
Abstract
Age-related cellular changes and limited replicative capacity of adult mesenchymal stem cells (MSCs) are few of the challenges confronting stem cell research. MSCs from human fetal membranes (hFM-MSCs), including placental, umbilical cord, and amniotic membrane, are considered an alternative to adult MSCs. However, the effect of mothers' age on hFM-MSC cellular properties is still not clearly established. This study aimed to evaluate the effect of mothers' age on hFM-MSC telomere length, telomerase activity, and proliferation ability in three different age groups: GI (20–29 years), GII (30–39 years), and GIII (≥40 years). hFM samples were collected from pregnant women ≤37 weeks after obtaining consent. hFM-MSCs were isolated and cultured to characterize them by flow cytometry and assess proliferation by MTT assay and doubling time. Telomere length and expression levels of human telomerase reverse transcriptase were assessed by quantitative real-time polymerase chain reaction (qRT-RCR). hFM-MSCs in the three age groups were spindle-shaped, plastic-adherent, and exhibited high proliferation rates and strong expression of hMSC markers. GI showed the longest telomere length in hMSCs in various FM regions, whereas GIII showed the highest level of telomerase expression. There was no difference in telomere length between GII and GIII, and both groups showed the same hMSC characteristics. In conclusion, although the hFM-MSCs derived from different fetal membranes maintained the MSC characteristics in all study groups, the hFM-MSCs of older mothers had shorter telomeres and higher telomerase activity and proliferation rate than did those derived from younger mothers. Thus, the hFM-MSCs of older mothers could be unsuitable for expansion in vitro or stem cell therapy. Determination of telomere length and telomerase expression level of hFM might help characterizing and understanding the biological differences of hFM-MSCs in different age groups.
Introduction
Mesenchymal stem cells (MSCs) are multipotent stem cells that have the ability of self-renewal and potential to differentiate into multiple stem cell lineages [1,2]. MSCs exist in various tissues, mainly in the bone marrow and other types of adult tissues, and in fetal tissues including placenta, amniotic membrane, Wharton's jelly, and umbilical cord blood [3 –8]. The key advantages of MSCs of human fetal membranes (hFM-MSCs) over those from adult tissues are high proliferation, long lifespan, wide differentiation potential, availability, and easy procurement with less ethical issues [1,2,9 –12]. In addition, hFM-MSCs can migrate, immunomodulate, and differentiate better than adult tissue-derived MSCs [13,14]. Given their versatility, stem cells are widely used for clinical applications, and fetal and hFM-MSCs are considered an attractive choice for cell-based therapies and tissue engineering [9,11,15 –17].
The lifespan of cells is directly proportional to the telomere length. Telomeres are special nucleoproteins that exist at the end of each chromosome and protect the chromosomal ends from DNA fusion, DNA recombination, and terminal DNA degradation [18,19]. Telomeres are maintained through the activation of telomerase enzyme, which comprises telomerase reverse transcriptase (TERT) and telomerase RNA component [20]. Telomere shortening occurs as a result of an incomplete replication of linear chromosomes owing to telomerase activity repression during somatic cell division, which serves as a mitotic clock and ultimately leads to aging and loss of cell viability [21].
In contrast to somatic cells, germ cells, embryonic stem cells, and tumor cells express high levels of telomerase and are, therefore, able to proliferate endlessly. Adult MSCs isolated from various tissues show a low-to-absent level of telomerase activity. This variation is because of the different cellular turnover rates in organs [22].
Previous studies reported the effect of MSC sources and donor age on the phenotype of hMSCs in culture. These studies revealed degradation of MSC characteristics from adult tissues with age, including loss of differentiation, self-renewal, and organ repair ability, shortening of telomeres, change in gene expression, and enhanced aging features [21,23 –26]. Altogether, these findings highlight the limitations of the use of MSCs in cell therapy and regenerative medicine. Because there is a close relationship between the effect of sources and increase of donor age on MSC capacities, selecting an appropriate source and studying the effect of donor age on the biological characteristics of human MSCs (hMSCs) are essential before clinical applications.
To this end, we hypothesized that mother's age might have an effect on telomere length, telomerase expression, and proliferation rates of hMSCs isolated from different fetal membranes. Therefore, this study aimed to isolate MSCs from hFMs including placenta, Wharton's jelly of umbilical cord, and amniotic membrane of mothers in three different age groups to evaluate the impact of mother's age on telomere length, telomerase activity, and cellular proliferative potential of MSCs.
Materials and Methods
Ethical considerations
This study was reviewed and approved by the Biomedical Research Ethics Committee at the Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia (No: 21–71, KAU). Tissue samples were collected after obtaining informed consent from all the pregnant women enrolled in this study.
Sample collection
hFMs were collected from 48 pregnant women, following normal full-term (≤37 weeks) delivery at King Abdulaziz University Hospital (KAUH). History of gestational diabetes, preeclampsia, hepatitis B, hepatitis C, human immunodeficiency virus, or other microbial infections were considered potential confounders and were excluded. No effect modifiers were spotted. The samples were collected between March 2017 and April 2018 and were divided into categories according to the mother's age as follows: GI (20–29 years), GII (30–39 years), and GIII (≥40 years).
