end-organ injury.
Mesenchymal stromal cells (MSCs) are nonhematopoietic stromal cells originally isolated from bone marrow. MSCs can modulate proliferation, apoptosis, and tissue inflammation, partially through the secretion or stimulation of soluble factors [9]. Their properties revealed them as a promising therapeutic tool to treat sepsis [9,10]. Indeed, several studies have demonstrated a beneficial effect of the injection of MSCs following a sepsis: liver [10], kidney [10,11], pulmonary [11], and heart [12,13]. In addition, beyond the improvement of prevention of organ dysfunction, three of these studies have shown that MSCs injection had a beneficial effect on the animal survival [10 –12]. MSCs function also seems to be very much dependent on the information provided by their environment. In vivo, Polchert et al. [14] have shown the beneficial effect of MSCs' preconditioning by interferon-γ (IFNγ) in the treatment of the graft-versus-host disease in mice. This beneficial effect was positively correlated with the dose of IFNγ used to stimulate the MSCs. Furthermore, Ryan et al. [15] showed that in vitro the preconditioning by IFNγ increased the immunosuppressive profile of MSCs, including increased expression of anti-inflammatory factors such as TGFβ1 and HGF.
In this study, we made the hypothesis that preconditioned MSCs could protect the microcirculatory function in the early phase of sepsis. Our objective was to evaluate the effect of naive or IFNγ-preconditioned MSCs on leukocyte–endothelium interactions in a polymicrobial sepsis model induced by intraperitoneal feces injection (FIP). We used intravital microscopy in cremaster muscle of mice to study the flow behavior of leukocytes.
Materials and Methods
MSC preparation
MSCs from one donor were extracted from bone residues from patients undergoing a total hip replacement surgery after written informed consent (HIA, Hôpital Percy, Clamart, France). Human samples were collected and handled in the full respect of the Declaration of Helsinki. They were isolated by plastic adhesion, amplified up to 80–90% confluence in a medium containing MEMalpha (Biological Industries) supplemented with 10% fetal calf serum (FCS; HyClone) +0.01 mg/mL ciprofloxacine (Panpharma), then frozen in MEMalpha +10% human albumin (LFB Biomedicaments) +0.01 mg/mL ciprofloxacine +10% dimethylsulfoxide (Sigma), and finally preserved in liquid nitrogen. For experimental purposes, MSCs were thawed and seeded at 4,000 cells/cm2 before in vivo experiments. When the cells reached 80% of confluence the medium was changed: MEMalpha +10% FCS with or without 25 ng/mL IFNγ (Peprotech) for 48 h. On the day of in vivo transplantation, cells were washed and harvested and then suspended at a concentration of 1.25 × 106 cells in 250 μL Ringer lactate solution (Fresinus).
Immunophenotyping of MSC
MSC surface antigen expression was analyzed using a Navios flow cytometer (Beckman Coulter). MSCs were incubated in phosphate-buffered saline (PBS) +2% of human serum albumin +1 μg/106 cells of purified human immunoglobulin G (R&D systems) for 20 min at +4°C, then incubated with primary antibodies at a saturating concentration for 20 min at +4°C, and finally fixed in 1% formol/PBS. MSCs were labeled with CD29 [Ref. IM0791 clone K20, monoclonal antibody (mAb)], CD90 (Ref. IM1839U, clone F15-42-1-5, mAb), CD105 (Ref. A07414, clone 1G2, mAb), human leukocyte antigen locus DR (HLA-DR) (IM0464U, clone B8.12.2, mAb), CD106 (Ref. A66085, clone 1G11, mAb) (all from Beckman Coulter), CD54 (Ref. BBA20, clone BBIG-I1, mAb from R&D), CD73 (Ref. 550257, clone AD2, mAb from BD) antibodies. The percentage of positive binding cells (evaluated by comparison with isotypic control) and the mean intensity of fluorescence (MFI) were analyzed for each parameter using Kaluza software (Beckman Coulter). Dilutions applied to the antibodies were in accordance with the manufacturer's instructions.
Animal experiments
All experimental protocols, procedures, and endpoints criteria were approved by the institutional animal committee: animal experiments ethics committee of Paris VII University. The authorization number CEEALV/#2978 was issued from the French Ministry of Research according to European Directive 2010/63/EU (Directive of the European Parliament and the Council of the European Union relating to protection of animals used for scientific purposes).
All animals were male BALB/c mice (Janvier Labs, St Berthevin, France) weighting 20–25 g, and were acclimated for 3–5 days with standard mouse diet before use. Mice had free access to water and food until they were anesthetized for intravital microscopy experiment. Twenty-four mice were included in this protocol and distributed in four groups of six mice, two additional mice were used for MSC tracking analysis. Health and behavior of animals were monitored every hour.
