
Abstract
Human mesenchymal stem cells (hMSCs) are considered to be able to adapt to environmental changes induced by gravity during cell expansion. In this study, we investigated neurogenic differentiation potential of passaged hMSCs under conventional gravity and simulated microgravity conditions. Immunostaining, quantitative real-time reverse transcription–polymerase chain reaction (qRT-PCR), and western blot analysis of neurogenic differentiation markers, neurofilament heavy (NF-H), and microtubule-associated protein 2 (MAP2) revealed that differentiated cells from the cells cultured under simulated microgravity conditions expressed higher neurogenic levels than those from conventional gravity conditions. The levels of NF-H and MAP2 in the cells from simulated microgravity conditions were consistent during passage culture, whereas cells from conventional gravity conditions exhibited a reduction of the neurogenic levels against an increase of their passage number. In growth culture, cells under simulated microgravity conditions showed less apical stress fibers over their nucleus with fewer cells having a polarization of lamin A/C than those under conventional gravity conditions. The ratio of lamin A/C to lamin B expression in the cells under simulated microgravity conditions was constant; however, cells cultured under conventional gravity conditions showed an increase in the lamin ratio during passages. Furthermore, analysis of activating H3K4me3 and repressive H3K27me3 modifications at promoters of neuronal lineage genes indicated that cells passaged under simulated microgravity conditions sustained the methylation during serial cultivation. Nevertheless, the enrichment of H3K27me3 significantly increased in the passaged cells cultured under conventional gravity conditions. These results demonstrated that simulated microgravity-coordinated cytoskeleton–lamin reorganization leads to suppression of histone modification associated with neurogenic differentiation capacity of passaged hMSCs.
Introduction
Human mesenchymal stem cells (hMSCs) play a promising role in tissue engineering and regenerative medicine with their self-renewal and differentiation abilities [1]. The multilineage differentiation potential of hMSCs has also raised the possibility of autologous hMSC transplantation as therapy for human illnesses [2]. The characteristic of expanded hMSCs for cellular application is the most important factor to ensure that cell-based therapy can be beneficial for human diseases [3]. However, during in vitro expansion under conventional gravity culture conditions, cells with an increased passage number can lose their abilities of proliferation and potency of multilineage differentiation [4 –6], along with a reduction in neurogenic gene expressions in the late-passaged differentiated neural cells [7]. Therefore, alternative culture strategies and culture systems are needed for preparation of the required quantities of cells, which allow them to preserve the undifferentiated state or direct stem cell differentiation into the desired lineage.
With the rapid advances in our understanding of cellular capacity due to adaptation in the microgravitational field, the physical microenvironment of the cell culture system is known to be a critical cue that directs cellular growth and differentiation [8]. As fundamental investigations began in the microgravity space environment, several specific devices for cell culture system in the space have been developed [9]. This raises questions about the cellular adaptation to an altered gravity environment shown as alterations in perception and transduction of cellular mechanical forces through actin filaments, and also the changes in the dynamics of enzyme kinetics and self-assembly of protein networks [10].
Intercellular and intracellular mechanical forces influence the response of cell nucleus owing to transduction of the mechanical signaling through the nucleoskeleton, affecting genome expression and organization [11]. It is well known that modification in epigenetic markers in chromatin, such as histone methylation, plays an important role in the promotion or inhibition of appropriate transcriptional pathways without changes in the DNA sequence [12]. These epigenetic modifications play an essential part in the regulation of numerous cellular processes involved with various cellular phenotypes [13]. In addition, it has been reported that microgravity exposure affects chromatin remodeling and contributes to altered gene expressions [14]. However, the association between gravity culture conditions and epigenetics is still unclear. The mechanical transductions from the extracellular matrix (ECM) to nucleus through cytoskeleton and nucleoskeleton may be an important regulator for the modification of histone methylation in cells.
A three-dimensional clinostat is one of the common devices that have been used to provide simulated microgravity culture conditions [15], and it has been shown to be an effective method on ground for attenuating gravitational effects similar to spaceflight experiments [16]. The mechanisms by which epigenetic histone modification patterns are inherited during cell expansion under simulated microgravity conditions still remain elusive [17]. Furthermore, until now, there is a lack of understanding about influence of passage culture of hMSCs on their differentiation capacity into neurons. Therefore, the aim of this study was to investigate the effect of simulated microgravity on neurogenic differentiation potential of passaged hMSCs during growth expansion culture. We also discussed the contribution of epigenetic memory of histone modifications on neurogenic differentiation capacity of hMSCs during serial cultivation. The results of this study led us to propose alternative culture processes, which possibly maintain the quality of hMSCs during long-term culture in terms of differentiation into neural lineage.
Materials and Methods
Cells and culture conditions
Bone marrow-derived hMSCs (Lot no. 0000654251; Lonza Walkersville, Inc., Walkersville, MD) were cultured according to the manufacturer's instruction in hMSC growth medium (Lonza) at 37°C under 5% CO2 in a humidified atmosphere. At 70% confluency, the cells were enzymatic detached with a 0.1% trypsin/0.02% EDTA solution (Sigma-Aldrich, St. Louis, MO). Then, the cells were seeded at a density of 4.0 × 103 cells/cm2 in every passage and a viable cell number was estimated by counting of suspended cells with a hemacytometer, by a Trypan Blue dye exclusion test.