Tissue samples were transferred immediately after delivery to the stem cell laboratory in transport medium (Hank's balanced salt solution containing 2% penicillin–streptomycin). The hFM samples were carefully examined to identify different anatomical regions: decidua basalis (hDB), chorionic plate (hCP), amniotic membrane (hAM), and umbilical cord Wharton's jelly (hWJSCs). In this study, the maternal age was the predictor, telomere length and hTERT gene expression in the MSCs were the outcomes.
Control samples and cell lines
In addition to the hFM-MSCs isolated from young mothers, two cell lines were used as controls: human leukemic cancer cell line (K562; passage 6), which was procured from American Type Culture Collection (ATCC® CCL-243™) (Prince William, Manassas, VA) and human bone marrow mesenchymal stem cells (hBM-MSC; passage 6) from an adult human donor (60 years old). The two cell lines were kindly provided by Professor Saleh Al-Karim, the head of the Embryonic Stem Cells Unit, King Fahd Center for Medical Research, Jeddah, Saudi Arabia.
Isolation and cell culture
Isolation of hFM-MSCs were performed under sterile conditions in a level II Biological Safety Cabinet (BSC-Level II) as previously described by Gauthaman et al. [16]. In brief, the fetal membrane pieces were exposed to 5 mL of enzymatic cocktail solution and filtered using 1,000 μL filter pipette tips (Greiner, Hamburg, Germany). Then, cell culture dishes, PS, 35/10 (Greiner) were incubated at 37°C for 45 min in a 5% CO2 incubator (Life Technologies, Carlsbad, CA). After the incubation period, 5 mL of neutralizing medium was added to the hFM pieces in the Petri dish to neutralize the enzyme effects.
The medium containing the hMSCs was collected and resuspended in Dulbecco's modified Eagle's medium–low glucose (DMEM-LG) (Gibco, Waltham, MA), 5% fetal bovine serum (FBS) (Sigma, St. Louis, MO), 1 mL (
On day 6 of the primary culture, many colonies contained cells, acquiring long spindle-shaped fibroblast-like cells with variation in their size. On day 9 of the primary culture, the density of the cells was medium and they covered ∼50% of the flask surface. On day 12 of the primary culture, the density of the cells was above medium and they covered ∼85%–90% of flask surface. Then, the cultured hFM-MSCs were expanded in vitro until the 6th passage. The hBM-MSCs (passage 6), isolated as described by Bernardo et al. [5], were thawed in a 37°C water bath and cultured in DMEM-LG. The K562 passage 6 was suspended in fresh warm RPMI media according to the manufacturer's protocol (ATCC CCL).
Analysis of MSCs surface markers
The cultured hFM-MSCs were analyzed using fluorescent-activated cell sorting (FACS) for detection of the related CD surface markers [5,16,27]. After trypsinization and washing twice in phosphate-buffered saline (PBS), 1 × 106 cells were incubated with MSC-related CD markers: CD45-APC, CD105-FITC, CD73-PERCP, CD29- PERCP, CD34-PE, CD44-PE-CY, and CD90-FITC. The cells were analyzed using an FACS Aria III Instrument (BD BioSciences, San Jose, CA) and data processing software (BD FACSDiva™ Flow Cytometry Software; BD BioSciences).
Assessment of proliferation capacity
Cell morphology was evaluated using phase contrast optics (Nikon Instruments, Tokyo, Japan), whereas the viability rates were evaluated using the MTT reagent kit [3-(4, 5-dimethylthiazolyl2)-2, 5-diphenyltetrazolium bromide] (Sigma) according to the manufacturer's instructions. At ∼80%–90% confluence, cells were detached by treating them with 0.25% trypsin–ethylenediaminetetraacetic acid (EDTA) (Gibco). Based on the viability rates, 2 × 105 cells per well were seeded in 24-well culture plates (Greiner) containing DMEM-LG and then incubated at 37°C in a 5% CO2 atmosphere for 24, 48, and 72 h. Absorbance was measured using a spectrophotometer (SpectraMax i3 Multi-Mode microplate reader; Molecular Devices, Sunnyvale, CA) at 750 and 630 nm. Doubling time (DT) was calculated as earlier outlined by Choi et al., to assess hFM-MSC growth rates from different fetal membranes [17].
Cells were separated by treating them with 0.25% trypsin-EDTA (Gibco) at ∼80%–90% confluence. Based on the trypan blue assay, 2 × 104 cells were plated in T-75 culture flasks (Greiner). When cells reached 80%–90% confluence, final numbers of cells were counted. The doubling time calculation for hFM-MSCs was performed using an online doubling time calculator (
RNA isolation and quantitative real-time polymerase chain reaction of telomerase activity
Total RNA isolation was performed by column purification technology using an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. RNA purity and quantity were measured as the absorbance ratio A260/A280 nm by a NanodropTM spectrophotometer (Nanodrop Technologies, Wilmington, DE). An aliquot of 1 μg of RNA was used to synthesize cDNA using a cDNA reverse transcription kit (Promega, Hilden, Germany). The relative hTERT expression was estimated using 1 mL of cDNA, with QuantiFast SYBR Green PCR Kit (Qiagen) on an ABI StepOne Plus Real-Time PCR System (Applied Biosystems, Foster City, CA). The data were normalized to the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
The primers of hTERT and GAPDH were designed and checked using the in-perform Primer-BLAST at
The 2−ΔΔCT method was used for relative quantification. The change in gene expression of hTERT was presented as a fold increase or decrease. The latter was classified into three classes: fold changes <0.5 were considered as downregulation, fold changes of >2 were considered as upregulation, and fold changes ranging from 0.5 to <2 were presented as unchanged gene expression [28].