Sepsis induction H0
Sepsis was induced by feces injection into the peritoneum (FIP). Feces were collected from the cecum of a donor mouse, mixed with sterile saline at 50 mg/mL concentration, and stored at −30°C. Bacteriological analysis of feces at Percy's Hospital (Clamart, France) detected Enterococcus faecalis at 108 CFU/mL, Staphylococcus warneri at 106 CFU/mL, and Bacillus cereus at 104 CFU/mL. The mixture was injected intraperitoneally (40 mg/mL in 1 mL) and time H0 of the experiment was determined. For the dose of 40 mg, ∼57% mortality was expected at 24 h according to previous assessment. Before FIP, analgesic buprenorphine (200 μg/kg in 0.8 mL of NaCl, Buprecare; Axience) was injected subcutaneously. Sham mice received subcutaneous injection of buprenorphine and intraperitoneal injection of sterile sodium chloride (NaCl; 0.9%) solution. The experimental procedure is given in Fig. 1.

Synopsis of the experimental procedure in vivo.
Treatment administration H1
Forty-five minutes after FIP or saline injection, the mice were anesthetized with 1.5–2% isoflurane (IsoVet; Piramal Healthcare, United Kingdom) for 20 min to inject the treatments. The right saphenous vein was cannulated (PE10 stretched by warming, ED = 0.25 μm) and treatments (106 cells in 200 μL PBS or PBS alone) were slowly injected 1 h after sepsis induction (H1). Catheter was then removed and saphenous vein ligated. The skin was disinfected and sutured with silk thread (5/0) (Fig. 1).
Treatment groups
The animals were divided into four groups (n = 6 animals/group). Group 1 served as Sham and received an intraperitoneal (H0) and intravenous (H1) injection of normal saline solution. The other three groups were injected with feces intraperitoneally (H0). At H1, group 2 (sepsis group) received an intravenous injection of normal saline solution PBS. The groups 3 and 4 (sepsis-MSC and sepsis-MSC-IFNγ respectively) received an intravenous injection of MSC or MSC-IFNγ.
Microcirculatory model (H4–H6)
Induction of anesthesia was performed with isoflurane (1.5–2%) followed by an intraperitoneal injection of ketamine (150 mg/kg, Imalgène 1000; Merial, France), xylazine hydrochloride (5 mg/kg; Sigma, France), and atropine (1 mg/kg; Renaudin, France). Anesthesia was maintained throughout the experiment with additional injections of the same drug preparation (a quarter of the initial dose). Animals were lying on a heating blanket (temperature 38°C) in supine position. A tracheotomy was performed and mice were immediately connected to a ventilator for small animals (Harvard Rodent Ventilator, model 683) with room air. Ventilation parameters were a tidal volume of 240 μL, a respiratory rate of 90 breaths/min, and a positive end expiratory pressure of 1 cm H2O. The right carotid artery was catheterized and connected to a pressure transducer (MP30; BIOPAC Systems) for continuous recording of systemic arterial blood pressure (MAP). A slow intra-arterial infusion (80 μL/h/10g) of physiological saline solution was given throughout the experiment to prevent clotting in the catheter.
The left cremaster muscle was prepared according to our previously described technique [16,17]. In brief, the scrotum was opened, the cremaster exteriorized, and spread over an optic glass. The muscle preparation was covered with transparent film and continuously superfused (2 mL/min) with Krebs solution (in mM, NaCl 118, KCl 5.9, MgSO4 0.5, NaH2CO3 28, CaCl2 1.25, glucose 10; Sigma, France) warmed at 34.5°C, and bubbled with gas mixture (O2 0%/CO2 5%/N2 95%) throughout the experiment.
Observation of venules was conducted on a microscope (Leitz, Germany) with a saline immersion objective ( × 50; Leitz). Experiments were recorded with HCImage software (Hamamatsu Photonics, Japan) connected to a digital camera Hamamatsu (C11440). Red blood cell velocity (VRBC) in microvessels was measured live using a photodiode system (IPM, San Diego, CA) connected to a cross-correlator (Model 102B; IPM). All the other microcirculation data analyses were performed offline using a free software open source (GPL2) Kinovea (Kinovea–0.8.19; France).
Intravital microscopy (H5 to H6)
After surgery and a stabilization period, venule videos recording started for a period of 1 h.