For simulated microgravity culture, the Gravite® (Space Bio-Laboratories Co., Ltd., Hiroshima, Japan) generates a simultaneous rotation on two axes with constant angular speed. At the center of the rotating frames, the Gravite cancels the cumulative gravity at the center of machine that simulates microgravity environment of 10–3 G in 8 min after the operation estimated by an installed acceleration sensor. Thus, this is accomplished to scatter the gravitational vector identically within a spherical boundary.
In every passage of growth culture, on day 0, hMSCs were seeded into a 12.5-cm2 T-flask with a vented filter cap (Corning, Tewksbury, MA) and cultured under conventional gravity environment for a day in a manner in which cells are attached on the bottom culture surface. Then, on day 1, the medium was removed from the flask. Subsequently, the flask was fully filled with fresh medium without any bubbles. After that, the cells were cultured under conventional gravity culture or simulated microgravity environment at the center of the Gravite until day 5 without any medium change. On day 5 of every passage, the cells were subcultured into a new culture vessel, and the cells were cultured for three passages for 15 days.
Proliferation ability
In every passage of growth culture, adhesion efficiency and specific growth rate of the cells were evaluated for estimation of proliferation ability. After seeding for 24 h, the cells attached on the culture surface and those were monitored through a phase-contrast microscope with magnification at 4 × . The attachment efficiency, α (−), was calculated as follows: α = X 24/X 0, where X 24 is the cell density after seeding for 24 h, and X 0 is the cell density at the seeding. The specific growth rate, μ (h−1), in an exponential growth phase in each passage was determined as follows: μ = ln(X 120/X 24)/96, where X 120 represents the cell density after seeding for 120 h as estimated by a Trypan Blue dye exclusion test.
Neurogenic differentiation potential
For neurogenic differentiation culture, hMSCs that have been cultured under conventional gravity or simulated microgravity conditions on day 5 of every passage, were transferred into a new culture vessel coated with fibronectin (PromoCell, Germany) with seeding density of 5.0 × 103 cells/cm2. The cells were incubated with hMSC growth medium for a day under conventional culture environment. After that, the medium was replaced with neurogenic differentiation medium (PromoCell). Then, the cells were cultured under conventional gravity conditions for 7 days. The differentiation medium was refreshed after every 3 days.
Fluorescent staining
Immunostaining was done as described previously [18]. Concisely, the cells were washed twice with phosphate-buffered saline (PBS), and then, were fixed with 4% paraformaldehyde in PBS (Wako Pure Chemical Industries, Tokyo, Japan) for 10 min at room temperature. After that, the cells were permeabilized with 0.5% polyoxyethylene octylphenyl ether in PBS for 10 min. Then, the samples were masked by the nonspecific proteins by overnight incubation with Block Ace (Dainippon Sumitomo Pharma Co., Ltd., Osaka, Japan) at 4°C. Consequently, the cells were incubated overnight with primary antibodies at 4°C. In this study, we used primary antibodies against lamin A/C (Santa Cruz), lamin B (Abcam, Cambridge, UK), neurofilament heavy (NF-H; Abcam), and microtubule-associated protein 2 (MAP2; Abcam) that were diluted in deionized water containing 10% Block Ace. After the incubation, the samples were washed twice with Tris-buffered saline (TBS) and immunolabeled with the appropriate secondary antibody (Alexa Flour-conjugated IgG; Life Technologies) at room temperature for an hour. The F-actin and nuclei were fluorescent stained with rhodamine phalloidin and 4′,6-diamidino-2-phenylindole (Life Technologies), respectively. The stained samples were observed manually by a confocal laser scanning microscope (FV-1000; Olympus, Tokyo, Japan).
Protein extraction and western blot analysis
The extraction and blotting analysis were performed as described previously [19]. Briefly, total protein was extracted from the cells by incubation on ice with a RIPA lysis buffer (Sigma-Aldrich) containing a Halt™ protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific, Waltham, MA) for 30 min. Protein concentration was estimated using a BCA Protein Assay Kit (Thermo Fisher Scientific). Equal amounts of total proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE; Wako), and then, transferred to nitrocellulose membrane (Bio-Rad). The membrane was blocked with ECL™ blocking agent (GE Healthcare, Chicago, IL) at room temperature for an hour. After that, the membrane was immunolabeled with primary antibodies that were diluted in TBS containing ECL blocking agent at 4°C overnight. In this analysis, we used the antibodies against lamin A/C (Santa Cruz), lamin B (Abcam), GAPDH (Sigma-Aldrich), H3K4me3, H3K27me3, H3 (Cell Signaling Technology, Inc., Danvers, MA), NF-H, and MAP2 (Abcam). Later, fluorescence staining was performed for 1 h at room temperature using the following secondary antibodies: DyLight® 800 anti-Rabbit IgG (Thermo Fisher Scientific) and Starbright™ Blue 700 Anti-Mouse IgG (Bio-Rad). Fluorescent signals were displayed using a ChemiDoc™ MP Imaging system (Bio-Rad). Finally, the optical intensity of protein signals was quantified using an image analysis software (Image Lab™; Bio-Rad).
Quantitative real-time reverse transcription–polymerase chain reaction
RNA isolation and quantitative real-time reverse transcription–polymerase chain reaction (qRT-PCR) were conducted as described previously [20]. Shortly, RNA was isolated from the cells using an RNeasy Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer's procedure. The RNA was then reverse transcribed to cDNA using a PrimeScript® RT Reagent Kit (TaKaRa Bio, Inc., Shiga, Japan). The qRT-PCR assays were conducted using SYBR® Premix Ex Taq (TaKaRa Bio, Inc.) with specific primers. The qRT-PCR primer sequence is listed in Supplementary Table S1. The assay was performed on a real-time PCR system (Applied Biosystems 7300; Thermo Fisher Scientific) and the relative mRNA expression of the target genes was evaluated by the cycle threshold value (C t) and normalized to that of GAPDH using the 2−ΔΔCt method.