Genomic DNA extraction and evaluation of telomere length by quantitative real-time polymerase chain reaction (qRT-RCR)
Genomic DNA was isolated using a Qiagen DNeasy kit (Qiagen) according to the manufacturer's instructions. The DNA quantity was measured as the absorbance ratio A260/A280 nm by a Nanodrop spectrophotometer (Nanodrop Technologies). Agarose gel electrophoresis was used to determine DNA sample integrity and to resolve PCR products. qRT-PCRs were applied using Absolute Human Telomere Length Quantification PCR Assay Kit (ScienCells, Carlsbad, CA), with Fast Start Essential DNA Green Master mixes (Roche, Basel, Switzerland), on a StepOnePlus real-time PCR system (Thermo Fisher Scientific) according to the manufacturer's protocol.
The 2−ΔΔCq method was used to calculate the average telomere length. The change in telomere length was presented as an increase or decrease as follows: values that showed an increase of <5 kb were represented as short, and values that showed an increase of >5 kb were considered as long. Total telomere length was derived from reference sample telomere length (233 ± 10 kb) × 2−ΔΔCq according to Tsamou et al. [29].
Statistical analysis
The statistical analysis was performed using the Statistical Package for Social Science (SPSS version 20; Chicago, IL). In PCR analysis, the differences between the measured parameters in different studied groups were compared using analysis of variance (ANOVA) [least significant difference (LSD)] test. In MTT assay, the difference in proliferation level parameters in different experimental groups and different duration was made using two-way ANOVA (LSD) test. P < 0.05 was considered significant. While in doubling time, the differences in expansion rate between measured parameters in different studied groups were compared using multivariate analysis, then, the difference between measured parameters in different experimental groups and different duration was made using one-way ANOVA (LSD) test. P < 0.05 was considered statistically significant.
Results
Morphology of hFM-MSCs after isolation and cell culture
At passage 6 of the cell culture, the growth of cells from each of the studied regions of the hFM was normal, fast, and productive. Under a phase-contrast microscope, the hMSCs cells seemed adherent and they had the characteristics of fibroblast-like cells as they became enlarged and flattened with expanded cytoplasm. In addition, the density of the cells was medium and was covered ∼50% of the flask surface. After 3 days, the cells covered 95%–100% of the flask surface (Fig. 1).

Cellular morphology of hFM-MSCs. Representative images of hFM-MSCs [decidua basalis (hDB), chorionic plate (hCP), amniotic membrane (hAM), and umbilical cord Wharton's jelly (hWJSCs)] from the three different age groups during cell culture at the initial passage (P0) and late passage (P6). At day 3 and 6, hFM-MSCs appear short, then from day 9 until 6th passages the cells appear fibroblast-like cells. They were longer, flattened, and have an expanded cytoplasm. (magnification × 10). hFM-MSCs, human fetal membranes–mesenchymal stem cells. Color images are available online.
K562 cells from passage 6 grew in suspension culture in RPMI-1640 culture medium. Cells showed rapid division, leading to splitting and subculturing of the cells. Under a phase contrast microscope, K562 cells appeared spherical and had the characteristics of undifferentiated blast cells. The K562 cells in suspension cultures appeared either as single or small clusters (5–7 cells) (Fig. 2). The adult hBM-MSCs from passages 6 grew in DMEM-LG. The cells appeared adherent and had the characteristics of fibroblast-like cells with expanded cytoplasm. The adult hBM-MSCs were different from hFM-MSCs in the same passage, with regard to the number and proliferation of the cells as fewer and more differentiated adult hBM-MSCs were observed (Fig. 2).