The studied venules were selected according to several criteria: Their diameter should be between 22 and 35 μm and direction straight and unbranched for at least 100 μm in length. The VRBC in these venules should be >500 μm/s to allow leukocyte visualization. Six to fifteen venules, distributed over the entire cremaster muscle preparation, were studied for each animal.
Venular diameter and VRBC are expressed in μm and μm/s, respectively. Rolling leukocyte velocity (VWBC) was determined from the time required for a leukocyte to traverse a given distance along the length of a venule (100 μm) and expressed as μm/s. Rolling leukocyte flux was calculated from the number of rolling leukocytes that crossed a perpendicular line through a given vessel within 1 min (n/min). The number of adherent leukocytes was determined by counting the number of firmly adherent leukocytes in 100 μm of venules length (n/100 μm). We considered leukocytes adhering >10 s as firmly adherent in each minute [17]. We also studied the heterogeneity of leukocyte distribution in venular network. We quantified this heterogeneity by measuring the percentage of venules with flowing leukocytes. This new parameter represents, among the venule studied, the venule with rolling leukocytes during the observation period and expressed as a percentage per mouse.
Blood analysis (H6)
At the end of the microcirculatory experimentation (H6), arterial blood samples were harvested for blood–gas [pH, pO2, pCO2, 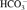 , base excess (BE), and hematocrit] and lactate analysis through carotid artery. The samples were analyzed in blood–gas analyzer (Rapidlab 348; Siemens Healthcare Diagnostic) and lactate meter (EDGE; Apex Biotechnology) with EDGE lactate strips.
, base excess (BE), and hematocrit] and lactate analysis through carotid artery. The samples were analyzed in blood–gas analyzer (Rapidlab 348; Siemens Healthcare Diagnostic) and lactate meter (EDGE; Apex Biotechnology) with EDGE lactate strips.
Plasma protein analysis (H6)
Mice blood was collected in 3.2% sodium citrate tubes and immediately centrifuged. Plasma was harvested and stored at −30°C until used. Circulating concentrations of adhesion molecules and anti- and proinflammatory cytokines [E-selectin, intercellular adhesion molecule-1 (ICAM-1), vascular endothelial growth factor (VEGF), vascular endothelial growth factor receptor 1 (VEGFR1), interleukin (IL)-1β, IL-12, IL-6, and IL-10] were performed using multiplex sandwich enzyme-linked immunosorbent assay (ELISA) arrays (Custom Quantibody Array, Raybiotech, Inc.) (Tebu-Bio) and ELISA analysis according to the manufacturer's instructions. Results per mice represent averaged technical quadruplicates.
Stromal cells localization after intravenous injection
To locate the cells after treatment, we injected two septic mice, MSCs labeled with carboxyfluorescein succinimidyl ester (CFSE), a fluorescent cell staining dye. In brief, MSCs were cultivated as described previously and harvested by trypsin (Gibco). MSCs were labeled with 5 μM CFSE (Life Technologies) during 5 min at room temperature and centrifugated before being suspended in Ringer lactate solution. Labeled MSC injection was performed according to the same procedure as previously described. The cremaster muscle was prepared and simultaneously observed using epifluorescence and transillumination for 5 h after MSC administration. Then several tissue samples (lung, kidney, spleen, and the liver) were collected and immediately observed.
Histological analysis
At H6 the tissue samples were harvested (left lung, left kidney, distal part of the spleen. and the left lateral lobe of the liver), fixed in formalin, and embedded in paraffin. Sections (5 μm) were prepared from paraffin blocks and stained with hematoxylin–eosin–safran (HES) by the HistIM platform of the Cochin Institute. The slides were scanned with a “Lamina” from PerkinElmer.
Statistical analyses
Statistical analysis was performed using R software (R version 3.0.0) with Rcmdr package. All data are reported as mean ± standard error of the mean (SEM) or median and interquartile range (IQR). All microvascular measurements were averaged by mouse before statistical testing. Thus for all tests n refers to the number of mice. We compared Sham and untreated sepsis groups by Wilcoxon test global. Comparisons between the two treated (MSC and MSC-IFNγ) and untreated sepsis groups were made with nonparametric permutation test with untransformed response values (Monte-Carlo approximation; Coin library), followed by Dunnett's post-test (nparcomp library) that adjusts 5% significance level for multiplicity. Considering a standard deviation of leukocytes flux estimated from previous experiments ∼2 n/min, we calculated that n = 6 animals per group will allow a 80% power to detect a difference of 4 n/min between groups at a 5% level adjusted for multiplicity.