Chromatin immunoprecipitation analysis
Chromatin immunoprecipitation (ChIP) analysis was conducted using a SimpleChIP® Enzymatic Chromatin Immunoprecipitation Kit (Cell Signaling Technology, Inc.), according to the manufacturer's protocol. In this study, we used three antibodies (anti-H3K4me3, anti-H3K27me3, and anti-IgG control; all purchased from Cell Signaling Technology, Inc.) for immunoprecipitation. Immunoselected DNA was quantified by RT-PCR method with the primers against the promoters of MAP2 and NF-H genes. The ChIP-quantitative PCR primer sequence is listed in Supplementary Table S2. The assays were analyzed using SYBR Premix Ex Taq (TaKaRa Bio, Inc.) on a real-time PCR system (Applied Biosystems 7300). Cycle threshold value (C t) in each condition was used to calculate relative fold enrichment of H3K4me3 and H3K27me3 at each specific promoter that was normalized to condition of IgG control.
Statistical analysis
All quantitative data were obtained from triplicate experiments, represented as the mean ± standard deviation. Comparisons between the conditions were evaluated by one-way analysis of variance with the Tukey's honest significance test. The statistical significance was considered at the P values less than 0.05.
Results
Proliferative ability and differentiation potential of hMSCs
Proliferation indices were considered as adhesion efficiency (α) and specific growth rate (μ) evaluated during a serial passage culture under conventional gravity conditions and simulated microgravity conditions. The α and μ values of hMSCs under both gravitational conditions were consistent at least up to the three passages. There was no significant difference in adhesion efficiency and specific growth rate in the cells between both the gravitational culture conditions (Supplementary Fig. S1).
To clarify the change in phenotypes of passaged hMSCs during growth expansion culture under both gravitational conditions, we analyzed hMSC-specific markers (CD73, CD90), pluripotent markers (Oct4, Nanog), and neurogenic markers (NF-H, MAP2) in the cells on day 5 of each passage. The expressions of CD73 and CD90 exhibited no significant levels between cells cultured under both gravitational conditions, which were maintained through continuous passages (Supplementary Fig. S2A, B). The expressions of Oct4 and Nanog in cells cultured under simulated microgravity conditions were higher than those grown under conventional gravity conditions and kept at a high level at least up to the three passages (Supplementary Fig. S2C, D). However, the pluripotent levels in cells grown under conventional gravity conditions gradually decreased against an increase of their passage number. At the third passage, the relative expressions of Oct4 and Nanog in the cells cultured under simulated microgravity conditions were 2.3 and 2.1-fold higher than those grown under conventional culture conditions, respectively. Furthermore, the analysis of neurogenic markers during growth expansion culture revealed that cells passaged under both gravitational culture conditions had no significant difference in their neurogenic expressions (Supplementary Fig. S2E, F).
For evaluation of the neurogenic differentiation potential in passaged hMSCs cultured under both gravitational conditions, the cells were cultured with neurogenic differentiation medium for 7 days, and the expressions of neuronal markers (NF-H and MAP2) were evaluated by immunostaining, western blot, and qRT-PCR. Fluorescent microscopy revealed that a large number of NF-H- and MAP2-positive cells were found in all cultures of passaged cells from both the gravitational conditions (Supplementary Fig. S3A). Western blot analysis of NF-H and MAP2 showed notable differences in their protein expression levels that were dependent on gravitational conditions and passage numbers (Supplementary Fig. S3B). At the first passage of growth culture, the relative ratios of NF-H and MAP2 to GAPDH in the differentiated cells from the cells cultured under simulated microgravity conditions exhibited 1.3 and 1.4-fold higher than those from conventional gravity conditions, respectively (Fig. 1). The relative levels of NF-H and MAP2 in the cells from the passaged under simulated microgravity conditions were consistent. However, cells from conventional gravity conditions presented the reduction in NF-H and MAP2 levels in accordance with their increasing passage numbers; especially, the NF-H and MAP2 levels showed drastic changes after the first passage of the culture. At the third passage of growth culture, the relative levels of NF-H and MAP2 in cultures from passaged cells under simulated microgravity conditions showed 1.9 and 2.6-fold higher than those from conventional gravity conditions, respectively.