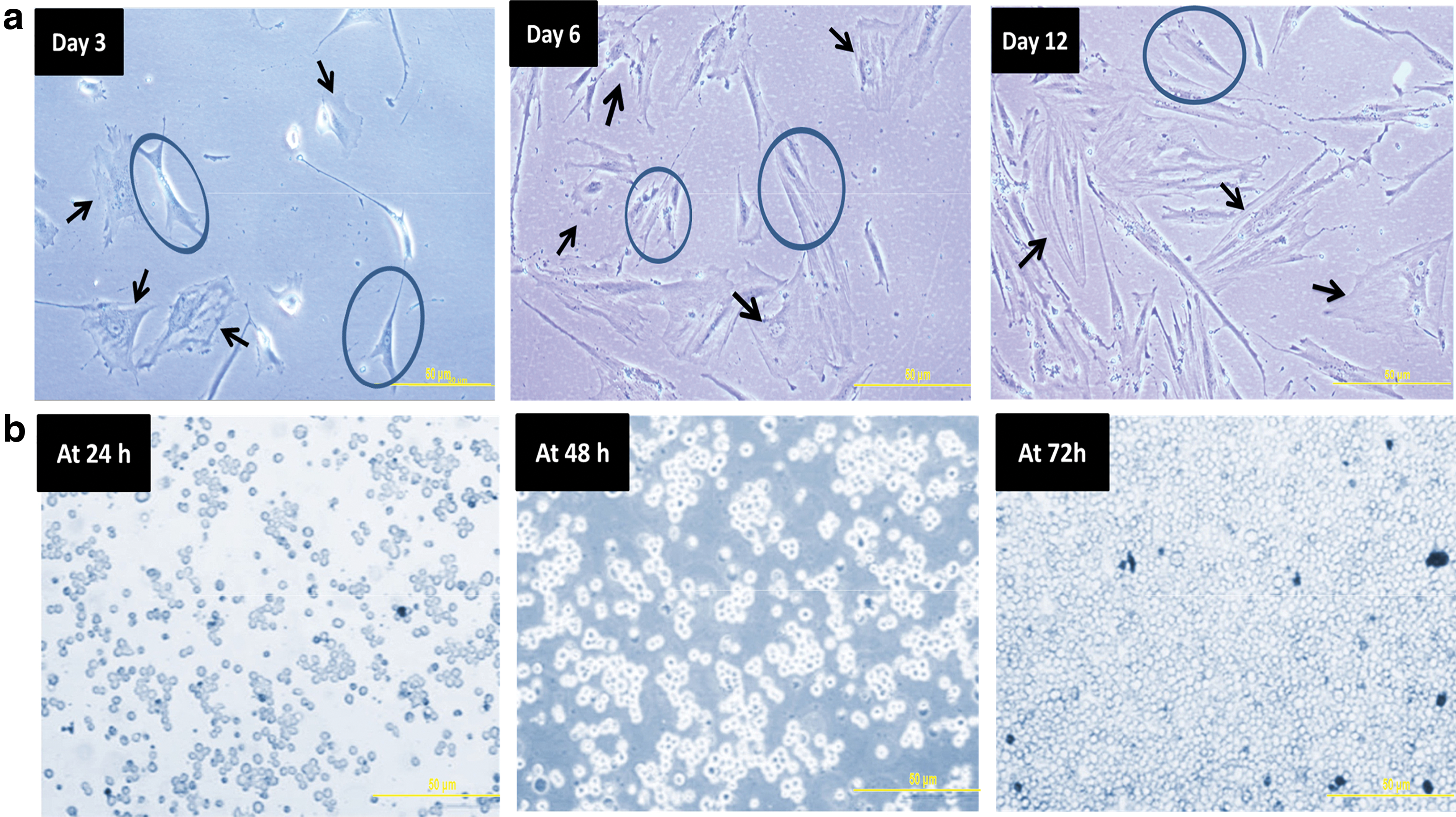
Cellular morphology of hBM-MSC and K562 at passage 6. Representative images of
Analysis of MSC surface markers
The hFM-MSCs were recognized with MSC-positive CD surface markers using flow cytometry analysis. The cultured hFM-MSCs of early passages (P3) showed strong expression (80%–100%) of hMSC CD markers CD29, CD73, CD90, CD44, and CD105. In addition, these cultured hFM-MSCs were negative for the hematopoietic markers CD34 and CD45 (Table 1 and Fig. 3).