Results
IFNγ preconditioning increased membrane expression of leukocyte cell adhesion molecules and the expression of immunomodulatory molecules on MSCs, without increasing the costimulatory molecules CD80 and CD86
Flow cytometric analysis revealed that after exposure to IFNγ, MSC maintained the same expression levels of CD29, CD44, CD73, CD90, and CD105, markers classically used for their definition (Fig. 2). However, IFNγ preconditioning led to an increase of HLA-DR expression (0.40% of positive cells in naive MSC population vs. 58% of positive cells in MSC-IFNγ population) (Fig. 2). We also observed an increase in CD54 (ICAM-1) (7% of positive cells in naive MSC population vs. 99% of positive cells in MSC-IFNγ population) (Fig. 2). No difference was observed for CD106 (VCAM-1) between groups (Fig. 2).

Phenotypic characterization of MSC and MSC-IFNγ. The values represented the percentage gated of positive or negative cells and the MFI for each marker on a logarithmic scale. “Mesenchymal” markers: CD29, CD44, CD54; CD73, CD90, CD105, and CD106. HLA-DR. These data are from a representative experiment. IFNγ, interferon-γ; MFI, mean fluorescence intensity; MSC, mesenchymal stromal cell.
Sepsis and treatment had no effect on animal's body weight or temperature
Mice body weight were similar before their randomization in the four experimental groups with 22.7 g ± 0.26. Four hours after feces administration, the treated and untreated mice have a swollen and bloated abdomen with a tendency to weight gain but this weight variation remains statistically nonsignificant compared with the sham group (Table 1). Moreover, body temperature, measured before placing the mice on the heating blanket (H4), showed no significant difference between sham, septic, or MSC-treated groups (Table 1).
Weight Variation (H0–H4) and Temperature Measured at H4 Non Parametric Values Were Expressed with Median and Interquartile Range
Six mice were used in each group. Sepsis was compared with sham group with a nonparametric exact Wilcoxon test. Effects of treatments were made by comparing the three sepsis groups (untreated, MSC, and MSC-IFNγ) with two/K sample permutation test followed by Dunnett's post-test. No difference was found in weight variation and body temperature among the four groups.
IFNγ, interferon-γ; MSC, mesenchymal stromal cell.
No histological sign of tissue damage was found at H6
We performed histological analyses of several organs that are commonly affected during sepsis: liver, lung, spleen, and kidney. Representative pictures of the organ we examined are given in Supplementary Figure S1. The examination of the HES staining showed no histological alteration of the tissues at H6. Specifically, we found no sign of swelling, necrosis, or mononuclear cell infiltration in the organs of the sepsis group. Therefore, we conclude that in our model, the microcirculation is affected before any objective histological sign of organ failure.
MSC-IFNγ had a beneficial effect on the metabolic acidosis induced by sepsis
As given in Table 2 sepsis induced acidosis with a significant decrease of bicarbonates ( ) and an increase in the absolute value of BE level compared with the sham group (P < 0.04 and P < 0.004, respectively). No significant changes were observed for pH, pO2, pCO2, hematocrit, and lactate plasma levels (Table 2).
) and an increase in the absolute value of BE level compared with the sham group (P < 0.04 and P < 0.004, respectively). No significant changes were observed for pH, pO2, pCO2, hematocrit, and lactate plasma levels (Table 2).
Blood Gas and Lactate Measured at H6 Nonparametric Values Were Expressed with Median and Interquartile Range
The number of mice used in the sham, sepsis, sepsis-MSC, and sepsis-MSC-IFNγ groups was 6, 6, 5, and 6 respectively.
Sepsis was compared with sham group: * P < 0.05; *** P < 0.005.
Effects of treatments were made by comparing the three sepsis groups (untreated, MSC, and MSC-IFNγ): §§ P < 0.01.
MSC-IFNγ treatment induced a significant decrease of the BE absolute value compared with sepsis group (P < 0.0001; Table 2). MSC and MSC-IFNγ treatments had no significant effect on pH, pO2, pCO2, hematocrit, and lactate values. However, it should be noted that MSC treatments, regardless of IFNγ preconditioning, resulted in a slight decrease of pO2 but statistically not significant (P = 0.051; Table 2).
MSC-IFNγ had a beneficial effect on the microcirculation disturbance induced by sepsis
Venules mean diameter was 27.45 ± 0.15 μm and was not significantly different between the four studied groups.