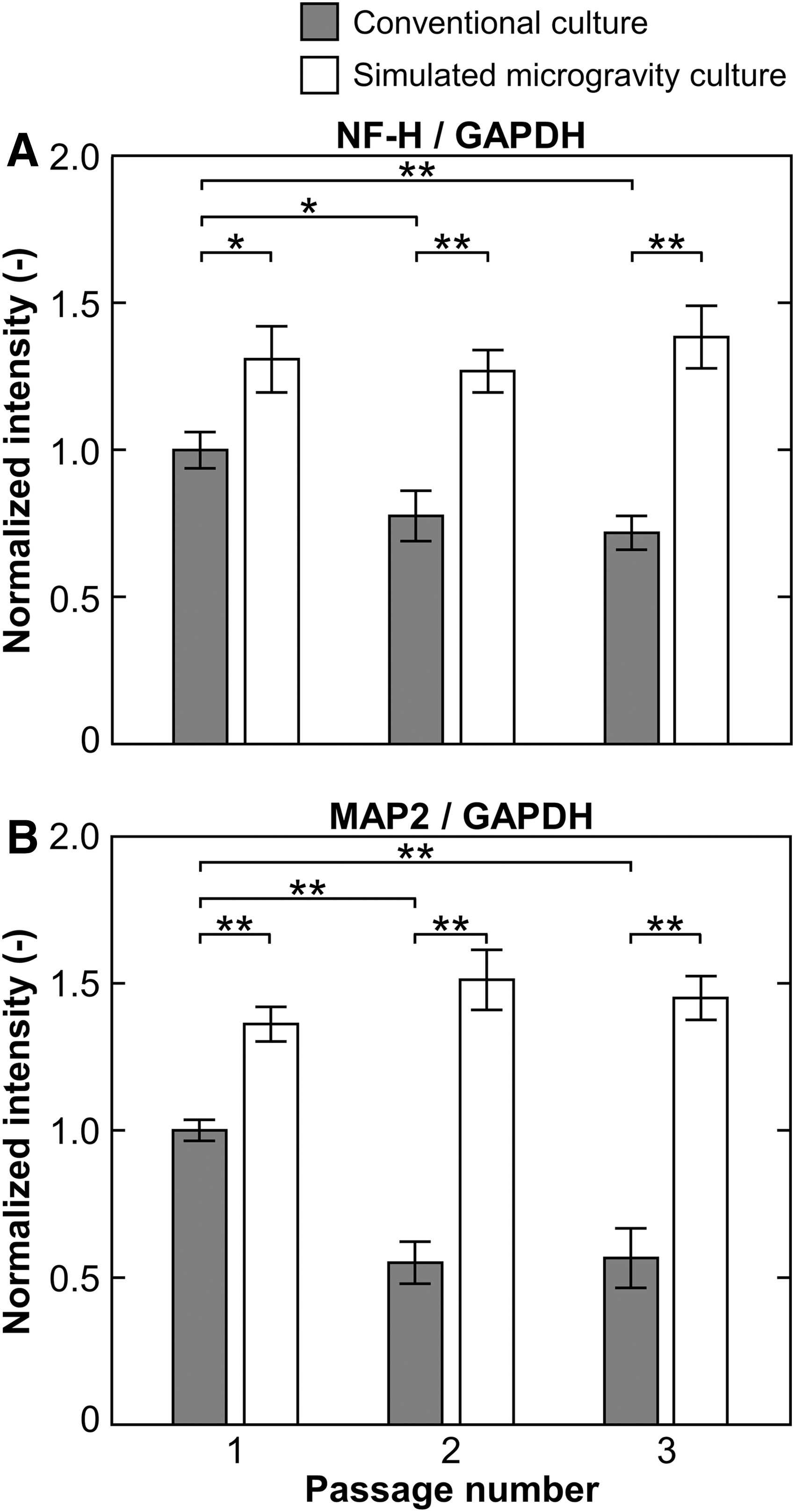
Neurogenic differentiation potential in passaged hMSCs evaluated by western blot analysis of the differentiated cells after culture in neurogenic differentiation medium under conventional conditions for 7 days. Quantitative data showing normalized intensities of NF-H
In addition, we also examined the expressions of two neural-related genes, NF-H and MAP2, in the cells during differentiation culture using qRT-PCR. The gene expression levels of NF-H and MAP2 in cells from simulated microgravity conditions were higher than those from conventional gravity conditions, at the same differentiation culture time (Supplementary Fig. S4). On day 7 of differentiation culture of cells from the first passage, cells from simulated microgravity conditions expressed relative levels of NF-H and MAP2 that were 1.5 and 1.8-fold higher than those from conventional gravity conditions, respectively. By increasing the passage number in growth culture, cells from simulated microgravity showed constant expressions of the neural-related genes; however, cells from conventional gravity conditions decreased their expressions with longer time in growth culture. At the third passage, the expression levels of NF-H and MAP2 in cells from simulated microgravity conditions were 2.7 and 2.5-fold higher than those from conventional gravity conditions, respectively.
Cytoskeleton and nucleoskeleton organization
To examine whether the cytoskeletal formation influences the nucleoskeletal response to gravitational conditions, fluorescent staining for F-actin and mechanosensitive nuclear lamin A/C was conducted in cells cultured under both the gravitational conditions on day 3 in every passage. Cells under conventional gravity conditions exhibited longitudinal actin stress fibers along both the apical and basal sides. They exhibited apical stress fibers over the nucleus together with appearance of lamin A/C at the apical site but not basal site, known as the polarization of lamin A/C. However, cells grown under simulated microgravity conditions had stress fibers located mainly along the basal side, and they presented larger cortical actin fibers in the filopodia and lamellipodia regions than those in conventional gravity conditions (Fig. 2A, B). Moreover, cells under simulated microgravity conditions mostly presented less apical stress fibers, with fewer cells having a polarization of lamin A/C than those grown under conventional gravity conditions.