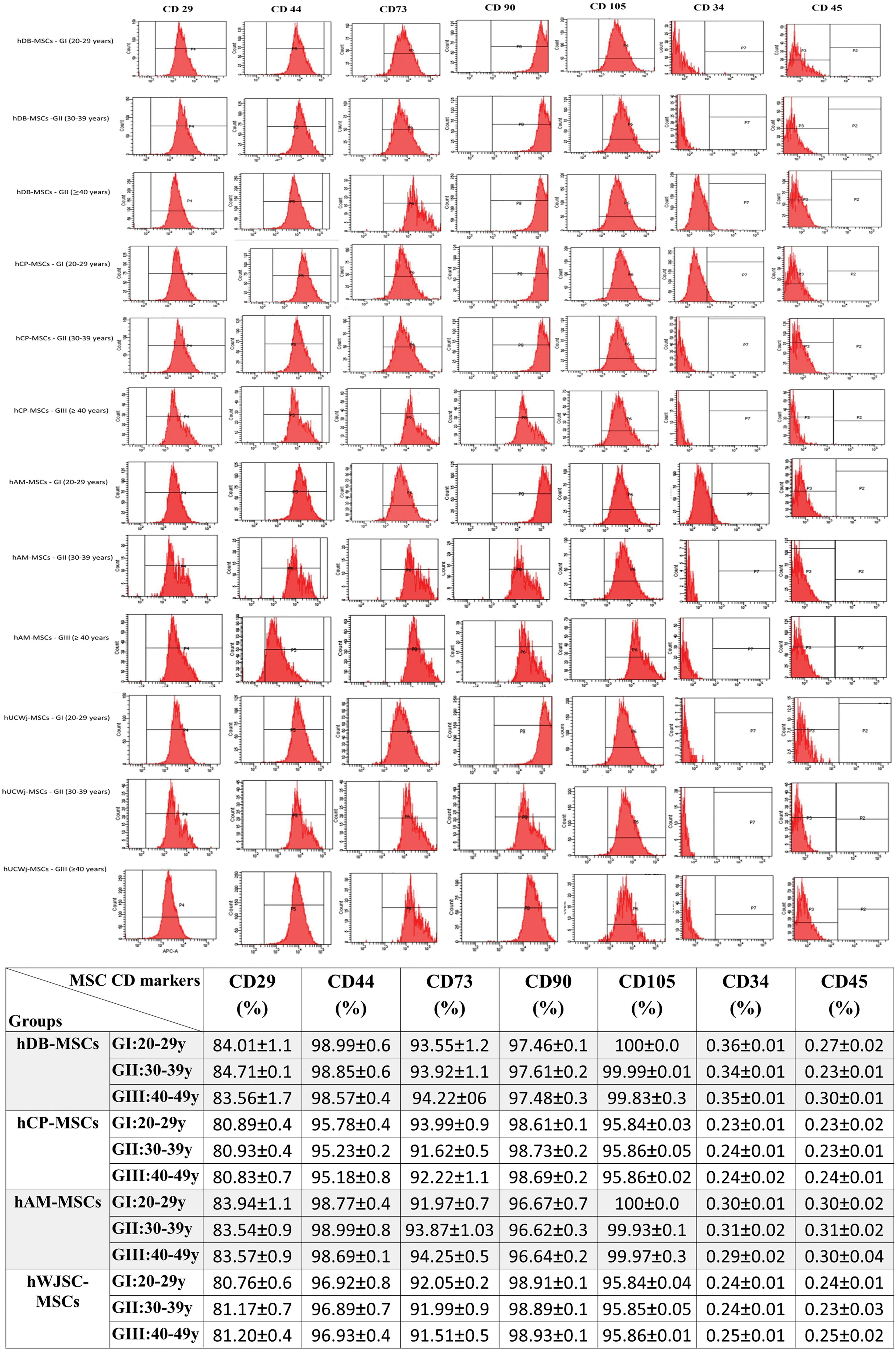
Mesenchymal stem cells from fetal membranes (hFM-MSCs) CD markers analysis using FACS. Representative histogram of the flow cytometry analysis (FACS) of MSC-related CD surface markers expression on hFM-MSCs [decidua basalis (hDB), chorionic plate (hCP), amniotic membrane (hAM), and umbilical cord Wharton's jelly (hWJSCs)] from three different age groups, at passages 3. Color images are available online.
Comparison hFM-MSCs Expression of MSC CD Markers in Different Studied Groups
Data are given as mean ± standard deviation of the flow cytometry analysis (FACS) of MSC-related CD surface markers expression compare hFM-MSCs expression of MSC CD markers in different studied groups on hFM-MSCs [decidua basalis (hDB), chorionic plate (hCP), amniotic membrane (hAM), and umbilical cord Wharton's jelly (hWJSCs)] from three different age groups, at passages 3.
hFM-MSC, human fetal membranes–mesenchymal stem cell.
Assessment of proliferation capacity
MTT assay was performed in the 6th passage to determine the proliferation capacity of hFM-MSCs in different age groups at different durations of culture (at 24, 48, and 72 h). The hFM-MSCs from different fetal membranes showed that there was significant difference in proliferation level between different groups, between different durations, and when the groups and duration intersect. Where the hDB-MSCs showed the significant difference in the proliferation level between different groups (mean square = 11.835, F = 771.619), between different durations (mean square = 1.550, F = 101.077) and when the groups and duration intersect (mean square = 0.146, F = 9.494) and (P < 0.05) between all.
As well, the hCP-MSCs showed the significant difference in the proliferation level between different groups (mean square = 10.053, F = 510.857), between different durations (mean square = 2.410, F = 122.464) and when the groups and duration intersect (mean square = 0.274, F = 13.925) and (P < 0.05) between all. The hAM-MSCs showed the significant difference in the proliferation level between different groups (mean square = 11.043, F = 533.683), between different durations (mean square = 2.042, F = 98.684) and when the groups and duration intersect (mean square = 0.290, F = 14.001) and (P < 0.05) between all. In addition, the hWJMSC-MSCs showed the significant difference in the proliferation level between different groups (mean square = 9.910, F = 621.015), between different duration (mean square = 2.553, F = 159.991) and when the groups and duration intersect (mean square = 0.283, F = 17.762) and (P < 0.05) between all.