At the hemodynamic level, injection of feces resulted in a significant decrease in MAP measured at H5-H6 in sepsis group compared with sham (P < 0.01; 72.48 ± 2.95 vs. 56.73 ± 2.33 mmHg in sham and septic groups, respectively) (Fig. 3A). This blood pressure reduction was associated with an important decrease of VRBC (−59%) in venules of cremaster muscle (P < 0.004; 1791.77 ± 314.8 vs. 743.19 ± 93.94 μm/s in sham and septic groups, respectively; Fig. 3B). In our sepsis model, leukocyte trafficking was modified in the cremaster muscle venule. Sepsis significantly decreased rolling leukocyte flux and the percentage of venules with flowing leukocytes (P < 0.01 and P < 0.006, respectively; Fig. 3B). VWBC was also significantly reduced in sepsis group (P < 0.04; Fig. 3). This decrease paralleled a nonsignificant trend toward an increase in leukocyte–endothelial adherence in cremaster muscle venules (Fig. 3A, B).
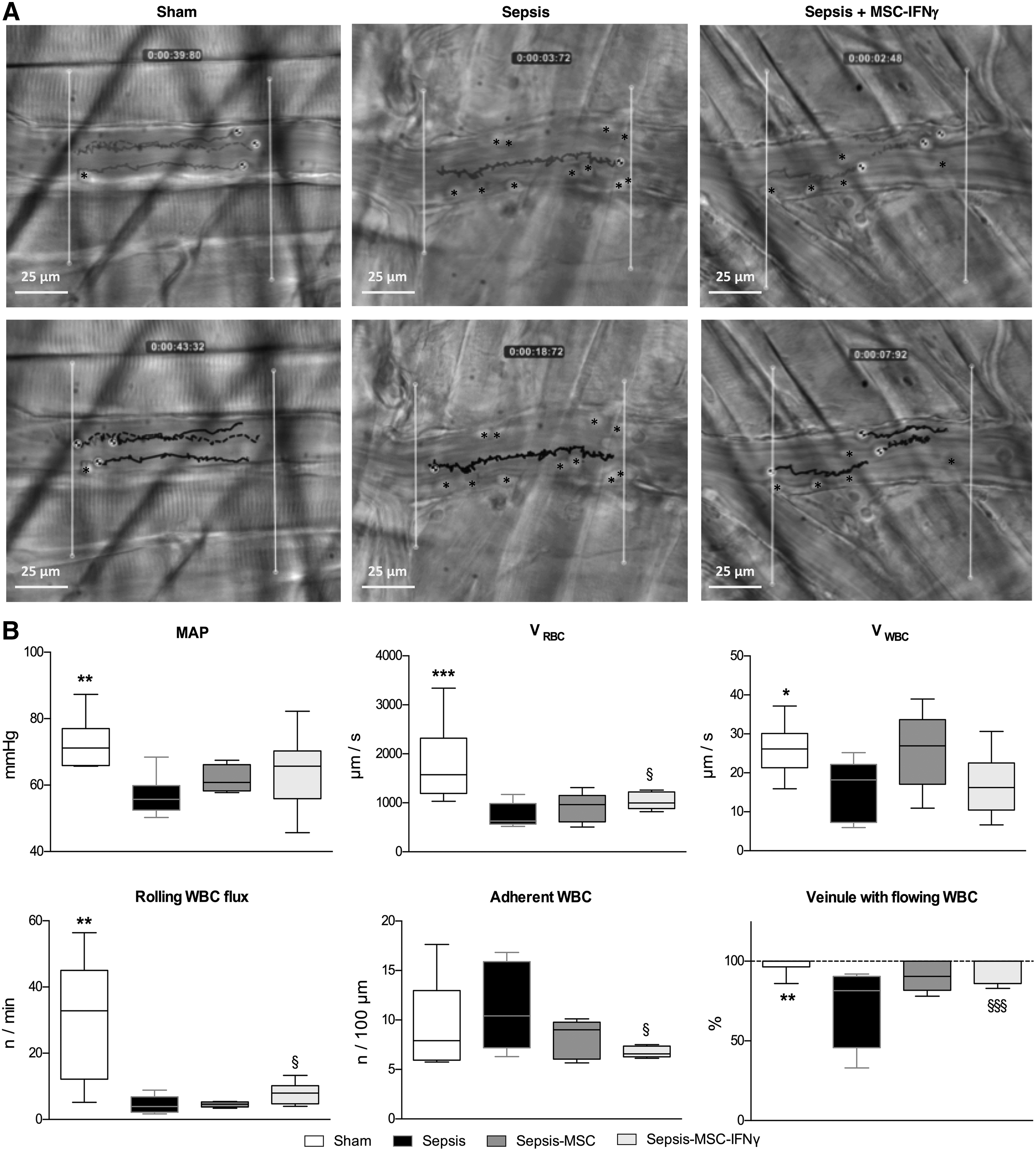
Systemic MAP, red blood cell velocity (VRBC) and leukocyte endothelium interaction in cremaster muscle venules (H5-H6).