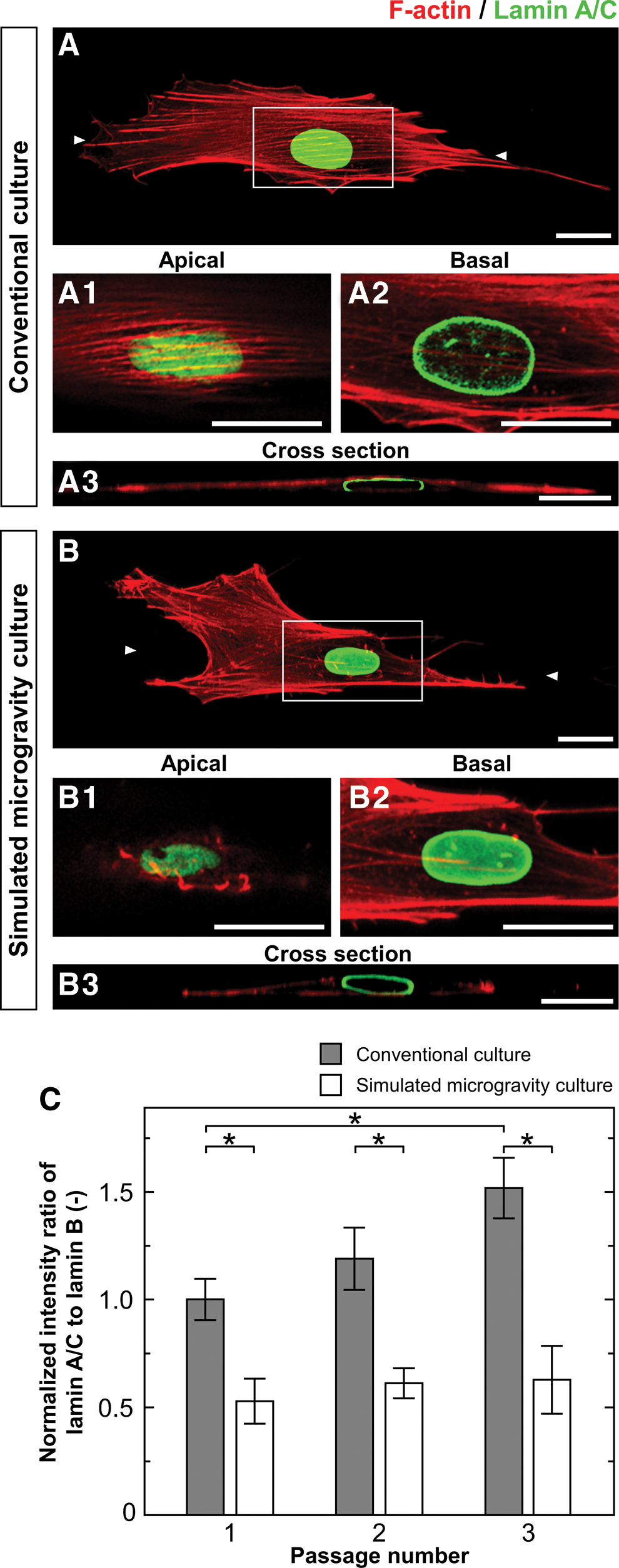
Cytoskeleton and nucleoskeleton organization in passaged hMSCs. Fluorescent staining images show F-actin (red) and lamin A/C (green) in the cells cultured under conventional conditions
The levels of nucleoskeletal proteins (lamin A/C and lamin B) were examined by western blot analysis of cells on day 5 in every passage (Supplementary Fig. S5) and the intensity ratio of lamin A/C to lamin B relative to GAPDH was calculated (Fig. 2C). At the first passage, the lamin ratio in cells cultured under simulated microgravity conditions was 1.9-fold lower than those cultured under conventional gravity conditions. Cells cultured under simulated microgravity conditions had no significant difference in the lamin ratio with the increasing passage numbers. However, the lamin ratio in the cells grown under the conventional gravity conditions gradually increased with the increasing passage numbers. At the third passage, the ratio of lamin A/C to lamin B in cells cultured under simulated microgravity conditions was 2.4-fold lower than those cultured under conventional gravity conditions.
Histone modification
To investigate the role of epigenetic chromatin markers, such as histone H3 trimethylation at lysine 4 (H3K4me3) and histone H3 trimethylation at lysine 27 (H3K27me3), cells cultured under both gravitational conditions on day 5 in every passage were investigated by western blot analysis for global analysis of histone modification (Supplementary Fig. S6). By calculation of the relative intensity of H3K4me3 and H3K27me3 to H3, there was no significant difference in the cells in terms of changes in passage numbers, and altered gravitational conditions (Fig. 3).