On the contrary, the LSD test of significance in proliferation capacity between different age groups at different durations showed significantly increase in K562 proliferation levels at 24 h (1.73 ± 0.21), 48 h (2.71 ± 0.19), and 72 h (3.16 ± 0.20), compared with levels in hFM-MSCs of different age groups. Meanwhile, hBM-MSC proliferation levels were significantly decreased at 24 h (0.13 ± 0.01), 48 h (0.17 ± 0.02), and 72 h (0.21 ± 0.01) compared with levels in hFM-MSCs of different age groups (Table 2). In addition, the MSCs from fetal membranes of GII and GIII showed an increase in proliferation compared with hFM-MSCs of GI at 24, 48, and 72 h (Table 2). The hDB-MSCs and hAM-MSCs from all the studied age groups showed significant increase in proliferation rates in culture at 24, 48, and 72 h compared with those of hCP-MSCs and hWJSCs (Table 2).
Comparison of hFM-MSCs Proliferation Levels in Different Studied Groups by MTT Assay
Data are given as mean ± standard deviation to compare proliferation level of hFM–MSCs. Comparison was made using two-way ANOVA (LSD) test and P < 0.05 was considered significant. hMSCs, mesenchymal stem cells from different fetal membranes (hDB, hCP, hAM, and hWJSCs); K562, human leukemic cancer cell; hBM-MSCs, mesenchymal stem cells from bone marrow. 1P, significance versus hMSCs of GI (20–29 years); 2P, significance versus K562 cells; 3P, significance versus hBM-MSCs; 4P, significance versus h-MSCs of GII (30–39 years). # P: significance versus 24 h of the same group; ## P: significance versus 48 h of the same group.
ANOVA, analysis of variance; LSD, least significant difference.
hFM-MSC expansion ability was evaluated using doubling time assay. The mean doubling time of K562 was significantly decreased (19.39 ± 0.31) compared with that of hFM-MSCs of different age groups, whereas hBM-MSC mean doubling time was significantly increased (75.42 ± 0.04) compared with that of hFM-MSCs in different age groups. Furthermore, MSCs from fetal membranes of GII and GIII showed a decrease in mean doubling time compared with that of hFM-MSCs of GI (Table 3). Moreover, hDB-MSCs and hAM-MSCs from all the studied age groups showed a significant decrease in mean doubling time compared with that of hCP-MSCs and hWJSCs (Table 3).
Comparison of hFM-MSCs Proliferation Rates in Different Studied Groups by Doubling Time
Data are given as mean ± standard deviation to compare hMSCs growth rates of hFM-MSCs (D) with different studied groups by doubling time (DT). Significance was determined using multivariate analysis and then significant data were analyzed by one-way analysis of variance (LSD) test. P < 0.05 was considered significant. IP, significance versus hMSC from GI of the same region; 2P, significance versus K562; 3P, significance versus hBM-MSC, 4P, significance versus hMSC from GII of the same region.
Assessment of hTERT gene expression
Gene expression of hTERT was assessed using qRT-PCR. It was noticed that expression of hTERT was upregulated in K562 compared with that in hFM-MSCs of different age groups, and in different FM regions. On the contrary, expression of hTERT was downregulated in adult hBM-MSCs compared with that in hFM-MSCs in different age groups and in different FM regions (Table 4).
Comparison of hTERT Expression Levels of hMSCs in Different Studied Groups
Gene expression level of hTERT assessed by comparative Ct method (ΔCt). The cutoff of fold change >±2. Comparison was made using one-way ANOVA (LSD) test and P < 0.05 was considered significant. hMSCs, mesenchymal stem cells from different fetal membranes (hDB, hCP, hAM, and hWJSC); K562, human leukemic cancer cell; hBM-MSCs, mesenchymal stem cells from bone marrow; 1P, significance versus hMSCs of GI (20–29 years); 2P, significance versus K562 cells; 3P, significance versus hBM-MSCs, 4P, significance versus h-MSCs of GII (30–39 years).
In terms of the effect of age, expression of hTERT was significantly upregulated in hDB-MSCs, hCP-MSCs, hAM-MSCs, and hWJSCs of GII (P < 0.05) and GIII (P < 0.05) compared with that in those of GI (Table 4). There was also a differential expression of hTERT in hMSCs among different fetal membranes at GII and GIII, as hDB-MSCs and hAM-MSCs showed higher expression levels than did hCP-MSCs and hWJSCs, whereas hAM-MSCs showed higher expression levels than did hDB-MSCs (Table 4).
Assessment of telomere length in hFM-MSCs
The telomere length was assessed using qRT-PCR. It was noticed that the telomere was longer in K562 (52.86 ± 2.26) than in hFM-MSCs in different age groups and in different studied FM regions. Moreover, the telomere was shorter in adult hBM-MSCs (1.01 ± 0.01) than in hFM-MSCs in different age groups and in different studied FM regions (Table 5).
Comparison of Telomere Length of hMSCs Different Studied Groups
Length of telomere assessed by quantification cycle value method (ΔΔCq). The cutoff of telomere length >±1. Comparison was made using one-way ANOVA (LSD) test and P < 0.05 is considered significant. hMSCs, mesenchymal stem cells from different fetal membranes (hDB, hCP, hAM, and hWJSCs); K562, human leukemic cancer cell; hBM-MSCs, mesenchymal stem cells from bone marrow; 1P, significance versus hMSCs of GI (20–29 years); 2P, significance versus K562 cells; 3P, significance versus hBM-MSCs; 4P, significance versus hMSCs of GII (30–39 years).