As given in Figure 3B, MAPs were not significantly increased in the MSC-treated mice (MSC and MSC-IFNγ) compared with untreated sepsis group.
In animals treated with naive MSC (sepsis-MSC group), no significant difference was observed on the macro and microcirculatory parameters compared with the sepsis group.
In contrast, MSC-IFNγ treatment improved several systemic and microcirculatory parameters altered by sepsis. The injection of MSC preconditioned with IFNγ significantly increased the VRBC in venules (P < 0.02; Fig. 3A, B). As given in Figure 3 this treatment reversed the sepsis effect on the percentage of venules with flowing leukocytes (95% ± 3% vs. 74% ± 8% in sepsis-MSC-IFNγ and sepsis groups, respectively, P < 0.0007; Fig. 3A, B). This improvement was associated with a significant increase in the rolling leukocyte flux (7.92 ± 1.23 vs. 4.47 ± 0.99 n/min in sepsis-MSC-IFNγ and sepsis groups, respectively, P < 0.003; Fig. 3A, B). Moreover MSC-IFNγ injection did not significantly affect the VWBC but reduced the leukocyte–endothelial adherence observed in sepsis group (P < 0.005; Fig. 3A, B).
MSC treatment modulated the concentration of several biomarkers of endothelial dysfunction
To test whether the beneficial effects of the treatments were correlated to the modulation of inflammatory cytokines or biomarkers of endothelial dysfunction, the concentration of various soluble plasma factors were assayed.
Six hours after sepsis induction, the levels of adhesion molecule (E-selectin) and the cytokines IL-10 and IL-12 were increased significantly in septic mice compared with sham group (P < 0.01; Fig. 4). In contrast, soluble ICAM-1, VEGF and VEGFR1 rates were not significantly different between sham and sepsis groups (Fig. 4). IL-1β level remained below the detection threshold in sham and septic groups (<40 pg/mL; data not shown). Moreover, tumor necrosis factor α expression was not higher in our septic group than in our sham group (data not shown). In contrast, IL-6 expression level, undetectable in the sham group was out of range above the threshold in the sepsis group (data not shown). Concerning the level of IFNγ, the values obtained from some samples were below the lower threshold of the range. The samples we were able to assay do not show any significant difference between groups (data not shown).

Plasma protein analysis (H6). Circulating concentrations of adhesion molecules and anti and proinflammatory cytokines (E-selectin, ICAM-1, VEGF, VEGFR1, IL-12, and IL-10) were expressed with median and min/max range. The number of mice used in the sham, sepsis, sepsis-MSC, and sepsis-MSC-IFNγ groups was 5, 5, 6, and 6 respectively. Sepsis was compared with sham group: **P < 0.01. Effects of treatments were made by comparing the three sepsis groups (untreated, MSC, and MSC-IFNγ): § P < 0.05 and §§ P < 0.01. IL, interleukin.
As given in Figure 4, the injection of MSC and MSC-IFNγ significantly reduced the rate of soluble E-selectin compared with the septic group. In addition, preconditioning by IFNγ also resulted in a significant increase in plasma level of ICAM-1. None of the other markers studied showed significant expression change either by sepsis or by MSC treatment (data not shown) and the IL-1β levels remained below the detection limit of the assay for all treatments studied (data not shown).
Labeled MSCs were found mainly in the lung 5 h after injection
Five hours after injection of fluorescent MSCs through the saphenous vein, the cells were trapped mainly in the lung (Fig. 5). Only a few fluorescent cells were found in the other tissues (cremaster, liver, kidney, and spleen). In cremaster muscle, the few labeled cells were located in the capillaries (Fig. 5).

Observation of fluorescent MSC in the lung and cremaster muscle. MSCs were labeled with CFSE and analyzed at × 25 magnification with an immersion objective. Observation was realized 6 h postsepsis induction.
Discussion
Our study using a mice polymicrobial sepsis model by intraperitoneal feces injection showed that MSC-IFNγ seem to have a beneficial effect on microvascular hemodynamics: by increasing the flow of white blood cells (WBCs) and the percentage of venules containing flowing WBCs, by significantly reducing the adhesion of WBCs, and increasing the average VRBC.