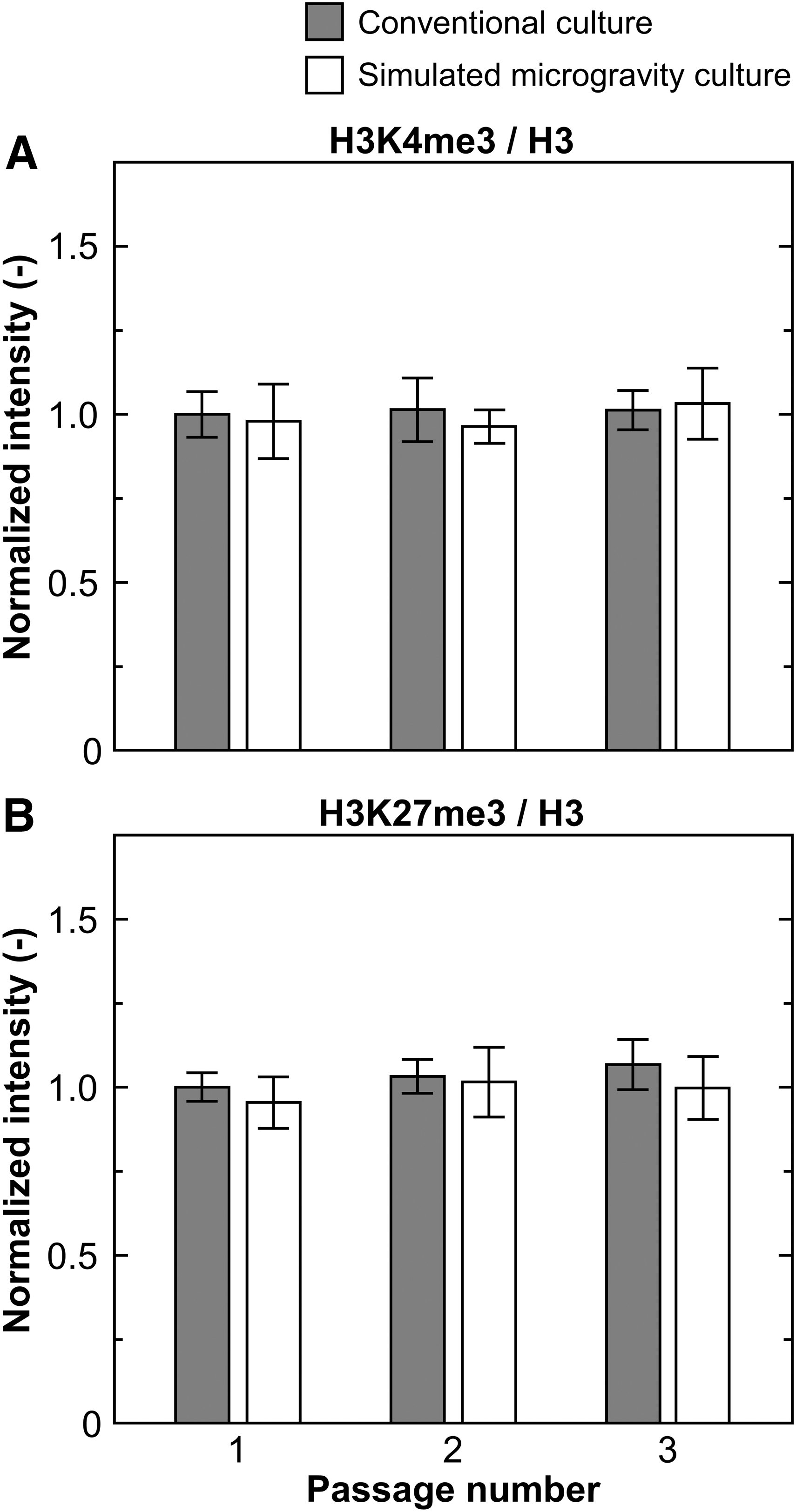
Global histone modification in passaged hMSCs evaluated by western blot analysis with quantitative data showing normalized intensities of H3K4me3
To elucidate the histone modification at specific regions, ChIP analysis of activation-associated H3K4me3 and repression-associated H3K27me3 at promoters of genes related to neuronal lineage (NF-H and MAP2) was performed in hMSCs cultured under both the gravitational conditions on day 5 in every passage. In case of H3K4me3 modification at NF-H and MAP2 promoters (Fig. 4A, B), there were no significant differences in the enrichment of H3K4me3 by variation of passage numbers or altered gravitational conditions. In case of H3K27me3 modification at neuronal promoters (Fig. 4C, D), at the first passage of growth culture, cells cultured under simulated microgravity exhibited the enrichment of H3K27me3 at promoters of NF-H and MAP2 that were 2.0 and 1.7-fold lower than those cultured under conventional gravity conditions, respectively. Cells passaged under simulated microgravity conditions sustained the enrichment of H3K27me3 at the neuronal promoters during passage, whereas those cultured under conventional gravity conditions significantly enhanced the enrichment of H3K27me3 with their increasing passage numbers. At the third passage, the enrichment level of H3K27me3 at promoters of NF-H and MAP2 in cells cultured under simulated microgravity conditions were 2.8 and 3.1-fold lower than those cultured under conventional gravity conditions, respectively.
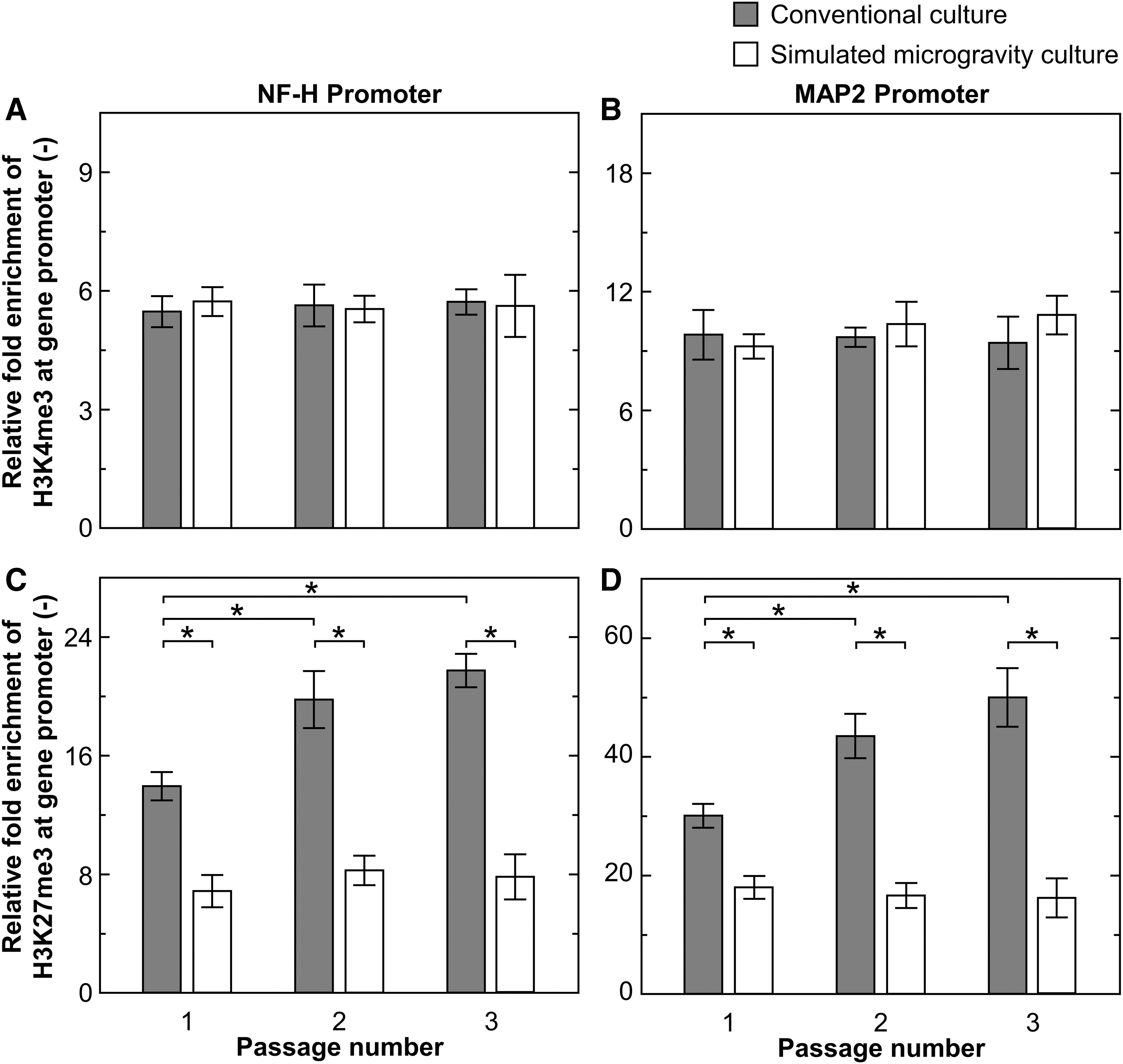
Specific histone modification in passaged hMSCs representing relative fold enrichments of H3K4me3
Discussion
Physical forces arising in the space have a profound impact on proliferation and differentiation potency of hMSCs during expansion culture, and the mechanisms need to be elucidated to control these biological functions. Nuclei respond to physical cues from their culture environment, which possibly induce the diverse cellular activities [21]. Lamin is a mechanosensitive protein that is known to control and regulate nuclear properties by the fibrillar network at the inner nucleus interacting with actin cytoskeleton [22].
Current development of a multidirectional gravity device for simulating microgravity has triggered various studies targeting to elucidate cellular adaptation to the environment of the gravitational conditions. Our previous report indicated that cells cultured under conventional gravity conditions exhibit a stretched morphology and undergo unidirectional migration, whereas cells cultured under simulated microgravity conditions present multidirectional migration with high frequency of directional changes in their movement [23]. Furthermore, cells cultured under conventional gravity conditions maintain their spindle-shape through fibronectin fibril formation in cell bodies and stabilization of focal adhesion with enriched stress fibers. Cells cultured under simulated microgravity conditions show an active extension of leading edges together with the inhibition of fibronectin fibrillogenesis by upregulation of MT1-MMP. Phosphorylation of paxillin in the cells cultured under simulated microgravity conditions was more intense at the cell edges but was less intense in the cell bodies compared with those under the conventional gravity conditions. Therefore, simulated microgravity influences cell behavior and mechanotransduction in hMSCs with changes in ECM assembly and spatial reorganization of the cytoskeleton [23].