In terms of the effect of age, it was noticed that the telomere length significantly decreased in MSCs of GII (P < 0.05) and GIII (P < 0.05) in hDB, hCP, hAM, and hWJSCs compared with that in those of GI (Table 5). At the same time, hDB-MSCs and hAM-MSCs showed increased telomere length compared with that in other regions in all the studied age groups, whereas hAM-MSCs showed higher expression levels than did hDB-MSCs (Table 5).
Discussion
The aging features shown by MSCs derived from old tissues or old donors pose limitations to their use in cell therapy [30 –32]. Thus, determining the characteristics of hMSCs derived from fetal membranes of mothers in different age groups may be useful to select the optimal fetal membranes and mother's age to maintain the placenta and umbilical cord in cell banks and to identify their potential uses in therapeutic applications.
For this, the study was designed to evaluate the impact of maternal age on the characteristics of hMSCs (expression of hTERT, telomere length, and proliferation potential) derived from hFMs, including hDB-MSCs, hCP-MSCs, hAM-MSCs, and hWJSCs. Previous studies have explored donor's age effects of hMSCs derived from adult tissues [5,17,32,33]. However, to the best of our knowledge, this is the first report to screen the impact of maternal age on the characteristics of hMSCs derived from hFMs and to unveil their potential use in cellular therapy and regenerative medicine.
We isolated and characterized hFM-MSCs using human MSC-related cell surface markers, whereas the proliferation level was estimated by MTT assay and doubling time. Telomere length and hTERT expression were analyzed using quantitative RT-PCR. Moreover, we used K562 and hBM-MSCs cells as controls to be certain about the risk and critical range of telomere length from hFM-MSCs by comparing their characteristics with K562 and hBM-MSCs. K562 cells are a rich source of telomerase, have long telomeres, and are able to undergo proliferation in long-term culture in vitro [34]. Thus, they were used as positive control. In contrast, hBM-MSCs are a poor source of telomerase, have short telomeres, and tend to progressively lose the proliferative ability and undergo rapid aging after minimal expansion in vitro [33,35]. Hence, they were used as the negative control.
In this study, hFM-MSCs from different age groups maintained their fibroblast-like shape and attached to the culture flask. In addition, our flow cytometry analysis revealed strong positive expression of CD29, CD44, CD73, CD90, and CD105, and negative expression of CD34 and CD45. These results indicate that the MSCs derived from fetal membranes meet the characterization criteria for MSCs that have been standardized by the International Society for Cellular Therapy (ISCT) [27,36].
We also observed an increase in the proliferation rate of hFM-MSCs from all age groups with the increase in culture duration (24, 48, and 72 h), and decrease in mean of doubling time with increase age and culture time. This decrease in mean doubling time indicates that hFM-MSCs show the highest growth rate and the greatest proliferative capacity with increase in age when grown in vitro. Moreover, a significant increase in proliferation and growth rate of hFM-MSCs was observed in GII and GIII, and also observed that hFM-MSCs had significantly increased proliferation rates in hDB and hAM regions in GII and GIII and growth rate. This is consistent with Shaer et al. who studied the characteristics of hMSCs derived from human placenta and indicated a high proliferation rate of hMSCs up to passage 20, except MSCs derived from the amniotic membrane, which had low proliferation rates after passage 14 [2]. This was confirmed by Choi et al. [17].
The difference in proliferation intensity in different regions between young and older age observed in this study might be because of the nature of human stem cell proliferation, as they have a slow rate of proliferation and do not divide unless there is a need to it [21,23]. Although the difference in hMSCs properties from different fetal membrane (hDB, hCP, hAM, and hWJSCs) regions might be attributed to the origin of these membranes and the complex nature of their development, placenta and fetal membrane are complex organs made up of maternal and fetal. Whereas the chorionic plate (CP), amniotic membrane (AM), and umbilical cord Wharton's jelly (WJMSCs) are of fetal origin and come from the inner layer of blastocyst called epiblast, decidua basalis is of maternal origin, and come from an outer layer of blastocyst called trophoblast.
Trophoblast differentiates into two layers and invade, proliferate, and migrate to the endometrium, and undergo different developmental stages to form the chorion frondosum. For that, the hCP, hAM, and hWJSC regions provide more primitive hMSCs compared with hMSCs from decidua basalis [31,37,38]. Besides, we found differences in proliferation capacity of MSCs from hAM compared with hCP and hWJSCs-MSCs that have the same origin; this might be owing to differences in patterns of gene expression of these cells in hAM region, as confirmed by Kim et al., Wegmeyer et al., and Ma et al. [39 –41].
The most striking results in this study were the ones revealing that the hFM-MSCs from GII and GIII presented comparable telomere length in different fetal membranes despite the decreased telomere length compared with the ones from GI. Nevertheless, hTERT expression was upregulated in hFM-MSCs from fetal membranes of mothers in GII and GIII, in which these cells undergo high proliferation with preservation of adherence to plastic and have a normal spindle shape in culture. In contrast, the hBM-MSCs derived from older donors have a short telomeres and lose the proliferative ability and undergo rapid aging.