In some of these studies naive MSCs have been shown to improve survival of septic animals [10,18,19] and reduce organ damage [10,18]. The objective of our study was to evaluate if the administration of MSCs could limit the microcirculatory disorders, which participate in the occurrence of the organ failure. We deliberately chose to measure the microcirculatory parameters at H6 to avoid any bias related to the mortality of the animals. However, the 6h post-sepsis timepoint is too early to observe organ failures with this model. Moreover, the model of sepsis can also potentially influence the MSC effect. In our experiments, we found no effect of the naive MSCs on the microcirculatory parameters we measured. Most articles using MSCs in sepsis models use the technique of cecal ligation and puncture (CLP) or the administration of lipopolysaccharide (LPS) or endotoxins. Note however that bolus injection of LPS, since as discussed by Laroye et al., does not reproduce the hemodynamic changes observed in human sepsis. As regards CLP model, the gauge of the needle or the number of perforations could induce differences of sepsis severity and the host response to CLP is to mark off the infected/inflamed area by creating an abscess. In addition, several animals contain the infection and do not progress to septic shock and recover fully [20,21]. For all these reasons, we chose the FIP approach: polymicrobial, severe, and reproducible.
Moreover, it is likely that difference in sepsis model can affect the effect of naive MSC since their biological activity is highly environment dependent. Indeed, previous published data from our laboratory showed that MSCs exhibit a differential effect according to the status of T cells: an inhibitory activity in contact with healthy highly proliferative T cells and a stimulating activity in the presence of low proliferative T cells (injured by LPS) [22]. It is therefore reasonable to think that signals received by our cells at an early time, before the appearance of organ injuries, gives rise to different mechanisms of action. However, this does not prejudge the potential effect of administering these same cells at later times. Finally, several data of literature demonstrate that MSC engrafted and functioned across the species barrier in lot of different experimental models, notably in pathophysiological contexts with a strong inflammatory component [23]. We believe that it is unlikely that the lack of effect observed with the naive MSCs was the consequence of a rejection.
During sepsis, endothelium is thought to be pivotal in several pathophysiological waypoints leading to multiples organ failure:
- Both through its ability to actively participate in the release of inflammatory mediators, leukocyte activation and recruitment, and hemostasis induction.
- And through the decrease in perfusion arising from microvascular injury. [2,6].
Beneficial effects of MSCs on skin or intestine microvascular perfusion have been reported in ischemia by decreasing interactions between leukocytes and the vascular endothelium [24,25]. However, to our knowledge no publication exists on the in vivo effect of MSCs on interactions between leukocytes and vascular endothelium in context of sepsis.
In our model of sepsis, we observed an important decrease in rolling leukocyte flux (−85%) and a slight reduction of their mean velocity (VWBC). These results were in agreement with the experimental literature data in mice models of sepsis by CLP [26 –28]. Naive MSCs did not induce beneficial effects on microcirculatory parameters, but when mice were treated with MSC-IFNγ 1 h after sepsis induction, the rolling leukocytes flux increased significantly in venules. In addition, the percentage of venules with flowing leukocytes, that largely decreased during sepsis, which reflects profound disturbances in leukocytes trafficking, was almost to sham situation with MSC-IFNγ.
Although it is commonly accepted that leukocyte adhesion increases in endotoxemia [28,29], results are more controversial in sepsis associated to peritonitis where the leukocytes adhesion has been shown to increase [6], decrease [30], or remain unchanged [26,27]. In this context, it is interesting to note that we found a significant decrease in adhesion of leukocytes in septic mice treated with MSC-IFNγ.
It is well known that systemic and local hemodynamic play an important role in leukocyte trafficking in the microcirculation. However, it should be noted that the effects on leukocytes adhesion or red blood cell dynamics in the MSC-IFNγ-treated mice were found in the absence of significant effect of this treatment on the sepsis-induced MAP reduction. This observation is in line with recent works that demonstrated improvement of microhemodynamic after systemic administration of MSC in ischemia–reperfusion model [24,25].
These effects were associated with a significant decrease in plasma soluble E-selectin. During sepsis or other severe injuries, the systemic inflammation contributes to the release of adhesion molecules (eg, E-selectin). These molecules may serve as biomarkers for the detection of general endothelial activation and sepsis, and therefore could be useful to monitor organ's endothelium damage [31]. In our experiments, we found an increase in the soluble E-selectin during sepsis, which is in line with the decrease in leukocyte flux and leukocyte velocity. Soluble E-selectin was significantly reduced by MSC-IFNγ.
The significant increase in soluble ICAM-1 associated to MSC or MSC-IFNγ might also participate in the decreased in leukocyte adherence observed in these groups. Indeed, Kim et al. [32] have shown that MSCs can produce soluble ICAM-1. Moreover, we and others already showed that proinflammatory molecules lead to an overexpression of ICAM-1 by MSCs. This overexpression could potentiate the accumulation of immune cells at the proximity of MSCs, leading to the enhancement of their immunosuppressive effects [33,34]. A study by Ren et al. [35] also showed that MSC-mediated immunosuppression was significantly reversed in vitro and in vivo when the adhesion molecules were genetically deleted. Finally, Boldt et al. [36] explained that soluble ICAM-1 might compete with membrane-bound ICAM-1 and thus prevent leukocyte attachment. Therefore, it has been suggested that elevated plasma levels of adhesion molecules like ICAM-1 may have protective effects (28).