This study investigated the neurogenic differentiation potential of passaged hMSCs under simulated microgravity conditions. Figure 5 shows a schematic illustration of our working hypothesis based on the results obtained in this study, with two main questions that have been addressed. First, how do different cytoskeletal formations get subjected to epigenetic modification under simulated microgravity conditions? Second, how can these mechanisms maintain neural differentiation potency under simulated microgravity conditions during passage culture?

Schematic illustration showing our working hypothesis of alterations in neurogenic differentiation potential in passaged hMSCs cultured under conventional conditions and simulated microgravity conditions due to changes in mechanotransduction, organization of nuclear lamins, and specific histone modification. Color images are available online.
Simulated microgravity-induced cytoskeletal reorganization leads to suppression of epigenetic modification of hMSCs
Changes in cell adhesion and cytoskeletal reorganization in response to environmental change are linked to epigenetic modification [24]. It has been suggested that structural pathway linking ECM to the nucleus, defined as mechanotransduction pathway, might be an essential biochemical transduction pathway for bringing the biophysical signaling into the nuclear interior [25]. In this study, we found differential cytoskeletal and nucleoskeletal changes in hMSCs when cultured under simulated microgravity conditions. The perinuclear actin cap forms a dome-like actin cap on top of the nucleus and has been shown to regulate nuclear shape and nuclear shape-determining function [26]. The basal-to-apical polarization of lamin A/C is tightly regulated by the assembly of the actin cap connecting to the nucleus through a linker of nucleoskeleton and cytoskeleton complex [27]. Some researchers have reported that the assembly of lamin A/C and lamin B is modified due to mechanical signaling to the nucleus [28,29]. Cells under conventional gravity conditions had apical stress fibers over the nucleus together with appearance of lamin A/C polarization (Fig. 2). However, cells cultured under simulated microgravity conditions exhibited few apical stress fibers over the nucleus that repressed the lamin A/C polarization and sustained the ratio of lamin A/C to lamin B during an increase of their passage number (Fig. 2). It is most likely that actin stress fibers that consist of the actin cap, a link to apical site of the nuclear surface, continuously generate compressive forces on the nucleus. Inhibition of actin cap formation decreases the level of nuclear pressure applied to the nuclear lamina, enforces polarized lamin A/C network at the apical site to remodel back to less tightly packed state [30].