The possible explanation for the appearance of the telomerase expression and increasing proliferate ability in fetal membranes through the shortening of telomere either owing to the use of growth factors (b-FGF) in hFM-MSCs culture, which stimulate hTERT gene expression, increase the length of originally short telomeres, and enhance differentiation, or the insufficient telomerase activity in adding DNA repeats. However, in the latter scenario, cells continue to take advantage of telomerase activity to protect the telomeres by slowing down the shortening of telomeres, and increasing proliferation that need to perform their functions until telomeres reach a critical length, in which case cells start aging.
This refers to the hFM-MSCs from older mother's age that increased their proliferation rates as a result to short of telomeres or stimulating by growth factors, might be unsuitable for expansion in vitro or their use in cellular therapy. Similar results were obtained by Schmelzer et al., who confirmed the role of growth factors, cytokines, and some antioxidants in stimulating the telomerase activity and increasing the telomere length in MSCs [42]. They suggested that one of the most essential function of telomerase is to avoid critical shortening of telomeres and loss of telomere function.
This was confirmed in previous studies [43,44]. In addition, the studies on MSCs from adult tissues, which reported that MSCs derived from younger donors yield more colonies containing larger numbers of cells and higher proliferative rate than those obtained from older donors that lose their stem cell characteristics. This proves the effect of donor age on MSC characteristics [33,35,45]. In addition, the experimental evidence shows that in the embryonic stem cells and stem cells from fetal tissues the reactions to DNA damage and activation of telomerase are distinct from the stem cells derived from adult tissues and somatic cells [46].
In this study, it was also observed that there was differential hTERT expression in hMSCs from different fetal membranes until the end of the study (passage 6); the hAM-MSCs showed high expression levels of hTERT compared with hDB-MSCs, hCP-MSCs, and hWJSCs. In addition, the telomere was longest in hAM-MSCs compared with that in hDB-MSCs, hCP-MSCs, and hWJSCs in all age groups. These results were in concordance with those of Karimi et al. who reported that the MSCs from umbilical cord underwent telomere loss within the first few passages [22]. However, this finding was contradictory to that obtained by Ventura et al. who found that MSCs from the chorion undergo minimal telomere loss in long-time cultures [12].
The difference between these studied FM regions might be attributed to the origin of these cells and the complex nature of their development, as described earlier. Although the difference between studies on hMSC characteristics might be because of differences in the lifestyle and age of MSC donors, mechanisms of telomere maintenance, or isolation protocols of these cells.
There were a few limitations in our study. Placental villi were not included as a part of fetal membranes. The study's goal was to isolate hMSCs from complex fetal membranes and maintain them without contamination, in a rapid manner, making isolation of placental villi very difficult. In addition, it could not be confirmed if telomere length and telomerase activity of hFM-MSCs could be maintained in long-term culture. Again, the primary goal was to obtain enough cells from hFM-MSCs to characterize them in the mothers of different age groups and this was achieved in the first six passages.
Moreover, the potential clinical application of hMSCs is generally recommended at early passage. In addition, several studies have reported the successful expansion of hMSCs from fetal membranes to >15–20 passages while maintaining hMSC proliferation and differentiation abilities [6,26,47]. Unfortunately, functional analysis, mesodermal differentiation, immunomodulation, and neuroprotective properties were not included in our study because the effect of the mother's age on these potentials of hMSCs was not in the study scope.
Conclusions
In conclusion, this study is the first to determine the effect of mother's age on hTERT gene expression, telomere length, and proliferation rate of MSCs derived from different hFMs. This study revealed that although hFM-MSCs derived from different fetal membranes maintained the MSCs characteristics in all study groups, the hAM-MSCs and hDB-MSC with different maternal ages presented better proliferation properties compared with other regions. Moreover, hFM-MSCs from older mother affected telomere length and telomerase expression in different fetal membranes, the hFM-MSCs showed a shorter telomere and increased the level of telomerase expression and high proliferation rate during culture compared with those of younger mothers. This might be attributed to the growth factors (b-FGF) used in hMSCs culture or a decrease in telomere length, which stimulated the activation of telomerase as a mechanism to maintain telomere length and increased proliferation rates in these regions. Moreover, the striking result was that there was no difference in telomere length and telomerase expression between group II and group III. Determination of telomere length, telomerase levels, the role of growth factors (b-FGF), and mechanisms underlying the activation of telomerase in hFM-MSC of older mothers calls for a more thorough investigation to provide a better understanding of the hFM-MSC characteristics.
Footnotes
Acknowledgments
The authors thank the Department of Obstetrics and Gynaecology at King Abdulaziz University Hospital and Embryonic Stem Cell Research Unit at King Fahad Medical Research Centre, King Abdulaziz University, Jeddah, Saudi Arabia for their support to the authors in conducting this study. The authors thank Editage for English language editing.
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This study was supported by King Abdulaziz City for Science and Technology by a research grant no. 103-122.