An interesting next step would be to test whether IFNγ increases MSCs supernatant content in sICAM-1 and resumes MSC activity.
Furthermore, it has been previously shown that IL-6 could contribute to the reduction of leukocyte adhesion on endothelial cells cultured in direct contact with BM-MSC [37]. The inhibition of IL-6 signaling using antibodies targeting soluble IL-6 or membrane IL-6R significantly reduced the inhibitory effects of BM-MSC [37]. Andreeva et al. also showed that IFNγ priming of MSCs significantly enhanced their production of IL-6 under “physiological” hypoxia [38], IL-6 could therefore be a mediator that partly explains the ability of MSC-IFNγ to improve microvascular hemodynamics.
A clinical trial also showed that treatment with 100 μg of IFNγ administered subcutaneously once daily for 21 days decreased the number of infection-related deaths in severely injured patients. IFNγ therapy seems to be beneficial in sepsis patients undergoing immunosuppression [39]. Concerning the plasma concentrations of mouse IFNγ we found no significant difference between our groups (data not shown). Moreover, we analyzed supernatant levels of IFNγ secreted after 6 h by naive or primed MSCs and it seems that IFNγ priming leads to a modest increase of IFNγ evaluated in MSC-IFNγ condition (data not shown). Whether this IFNγ originates from the secretion of the cell or the recombinant IFNγ treatment potentially stuck to our cells is unclear. However, it should be noted that the concentrations measured are probably still far removed from those used in clinical trials.
Finally, our experiments of labeled-MSC tracking revealed that very few MSC injected intravascularly were present in peripheral organs in contrast to the lung. This can also explain the observed trend toward decreased arterial pO2 in the treated mice compared with untreated mice (−28% or −31% decrease in MSC-naive or MSC-IFNγ treatment). These observations were in line with Schrepfer et al. [40] who also observed episodes of apnea, tachypnea, and hemodynamic alterations characteristic of pulmonary embolism after MSCs intravenous injection in mice. Despite the pulmonary trapping, several studies have demonstrated improvement of the function of certain organs (kidney, liver, and heart) early after injection of MSCs [10,11,13]. As proposed by other authors [10,13], our results are more likely because of paracrine effects.
Considering these findings, it would be interesting to test MSCs issued from spheroid culture aiming at modulating mean cell size and therefore lowering their possible embolism. Moreover, this culture condition is known to enhance anti-inflammatory, angiogenic, and tissue reparative/regenerative effects with improved cell survival after transplantation [41].
Conclusion
Our main results showed that intravenous injection of IFNγ-preconditioned MSC improves microvascular hemodynamics in early stages of sepsis by significantly increasing the VRBC, the rolling WBC flux and the number of venules with circulating WBCs. We also observed a significant reduction in the rate of soluble E-selectin. Other studies are now needed to confirm a potential beneficial effect of MSC treatment at later stages of sepsis, possibly requiring other types of preconditioning. Moreover, because the cell therapy strategy may be difficult to implement in such early times, it would be interesting to explore if secreted factors such as extracellular vesicles derived from IFNγ-preconditioned MSC and storable could recapitulate all or part of their therapeutic effect.
Footnotes
Acknowledgments
The authors thank Dr. Georges Uzan for allowing IRBA's team to initiate a collaboration with Prof. Vicaut's team. The authors specially thank Dr. Martinaud and the biology department of Percy's Hospital, Clamart, for their contribution in bacteriological evaluation of our pool of feces and Pr Saint Blancard and the department of anatomical pathology for their help. The authors also thank Véronique Sarilar for the help with statistical analysis of plasma assays and Cédric Thepenier for his scientific support. The authors extend their thanks to their colleagues for continuous support throughout this study. The authors also thank HistIM plateform, especially Marilyn Favier and Rachel Onifarasoaniaina from Institut Cochin. I also want to thank Marion Grosbot for her contribution in the preparation of MSCs. Finally I want to thank Elisabeth Laemmel for her help in developing the sepsis model.
Author Disclosure Statement
The authors do not declare any conflicting financial interests.
Funding Information
This study was supported by grants from DGA (Direction Générale de l'Armement, Ministère des armées) n° PDH-1-SMO-1-0207 and from University Paris VII.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.