Changes in organization of nuclear lamina affects chromatin/lamina attachments as well as genome organization [31]. Culture-induced epigenetic modifications affect the cell phenotypes and subsequent generations by alteration in gene expressions [32]. It has been reported that epigenetic switch involving histone methylation and demethylation at specific promoters of differentiation regions dictate hMSC lineage determination that influences the quality of differentiated cells [33]. It is well known that chromatin structure relates to histone methylation of H3K4me3 and H3K27me3 at specific sites, which associated with gene activation and repression, respectively, of transcriptions from those gene regions [34]. The histone markers can be maintained and transmitted to daughter cells through DNA replication and mitotic cell division [35]. Cells passaged under simulated microgravity conditions expressed significantly higher levels of NF-H and MAP2 proteins than those passaged under conventional gravity conditions (Fig. 1). Furthermore, analysis of repressive H3K27me3 modifications at promoters of NF-H and MAP2 indicated that cells from simulated microgravity conditions exhibited lower levels of these enrichments than those from conventional gravity conditions (Fig. 4). The findings of this study on simulated microgravity-induced alternation of cytoskeleton and nucleoskeleton indicated that epigenetic regulation might be a potential mechanistic basis for the changes in the expressions of some of the genes associated with phenotypic variation as a result of exposure to the cells cultured under simulated microgravity conditions.
Simulated microgravity-induced epigenetic memory plays an important role in the maintenance of neurogenic differentiation potency in hMSCs during passages
The epigenetic memory is influenced by the environmental changes and that histone modification impacts the differentiation of stem cells [36]. As mentioned above, mechano-adaptation of the nucleus respond to biophysical signaling that occurs in association with the actin cytoskeleton. The stem cell nucleus reacts to mechanical stresses by modification of the nucleoskeletal and chromatin architecture, and the mechanical loading that instills a mechanical memory in nuclei through changes in chromatin condensation [37].
During long-term exposure of physical input to the nucleus, force induces a more basal and persistent mechano-adaptation by synthesis of new or additional elements, which can control subsequent mechanical signals that are interpreted by the cells. Cellular memorization refers to a specific subdivision of mechanotransductive responses, defined as mechanical dosage, which is greater than some thresholds, resulting in persistence of mechano-adaptive response, continually long past the process of the original physical cues, which leads to influences on the mechano-adaption and functions of the cells [38]. hMSCs isolated from different sources, such as bone marrow and adipose tissue, alter epigenetic memory that contributes to changes in differentiation capacities into specific fates [39]. Thus, the condition of cell expansion is the buildup of these mechano-adaptive events that are stored in epigenetic memory.
In this study, we examined the epigenetic mechanisms that maintained their phenotypes while developing in passaged hMSCs under simulated microgravity conditions. Cells grown under simulated microgravity conditions showed maintenance of neurogenic differentiation potential during passage culture (Fig. 1, Supplementary Figs. S3 and S4); however, cells grown under conventional gravity conditions presented a reduction in the differentiation capacity due to accumulation of H3K27me3 at neurogenic promoter sites (Fig. 4). Some mechanical sensitive systems, molecules, and signaling pathways are involved in the response and adaptation of hMSCs to simulated microgravity. ECM-integrin-cytoskeleton system is an important system for mechanotransduction process [40 –42]. Simulated microgravity culture sustains the neurogenic differentiation capacity in passaged hMSCs by suppression of specific epigenetic modification at the neurogenic promoters together with controlling organization of the cytoskeletal and nuclear lamina during the passage culture. Therefore, design of the culture system, which is possible to regulate the epigenetics in hMSCs, is a critical challenge for controlling the quality of cellular differentiation related to the epigenetic mechanism in their memory storage. Further analysis of these adaptive and storage mechanisms will be required to fully elucidate the manner by which these signals influence stem cell differentiation and determine phenotypic commitment.
Conclusion
This study investigated the effect of simulated microgravity on neurogenic differentiation potential of hMSCs during expansion culture. Simulated microgravity plays an important role in the alternation of nuclear lamina–cytoskeleton interactions and the subsequent modification of epigenetic markers at specific regions that influence differentiation phenotypes. In addition, we found the role of simulated microgravity-induced epigenetic mechanisms on maintenance of neurogenic differentiation potential, which are heritable that have long-term effects on the regulation of gene expressions. Taken together, this study provided an insight into the fundamental epigenetic mechanism of effects of simulated microgravity during hMSC expansion along with being a practical guide for device design, operation, and application of the culture system in vitro.
Footnotes
Acknowledgments
The authors are grateful to Mr. Shinzou Sato, Mr. Ken-ichi Nishisako, Mr. Toshinori Tawa, and Mr. Yoshitaka Kanetada (Miyuki Ironworks Co., Ltd., Hiroshima, Japan) for preparation of the Gravite used in this study to generate the simulated microgravity conditions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported partially by the Kotozukuri Consortium for Cell Manufacturing and also the Japan Agency for Medical Research and Development (AMED) under grant number JP19be0604001.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.